
Transcriptomic Analysis of Gibberellin-Mediated Flower Opening Process in Tree Peony (Paeonia suffruticosa)
 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Phenotypic Changes in Tree Peony Under Different Exogenous GA3 Concentrations
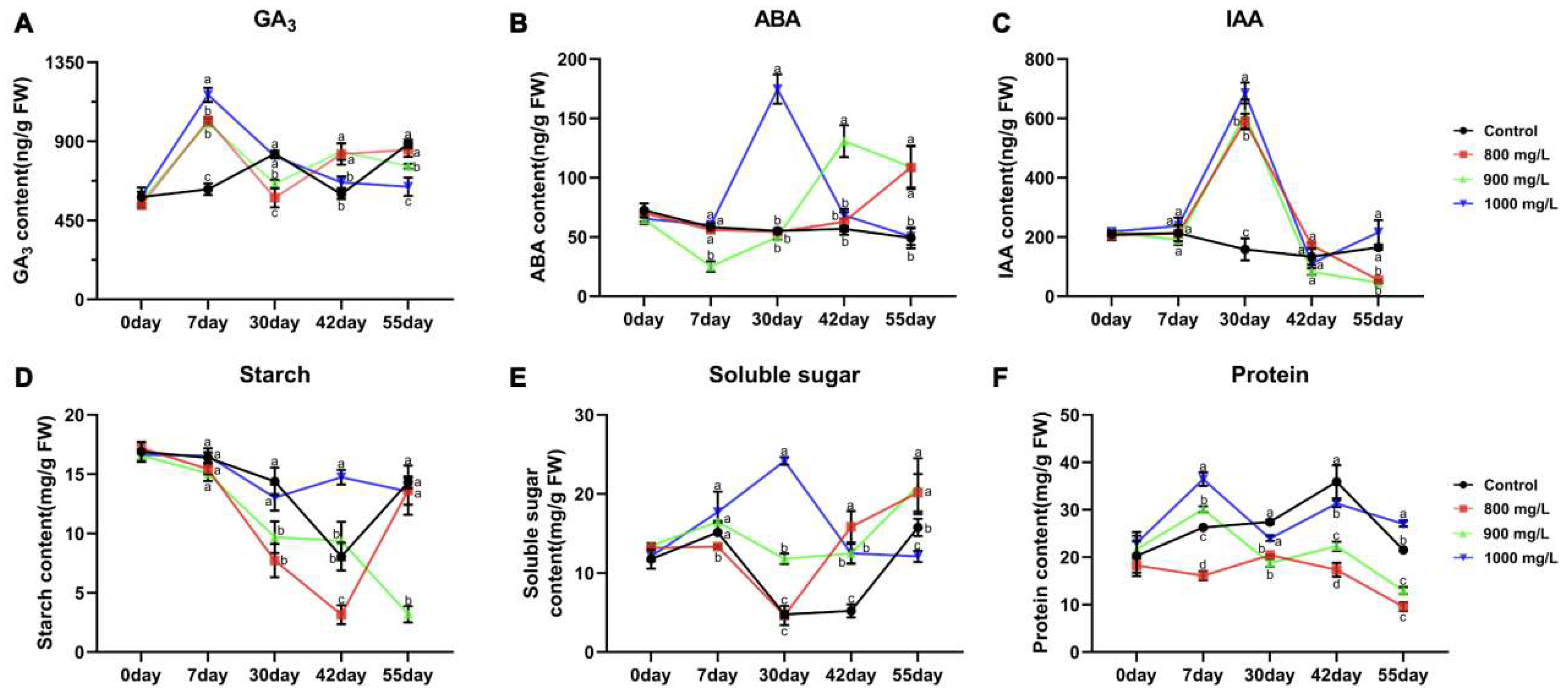
2.2. Analysis of Endogenous Hormone Content Changes
2.3. Analysis of Physiological Index Changes
2.4. Transcriptome Sequencing of Tree Peony Flower Buds Under Different Treatments
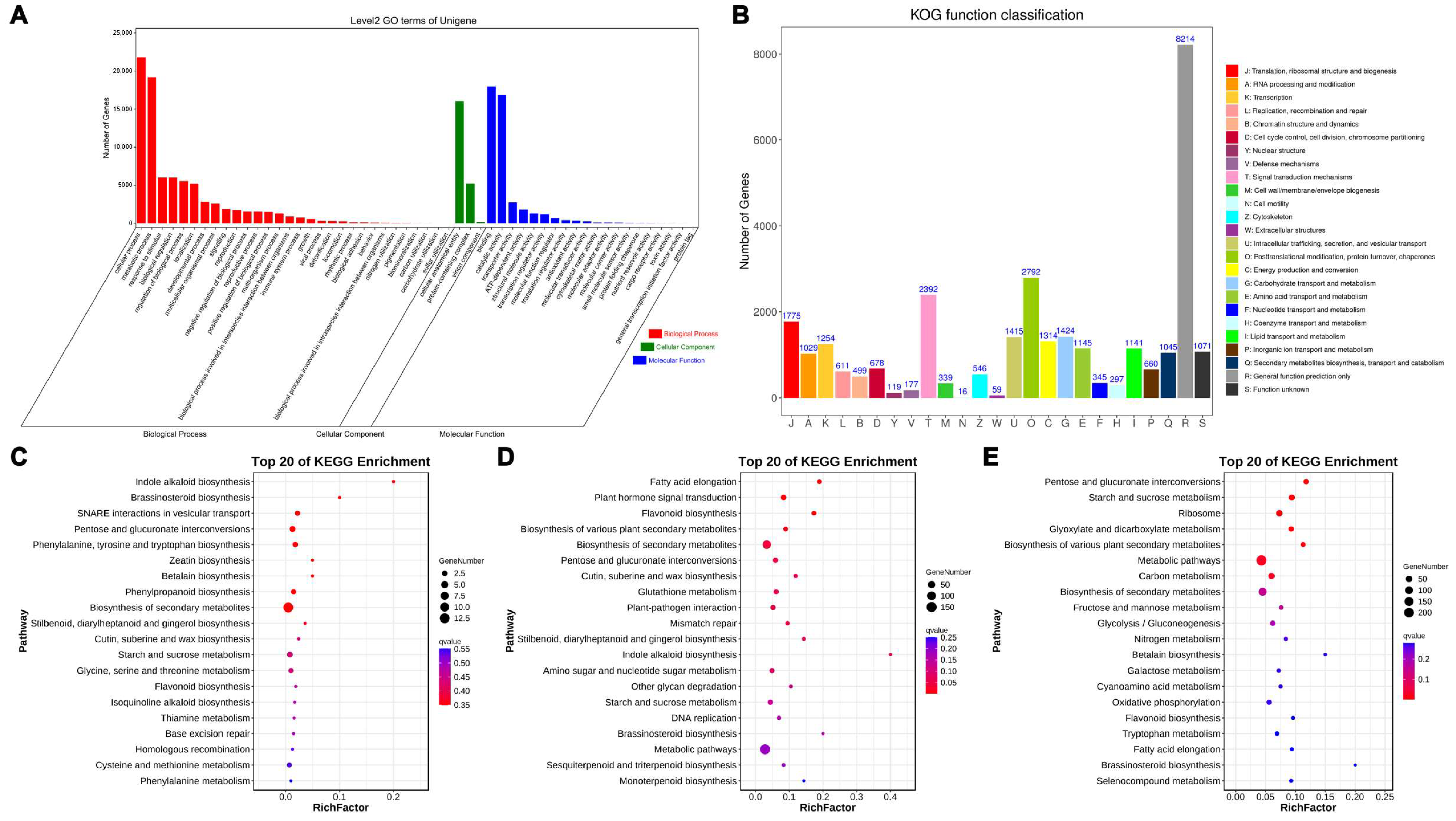
2.5. GO Enrichment and KEGG Pathway Analysis
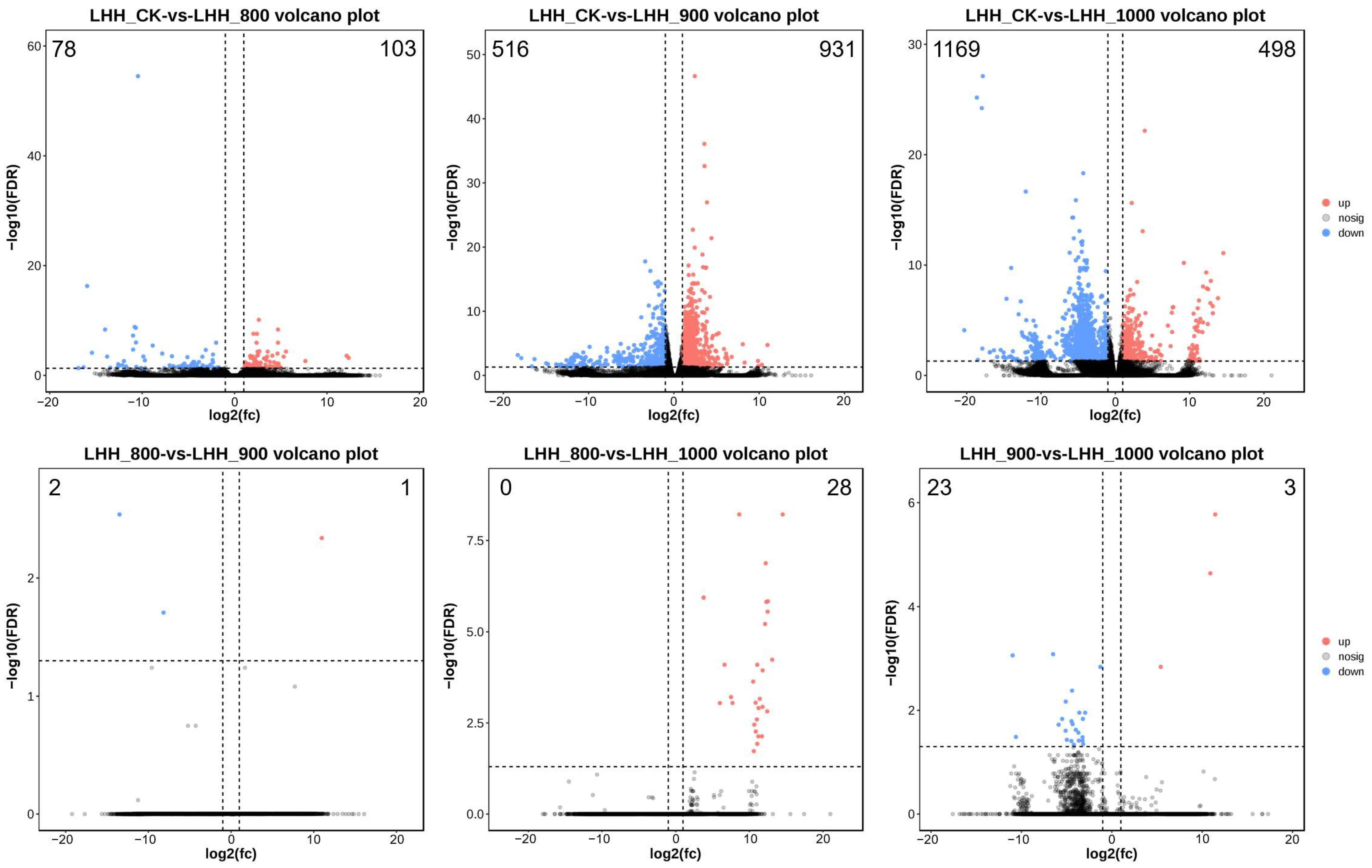
2.6. Analysis of DEGs in Tree Peony Under Different Treatments
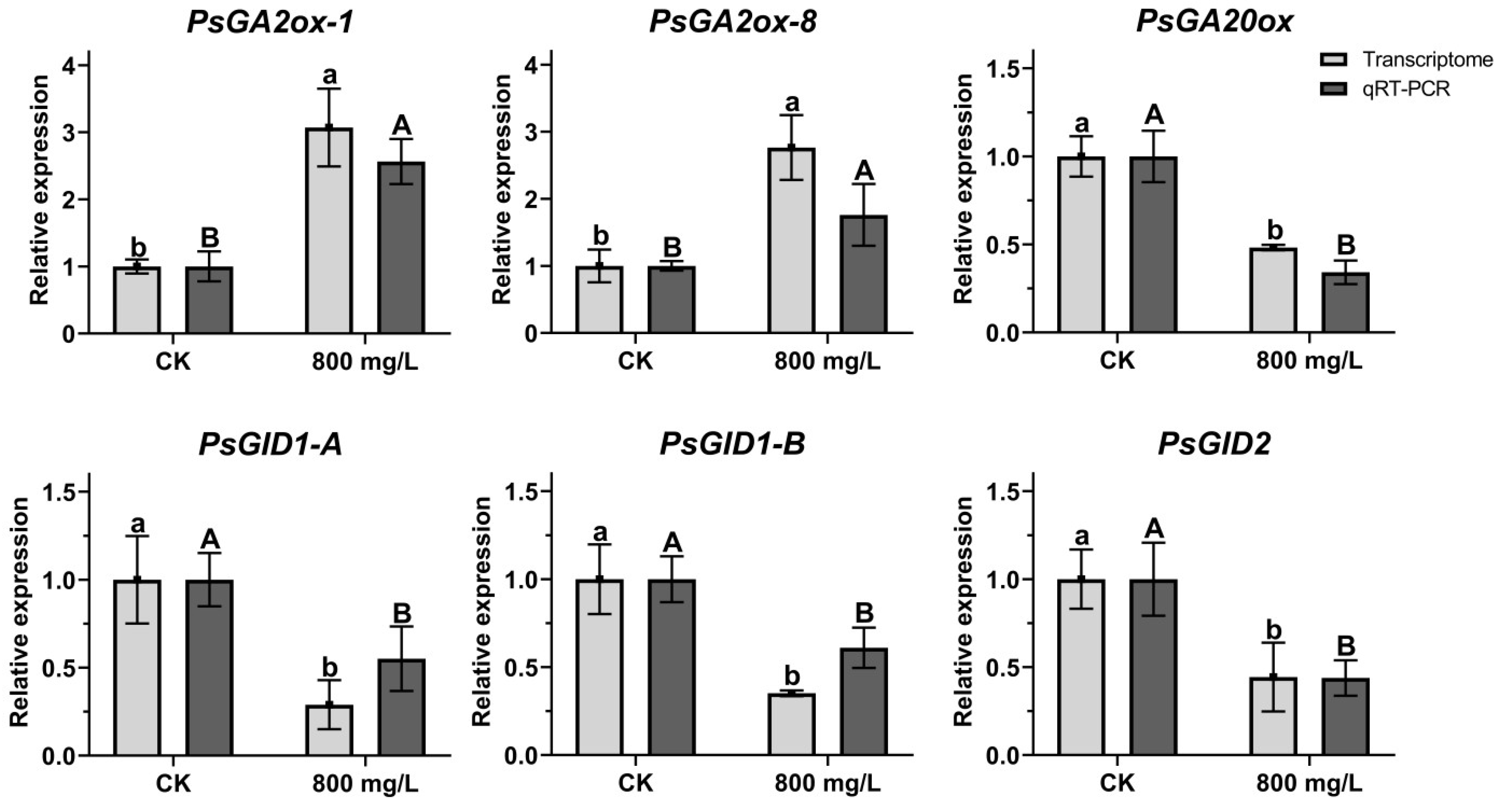
2.7. Verification of Relative Gene Expression During Flower Opening Process
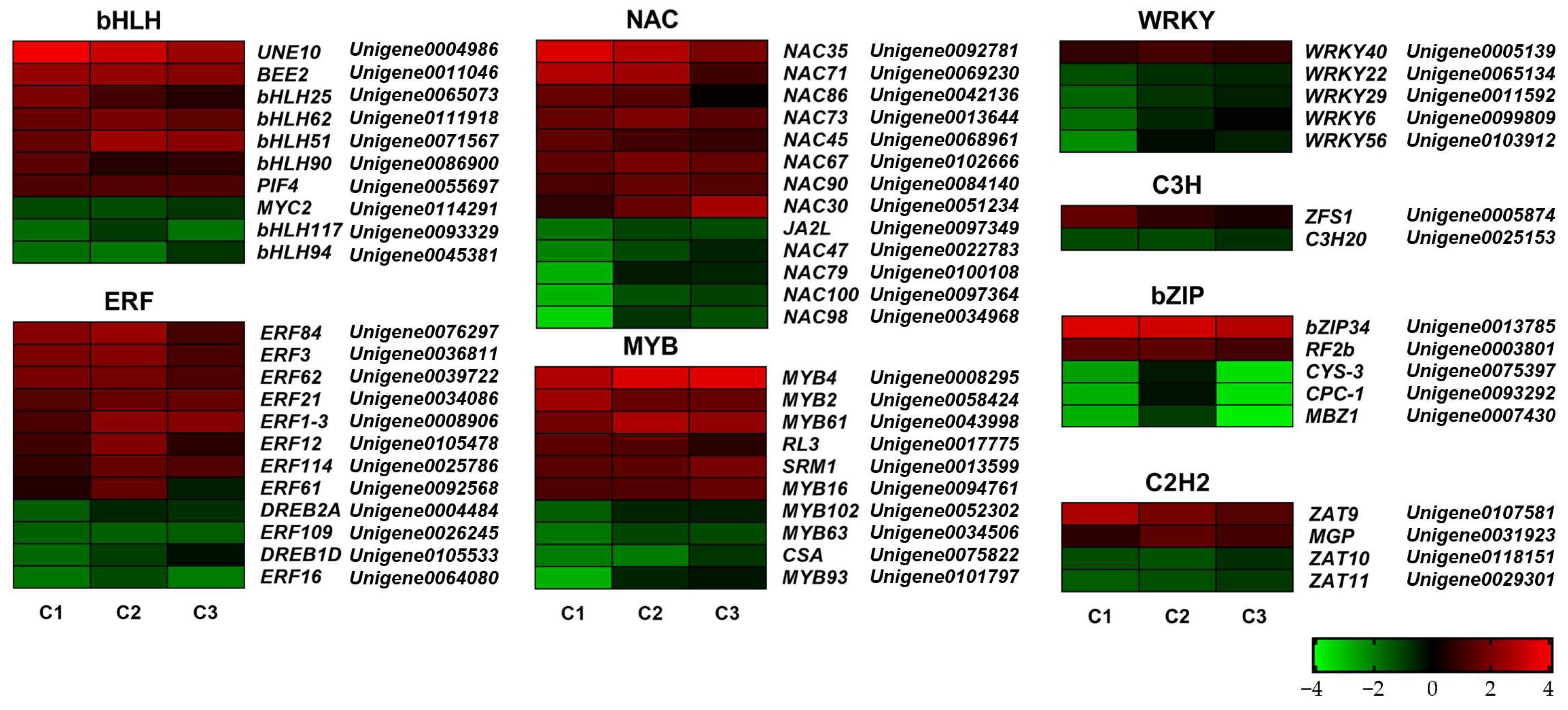
2.8. Analysis of DEGs in Plant Hormone Signal Transduction Pathways Under Different Treatments
2.9. Analysis of Gene Expression Related to GA Pathway
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatment Methods
4.2. Morphological Measurements
4.3. Physiological Measurements
4.4. RNA Extraction, Library Construction and Sequencing
4.5. Analysis of Differentially Expressed Genes (DEGs) and Enrichment
4.6. RNA Extraction for Quantitative Real-Time PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamenetsky-Goldstein, R.; Yu, X. Cut peony industry: The first 30 years of research and new horizons. Hortic. Res. 2022, 9, uhac079. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.Z.; Chao, X.; Jing, L.; Yan, Y.; Ping, W.; Tao, C.; Hong, D. Out of the Pan-Himalaya: Evolutionary history of the Paeoniaceae revealed by phylogenomics. J. Syst. Evol. 2021, 59, 1170–1182. [Google Scholar]
- Zhang, R. King of flowers: Reinterpretation of Chinese peonies in early modern Europe. J. South. Assoc. Hist. Med. Sci. 2021, 3, 41–54. [Google Scholar]
- Çelikel, G.F.; Doorn, V.G.W. Endogenous ethylene does not regulate opening of unstressed Iris flowers but strongly inhibits it in water-stressed flowers. J. Plant Physiol. 2012, 169, 1425–1429. [Google Scholar] [CrossRef]
