
Impacts of Mechanical Injury on Volatile Emission Rate and Composition in 45 Subtropical Woody Broad-Leaved Storage and Non-Storage Emitters

 ,
,  and
and
Abstract
1. Introduction
2. Results
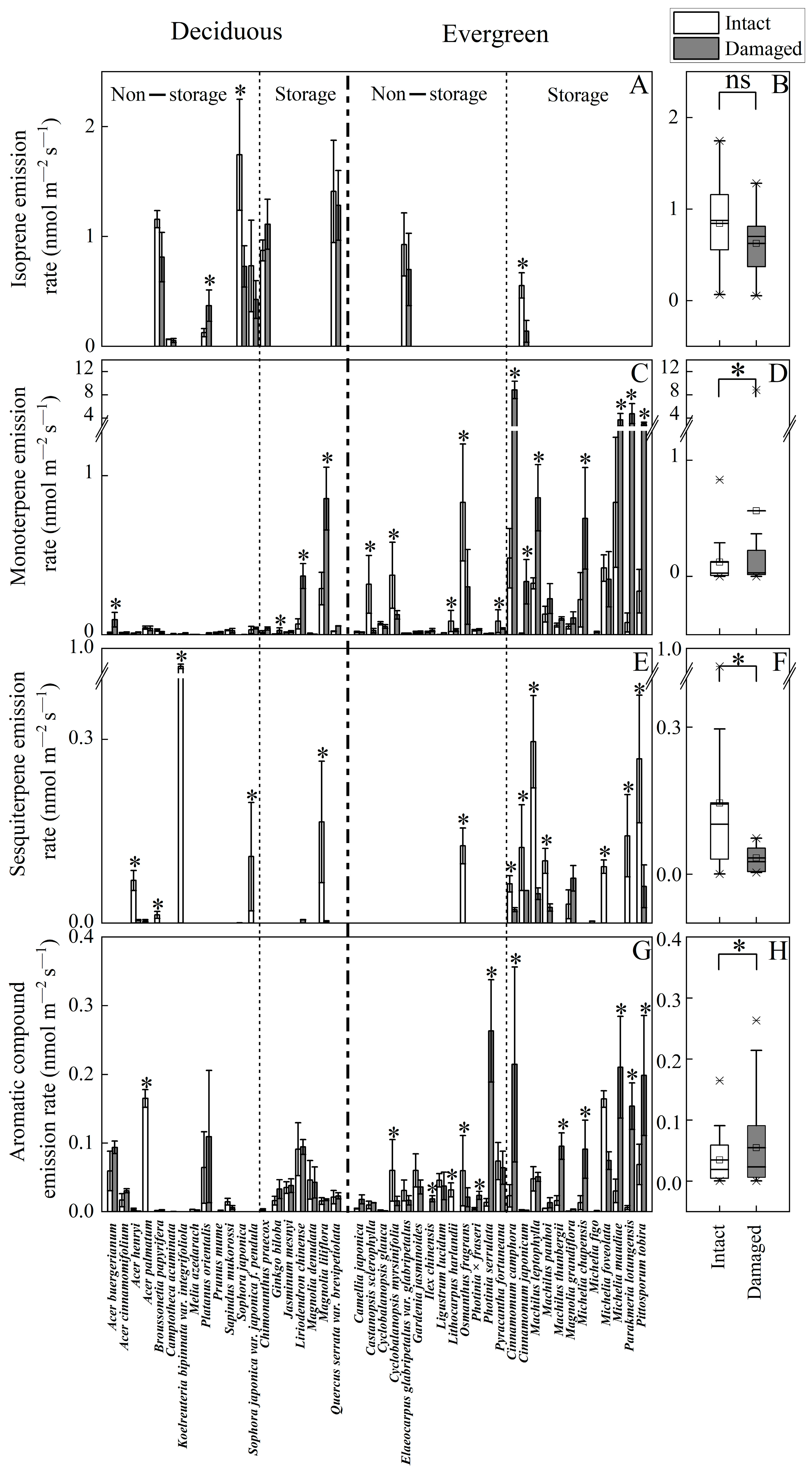
2.1. Impacts of Mechanical Injury on BVOC Emission Rates as Affected by the Presence of Storage Structures
2.1.1. Isoprene Emissions
2.1.2. Monoterpene Emissions
2.1.3. Sesquiterpene Emissions
2.1.4. Aromatic Compound Emissions
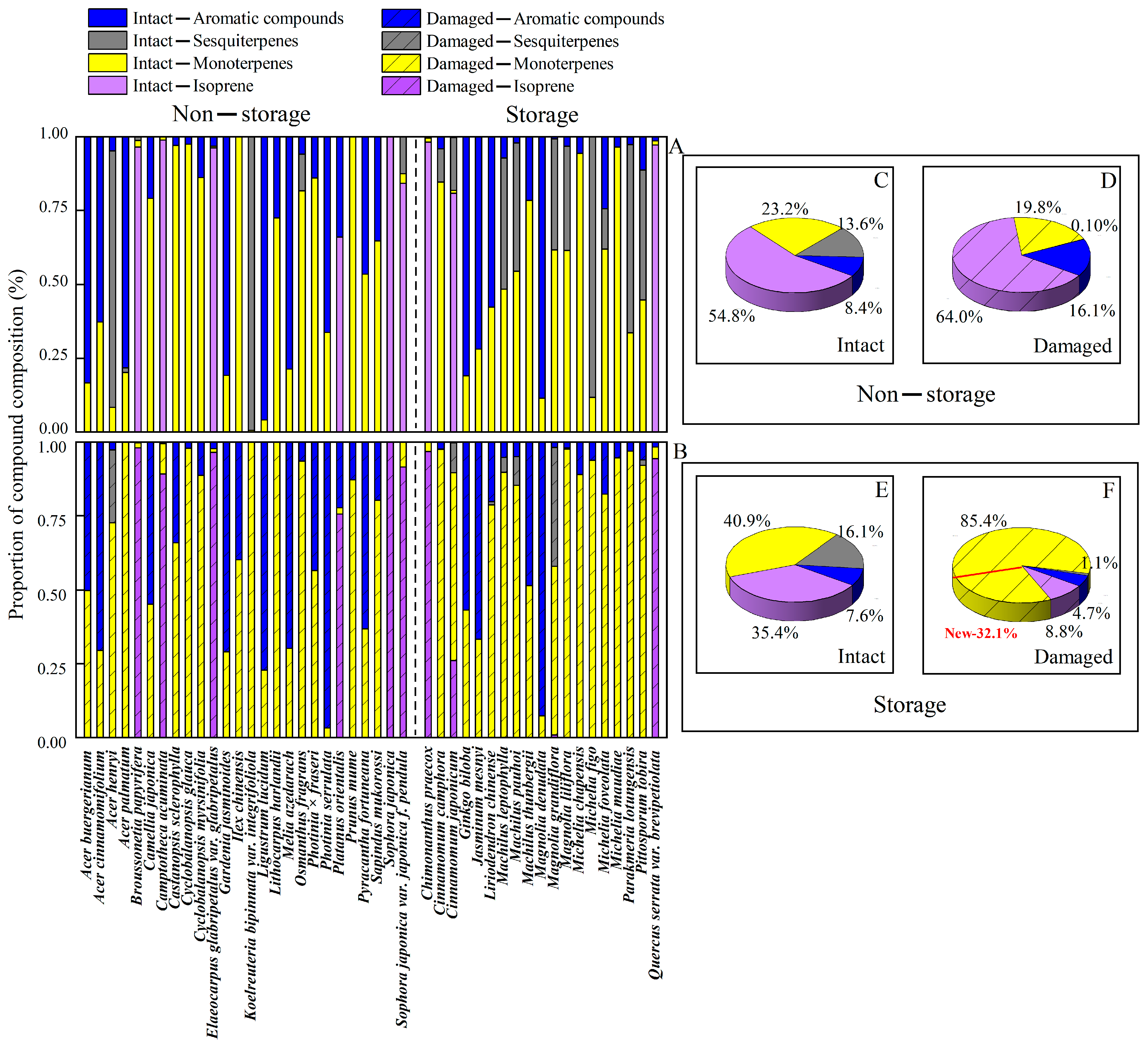
2.2. Impacts of Mechanical Injury on the BVOC Composition and Abundance in Storage and Non-Storage Species
2.2.1. Injury-Dependent Changes in the Composition of Emitted Monoterpenes
2.2.2. Changes in Sesquiterpene Composition in Response to Mechanical Injury
2.2.3. Modifications in the Composition of Aromatics After Mechanical Injury
2.3. Response of BVOC Emission to Mechanical Injury in Relation to Leaf Longevity and Species Ecological Adaptations
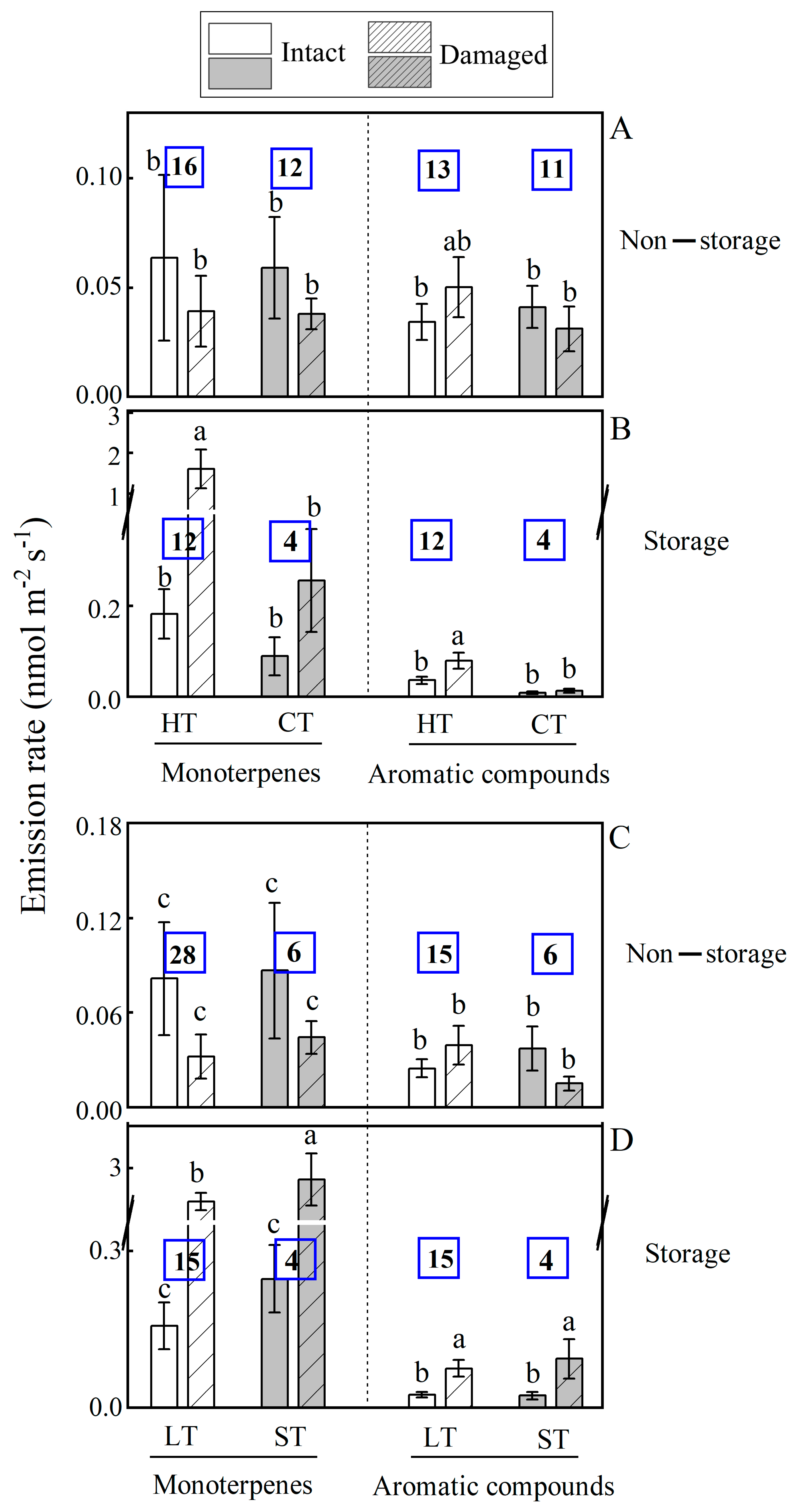
2.3.1. Changes in BVOC Emission Due to Mechanical Injury Associated with Leaf Longevity
2.3.2. Changes in BVOC Emission to Mechanical Injury in Species with Different Ecological Tolerance
3. Discussion
3.1. Changes in Isoprene Emission in Response to Mechanical Injury
3.2. Responses of Monoterpene Emissions to Mechanical Injury
3.3. Alterations in Sesquiterpene Emissions by Mechanical Injury
3.4. Aromatic Compound Responses to Mechanical Injury
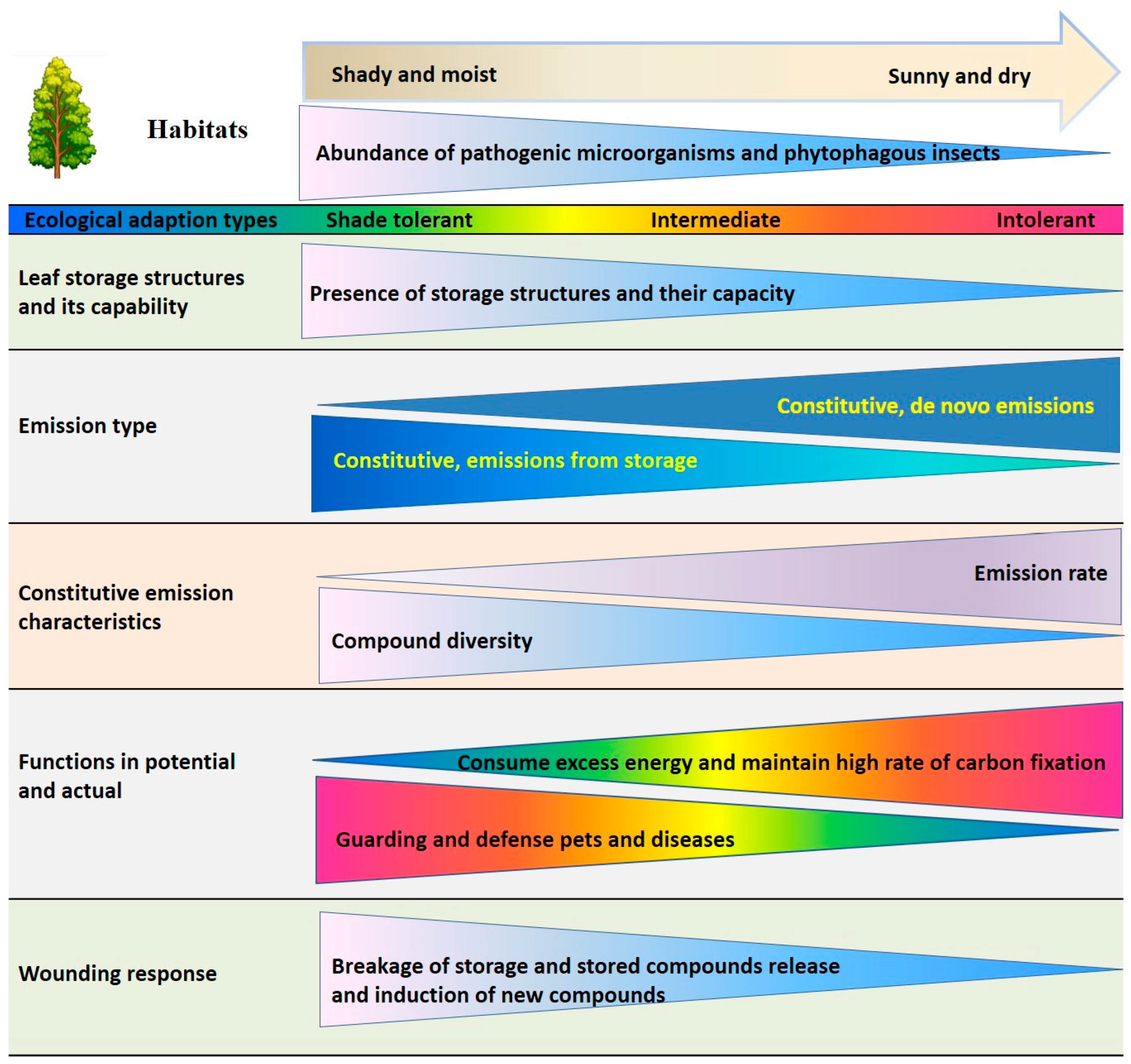
3.5. Effects of Plant Functional Types and Different Ecological Requirements on Species Emission After Mechanical Injury
4. Materials and Methods
4.1. Plant Selection and Sampling
4.2. Experimental Treatments
4.3. Compound Sampling and Analysis
4.4. Determination of Retention Index and Compound Identification
4.5. Classification and Identification of Leaf BVOC Storage Structures
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isoprene | Monoterpenes | Sesquiterpenes | Aromatic Compounds | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rate (nmol m−2 s−1) | Compound | Rate (nmol m−2 s−1) | Compound | Rate (nmol m−2 s−1) | Compound | Rate (nmol m−2 s−1) | |||||
| Intact | Injured | Intact | Injured | Intact | Injured | Intact | Injured | ||||
| Acer buergerianum | C10H16O Camphor | 0.012 ± 0.0021 | 0.014 ± 0.002 | C10H12 2,5-dimethylstyrene | 0.059 ± 0.014 | 0.092 ± 0.009 | |||||
| C10H16 α-pinene | 0.069 ± 0.0012 | C10H12O benzenebutanal | 0.0015 ± 0.00021 | ||||||||
| C10H16 Limonene | 0.024 ± 0.0010 | ||||||||||
| C10H16 α-phellandrene | 0.012 ± 0.0042 | ||||||||||
| C10H16 Pseudolimonene | 0.011 ± 0.0011 | ||||||||||
| C10H16 α-terpinolene | 0.0011 ± 0.00043 | ||||||||||
| Acer cinnamomifolium | C10H16 γ-terpinene | 0.0017 ± 0.00023 | 0.0041 ± 0.00062 | C10H12 2,5-dimethylstyrene | 0.021 ± 0.0056 | 0.031 ± 0.0020 | |||||
| C10H16 α-phellandrene | 0.00072 ± 0.00004 | * 0.0029 ± 0.0004 | |||||||||
| C10H16O Camphor | 0.0037 ± 0.0007 | 0.0059 ± 0.00081 | |||||||||
| C10H16 (E)-β-ocimene | 0.0020 ± 0.00033 | ||||||||||
| Acer henryi | C10H16 (E)-β-ocimene | 0.0031 ± 0.000010 | 0.0032 ± 0.0013 | C15H24 bicyclogermacrene | 0.028 ± 0.003 | C10H12 2,5-dimethylstyrene | * 0.0038 ± 0.0009 | 0.00061 ± 0.00025 | |||
| C10H16 α-pinene | 0.0018 ± 0.00020 | 0.0030 ± 0.00033 | C15H24 δ-guaiene | 0.024 ± 0.00034 | |||||||
| C10H16 Camphor | 0.011 ± 0.0018 | C15H24 aciphyllene | 0.015 ± 0.0013 | ||||||||
| C10H16 Limonene | 0.0022 ± 0.00049 | C15H24 α-muurolene | 0.018 ± 0.0053 | ||||||||
| C15H24 δ-selinene | 0.011 ± 0.00022 | ||||||||||
| Acer palmatum | C10H16 α-pinene | 0.012 ± 0.00083 | 0.015 ± 0.0028 | C15H24 cis-muurola-4(14),5-diene | 0.0032 ± 0.0012 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.17 ± 0.018 | ||||
| C10H16O Camphor | 0.030 ± 0.0055 | 0.029 ± 0.0035 | |||||||||
| Broussonetia papyrifera | 1.2 ± 0.051 | 0.81 ± 0.33 | C10H16 α-pinene | 0.0080 ± 0.0011 | 0.0054 ± 0.00096 | C15H24 α-muurolene | 0.011 ± 0.00001 | ||||
| C10H16O Camphor | 0.019 ± 0.0051 | 0.0085 ± 0.0016 | C15H24 bicyclogermacrene | 0.0078 ± 0.00072 | |||||||
| Camellia japonica | C10H16 Limonene | 0.0034 ± 0.00018 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.0032 ± 0.00063 | * 0.017 ± 0.00053 | ||||||
| C10H16 α-pinene | 0.0032 ± 0.00032 | 0.0025 ± 0.00033 | C7H6O benzaldehyde | * 0.0023 ± 0.0004 | 0.00071 ± 0.00038 | ||||||
| C10H16O Camphor | 0.011 ± 0.0013 | 0.011 ± 0.0010 | |||||||||
| Camptotheca acuminata | 0.064 ± 0.0062 | 0.051 ± 0.016 | C10H16 α-pinene | 0.00044 ± 0.00001 | 0.0031 ± 0.00023 | C13H18O2 2-methyl-3-phenylpropyl benzoate | 0.00043 ± 0.0000 | ||||
| C10H16O Camphor | 0.0014 ± 0.00013 | ||||||||||
| C10H16 Limonene | 0.0016 ± 0.00001 | ||||||||||
| Castanopsis sclerophylla | C10H16 α-pinene | * 0.097 ± 0.034 | 0.020 ± 0.0024 | C10H12 2,5-dimethylstyrene | 0.012 ± 0.0060 | ||||||
| C10H16 Limonene | * 0.022 ± 0.010 | 0.0034 ± 0.00076 | C13H18O2 2,4-butylethylbenzoate | 0.013 ± 0.00023 | |||||||
| C10H16 Cyclofeuchene | * 0.047 ± 0.015 | 0.0086 ± 0.00016 | |||||||||
| C10H16 γ-terpinene | * 0.046 ± 0.023 | 0.0079 ± 0.0020 | |||||||||
| C10H16 α-terpinene | 0.076 ± 0.017 | ||||||||||
| C10H16 camphene | 0.041 ± 0.023 | ||||||||||
| C10H16 Pseudolimonene | 0.021 ± 0.010 | ||||||||||
| C10H16 α-terpinolene | 0.013 ± 0.0049 | ||||||||||
| C10H16 (E)-β-ocimene | 0.011 ± 0.0056 | ||||||||||
| C10H16 α-phellandrene | 0.0086 ± 0.0027 | ||||||||||
