

Botanical-Based Strategies for Controlling Xanthomonas spp. in Cotton and Citrus: In Vitro and In Vivo Evaluation
 ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
2. Results
2.1. Plant Species Data Collection and Natural Product Yields
2.2. Characterization of Xanthomonas citri Strains
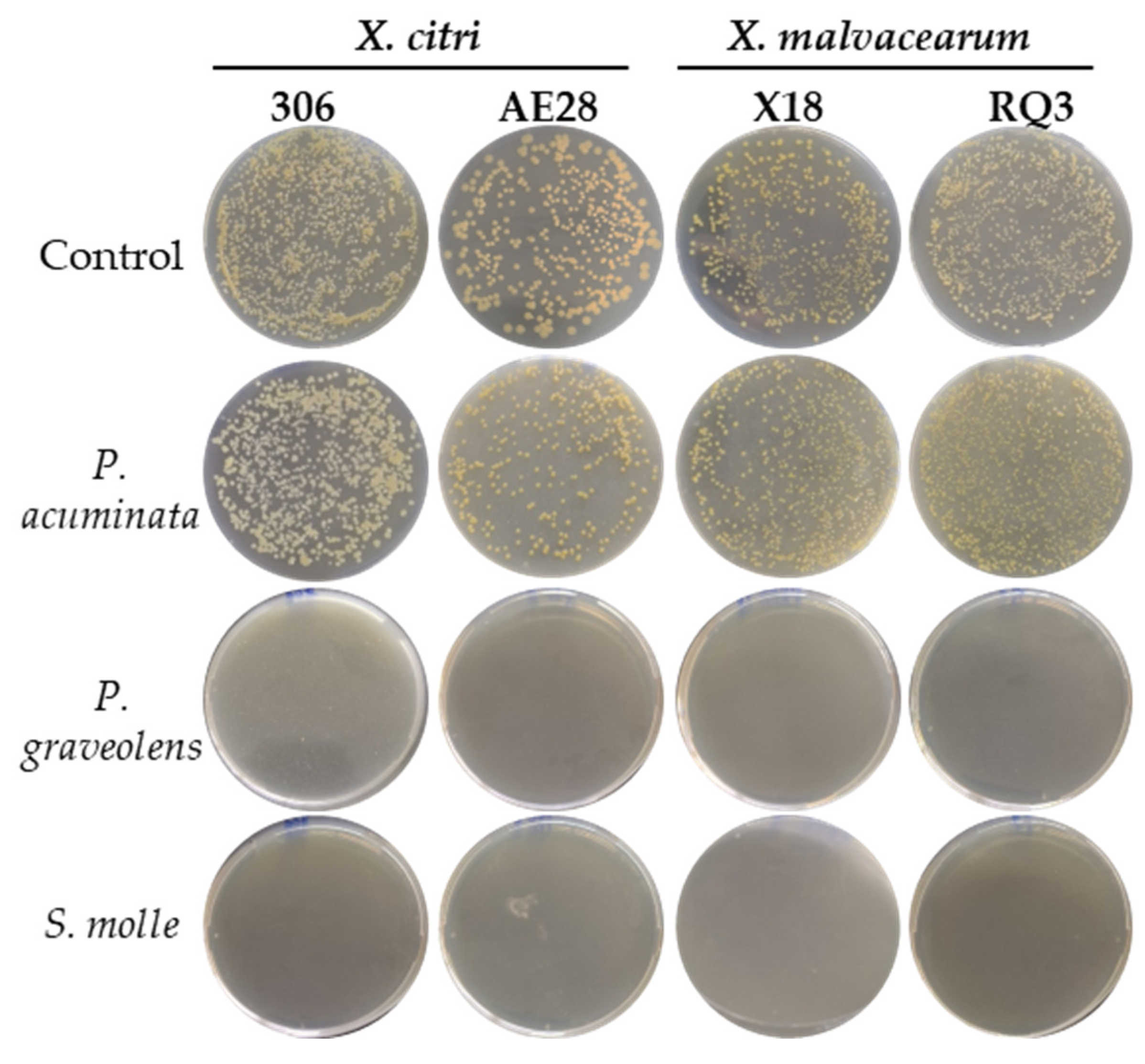
2.3. Effects of Natural Plant Products on Xanthomonas Strains Growth In Vitro
2.4. Effects of S. molle and P. graveolens Essential Oils on Citrus Canker and Cotton Bacterial Blight Disease
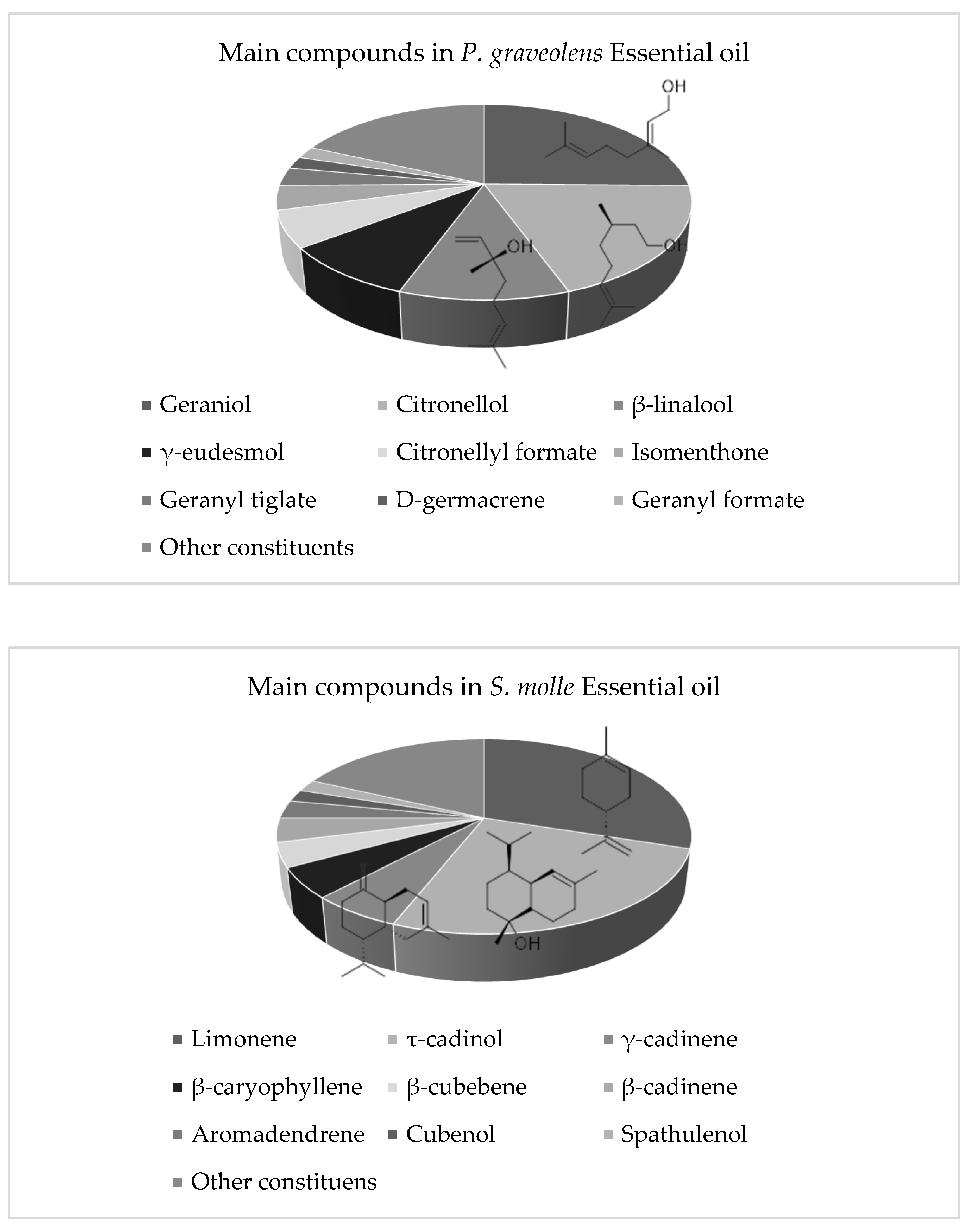
2.5. Phytochemical Profile of Active Natural Products
3. Discussion
4. Materials and Methods
4.1. Plant Material and Natural Products Obtaining
4.2. Bacterial Strains, Culture Media, and Growth Conditions
4.3. Molecular Identification of X. citri Regional Strains
4.4. In Vitro Susceptibility Test
4.5. Preventive and Curative Disease Control with Essential Oils
4.6. Chemical Profile of the Active Natural Products
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Citrus: World Markets and Trade. Available online: https://apps.fas.usda.gov/psdonline/circulars/citrus.pdf?utm_source (accessed on 12 February 2025).