- Chen, X.C.; Qin, Y.; Wang, Y.; Hong, W.; Yu, H.D.; Yu, Q.J.; Xiao, F.Z.; Yong, H.L.; Cai, Z.J.; Su, S.G.; et al. Ethylene-regulated asymmetric growth of the petal base promotes flower opening in rose (Rosa hybrida). Plant Cell 2021, 33, 1229–1251. [Google Scholar] [CrossRef]
- Kalam, A.A.; Shahabuddin, S.; Ishikawa, T.; Shibata, H. Phosphorylation of plasma membrane aquaporin regulates temperature-dependent opening of tulip petals. Plant Cell Physiol. 2004, 45, 608–617. [Google Scholar]
- Rizzo, J.; Harampolis, A. The Flower Recipe Book; Artisan: New York, NY, USA, 2013. [Google Scholar]
- Bouza-Morcillo, L.; Marquez, J.; Ghita, M.; Sánchez-Montesino, R.; Duran-Nebreda, S.; Gómez, L.; Holdsworth, M.J.; Bassel, G.; Oñate-Sánchez, L. A regulatory module controlling GA-mediated endosperm cell expansion is critical for seed germination in Arabidopsis. Mol. Plant 2019, 12, 71–85. [Google Scholar]
- Shahnejat-Bushehri, S.; Tarkowska, D.; Sakuraba, Y.; Balazadeh, S. Arabidopsis NAC transcription factor JUB1 regulates GA/BR metabolism and signalling. Nat. Plants 2016, 2, 16013. [Google Scholar] [CrossRef]
- Su, G.; Lin, Y.; Wang, C.; Lu, J.; Liu, Z.; He, Z.; Shu, X.; Chen, W.; Wu, R.; Li, B.; et al. Expansin SlExp1 and endoglucanase SlCel2 synergistically promote fruit softening and cell wall disassembly in tomato. Plant Cell 2024, 36, 709–726. [Google Scholar] [CrossRef]
- Song, C.; Shan, W.; Kuang, J.; Chen, J.; Lu, W. The basic helix-loop-helix transcription factor MabHLH7 positively regulates cell wall-modifying-related genes during banana fruit ripening. Postharvest Biol. Technol. 2020, 161, 111068. [Google Scholar] [CrossRef]
- Xue, J.; Li, T.; Wang, S.; Xue, Y.; Liu, X.; Zhang, X. Defoliation and gibberellin synergistically induce tree peony flowering with non-structural carbohydrates as intermedia. J. Plant Physiol. 2018, 233, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Li, T.; Wang, S.; Wang, S.; Xue, Y.; Hu, F.; Zhang, X. Elucidation of the mechanism of reflowering in tree peony (Paeonia suffruticosa) ‘Zi Luo Lan’ by defoliation and gibberellic acid application. Plant Physiol. Biochem. 2018, 132, 571–578. [Google Scholar]