| C10H16 α-fenchene | 0.0060 ± 0.0028 | ||||||||||
| C10H14 p-cymene | 0.069 ± 0.035 | ||||||||||
| Chimonanthus praecox | * 0.87 ± 0.077 | 0.14 ± 0.44 | C10H16 α-phellandrene | 0.0064 ± 0.00023 | C14H18O2 2,4-butylethylbenzoate | 0.0027 ± 0.00033 | |||||
| C10H16 α-pinene | 0.0023 ± 0.00063 | 0.0042 ± 0.00043 | C8H10 p-xylene | 0.0015 ± 0.0004 | |||||||
| C10H16 γ-terpinene | 0.0078 ± 0.00003 | * 0.024 ± 0.0033 | |||||||||
| C10H16 Pseudolimonene | 0.0037 ± 0.00058 | 0.0075 ± 0.00068 | |||||||||
| C10H16 β-phellandrene | 0.0025 ± 0.00022 | 0.0037 ± 0.00063 | |||||||||
| C10H16 Limonene | 0.0012 ± 0.000042 | * 0.0029 ± 0.0003 | |||||||||
| C10H16 α-terpinolene | 0.00061 ± 0.00001 | * 0.0015 ± 0.0002 | |||||||||
| C10H16O Camphor | 0.0022 ± 0.00050 | * 0.0058 ± 0.0011 | |||||||||
| C10H14 p-cymene | 0.0021 ± 0.00059 | * 0.010 ± 0.0034 | |||||||||
| C10H16 (E)-β-ocimene | 0.0071 ± 0.0013 | ||||||||||
| Cinnamomum camphora | C10H16 (E)-β-ocimene | 0.20 ± 0.085 | 0.37 ± 0.0093 | C15H24 γ-humulene | 0.039 ± 0.0057 | C10H12 2,5-dimethylstyrene | 0.013 ± 0.0063 | * 0.21 ± 0.083 | |||
| C10H16 α-pinene | 0.041 ± 0.015 | * 0.57 ± 0.040 | C15H24 bicyclogermacrene | 0.015 ± 0.0041 | C14H18O2 2,4-butylethylbenzoate | * 0.030 ± 0.011 | 0.012 ± 0.00023 | ||||
| C10H16 γ-terpinene | 0.024 ± 0.0097 | * 0.27 ± 0.0056 | C15H24 β-longipinene | 0.0089 ± 0.0032 | C10H12O2 dicyclopentadiene diepoxide | 0.012 ± 0.00010 | |||||
| C10H16 α-terpinene | 0.018 ± 0.0070 | * 0.30 ± 0.14 | C15H24 cis-muurola-4(14),5-diene | 0.0050 ± 0.0000 | |||||||
| C10H16 Limonene | 0.011 ± 0.0037 | * 0.56 ± 0.040 | C15H24 δ-selinene | 0.0053 ± 0.00001 | * 0.022 ± 0.0016 | ||||||
| C10H16 α-phellandrene | 0.0048 ± 0.00073 | 0.17 ± 0.066 | |||||||||
| C10H16 Pseudolimonene | 0.011 ± 0.0028 | * 0.18 ± 0.055 | |||||||||
| C10H16 α-terpinolene | 0.0069 ± 0.0027 | * 0.21 ± 0.085 | |||||||||
| C10H16 α-fenchene | 0.0010 ± 0.0001 | * 0.026 ± 0.0046 | |||||||||
| C10H16O Camphor | 0.30 ± 0.000012 | * 2.6 ± 0.79 | |||||||||
| C10H18O eucalyptol | 0.16 ± 0.095 | * 1.1 ± 0.39 | |||||||||
| C10H14 p-cymene | 0.058 ± 0.029 | * 0.49 ± 0.13 | |||||||||
| C10H16 camphene | 0.52 ± 0.068 | ||||||||||
| C10H16 3-carene | 0.27 ± 0.046 | ||||||||||
| C10H16 β-phellandrene | 0.25 ± 0.090 | ||||||||||
| C10H18O α-terpineol | 0.45 ± 0.13 | ||||||||||
| C10H18O trans-4-thujanol | 0.23 ± 0.074 | ||||||||||
| C10H18O endo-borneol | 0.099 ± 0.0081 | ||||||||||
| Cinnamomum japonicum | * 0.55 ± 0.071 | 0.16 ± 0.099 | C10H16 α-pinene | 0.0055 ± 0.0014 | * 0.075 ± 0.028 | C15H24 cis-muurola-4(14),5-diene | 0.0039 ± 0.0010 | C10H12 2,5-dimethylstyrene | 0.0020 ± 0.0011 | 0.0017 ± 0.00066 | |
| C10H16 camphene | 0.00090 ± 0.00009 | * 0.030 ± 0.012 | C15H24 γ-gurjunene | 0.026 ± 0.012 | 0.018 ± 0.00019 | ||||||
| C10H14 p-cymene | 0.00053 ± 0.00005 | * 0.0065 ± 0.0030 | C15H24 β-copaene | 0.11 ± 0.0069 | |||||||
| C10H16 Limonene | 0.035 ± 0.010 | C15H24 caryophyllene | 0.059 ± 0.000013 | ||||||||
| C10H16 Pseudolimonene | 0.027 ± 0.0097 | C15H24 bicyclogermacrene | 0.055 ± 0.0013 | ||||||||
| C10H16 β-phellandrene | 0.011 ± 0.0055 | C15H24 δ-guaiene | 0.033 ± 0.014 | ||||||||
| C10H16 α-phellandrene | 0.011 ± 0.0039 | C15H24 γ-humulene | 0.017 ± 0.00012 | ||||||||
| C10H16 (E)-β-ocimene | 0.0056 ± 0.0010 | C15H24 β-maaliene | 0.014 ± 0.0044 | ||||||||
| C10H16 γ-terpinene | 0.0027 ± 0.00025 | C15H24 δ-selinene | 0.0052 ± 0.0017 | 0.0072 ± 0.00052 | |||||||
| C10H16O Camphor | 0.24 ± 0.073 | ||||||||||
| C10H18O endo-Borneol | 0.010 ± 0.00064 | ||||||||||
| Cyclobalanopsis glauca | C10H16 α-pinene | 0.047 ± 0.0053 | 0.035 ± 0.016 | C10H12 3,4-dimethylstyrene | 0.00093 ± 0.0002 | 0.00070 ± 0.00002 | |||||
| C10H16 Pseudolimonene | 0.0050 ± 0.0013 | 0.0088 ± 0.00007 | C10H12O benzenebutanal | 0.0012 ± 0.00009 | |||||||
| C10H16 camphene | 0.0026 ± 0.00022 | 0.0039 ± 0.00019 | C13H18O2 2,4-butylethylbenzoate | 0.0018 ± 0.00021 | |||||||
| C10H16 Limonene | 0.0025 ± 0.00023 | 0.0032 ± 0.00086 | C8H10 p-xylene | 0.00062 ± 0.000061 | |||||||
| C10H16 α-terpinolene | 0.00051 ± 0.00003 | * 0.0013 ± 0.0001 | C15H14O2 2-phenylethyl benzoate | 0.00086 ± 0.00002 | |||||||
| C10H16O Camphor | 0.013 ± 0.0015 | 0.016 ± 0.0024 | |||||||||
| C10H14 p-cymene | 0.00073 ± 0.00022 | 0.00077 ± 0.0002 | |||||||||
| Cyclobalanopsis myrsinifolia | C10H16 α-pinene | 0.11 ± 0.0098 | 0.052 ± 0.023 | C10H12 3,4-dimethylstyrene | 0.010 ± 0.00021 | ||||||
| C10H16 γ-terpinene | * 0.050 ± 0.026 | 0.015 ± 0.0069 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.065 ± 0.025 | 0.016 ± 0.0040 | ||||||
| C10H16 Cyclofeuchene | 0.055 ± 0.016 | ||||||||||
| C10H16 α-terpinene | 0.032 ± 0.16 | 0.016 ± 0.0061 | |||||||||
| C10H16 Pseudolimonene | 0.027 ± 0.0066 | 0.016 ± 0.0039 | |||||||||
| C10H16 Limonene | 0.015 ± 0.0040 | 0.011 ± 0.0038 | |||||||||
| C10H16 camphene | * 0.014 ± 0.0040 | 0.0040 ± 0.0013 | |||||||||
| C10H16 α-terpinolene | * 0.011 ± 0.0047 | 0.0043 ± 0.0010 | |||||||||
| C10H16 β-phellandrene | * 0.010 ± 0.0042 | 0.0036 ± 0.00091 | |||||||||
| C10H16 α-phellandrene | 0.0065 ± 0.0028 | 0.0036 ± 0.00090 | |||||||||
| C10H16 α-fenchene | 0.0023 ± 0.00046 | ||||||||||
| C10H16O Camphor | 0.0032 ± 0.00063 | 0.0024 ± 0.00017 | |||||||||
| C10H14 p-cymene | * 0.069 ± 0.032 | 0.025 ± 0.012 | |||||||||