- Khan, M.A.; Wahid, A.; Ahmad, M.; Tahir, M.T.; Ahmed, M.; Ahmad, S.; Hasanuzzaman, M. World cotton production and consumption: An overview. In Cotton Production and Uses: Agronomy, Crop Protection, and Postharvest Technologies; Ahmad, S., Hasanuzzaman, M., Eds.; Springer: Singapore, 2020; pp. 1–7. [Google Scholar]
- The Argentine Citrus Industry. Available online: https://www.federcitrus.org/wp-content/uploads/2024/11/Federcitrus-Actividad-Citricola-2024.pdf (accessed on 12 February 2025).
- Scarpin, G.J.; Dileo, P.N.; Winkler, H.M.; Cereijo, A.E.; Lorenzini, F.G.; Muchut, R.J.; Roeschlin, R.A.; Acuña, C.; Paytas, M. Genetic progress in cotton dry matter partitioning in Argentina. Ind. Crop. Prod. 2025, 225, 120419. [Google Scholar] [CrossRef]
- An, S.Q.; Potnis, N.; Dow, M.; Vorhölter, F.J.; He, Y.Q.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Rev. 2020, 44, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, D.; Gottwald, T.; Lopes, S.A.; Wulff, N.A. Bacterial pathogens of citrus: Citrus canker, citrus variegated chlorosis, and Huanglongbing. In The Genus Citrus; Talon, M., Caruso, M., Fred, G., Gmitter, F.G., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 371–389. [Google Scholar]
- Delannoy, E.; Lyon, B.; Marmey, P.; Jalloul, A.; Montillet, J.; Daniel, J.; Essenberg, M.; Nicole, M. Resistance of cotton to Xanthomonas campestris pv. malvacearum. Annu. Rev. Phytopathol. 2005, 43, 62–82. [Google Scholar]
- Favaro, M.A.; Roeschlin, R.A.; Ribero, G.G.; Maumary, R.L.; Fernandez, L.N.; Lutz, A.; Sillon, M.; Rista, L.M.; Marano, M.R.; Gariglio, N.F. Relationships between copper content in orange leaves, bacterial biofilm formation, and citrus canker disease control after different copper treatments. Crop Prot. 2017, 92, 182–189. [Google Scholar] [CrossRef]
- Favaro, M.A.; Molina, M.C.; Roeschlin, R.A.; Gadea, J.; Gariglio, N.F.; Marano, M.R. Different responses in mandarin cultivars uncover a role of cuticular waxes in the resistance to citrus canker. Phytopathology 2020, 110, 1791–1801. [Google Scholar] [CrossRef]
- Behlau, F.; Gochez, A.M.; Jones, J.B. Diversity and copper resistance of Xanthomonas affecting citrus. Trop. Plant Pathol. 2020, 45, 200–212. [Google Scholar] [CrossRef]
- Mačionienė, I.; Čepukoit, D.; Šalomskienė, J.; Černauskas, D.; Burokienė, D.; Šalaševičienė, A. Effects of natural antimicrobials on Xanthomonas strains growth. Horticulturae 2021, 8, 7. [Google Scholar] [CrossRef]
- Zhang, J.; Elassbli, H.; Zhu, Y.; Wheeler, T.; Bourland, F. Evaluation methods, resistant germplasm, and breeding for resistance to bacterial blight in cotton: A review. J. Cotton Sci. 2024, 28, 27–57. [Google Scholar] [CrossRef]
- Chiesa, M.A.; Siciliano, M.F.; Ornella, L.; Roeschlin, R.A.; Favaro, M.A.; Delgado, N.P.; Sendín, L.N.; Orce, I.G.; Ploper, L.D.; Vojnov, A.A.; et al. Characterization of a variant of Xanthomonas citri subsp. citri that triggers a host-specific defense response. Phytopathology 2013, 103, 555–564. [Google Scholar]