- Zhang, J.; Wang, X.; Zhang, D.; Qiu, S.; Wei, J.; Guo, J.; Li, D.; Xia, Y. Evaluating the Comprehensive Performance of Herbaceous Peonies at Low Latitudes by the Integration of Long-running Quantitative Observation and Multi-Criteria Decision Making Approach. Sci. Rep. 2019, 9, 15079. [Google Scholar]
- Cohen, M.; Eitan, R.; Din, G.Y.; Kamenetsky, R. Effect of constant and alternating temperature regimes on post-dormancy development of herbaceous peony. Acta Hortic. 2017, 1171, 89–98. [Google Scholar]
- Hosoki, T.; Hamada, M.; Inaba, K. Forcing of Tree Peony for December Shipping by Pre-chilling and Chemical Treatments. J. Jpn. Soc. Hortic. Sci. 1984, 53, 187–193. [Google Scholar]
- Kumar, M.; Kesawat, S.M.; Ali, A.; Lee, S.; Gill, S.S.; Kim, H.U. Integration of Abscisic Acid Signaling with Other Signaling Pathways in Plant Stress Responses and Development. Plants 2019, 8, 592. [Google Scholar] [CrossRef]
- Sheng, J.B.; Chang, M.H.; Li, S.S.; Hao, Y. New insights into gibberellin signaling in regulating flowering in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 118–131. [Google Scholar]
- Mara, C.; Alex, C.; William, J.C.; Lucia, C. Auxin and Flower Development: A Blossoming Field. Cold Spring Harb. Perspect. Biol. 2020, 13, a039974. [Google Scholar]
- Hatamzadeh, A.; Shafyii-Masoule, S.; Samizadeh, H. Gibberellin Effects on Development Process from Visible Bud to Flowering in Lilium. Hortic. Environ. Biotechnol. 2010, 51, 449–452. [Google Scholar]
- Yu, Y.Y.; Ji, L.; Bei, Y.G.; Lin, L.; Ma, G.H.; Wu, K.L.; Yang, F.X.; Zhu, G.F.; Fang, L.; Zeng, S.J. Exogenous GA3 promotes flowering in Paphiopedilum callosum (Orchidaceae) through bolting and lateral flower development regulation. Hortic. Res. 2022, 9, uhac091. [Google Scholar]