| Elaeocarpus glabripetalus var. glabripetalus | 0.93 ± 0.18 | 0.70 ± 0.37 | C10H16 α-pinene | 0.0016 ± 0.00031 | 0.0025 ± 0.00022 | C13H18O2 2,4-butylethylbenzoate | * 0.031 ± 0.0089 | 0.013 ± 0.0040 | |||
| C10H16O Camphor | 0.0045 ± 0.0010 | 0.0067 ± 0.0010 | C15H14O2 2-phenylethyl benzoate | 0.00048 ± 0.000073 | |||||||
| C16H16O2 2-phenylethyl benzoate | 0.00086 ± 0.00009 | ||||||||||
| Gardenia jasminoides | C10H16O camphor | 0.013 ± 0.0035 | 0.014 ± 0.0037 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.060 ± 0.014 | 0.036 ± 0.0069 | |||||
| C10H16 α-pinene | 0.0014 ± 0.00013 | ||||||||||
| Ginkgo biloba | C10H16 α-pinene | 0.0038 ± 0.0005 | * 0.014 ± 0.0043 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.015 ± 0.0034 | * 0.040 ± 0.0084 | |||||
| C10H16O Camphor | 0.035 ± 0.00078 | ||||||||||
| Ilex chinensis | C10H16 α-pinene | 0.0015 ± 0.00045 | * 0.0081 ± 0.0016 | C11H14O2 2-phenylethyl benzoate | 0.0031 ± 0.00076 | ||||||
| C10H16O Camphor | 0.012 ± 0.0020 | 0.025 ± 0.011 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.016 ± 0.0034 | |||||||
| Jasminum mesnyi | C10H16 α-pinene | 0.0035 ± 0.00051 | 0.0055 ± 0.00030 | C15H14O2 2-phenylethyl benzoate | 0.0014 ± 0.00001 | ||||||
| C10H16O Camphor | 0.010 ± 0.0015 | 0.013 ± 0.0020 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.034 ± 0.0059 | 0.038 ± 0.0057 | ||||||
| Koelreuteria bipinnata var. integrifoliola | C10H16 α-pinene | 0.0047 ± 0.00013 | 0.0046 ± 0.00052 | C15H24 β-longipinene | 0.36 ± 0.0019 | ||||||
| C15H24 valerena-4,7(11)-diene | 0.17 ± 0.010 | ||||||||||
| C15H24 γ-himachalene | 0.099 ± 0.00044 | ||||||||||
| C15H24 δ-guaiene | 0.059 ± 0.013 | ||||||||||
| C15H24 β-elemene | 0.056 ± 0.017 | ||||||||||
| C15H24 α-guaiene | 0.021 ± 0.00086 | ||||||||||
| C15H24 bicyclogermacrene | 0.017 ± 0.00063 | ||||||||||
| C15H24 β-copaene | 0.016 ± 0.00058 | ||||||||||
| C15H24 aromadendrene | 0.013 ± 0.000064 | ||||||||||
| Ligustrum lucidum | C10H16 α-pinene | 0.0020 ± 0.00045 | * 0.011 ± 0.00022 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.046 ± 0.0069 | 0.051 ± 0.0077 | |||||
| C9H10O2 2-phenylethyl benzoate | 0.0036 ± 0.00025 | ||||||||||
| Liriodendron chinense | C10H16 Pseudolimonene | 0.027 ± 0.0073 | * 0.12 ± 0.025 | C15H24 cis-muurola-3,5-diene | 0.0054 ± 0.00027 | C13H18O2 2,4-butylethylbenzoate | 0.091 ± 0.022 | 0.093 ± 0.015 | |||
| C10H16 α-pinene | 0.024 ± 0.0060 | 0.049 ± 0.018 | C10H12 3,4-dimethylstyrene | 0.0041 ± 0.00063 | |||||||
| C10H16 Limonene | 0.013 ± 0.0044 | 0.016 ± 0.0040 | |||||||||
| C10H16 γ-terpinene | 0.0052 ± 0.0020 | * 0.055 ± 0.0057 | |||||||||
| C10H16 α-fenchene | 0.0022 ± 0.00064 | 0.0031 ± 0.00005 | |||||||||
| C10H16 (E)-β-ocimene | 0.082 ± 0.020 | ||||||||||
| C10H16 α-terpinene | 0.034 ± 0.0049 | ||||||||||
| C10H16 camphene | 0.025 ± 0.00083 | ||||||||||
| C10H16 α-phellandrene | 0.022 ± 0.011 | ||||||||||
| C10H16 3-carene | 0.0084 ± 0.00063 | ||||||||||
| C10H18O eucalyptol | 0.0095 ± 0.00044 | ||||||||||
| C10H14 p-cymene | 0.0099 ± 0.0026 | ||||||||||
| Lithocarpus harlandii | C10H16 (E)-β-ocimene | * 0.050 ± 0.021 | 0.016 ± 0.0039 | C15H24 cis-muurola-3,5-diene | 0.0030 ± 0.0000011 | C10H12 3,4-dimethylstyrene | 0.0010 ± 0.00001 | ||||
| C10H16 α-pinene | 0.012 ± 0.0051 | 0.0050 ± 0.00083 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.041 ± 0.010 | |||||||
| C10H16O Camphor | 0.0080 ± 0.00031 | 0.0063 ± 0.00027 | |||||||||
| C10H16 α-terpinolene | 0.015 ± 0.0010 | ||||||||||
| C10H16 Pseudolimonene | 0.012 ± 0.00015 | ||||||||||
| C10H16 γ-terpinene | 0.010 ± 0.0039 | ||||||||||
| C10H16 Limonene | 0.0032 ± 0.000017 | ||||||||||
| C10H16 α-phellandrene | 0.0028 ± 0.000070 | ||||||||||
| C10H14 p-cymene | 0.0077 ± 0.0020 | ||||||||||
| Machilus leptophylla | C10H16 α-pinene | 0.11 ± 0.0028 | 0.21 ± 0.028 | C15H24 β-copaene | 0.094 ± 0.00087 | C10H12 3,4-dimethylstyrene | 0.019 ± 0.0059 | 0.022 ± 0.0014 | |||
| C10H16 camphene | 0.054 ± 0.019 | 0.072 ± 0.013 | C15H24 cis-muurola-4(14),5-diene | 0.040 ± 0.014 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.058 ± 0.010 | 0.029 ± 0.0029 | ||||
| C10H16 Pseudolimonene | 0.036 ± 0.017 | * 0.071 ± 0.013 | C15H24 σ-selinene | 0.054 ± 0.0051 | |||||||
| C10H16 α-terpinene | 0.029 ± 0.0095 | 0.058 ± 0.017 | C15H24 β-maaliene | 0.040 ± 0.0043 | |||||||
| C10H16 Limonene | 0.029 ± 0.0088 | 0.059 ± 0.0095 | C15H24 bicyclogermacrene | 0.018 ± 0.0062 | |||||||
| C10H16 γ-terpinene | 0.028 ± 0.011 | 0.034 ± 0.0066 | C15H24 β-elemene | 0.049 ± 0.014 | |||||||
| C10H16 β-phellandrene | 0.0086 ± 0.0010 | * 0.040 ± 0.011 | C15H24 aromadendrene | 0.018 ± 0.0062 | |||||||
| C10H16 α-terpinolene | 0.0068 ± 0.0017 | 0.011 ± 0.0018 | C15H24 α-bulnesene | 0.012 ± 0.00073 | 0.0027 ± 0.00034 | ||||||
| C10H16 α-phellandrene | 0.0083 ± 0.0036 | * 0.16 ± 0.0049 | C15H24 α-ylangene | 0.0065 ± 0.0019 | |||||||
| C10H16 α-fenchene | 0.0058 ± 0.0022 | 0.0076 ± 0.0011 | C15H24 germacrene D | 0.0030 ± 0.0010 | |||||||
| C10H14 p-cymene | 0.050 ± 0.014 | 0.10 ± 0.016 | C15H24 β-longipinene | 0.0066 ± 0.0019 | |||||||
| C10H16 (E)-β-ocimene | 0.011 ± 0.0011 | C15H24 aromadendrene | 0.017 ± 0.0021 | ||||||||
| C10H16 β-fenchene | 0.0035 ± 0.00032 | C15H24 cis-muurola-3,5-diene | 0.041 ± 0.000011 | ||||||||
| C10H18O eucalyptol | 0.015 ± 0.0012 | C15H24 β-longipinene | 0.026 ± 0.0044 | ||||||||
| C10H16 Cyclofeuchene | 0.029 ± 0.00033 | 0.019 ± 0.0034 | C15H24 (-)-aristolene | 0.0038 ± 0.00081 | |||||||