- Roeschlin, R.A.; Favaro, M.A.; Chiesa, M.A.; Alemano, S.; Vojnov, A.A.; Castagnaro, A.P.; Filippone, M.P.; Gmitter, F.G.; Gadea, J.; Marano, M.R. Resistance to citrus canker induced by a variant of Xanthomonas citri ssp. citri is associated with a hypersensitive cell death response involving autophagy-associated vacuolar processes. Mol. Plant Pathol. 2017, 18, 1267–1281. [Google Scholar]
- Huang, X.; Zhai, J.; Luo, Y.; Rudolph, K. Identification of a highly virulent strain of Xanthomonas axonopodis pv. malvacearum. Eur. J. Plant Pathol. 2008, 122, 461–469. [Google Scholar] [CrossRef]
- Phillips, A.Z.; Berry, J.C.; Wilson, M.C.; Vijayaraghavan, A.; Burke, J.; Bunn, J.I.; Bart, R.S. Genomics-enabled analysis of the emergent disease cotton bacterial blight. PLoS Genet. 2017, 13, e1007003. [Google Scholar] [CrossRef]
- Chavhan, R.L.; Mondal, K.K.; Karuppayil, S.M.; Chakrabarty, P.K. Evolution of biotypes within race 18 population of Xanthomonas citri subsp. malvacearum and their predominance in Indian cotton belts. Physiol. Mol. Plant Pathol. 2021, 116, 101721. [Google Scholar] [CrossRef]
- Di Liberto, M.; Stegmayer, M.I.; Svetaz, L.; Derita, M. Evaluation of Argentinean medicinal plants and isolation of their bioactive compounds as an alternative for the control of postharvest fruits phytopathogenic fungi. Br. J. Pharmacog. 2019, 29, 686–689. [Google Scholar] [CrossRef]
- Basaid, K.; Chebli, B.; Mayad, E.H.; Furze, J.N.; Bouharroud, R.; Krier, F.; Paulitz, T. Biological activities of essential oils and lipopeptides applied to control plant pests and diseases: A review. Int. J. Pest Manag. 2021, 67, 155–177. [Google Scholar] [CrossRef]
- Naqvi, S.A.H.; Iqbal, S.; Farooq, U.; Hassan, M.Z.; Shahid, M.N.; Noor Shah, A.; Abbas, A.; Mubeen, I.; Farooq, A.; Ghareeb, R.Y.; et al. Evaluation of bacterial perpetuation assays and plant biomolecules antimicrobial activity against cotton blight bacterium Xanthomonas citri subsp. malvacearum; an alternative source for food production and protection. Plants 2022, 11, 1278. [Google Scholar] [PubMed]
- Košćak, L.; Lamovšek, J.; Đermić, E.; Prgomet, I.; Godena, S. Microbial and plant-based compounds as alternatives for the control of phytopathogenic bacteria. Horticulturae 2023, 9, 1124. [Google Scholar] [CrossRef]
- Aslam, M.N.; Khaliq, H.; Zhao, H.; Moosa, A.; Maqsood, A.; Farooqi, M.A.; Bilal, M.S.; Mahmood, T.; Mukhtar, T. Thymol as a Novel Plant-Derived Antibacterial Agent for Suppressing Xanthomonas citri pv. malvacearum in Cotton. Curr. Microbiol. 2025, 82, 99. [Google Scholar] [CrossRef]
- Di Liberto, M.; Seimandi, G.; Fernández, L.; Ruiz, V.; Svetaz, L.; Derita, M. Botanical control of citrus green mold and peach brown rot on fruits assays using a Persicaria acuminata phytochemically characterized extract. Plants 2021, 10, 425. [Google Scholar] [CrossRef]