- Lin, S.Y.; Agehara, S. Exogenous Gibberellic Acid and Cytokinin Effects on Budbreak, Flowering, and Yield of Blackberry Grown under Subtropical Climatic Conditions. HortScience 2020, 55, 1938–1945. [Google Scholar] [CrossRef]
- Mornya, M.P.; Cheng, F. Effect of Combined Chilling and GA3 Treatment on Bud Abortion in Forced ‘Luoyanghong’ Tree Peony (Paeonia suffruticosa Andr.). Hortic. Plant J. 2018, 4, 250–256. [Google Scholar] [CrossRef]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Naveen, K.; Chand, G.S.; Kiran, D. Flower bud opening and senescence in roses (Rosa hybrida L.). Plant Growth Regul. 2008, 55, 81–99. [Google Scholar]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef]
- Li, L.F.; Zhang, W.B.; Zhang, L.L.; Li, N.; Peng, J.Z.; Wang, Y.Q.; Zhong, C.M.; Yang, Y.P.; Sun, S.L.; Liang, S.; et al. Transcriptomic insights into antagonistic effects of gibberellin and abscisic acid on petal growth in Gerbera hybrida. Front. Plant Sci. 2015, 6, 168. [Google Scholar] [CrossRef]
- Khan, I.M.; Giridhar, P. Plant betalains: Chemistry and biochemistry. Phytochemistry 2015, 117, 267–295. [Google Scholar] [CrossRef]
- Yoshikazu, T.; Nobuhiro, S.; Akemi, O. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. PlantJ. Cell Mol. Biol. 2008, 54, 733–749. [Google Scholar]
- Ballerini, E.S.; Min, Y.; Edwards, M.B.; Kramer, E.M.; Hodges, S.A. POPOVICH, encoding a C2H2 zinc-finger transcription factor, plays a central role in the development of a key innovation, floral nectar spurs, in Aquilegia. Proc. Natl. Acad. Sci. USA 2020, 117, 22552–22560. [Google Scholar] [CrossRef]
- Hongna, Z.; Xiaolu, P.; Shenghui, L.; Lin, W.Q.; Li, Y.H.; Zhang, X.M. Genome-wide analysis of AP2/ERF transcription factors in pineapple reveals functional divergence during flowering induction mediated by ethylene and floral organ development. Genomics 2020, 113, 474–489. [Google Scholar]
- Xiao, J.X.; Jia, F.J.; Yao, Y.H.; Zhang, Z.X.; Song, A.P.; Ding, L.; Wang, H.B.; Yao, J.J.; Chen, S.M.; Chen, F.D.; et al. The Constitutive Expression of a Chrysanthemum ERF Transcription Factor Influences Flowering Time in Arabidopsis thaliana. Mol. Biotechnol. 2019, 61, 20–31. [Google Scholar]