| Machilus pauhoi | C10H16 α-pinene | 0.039 ± 0.022 | * 0.096 ± 0.021 | C15H24 σ-selinene | 0.055 ± 0.015 | C13H18O3 methanol, [4-(1,1-dimethylethyl)phenoxy]-, acetate | 0.0015 ± 0.00004 | ||||
| C10H16 γ-terpinene | 0.052 ± 0.0090 | 0.064 ± 0.012 | C15H24 (-)-aciphyllene | 0.018 ± 0.0065 | C10H12 o-cumenene | 0.0045 ± 0.0011 | 0.0068 ± 0.0015 | ||||
| C10H16 Pseudolimonene | 0.017 ± 0.0023 | 0.020 ± 0.0063 | C15H24 β-copaene | 0.018 ± 0.0078 | C16H16O2 2-phenylethyl benzoate | 0.019 ± 0.0071 | |||||
| C10H16 α-terpinolene | 0.0058 ± 0.00093 | 0.0066 ± 0.0016 | C15H24 α-muurolene | 0.0086 ± 0.0026 | |||||||
| C10H16 β-phellandrene | 0.0056 ± 0.000057 | 0.0063 ± 0.0017 | C15H24 aromadendrene | 0.0025 ± 0.0011 | |||||||
| C10H16 Limonene | 0.0056 ± 0.0024 | 0.012 ± 0.0028 | C15H24 β-patchoulene | 0.0055 ± 0.0011 | |||||||
| C10H16 camphene | 0.011 ± 0.0038 | 0.011 ± 0.00051 | C15H24 caryophyllene | 0.0030 ± 0.00060 | |||||||
| C10H16 α-phellandrene | 0.0070 ± 0.000052 | 0.0085 ± 0.0018 | |||||||||
| C10H16 (E)-β-ocimene | 0.0078 ± 0.0016 | ||||||||||
| C10H14 p-cymene | 0.038 ± 0.015 | 0.021 ± 0.0078 | |||||||||
| Machilus thunbergii | C10H16 α-terpinolene | 0.025 ± 0.0017 | 0.053 ± 0.0037 | C15H14O2 2-phenylethyl benzoate | 0.0012 ± 0.00026 | ||||||
| C10H16 γ-terpinene | 0.0039 ± 0.00023 | * 0.010 ± 0.0024 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.017 ± 0.0047 | * 0.094 ± 0.045 | ||||||
| C10H16 α-terpinene | 0.0029 ± 0.000010 | * 0.013 ± 0.00005 | C9H10O2 2-phenylethyl formate | 0.0037 ± 0.00033 | |||||||
| C10H16 Limonene | 0.00063 ± 0.00024 | * 0.012 ± 0.0038 | |||||||||
| C10H16 Pseudolimonene | 0.058 ± 0.0011 | ||||||||||
| C10H16 camphene | 0.0024 ± 0.00013 | ||||||||||
| C10H14 p-cymene | 0.0041 ± 0.0010 | ||||||||||
| C10H16 α-thujene | 0.011 ± 0.00083 | 0.0093 ± 0.00044 | |||||||||
| C10H16O Camphor | 0.023 ± 0.0046 | 0.021 ± 0.0054 | |||||||||
| Magnolia denudata | C10H16 α-pinene | 0.0013 ± 0.00025 | 0.0013 ± 0.00007 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.046 ± 0.018 | 0.038 ± 0.013 | |||||
| C10H16O Camphor | 0.0017 ± 0.00020 | 0.0020 ± 0.00023 | C15H14O2 2-phenylethyl benzoate | 0.0010 ± 0.000071 | |||||||
| C10H18O eucalyptol | 0.0092 ± 0.00034 | ||||||||||
| Magnolia grandiflora | C10H16 camphene | 0.0028 ± 0.000051 | 0.0071 ± 0.0014 | C15H24 β-elemene | 0.0078 ± 0.0044 | * 0.081 ± 0.0076 | C10H12 3,4-dimethylstyrene | 0.00066 ± 0.0002 | * 0.0023 ± 0.00090 | ||
| C10H16 Pseudolimonene | 0.0026 ± 0.00053 | * 0.011 ± 0.0041 | C15H24 β-copaene | 0.0040 ± 0.0007 | C12H16O 1-(4-methylphenyl)-1-Pentanone | 0.0033 ± 0.0012 | |||||
| C10H16 Limonene | 0.0014 ± 0.0003 | * 0.0064 ± 0.0016 | C15H24 cis-muurola-4(14),5-diene | 0.028 ± 0.0079 | |||||||
| C10H16 γ-terpinene | 0.0013 ± 0.0002 | * 0.0048 ± 0.0009 | C15H24 caryophyllene | 0.023 ± 0.00001 | |||||||
| C10H16 α-terpinene | 0.00093 ± 0.00006 | * 0.0032 ± 0.0005 | C15H24 bicyclogermacrene | * 0.015 ± 0.0041 | 0.0014 ± 0.0005 | ||||||
| C10H14 p-cymene | 0.0021 ± 0.0005 | 0.0031 ± 0.0012 | C15H24 γ-gurjunene | 0.015 ± 0.0062 | 0.0087 ± 0.0028 | ||||||
| C10H16 α-fenchene | 0.0014 ± 0.0002 | ||||||||||
| C10H16O Camphor | 0.058 ± 0.0311 | ||||||||||
| C10H16 α-pinene | 0.042 ± 0.0069 | 0.040 ± 0.016 | |||||||||
| C10H16 α-thujene | 0.0025 ± 0.0002 | ||||||||||
| Magnolia liliflora | C10H16 α-pinene | 0.076 ± 0.032 | 0.13 ± 0.018 | C15H24 γ-gurjunene | 0.14 ± 0.038 | C14H18O2 2-phenylethyl benzoate | 0.013 ± 0.0012 | 0.014 ± 0.0031 | |||
| C10H16 Pseudolimonene | 0.063 ± 0.035 | * 0.18 ± 0.019 | C15H24 γ-humulene | 0.040 ± 0.0075 | C10H12 3,4-dimethylstyrene | 0.0036 ± 0.0014 | 0.0037 ± 0.0006 | ||||
| C10H16 γ-terpinene | 0.035 ± 0.016 | * 0.14 ± 0.031 | C15H24 β-longipinene | 0.11 ± 0.0002 | C9H12O 3,4-dimethylbenzyl alcohol | 0.011 ± 0.0033 | |||||
| C10H16 Limonene | 0.027 ± 0.011 | * 0.068 ± 0.0098 | C15H24 δ-guaiene | 0.016 ± 0.0037 | |||||||
| C10H16 camphene | 0.025 ± 0.011 | 0.040 ± 0.0061 | C15H24 germacrene D | 0.012 ± 0.0031 | |||||||
| C10H16 (E)-β-ocimene | 0.0035 ± 0.0004 | * 0.011 ± 0.0034 | C15H24 β-bourbonene | 0.0032 ± 0.0004 | |||||||
| C10H16 α-terpinolene | 0.0029 ± 0.0003 | 0.0044 ± 0.0008 | |||||||||
| C10H16 α-thujene | 0.0013 ± 0.00006 | 0.0025 ± 0.0004 | |||||||||
| C10H16 β-phellandrene | 0.00063 ± 0.0001 | 0.0014 ± 0.0003 | |||||||||
| C10H18O eucalyptol | 0.012 ± 0.0019 | 0.020 ± 0.0019 | |||||||||
| C10H14 p-cymene | 0.047 ± 0.022 | * 0.13 ± 0.017 | |||||||||
| C10H16 α-terpinene | 0.022 ± 0.0052 | ||||||||||
| C10H16O Camphor | 0.13 ± 0.042 | ||||||||||
| C10H18O α-terpineol | 0.0090 ± 0.0003 | ||||||||||
| Melia azedarach | C10H16 α-pinene | 0.00052 ± 0.00009 | 0.00094 ± 0.0002 | C8H10 p-xylene | 0.0032 ± 0.0002 | ||||||
| C10H16O camphor | 0.00062 ± 0.00001 | C10H18O2 cis-3-Hexenyl iso-butyrate | 0.00093 ± 0.00005 | ||||||||
| C15H14O2 2-phenylethyl benzoate | 0.0014 ± 0.0030 | ||||||||||
| Michelia chapensis | C10H16 α-pinene | 0.090 ± 0.031 | 0.11 ± 0.027 | C10H12 3,4-dimethylstyrene | 0.013 ± 0.0050 | * 0.049 ± 0.021 | |||||
| C10H16 α-terpinene | 0.061 ± 0.029 | 0.11 ± 0.042 | C9H12O 3,4-dimethylbenzyl alcohol | 0.082 ± 0.032 | |||||||
| C10H16 camphene | 0.054 ± 0.017 | 0.055 ± 0.023 | C9H10O2 (2-methylphenyl)methyl formate | 0.0031 ± 0.0003 | |||||||
| C10H16 Limonene | 0.041 ± 0.019 | 0.061 ± 0.027 | |||||||||
| C10H16 α-thujene | 0.029 ± 0.018 | * 0.11 ± 0.031 | |||||||||
| C10H16 Pseudolimonene | 0.027 ± 0.013 | 0.052 ± 0.025 | |||||||||