- Stegmayer, M.I.; Fernández, L.; Alvarez, N.; Olivella, L.; Gutiérrez, H.; Favaro, M.A.; Derita, M. Aceites esenciales provenientes de plantas nativas para el control de hongos fitopatógenos que afectan a frutales. Rev. FAVE-Cienc. Agrar. 2021, 20, 317–329. [Google Scholar] [CrossRef]
- Stegmayer, M.I.; Alvarez, N.H.; Sager, N.; Buyatti, M.; Derita, M.G. Evaluation of Pelargonium graveolens essential oil to prevent gray mold in rose flowers. J. Plant Prot. Res. 2022, 62, 145–152. [Google Scholar]
- Alvarez, N.; Stegmayer, M.I.; Seimandi, G.; Pensiero, J.F.; Zabala, J.M.; Favaro, M.A.; Derita, M.G. Natural Products Obtained from Argentinean Native Plants Are Fungicidal against Citrus Postharvest Diseases. Horticulturae 2023, 9, 562. [Google Scholar] [CrossRef]
- da Silva, A.R.; Ferro, J.A.; Reinach, F.D.C.; Farah, C.S.; Furlan, L.R.; Quaggio, R.B.; Kitajima, J.P. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 2002, 417, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Favaro, M.A.; Micheloud, N.G.; Roeschlin, R.A.; Chiesa, M.A.; Castagnaro, A.P.; Vojnov, A.A.; Gmitter, F.G.; Gadea, J.; Rista, L.M.; Gariglio, N.F.; et al. Surface barriers of mandarin ‘Okitsu’ leaves make a major contribution to canker disease resistance. Phytopathology 2014, 104, 970–976. [Google Scholar] [CrossRef]
- Cunnac, S.; Bolot, S.; Forero Serna, N.; Ortiz, E.; Szurek, B.; Noël, L.D.; Arlat, M.; Jacques, M.A.; Gagnevin, L.; Carrere, S.; et al. High-quality draft genome sequences of two Xanthomonas citri pv. malvacearum strains. Genome Announc. 2013, 1, e00674-13. [Google Scholar]
- dos Santos, J.; Fernandes, C.; Silva, N.; Calefi, G.; Martins, C.; Volpini, G.; Crotti, A.; Ribeiro, A.; Esperandim, T.; Tavares, D.; et al. Volatile compounds of hexane extract from Pterodon pubescens Benth seeds and its significant in vitro potential against different bacterial strains. Nat. Prod. Res. 2025, 39, 1428–1433. [Google Scholar] [CrossRef]
- Ribeiro, A.M.R.; Fernandes, C.C.; Menezes, R.d.P.; Oliveira, A.M.; Gonçalves, D.S.; Martins, C.H.G.; Miranda, M.L.D. Antibacterial screening of hexane extracts from Psidium myrtoides, a Brazilian native plant. Ciência E Nat. 2024, 46, e84178. [Google Scholar] [CrossRef]
- Martin, A.P.; Martínez, M.F.; Chiesa, M.A.; Garcia, L.; Gerhardt, N.; Uviedo, F.; Torres, P.S.; Marano, M.R. Priming crop plants with rosemary (Salvia rosmarinus Spenn, syn Rosmarinus officinalis L.) extract triggers protective defense response against pathogens. Plant Physiol. Biochem. 2023, 197, 107644. [Google Scholar] [CrossRef]
- Derita, M.; Montenegro, I.; Garibotto, F.; Enriz, R.; Cuellar Fritis, M.; Zacchino, S. Structural Requirements for the Antifungal Activities of Natural Drimane Sesquiterpenes and Analogues, Supported by Conformational and Electronic Studies. Molecules 2013, 18, 2029–2051. [Google Scholar] [CrossRef]
- Derita, M.; Leiva, M.; Zacchino, S. Influence of plant part, season of collection and content of the main active constituent, on the antifungal properties of Polygonum acuminatum Kunth. J. Ethnopharmacol. 2009, 124, 377–383. [Google Scholar] [CrossRef]