- Ito, S.; Song, Y.H.; Josephson-Day, A.R.; Miller, R.J.; Breton, G.; Olmstead, R.G.; Imaizumi, T. FLOWERING BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 3582–3587. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Y.; Pan, J.; Lou, D.J.; Hu, Y.R.; Yu, D.Q. The bHLH Transcription Factors MYC2, MYC3, and MYC4 Are Required for Jasmonate-Mediated Inhibition of Flowering in Arabidopsis. Mol. Plant 2017, 10, 1461–1464. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, B.; Qiu, Z.; Lu, Z.Y.; Hang, Z.Y.; Wu, F.; Chen, X.; Zhu, X.T. Genome-wide identification of MYC transcription factors and their potential functions in the growth and development regulation of tree peony (Paeonia suffruticosa). Plants 2024, 13, 437. [Google Scholar] [CrossRef]
- Li, C.Z.; Guo, X.L.; Ji, Z.J.; Zhao, G.Y.; Xia, C.; Zhang, L.; Li, F.; Zhang, Q.; Dong, C.H.; Gao, S.C.; et al. The wheat MYB-related transcription factor TaMYB72 promotes flowering in rice. J. Integr. Plant Biol. 2016, 58, 701–704. [Google Scholar]
- Yu, T.L.; Zi, J.C.; Yu, H.T.; Sun, J.; Meng, J.S.; Tao, J.; Zhao, D.Q. Tree peony PsMYB44 negatively regulates petal blotch distribution by inhibiting dihydroflavonol-4-reductase gene expression. Ann. Bot. 2022, 131, 323–334. [Google Scholar]
- Nan, H.; Cao, D.; Zhang, D.; Li, Y.; Lu, S.J.; Tang, L.L.; Yuan, X.H.; Liu, B.H.; Kong, F.J. GmFT2a and GmFT5a redundantly and differentially regulate flowering through interaction with and upregulation of the bZIP transcription factor GmFDL19 in soybean. PLoS ONE 2017, 9, e97669. [Google Scholar] [CrossRef] [PubMed]
- Vroemen, C.W.; Mordhorst, A.P.; Albrecht, C.; Kwaaitaal, M.A.; Vries, S.C. The CUP-SHAPED COTYLEDON3 gene is required for boundary and shoot meristem formation in Arabidopsis. Plant Cell 2003, 15, 1563–1577. [Google Scholar] [CrossRef]
- Zhong, J.; Powell, S.; Preston, J.C. Organ boundary NAC-domain transcription factors are implicated in the evolution of petal fusion. Plant Biol. 2016, 18, 893–902. [Google Scholar] [CrossRef]