| C10H16 γ-terpinene | 0.018 ± 0.0068 | * 0.13 ± 0.048 | |||||||||
| C10H16 β-phellandrene | 0.013 ± 0.0045 | * 0.038 ± 0.016 | |||||||||
| C10H16 α-phellandrene | 0.0087 ± 0.0043 | 0.015 ± 0.0093 | |||||||||
| C10H16 α-fenchene | 0.0066 ± 0.0021 | 0.0087 ± 0.0036 | |||||||||
| C10H16 α-terpinolene | 0.0036 ± 0.0013 | * 0.040 ± 0.015 | |||||||||
| C10H18O eucalyptol | 0.031 ± 0.0096 | 0.037 ± 0.014 | |||||||||
| C10H14 p-cymene | 0.10 ± 0.028 | * 0.20 ± 0.033 | |||||||||
| C10H16 3-carene | 0.015 ± 0.0026 | ||||||||||
| C10H18O trans-4-thujanol | 0.010 ± 0.0011 | ||||||||||
| Michelia figo | C10H16 α-pinene | 0.00043 ± 0.00008 | * 0.0039 ± 0.0003 | C15H24 germacrene D | 0.0052 ± 0.00001 | C11H14O2 2-phenylethyl benzoate | 0.0011 ± 0.00041 | ||||
| C10H16 camphene | 0.013 ± 0.0014 | C15H24 cis-muurola-4(14),5-diene | 0.0023 ± 0.0002 | ||||||||
| C10H16 Limonene | 0.00076 ± 0.0003 | ||||||||||
| Michelia foveolata | C10H16 α-thujene | 0.039 ± 0.000022 | 0.068 ± 0.012 | C15H24 β-copaene | 0.036 ± 0.000011 | C14H18O2 4-cyclopentyl ethylbenzoate | * 0.14 ± 0.0087 | 0.064 ± 0.023 | |||
| C10H16 γ-terpinene | 0.068 ± 0.0052 | 0.11 ± 0.024 | C15H24 γ-elemene | 0.034 ± 0.0056 | C10H12 2,5-dimethylstyrene | 0.010 ± 0.0015 | 0.0096 ± 0.0054 | ||||
| C10H16 α-terpinene | 0.028 ± 0.0049 | 0.046 ± 0.0190 | C15H24 bicyclogermacrene | 0.026 ± 0.0010 | C9H12O 3,4-dimethylbenzyl alcohol | 0.027 ± 0.0028 | |||||
| C10H16 Limonene | 0.025 ± 0.0028 | 0.029 ± 0.017 | C15H24 α-muurolene | 0.021 ± 0.0060 | C9H10O2 2-phenylethyl benzoate | 0.0021 ± 0.00022 | |||||
| C10H16 camphene | 0.020 ± 0.0002 | * 0.055 ± 0.014 | C15H24 α-copaene | 0.017 ± 0.0033 | |||||||
| C10H16 α-terpinolene | 0.0073 ± 0.0012 | 0.016 ± 0.0078 | C15H24 -guaiene | 0.014 ± 0.00041 | |||||||
| C10H16 α-phellandrene | 0.0062 ± 0.0013 | 0.019 ± 0.0035 | C15H24 (-)-aristolene | 0.0097 ± 0.0022 | |||||||
| C10H16 β-phellandrene | 0.0060 ± 0.0008 | * 0.020 ± 0.0043 | C15H24 β-longipinene | 0.0084 ± 0.00008 | |||||||
| C10H16 α-fenchene | 0.0013 ± 0.00002 | * 0.0080 ± 0.0019 | C15H24 γ-humulene | 0.0065 ± 0.00004 | |||||||
| C10H18O eucalyptol | 0.0070 ± 0.0005 | 0.020 ± 0.0030 | C15H24 β-maaliene | 0.0045 ± 0.00005 | |||||||
| C10H16O Camphor | 0.012 ± 0.0020 | C15H24 isoledene | 0.0029 ± 0.00052 | ||||||||
| C10H14 p-cymene | 0.055 ± 0.0097 | 0.040 ± 0.020 | |||||||||
| C10H16 α-pinene | 0.14 ± 0.019 | 0.082 ± 0.029 | |||||||||
| C10H16 (E)-β-ocimene | 0.084 ± 0.0052 | 0.049 ± 0.023 | |||||||||
| Michelia maudiae | C10H16 camphene | 0.18 ± 0.041 | * 1.0 ± 0.19 | C10H12 2,5-dimethylstyrene | 0.018 ± 0.054 | * 0.15 ± 0.027 | |||||
| C10H16 α-pinene | 0.12 ± 0.031 | * 0.45 ± 0.079 | C13H18O2 2,4-butylethylbenzoate | 0.031 ± 0.0042 | |||||||
| C10H16 (E)-β-ocimene | 0.11 ± 0.037 | 0.12 ± 0.027 | C9H12O 3,4-dimethylbenzyl alcohol | 0.077 ± 0.029 | |||||||
| C10H16 Cyclofeuchene | 0.10 ± 0.023 | * 0.32 ± 0.021 | |||||||||
| C10H16 Limonene | 0.086 ± 0.032 | * 0.43 ± 0.090 | |||||||||
| C10H16 Pseudolimonene | 0.082 ± 0.028 | * 0.44 ± 0.052 | |||||||||
| C10H16 γ-terpinene | 0.036 ± 0.015 | * 0.17 ± 0.045 | |||||||||
| C10H16 α-terpinene | 0.028 ± 0.0095 | * 0.15 ± 0.051 | |||||||||
| C10H16 α-thujene | 0.019 ± 0.0074 | * 0.10 ± 0.037 | |||||||||
| C10H16 α-terpinolene | 0.013 ± 0.0039 | * 0.16 ± 0.054 | |||||||||
| C10H16 α-phellandrene | 0.0051 ± 0.00023 | * 0.11 ± 0.0092 | |||||||||
| C10H16 β-phellandrene | 0.0043 ± 0.0013 | * 0.076 ± 0.020 | |||||||||
| C10H18O eucalyptol | 0.023 ± 0.0093 | * 0.23 ± 0.068 | |||||||||
| C10H16O Camphor | 0.0017 ± 0.00072 | * 0.21 ± 0.030 | |||||||||
| C10H14 p-cymene | 0.083 ± 0.031 | * 0.23 ± 0.057 | |||||||||
| C10H16 β-fenchene | 0.17 ± 0.11 | ||||||||||
| C10H16 3-carene | 0.12 ± 0.0048 | ||||||||||
| C10H18O Fenchol | 0.053 ± 0.0017 | ||||||||||
| C10H18O endo-borneol | 0.050 ± 0.0017 | ||||||||||
| C10H18O trans-4-thujanol | 0.024 ± 0.0.0041 | ||||||||||
| C10H18O α-terpineol | 0.054 ± 0.016 | ||||||||||
| Osmanthus fragrans | C10H16O Camphor | 0.024 ± 0.000090 | C15H24 β-longipinene | 0.13 ± 0.021 | C14H18O2 4-cyclopentyl ethylbenzoate | * 0.017 ± 0.0038 | 0.0064 ± 0.00054 | ||||
| C10H16 α-pinene | 0.0049 ± 0.00051 | C10H12 3,4-dimethylstyrene | * 0.14 ± 0.039 | 0.029 ± 0.0069 | |||||||
| C10H16 Pseudolimonene | 0.0099 ± 0.0014 | ||||||||||
| C10H16 camphene | * 0.55 ± 0.047 | 0.20 ± 0.059 | |||||||||
| C10H16 Cyclofeuchene | * 0.32 ± 0.051 | 0.13 ± 0.00001 | |||||||||
| C10H16 Limonene | * 0.28 ± 0.00013 | 0.044 ± 0.0077 | |||||||||
| C10H16 α-terpinolene | * 0.020 ± 0.00014 | 0.0041 ± 0.0010 | |||||||||
| C10H14 p-cymene | * 0.36 ± 0.11 | 0.14 ± 0.058 | |||||||||
| C10H16 bornylene | 0.031 ± 0.0090 | ||||||||||
| C10H16 α-fenchene | 0.020 ± 0.000013 | ||||||||||
| Parakmeria lotungensis | C10H16 α-pinene | 0.028 ± 0.012 | * 0.45 ± 0.16 | C15H24 β-longipinene | 0.17 ± 0.036 | C10H12 3,4-dimethylstyrene | 0.012 ± 0.000014 | * 0.14 ± 0.037 | |||
| C10H16 Limonene | 0.021 ± 0.0077 | * 0.47 ± 0.21 | C15H24 α-copaene | 0.0040 ± 0.00011 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.0029 ± 0.00020 | |||||
| C10H16 Pseudolimonene | 0.019 ± 0.0099 | * 0.58 ± 0.19 | C9H12O p-cumenol | 0.022 ± 0.0048 | |||||||
| C10H16 β-phellandrene | 0.0020 ± 0.00051 | * 0.14 ± 0.052 | |||||||||
| C10H18O eucalyptol | 0.0057 ± 0.00062 | * 0.28 ± 0.051 | |||||||||
| C10H16 camphene | 0.46 ± 0.11 | ||||||||||