- Babu, K.G.; Kaul, V.K. Variation in essential oil composition of rose-scented geranium (Pelargonium sp.) distilled by different distillation techniques. Flav. Fragr. J. 2013, 20, 222–231. [Google Scholar] [CrossRef]
- Bouzenna, H.; Krichen, L. Pelargonium graveolens L’Her. and Artemisia arborescens L. essential oils: Chemical composition, antifungal activity against Rhizoctonia solani and insecticidal activity against Rhysopertha dominica. Nat. Prod. Res. 2013, 27, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Lira, M.H.P.D.; Andrade Júnior, F.P.D.; Moraes, G.F.Q.; Macena, G.D.S.; Pereira, F.D.O.; Lima, I.O. Antimicrobial activity of geraniol: An integrative review. J. Essent. Oil Res. 2020, 32, 187–197. [Google Scholar] [CrossRef]
- Pereira, F.D.O.; Mendes, J.M.; Lima, I.O.; Mota, K.S.D.L.; Oliveira, W.A.D.; Lima, E.D.O. Antifungal activity of geraniol and citronellol, two monoterpenes’ alcohols, against Trichophyton rubrum involves inhibition of ergosterol biosynthesis. Pharm. Biol. 2015, 53, 228–234. [Google Scholar] [CrossRef]
- Kačániová, M.; Vukic, M.; Vukovic, N.L.; Čmiková, N.; Verešová, A.; Schwarzová, M.; Garzoli, S. An in-depth study on the chemical composition and biological effects of Pelargonium graveolens essential oil. Foods 2023, 13, 33. [Google Scholar] [CrossRef]
- Gomes, V.; Agostini, G.; Agostini, F.; Atti dos Santos, A.C.; Rossato, M. Variation in the essential oils composition in Brazilian populations of Schinus molle L. (Anacardiaceae). Biochem. Syst. Ecol. 2013, 48, 222–227. [Google Scholar] [CrossRef]
- Do Rosário Martins, M.; Arantes, S.; Candeias, F.; Tinoco, M.T.; Cruz-Morais, J. Antioxidant, antimicrobial and toxicological properties of Schinus molle L. essential oils. J. Ethnopharmacol. 2014, 151, 485–492. [Google Scholar] [CrossRef]
- Do Prado, A.C.; Garces, H.G.; Bagagli, E.; Rall, V.L.M.; Furlanetto, A.; Fernandes Junior, A.; Furtado, F.B. Schinus molle essential oil as a potential source of bioactive compounds: Antifungal and antibacterial properties. J. Appl. Microbiol. 2019, 126, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.L.; Liao, P.C.; Wang, E.I.C.; Su, Y.C. Composition and antifungal activities of the leaf essential oil of Neolitsea parvigemma from Taiwan. Nat. Prod. Commun. 2011, 6, 1357–1360. [Google Scholar] [CrossRef]
- Mahdavi Omran, S.; Moodi, M.A.; Norozian Amiri, S.M.B.; Mosavi, S.J.; Ghazi Mir Saeed, S.A.M. The effects of limonene and orange peel extracts on some spoilage fungi. Int. J. Mol. Clin. Microbiol. 2011, 1, 82–86. [Google Scholar]
- Marei, G.I.K.; Rasoul, M.A.A.; Abdelgaleil, S.A. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pestic. Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]
- da Silva, I.R.R.; Fernandes, C.C.; Gonçalves, D.S.; Martins, C.H.G.; Miranda, M.L.D. Chemical composition and anti-Xanthomonas citri activities of essential oils from Schinus molle L. fresh and dry leaves and of its major constituent spathulenol. Nat. Prod. Res. 2023, 38, 3476–3480. [Google Scholar] [CrossRef] [PubMed]