- Yan, H.Y.; Zhen, H.L.; Long, W.; Kim, S.; Seo, P.J.; Qiao, M.; Wang, N.; Li, S.; Cao, X.F.; Park, C.; et al. WRKY71 accelerates flowering via the direct activation of FLOWERING LOCUS T and LEAFY in Arabidopsis thaliana. Plant J. 2016, 85, 96–106. [Google Scholar]
- Liu, H.; Huang, R.; Ma, J.; Sui, S.Z.; Guo, Y.L.; Liu, D.F.; Li, Z.N.; Lin, Y.C.; Li, M.Y. Two C3H Type Zinc Finger Protein Genes, CpCZF1 and CpCZF2, from Chimonanthus praecox Affect Stamen Development in Arabidopsis. Genes 2017, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Chen, Y.; Xu, J.; Liu, Y.; Zhang, H.; Li, R. Progress in the Study of Plant Gibberellin Oxidase and Its Functions. Biotechnol. Bull. 2024. [Google Scholar] [CrossRef]
- Ueguchi-Tanaka, M.; Ashikari, M.; Nakajima, M.; Itoh, H.; Katoh, E.; Kobayashi, M.; Chow, T.; Hsing, Y.C.; Kitano, H.; Yamaguchi, I.; et al. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin. Nature 2005, 437, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Hirano, Y.; Sun, T.-P.; Hakoshima, T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 2008, 456, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Shimada, A.; Ueguchi-Tanaka, M.; Nakatsu, T.; Nakajima, M.; Naoe, Y.; Ohmiya, H.; Kato, H.; Matsuoka, M. Structural basis for gibberellin recognition by its receptor GID1. Nature 2008, 456, 520–523. [Google Scholar] [CrossRef]
- Griffiths, J.; Murase, K.; Rieu, I.; Zentella, R.; Zhang, Z.; Powers, S.J.; Gong, F.; Phillips, A.L.; Hedden, P.; Sun, T.; et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 2006, 18, 3399–3414. [Google Scholar] [CrossRef]
- Willige, B.C.; Ghosh, S.; Nill, C.; Zourelidou, M.; Dohmann, E.M.N.; Maier, A.; Schwechheimer, C. The DELLA domain of GA INSENSITIVE mediates the interaction with the ga insensitive dwarf1a gibberellin receptor of Arabidopsis. Plant Cell 2007, 19, 1209–1220. [Google Scholar] [CrossRef]
- Dill, A.; Thomas, S.G.; Hu, J.; Steber, C.M.; Sun, T.-P. The Arabidopsis F-Box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation. Plant Cell 2004, 16, 1392–1405. [Google Scholar] [CrossRef]