| C10H16 α-terpinene | 0.54 ± 0.19 | ||||||||||
| C10H16 γ-terpinene | 0.32 ± 0.094 | ||||||||||
| C10H16 α-fenchene | 0.17 ± 0.047 | ||||||||||
| C10H16 (E)-β-ocimene | 0.10 ± 0.041 | ||||||||||
| C10H16 3-carene | 0.18 ± 0.059 | ||||||||||
| C10H16 Cyclofeuchene | 0.18 ± 0.061 | ||||||||||
| C10H16 α-phellandrene | 0.15 ± 0.059 | ||||||||||
| C10H16 β-fenchene | 0.068 ± 0.024 | ||||||||||
| C10H14 p-cymene | 0.41 ± 0.084 | ||||||||||
| C10H16O Camphor | 0.12 ± 0.047 | ||||||||||
| C10H18O endo-borneol | 0.10 ± 0.022 | ||||||||||
| C10H18O trans-4-thujanol | 0.082 ± 0.017 | ||||||||||
| C10H18O Fenchol | 0.064 ± 0.021 | ||||||||||
| C10H18O α-terpineol | 0.034 ± 0.0082 | ||||||||||
| C10H18O Linalool | 0.29 ± 0.16 | ||||||||||
| Photinia × fraseri | C10H16 α-pinene | 0.0056 ± 0.00080 | 0.0068 ± 0.0010 | C7H6O benzaldehyde | 0.0018 ± 0.00042 | * 0.016 ± 0.0047 | |||||
| C10H16O Camphor | 0.019 ± 0.0023 | 0.021 ± 0.0016 | C11H14O2 2-phenylethyl benzoate | 0.0042 ± 0.00071 | 0.0038 ± 0.00070 | ||||||
| C10H16 (E)-β-ocimene | 0.0068 ± 0.0010 | C8H10 p-xylene | 0.013 ± 0.0034 | ||||||||
| Photinia serrulata | C10H16 Limonene | 0.00073 ± 0.00005 | 0.00091 ± 0.0002 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.015 ± 0.0054 | 0.016 ± 0.0020 | |||||
| C10H16O Camphor | 0.0032 ± 0.0010 | 0.0042 ± 0.00065 | C15H14O2 2-phenylethyl benzoate | 0.00033 ± 0.000022 | * 0.00081 ± 0.000041 | ||||||
| C10H16 α-pinene | 0.0044 ± 0.0021 | 0.0032 ± 0.00071 | C7H6O benzaldehyde | 0.21 ± 0.042 | |||||||
| C7H8O benzyl alcohol | 0.050 ± 0.0081 | ||||||||||
| Pittosporum tobira | C10H16 camphene | 0.079 ± 0.012 | * 0.32 ± 0.082 | C15H24 thujopsene | 0.044 ± 0.0026 | C10H12 2,5-dimethylstyrene | 0.065 ± 0.0097 | * 0.17 ± 0.043 | |||
| C10H16 Cyclofeuchene | 0.025 ± 0.0040 | * 0.22 ± 0.012 | C15H24 β-longipinene | 0.14 ± 0.0020 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.018 ± 0.0023 | |||||
| C10H16 α-pinene | 0.039 ± 0.022 | * 0.93 ± 0.25 | C15H24 cis-thujopsene | 0.049 ± 0.0036 | C9H12O 3,4-dimethylbenzyl alcohol | 0.090 ± 0.0016 | |||||
| C10H16 bornylene | 0.0053 ± 0.00061 | * 0.10 ± 0.022 | C15H24 bicyclogermacrene | 0.0043 ± 0.0001 | |||||||
| C10H16 β-fenchene | 0.0038 ± 0.00064 | * 0.47 ± 0.19 | C15H24 γ-amorphene | 0.051 ± 0.0125 | |||||||
| C10H16 Limonene | 0.021 ± 0.0047 | * 0.18 ± 0.048 | C15H24 γ-cadinene | 0.036 ± 0.00030 | |||||||
| C10H16 α-fenchene | 0.12 ± 0.034 | C15H24 cis-muurola-4(14),5-diene | 0.012 ± 0.0028 | ||||||||
| C10H16 α-terpinene | 0.12 ± 0.041 | C15H24 γ-elemene | 0.013 ± 0.0028 | ||||||||
| C10H16 Pseudolimonene | 0.13 ± 0.029 | C15H24 β-copaene | 0.17 ± 0.016 | ||||||||
| C10H16 α-terpinolene | 0.12 ± 0.033 | C15H24 cis-muurola-3,5-diene | * 0.057 ± 0.0074 | 0.017 ± 0.0070 | |||||||
| C10H16 γ-terpinene | 0.075 ± 0.030 | C15H24 isoledene | 0.035 ± 0.016 | 0.028 ± 0.018 | |||||||
| C10H16 α-phellandrene | 0.050 ± 0.013 | ||||||||||
| C10H16 β-phellandrene | 0.030 ± 0.0058 | ||||||||||
| C10H16 3-carene | 0.12 ± 0.00050 | ||||||||||
| C10H16 (E)-β-ocimene | 0.0044 ± 0.0017 | ||||||||||
| C10H18O Fenchol | 0.049 ± 0.016 | ||||||||||
| C10H18O endo-borneol | 0.049 ± 0.0091 | ||||||||||
| C10H16O Camphor | 0.025 ± 0.00051 | ||||||||||
| C10H18O fenchone | 0.015 ± 0.0015 | ||||||||||
| C10H14 p-cymene | 0.21 ± 0.012 | ||||||||||
| Platanus orientalis | 0.13 ± 0.044 | * 0.37 ± 0.090 | C10H16 α-pinene | 0.0018 ± 0.00001 | C13H18O2 2,4-butylethylbenzoate | 0.064 ± 0.030 | 0.13 ± 0.057 | ||||
| C10H16O Camphor | 0.0085 ± 0.00062 | ||||||||||
| Prunus mume | C10H16 α-pinene | 0.0032 ± 0.00020 | 0.0048 ± 0.00063 | C15H14O2 2-phenylethyl benzoate | 0.0021 ± 0.000024 | ||||||
| C10H16O Camphor | 0.0086 ± 0.0010 | 0.011 ± 0.0018 | |||||||||
| Pyracantha fortuneana | C10H16 camphene | 0.011 ± 0.0044 | 0.012 ± 0.0018 | C13H18O2 2,4-butylethylbenzoate | 0.072 ± 0.015 | 0.064 ± 0.014 | |||||
| C10H16 (E)-β-ocimene | 0.13 ± 0.00033 | C16H16O2 2-phenylethyl benzoate | 0.0034 ± 0.0013 | ||||||||
| C10H16 α-pinene | 0.017 ± 0.0061 | 0.015 ± 0.0015 | |||||||||
| C10H16O Camphor | 0.012 ± 0.0050 | 0.010 ± 0.0013 | |||||||||
| Quercus serrata var. brevipetiolata | 1.4 ± 0.3024 | 1.3 ± 0.5391 | C10H16 α-pinene | 0.0074 ± 0.00060 | * 0.023 ± 0.0010 | C14H18O2 4-cyclopentyl ethylbenzoate | 0.019 ± 0.0064 | 0.023 ± 0.0026 | |||
| C10H16 Limonene | 0.0019 ± 0.00027 | * 0.0050 ± 0.0013 | |||||||||
| C10H16O Camphor | 0.013 ± 0.00073 | 0.023 ± 0.00066 | |||||||||
| C10H16 Sabinene | 0.0020 ± 0.00090 | ||||||||||
| C10H16 Pseudolimonene | 0.0018 ± 0.00006 | ||||||||||
| Sapindus mukorossi | C10H16 α-pinene | 0.0020 ± 0.00011 | 0.0034 ± 0.00070 | C13H18O2 2,4-butylethylbenzoate | 0.012 ± 0.0031 | 0.0057 ± 0.0015 | |||||
| C10H16 Limonene | 0.00053 ± 0.0001 | C10H18O2 4-hexen-1 butylethylbenzoate | 0.00063 ± 0.00013 | ||||||||
| C10H16O Camphor | 0.023 ± 0.0024 | 0.021 ± 0.0061 | |||||||||
| Sophora japonica | 1.7 ± 0.29 | 0.76 ± 0.29 | C10H14 p-cymene | 0.00084 ± 0.0002 | C15H24 cis-muurola-4(14),5-diene | 0.00063 ± 0.000023 | |||||
| Sophora japonica var. japonica f. pendula | 0.87 ± 0.22 | 0.49 ± 0.21 | C10H16 Cyclofeuchene | * 0.049 ± 0.00013 | 0.012 ± 0.0020 | C15H24 α-muurolene | 0.1498 ± 0.0024 | ||||
| C10H16 α-pinene | 0.0049 ± 0.0026 | 0.0054 ± 0.00011 | C15H24 cis-muurola-4(14),5-diene | 0.089 ± 0.040 | |||||||
| C10H16O Camphor | 0.016 ± 0.0056 | 0.028 ± 0.00053 | C15H24 β-copaene | 0.060 ± 0.000021 | |||||||