- Almeida, N.F.; Yan, S.; Cai, R.; Clarke, C.R.; Morris, C.E.; Schaad, N.W.; Vinatzer, B.A. PAMDB, a multilocus sequence typing and analysis database and website for plant-associated microbes. Phytopathology 2010, 100, 208–215. [Google Scholar] [CrossRef] [PubMed]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helminthosporium sativum. J. Agric. Res. 1923, 26, 195–218. [Google Scholar]
- Buttar, D.; Pawar, T.; Grewal, I. Impact of Priaxor (fluxapyroxad 167 g/L + pyraclostrobin 333 g/L SC) on fungal foliar leaf spots in upland cotton. Pl. Dis. Res. 2022, 37, 164–168. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Scientific Name | Voucher Specimen | Collection Data | Part Used | Natural Product Type | Yield (%) |
|---|---|---|---|---|---|
| P. acuminata | MD53 UNL | 19 March 2024, Salado River, Las Colonias, Santa Fe. | Dried leaves | EtOAc extract | 1.80 |
| P. graveolens | MD58 UNL | 26 May 2024, CECIF, Las Colonias, Santa Fe. | Freshly aerial parts | Essential oil | 0.46 |
| S. molle | MD57 UNL | 15 April 2024, FAVE, Las Colonias, Santa Fe. | Freshly aerial parts | Essential oil | 0.72 |
| Species | Strain | Origin | Reference |
|---|---|---|---|
| X. citri subsp. citri | 306, reference | Brazil | da Silva et al., 2002 [27] |
| X. citri subsp. citri | AE28 | Esperanza, Santa Fe, Argentina | Favaro et al., 2014 [28] |
| X. citri subsp. malvacearum | X18, reference | Burkina Faso | Cunnac et al., 2013 [29] |
| X. citri subsp. malvacearum | RQ3 | Reconquista, Santa Fe, Argentina | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roeschlin, R.A.; Favaro, M.A.; Bertinat, B.; Lorenzini, F.G.; Paytas, M.J.; Fernandez, L.N.; Marano, M.R.; Derita, M.G. Botanical-Based Strategies for Controlling Xanthomonas spp. in Cotton and Citrus: In Vitro and In Vivo Evaluation. Plants 2025, 14, 957. https://doi.org/10.3390/plants14060957
Roeschlin RA, Favaro MA, Bertinat B, Lorenzini FG, Paytas MJ, Fernandez LN, Marano MR, Derita MG. Botanical-Based Strategies for Controlling Xanthomonas spp. in Cotton and Citrus: In Vitro and In Vivo Evaluation. Plants. 2025; 14(6):957. https://doi.org/10.3390/plants14060957
Chicago/Turabian StyleRoeschlin, Roxana Andrea, María Alejandra Favaro, Bruno Bertinat, Fernando Gabriel Lorenzini, Marcelo Javier Paytas, Laura Noemí Fernandez, María Rosa Marano, and Marcos Gabriel Derita. 2025. "Botanical-Based Strategies for Controlling Xanthomonas spp. in Cotton and Citrus: In Vitro and In Vivo Evaluation" Plants 14, no. 6: 957. https://doi.org/10.3390/plants14060957
APA StyleRoeschlin, R. A., Favaro, M. A., Bertinat, B., Lorenzini, F. G., Paytas, M. J., Fernandez, L. N., Marano, M. R., & Derita, M. G. (2025). Botanical-Based Strategies for Controlling Xanthomonas spp. in Cotton and Citrus: In Vitro and In Vivo Evaluation. Plants, 14(6), 957. https://doi.org/10.3390/plants14060957