- Achard, P.; Herr, A.; Baulcombe, D.C.; Harberd, N.P. Modulation of floral development by a gibberellin-regulated microRNA. Development 2004, 131, 3357–3365. [Google Scholar] [CrossRef]
- Gocal, G.F.; Sheldon, C.C.; Gubler, F.; Moritz, T.; Bagnall, D.J.; MacMillan, C.P.; Li, S.F.; Parish, R.W.; Dennis, E.S.; Weigel, D.; et al. GAMYB-like genes, flowering, and gibberellin signaling in Arabidopsis. Plant Physiol. 2001, 127, 1682–1693. [Google Scholar]
- Blázquez, M.A.; Green, R.; Nilsson, O.; Sussman, M.R.; Weigel, D. Gibberellins promote flowering of Arabidopsis by activating the LEAFY promoter. Plant Cell 1998, 10, 791–800. [Google Scholar] [CrossRef]
- Lee, H.; Suh, S.-S.; Park, E.; Cho, E.; Ahn, J.H.; Kim, S.; Lee, J.S.; Kwon, Y.M.; Lee, I. The AGAMOUS-LIKE 20 MADS domain protein integrates floral inductive pathways in Arabidopsis. Genes Dev. 2000, 14, 2366–2376. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Shi, J.; Xu, H.W.; Zhang, S.; Huo, Z.; Wang, J. Changes of protein and proline content in different types of tree peony during flowering stage. J. Henan Inst. Sci. Technol. 2012, 40, 28–31. [Google Scholar]
- Wen, X.; Xin, G.; Li, Y.X. Study on the determination of total sugar in potato starch by anthrone colorimetry. Food Res. Dev. 2013, 17, 86–88. [Google Scholar]
- Guo, Y.H.; Yang, W.H.; Yu, Y.Z.; Xu, H.X.; Wang, Y.Q.; Zhou, D.Y.; Kuang, M. Determination of water soluble total sugar content in cotton mature fiber by anthrone colorimetry. China Cotton 2011, 38, 23–26. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotypic Observation of Flower Buds | ||||
|---|---|---|---|---|
| Treatment | Date of Floral Bud Initiation Floral Bud Diameter (cm) | Floral Bud Development Period Floral Bud Diameter (cm) | Bell-Shaped Bud Stage Bell-Shaped Bud Diameter (cm) | First Flowering Date Flower Diameter (cm) |
| Treatment 1 | 13 d, 0.83 ± 0.06 a | 22 d, 0.97 ± 0.03 a | 30 d, 3.40 ± 0.03 a | 42 d, 15.47 ± 0.682 a |
| Treatment 2 | 13 d, 0.90 ± 0.00 a | 22 d, 1.07 ± 0.06 a | 33 d, 2.80 ± 0.26 b | 45 d, 14.65 ± 0.679 a |