| C15H24 σ-cadinene | 0.0356 ± 0.0080 | ||||||||||
Appendix B
| Compounds | Molecular Formula | tx/min | Carbon Number (n) | tn/min | tn+1/min | Retention Index (RI) |
|---|---|---|---|---|---|---|
| benzyl alcohol | C7H8O | 20.872 | C8 | 25.521 | 29.855 | 692.7319 |
| p-xylene | C8H10 | 29.507 | C8 | 25.521 | 29.855 | 891.9705 |
| 2-phenylethyl benzoate | C11H14O2 | 30.714 | C9 | 29.855 | 34.8 | 917.3711 |
| 2-phenylethyl benzoate | C15H14O2 | 30.726 | C9 | 29.855 | 34.8 | 917.6138 |
| 2-phenylethyl benzoate | C9H10O2 | 30.735 | C9 | 29.855 | 34.8 | 917.7958 |
| cyclofeuchene | C10H16 | 31.294 | C9 | 29.855 | 34.8 | 929.1001 |
| α-pinene | C10H16 | 31.695 | C9 | 29.855 | 34.8 | 937.2093 |
| α-bulnesene | C15H24 | 32.283 | C9 | 29.855 | 34.8 | 949.1001 |
| α-fenchene | C10H16 | 32.425 | C9 | 29.855 | 34.8 | 951.9717 |
| camphene | C10H16 | 32.538 | C9 | 29.855 | 34.8 | 954.2568 |
| thujopsene | C15H24 | 33.098 | C9 | 29.855 | 34.8 | 965.5814 |
| 4-cyclopentyl ethylbenzoate | C14H18O2 | 33.532 | C9 | 29.855 | 34.8 | 974.3579 |
| 2,4-butylethylbenzoate | C13H18O2 | 33.561 | C9 | 29.855 | 34.8 | 974.9444 |
| pseudolimonene | C10H16 | 33.804 | C9 | 29.855 | 34.8 | 979.8584 |
| β-longipinene | C15H24 | 34.35 | C9 | 29.855 | 34.8 | 990.8999 |
| aciphyllene | C15H24 | 34.722 | C9 | 29.855 | 34.8 | 998.4226 |
| α-phellandrene | C10H16 | 34.809 | C10 | 38.719 | 34.8 | 1000.2297 |
| benzaldehyde | C7H6O | 35.052 | C10 | 38.719 | 34.8 | 1006.4302 |
| 3,4-dimethylbenzyl alcohol | C9H12O | 35.399 | C10 | 38.719 | 34.8 | 1015.2845 |
| limonene | C10H16 | 35.912 | C10 | 38.719 | 34.8 | 1028.3746 |
| p-cymene | C10H14 | 36.137 | C10 | 38.719 | 34.8 | 1034.1158 |
| β-phellandrene | C10H16 | 36.309 | C10 | 38.719 | 34.8 | 1038.5047 |
| (E)-β-ocimene | C10H16 | 36.451 | C10 | 38.719 | 34.8 | 1042.1281 |
| eucalyptol | C10H18O | 36.68 | C10 | 38.719 | 34.8 | 1047.9714 |
| 3-carene | C10H16 | 37.089 | C10 | 38.719 | 34.8 | 1058.4078 |
| γ-terpinene | C10H16 | 37.319 | C10 | 38.719 | 34.8 | 1064.2766 |
| aromadendrene | C15H24 | 37.745 | C11 | 42.954 | 38.719 | 1077.0012 |
| α-terpinolene | C10H16 | 38.885 | C11 | 42.954 | 38.719 | 1103.9197 |
| p-cumenol | C9H12O | 38.885 | C11 | 42.954 | 38.719 | 1103.9197 |
| γ-humulene | C15H24 | 39.052 | C11 | 42.954 | 38.719 | 1107.8630 |
| γ-himachalene | C15H24 | 39.086 | C11 | 42.954 | 38.719 | 1108.6659 |
| (-)-aristolene | C15H24 | 39.327 | C11 | 42.954 | 38.719 | 1114.3566 |
| 2,5-dimethylstyrene | C10H12 | 39.433 | C11 | 42.954 | 38.719 | 1116.8595 |
| β-maaliene | C15H24 | 40.158 | C11 | 42.954 | 38.719 | 1133.9787 |
| α-ylangene | C15H24 | 41.436 | C11 | 42.954 | 38.719 | 1164.1558 |
| β-copaene | C15H24 | 41.736 | C11 | 42.954 | 38.719 | 1171.2397 |
| δ-selinene | C15H24 | 41.828 | C11 | 42.954 | 38.719 | 1173.4120 |
| cis-muurola-3,5-diene | C15H24 | 41.837 | C11 | 42.954 | 38.719 | 1173.6246 |
| germacrene D | C15H24 | 42.338 | C11 | 42.954 | 38.719 | 1185.4545 |
| cis-muurola-4(14),5-diene | C15H24 | 42.392 | C11 | 42.954 | 38.719 | 1186.7296 |
| isoledene | C15H24 | 42.66 | C11 | 42.954 | 38.719 | 1193.0579 |
| valerena-4,7(11)-diene | C15H24 | 42.78 | C11 | 42.954 | 38.719 | 1195.8914 |
| 4-hexen-1-butylethylbenzoate | C10H18O2 | 42.868 | C11 | 42.954 | 38.719 | 1197.9693 |
| β-bourbonene | C15H24 | 43.123 | C12 | 44.8 | 42.954 | 1209.1549 |
| β-elemene | C15H24 | 43.511 | C12 | 44.8 | 42.954 | 1230.1733 |
| camphor | C10H16O | 43.515 | C12 | 44.8 | 42.954 | 1230.3900 |
| bicyclogermacrene | C15H24 | 44.246 | C12 | 44.8 | 42.954 | 1269.9892 |
| α-muurolene | C15H24 | 44.262 | C12 | 44.8 | 42.954 | 1270.8559 |
| endo-borneol | C10H18O | 44.308 | C12 | 44.8 | 42.954 | 1273.3478 |
| caryophyllene | C15H24 | 44.492 | C12 | 44.8 | 42.954 | 1283.3153 |
| α-terpineol | C10H18O | 44.731 | C12 | 44.8 | 42.954 | 1296.2622 |
References
- Loreto, F.; Schnitzler, J.P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Fineschi, S.; Loreto, F.; Staudt, M.; Peñuelas, J. Diversification of volatile isoprenoid emissions from trees: Evolutionary and ecological perspectives. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–20. [Google Scholar]
- Peñuelas, J.; Llusià, J. BVOCs: Plant defense against climate warming? Trends Plant Sci. 2003, 8, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Fehsenfeld, F.; Calvert, J.; Fall, R.; Goldan, P.; Guenther, A.B.; Hewitt, C.N.; Lamb, B.; Liu, S.; Trainer, M.; Westberg, H. Emissions of volatile organic compounds from vegetation and the implications for atmospheric chemistry. Glob. Biogeochem. Cycles 1992, 6, 389–430. [Google Scholar] [CrossRef]
- Calfapietra, C.; Fares, S.; Manes, F.; Morani, A.; Sgrigna, G.; Loreto, F. Role of Biogenic Volatile Organic Compounds (BVOC) emitted by urban trees on ozone concentration in cities: A review. Environ. Pollut. 2013, 183, 71–80. [Google Scholar] [CrossRef]
- Kulmala, M.; Nieminen, T.; Chellapermal, R.; Makkonen, R.; Bäck, J.; Kerminen, V.-M. Climate feedbacks linking the increasing atmospheric CO2 concentration, BVOC emissions, aerosols and clouds in forest ecosystems. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 489–508. [Google Scholar]
- Guenther, A.B.; Jiang, X.; Heald, C.L.; Sakulyanontvittaya, T.; Duhl, T.; Emmons, L.K.; Wang, X. The Model of Emissions of Gases and Aerosols from Nature version 2.1 (MEGAN2.1): An extended and updated framework for modeling biogenic emissions. Geosci. Model Dev. 2012, 5, 1471–1492. [Google Scholar] [CrossRef]