| Treatment 3 | 13 d, 0.90 ± 0.00 a | 22 d, 0.90 ± 0.02 a | −1.82 ± 0.08 c | - |
| CK | 13 d, 0.87 ± 0.06 a | 30 d, 0.87 ± 0.06 a | −1.50 ± 0.20 c | - |
| Tree Peony Growth Parameters | ||||||
|---|---|---|---|---|---|---|
| Vegetative Growth | Reproductive Growth | |||||
| Treatment | Leaf Width(cm) | Number of Compound Leaves | New Shoot Length (cm) | Total Number of Flowers | Flowering Rate | Flower Diameter (cm) |
| Treatment 1 | 4.99 ± 0.735 a | 210.21 ± 71.012 ab | 11.74 ± 2.063 a | 16 | 0.137 ± 0.152 a | 15.47 ± 0.682 a |
| Treatment 2 | 4.6 ± 0.893 ab | 287.14 ± 87.441 a | 9.38 ± 2.704 ab | 22 | 0.186 ± 0.128 a | 14.65 ± 0.679 a |
| Treatment 3 | 4.05 ± 0.512 bc | 290.14 ± 114.988 a | 9.81 ± 2.299 ab | 2 | 0.017 ± 0.043 b | - |
| CK | 3.49 ± 0.551 c | 181.07 ± 74.523 b | 7.66 ± 1.918 b | 1 | 0.01 ± 0.037 b | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Wang, Q.; Qiu, Z.; Lu, Z.; Zhang, J.; He, Q.; Yang, J.; Zhang, H.; Zhu, X.; Chen, X. Transcriptomic Analysis of Gibberellin-Mediated Flower Opening Process in Tree Peony (Paeonia suffruticosa). Plants 2025, 14, 1002. https://doi.org/10.3390/plants14071002
Li B, Wang Q, Qiu Z, Lu Z, Zhang J, He Q, Yang J, Zhang H, Zhu X, Chen X. Transcriptomic Analysis of Gibberellin-Mediated Flower Opening Process in Tree Peony (Paeonia suffruticosa). Plants. 2025; 14(7):1002. https://doi.org/10.3390/plants14071002
Chicago/Turabian StyleLi, Bole, Qianqian Wang, Zefeng Qiu, Zeyun Lu, Junli Zhang, Qionghua He, Jiajun Yang, Hangyan Zhang, Xiangtao Zhu, and Xia Chen. 2025. "Transcriptomic Analysis of Gibberellin-Mediated Flower Opening Process in Tree Peony (Paeonia suffruticosa)" Plants 14, no. 7: 1002. https://doi.org/10.3390/plants14071002
APA StyleLi, B., Wang, Q., Qiu, Z., Lu, Z., Zhang, J., He, Q., Yang, J., Zhang, H., Zhu, X., & Chen, X. (2025). Transcriptomic Analysis of Gibberellin-Mediated Flower Opening Process in Tree Peony (Paeonia suffruticosa). Plants, 14(7), 1002. https://doi.org/10.3390/plants14071002