- Calfapietra, C.; Pallozzi, E.; Lusini, I.; Velikova, V. Modification of BVOC emissions by changes in atmospheric [CO2] and air pollution. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 253–284. [Google Scholar]
- Tani, A.; Masui, N.; Chang, T.; Okumura, M.; Kokubu, Y. Basal emission rates of isoprene and monoterpenes from major tree species in Japan: Interspecies and intraspecies variabilities. Prog. Earth Planet. Sci. 2024, 11, 42. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, Z.; Kännaste, A.; Guo, M.; Zhou, G.; Niinemets, Ü. Isoprenoid and aromatic compound emissions in relation to leaf structure, plant growth form and species ecology in 45 East-Asian urban subtropical woody species. Urban For. Urban Green. 2020, 53, 126705. [Google Scholar] [CrossRef]
- Copolovici, L.; Niinemets, Ü. Environmental impacts on plant volatile emission. In Deciphering Chemical Language of Plant Communication; Springer: Berlin/Heidelberg, Germany, 2016; pp. 35–59. [Google Scholar]
- Brilli, F.; Ciccioli, P.; Frattoni, M.; Prestininzi, M.; Spanedda, A.F.; Loreto, F. Constitutive and herbivore-induced monoterpenes emitted by Populus × euroamericana leaves are key volatiles that orient Chrysomela populi beetles. Plant Cell Environ. 2009, 32, 542–552. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Arneth, A.; Kuhn, U.; Monson, R.; Peñuelas, J.; Staudt, M. The emission factor of volatile isoprenoids: Stress, acclimation, and developmental responses. Biogeosciences 2010, 7, 2203–2223. [Google Scholar] [CrossRef]
- Guenther, A.B. Biological and chemical diversity of biogenic volatile organic emissions into the atmosphere. Int. Sch. Res. Not. 2013, 2013, 786290. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Llusià, J.; Filella, I.; Niinemets, Ü.; Arneth, A.; Wright, I.J.; Loreto, F.; Peñuelas, J. Nutrient-rich plants emit a less intense blend of volatile isoprenoids. New Phytol. 2018, 220, 773–784. [Google Scholar] [CrossRef]
- Ciccioli, P.; Centritto, M.; Loreto, F. Biogenic volatile organic compound emissions from vegetation fires. Plant Cell Environ. 2014, 37, 1810–1825. [Google Scholar] [CrossRef] [PubMed]
- Hanson, D.T.; Swanson, S.; Graham, L.E.; Sharkey, T.D. Evolutionary significance of isopreneemission from mosses. Am. J. Bot. 1999, 86, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Chen, X.; Yeh, S. Isoprene increases thermotolerance of fosmidomycin-fed leaves. Plant Physiol. 2001, 125, 2001–2006. [Google Scholar] [CrossRef]
- Salomón, R.L.; Rodríguez-Calcerrada, J.; Staudt, M. Carbon losses from respiration and emission of volatile organic compounds—The overlooked side of tree carbon budgets. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 327–359. [Google Scholar]
- Niinemets, Ü.; Rasulov, B.; Talts, E. CO2-responsiveness of leaf isoprene emission: Why do species differ? Plant Cell Environ. 2021, 44, 3049–3063. [Google Scholar] [CrossRef]
- Fischbach, R.J.; Staudt, M.; Zimmer, I.; Rambal, S.; Schnitzler, J.P. Seasonal pattern of monoterpene synthase activities in leaves of the evergreen tree Quercus ilex. Physiol. Plant. 2002, 114, 354–360. [Google Scholar] [CrossRef]
- Niinemets, Ü. What are plant-released biogenic volatiles and how they participate in landscape-to global-level processes? In Ecosystem Services from Forest Landscapes: Broadscale Considerations; Springer: Berlin/Heidelberg, Germany, 2018; pp. 29–56. [Google Scholar]
- Loreto, F.; Förster, A.; Dürr, M.; Csiky, O.; Seufert, G. On the monoterpene emission under heat stress and on the increased thermotolerance of leaves of Quercus ilex L. fumigated with selected monoterpenes. Plant Cell Environ. 1998, 21, 101–107. [Google Scholar] [CrossRef]
- Misztal, P.K.; Hewitt, C.N.; Wildt, J.; Blande, J.D.; Eller, A.S.D.; Fares, S.; Gentner, D.R.; Gilman, J.B.; Graus, M.; Greenberg, J. Atmospheric benzenoid emissions from plants rival those from fossil fuels. Sci. Rep. 2015, 5, 12064. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Storage of defense metabolites in the leaves of Myrtaceae: News of the eggs in different baskets. Tree Physiol. 2018, 38, 1445–1450. [Google Scholar] [CrossRef]
- Grote, R.; Sharma, M.; Ghirardo, A.; Schnitzler, J.P. A new modeling approach for estimating abiotic and biotic stress-induced de novo emissions of biogenic volatile organic compounds from plants. Front. For. Glob. Chang. 2019, 2, 26. [Google Scholar] [CrossRef]
- Pazouki, L.; Kanagendran, A.; Li, S.; Kännaste, A.; Memari, H.R.; Bichele, R.; Niinemets, Ü. Mono-and sesquiterpene release from tomato (Solanum lycopersicum) leaves upon mild and severe heat stress and through recovery: From gene expression to emission responses. Environ. Exp. Bot. 2016, 132, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kanagendran, A.; Pazouki, L.; Li, S.; Liu, B.; Kännaste, A.; Niinemets, Ü. Ozone-triggered surface uptake and stress volatile emissions in Nicotiana tabacum ‘Wisconsin’. J. Exp. Bot. 2018, 69, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- Rasulov, B.; Talts, E.; Niinemets, Ü. A novel approach for real-time monitoring of leaf wounding responses demonstrates unprecedently fast and high emissions of volatiles from cut leaves. Plant Sci. 2019, 283, 256–265. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Niinemets, Ü. Massive release of volatile organic compounds due to leaf midrib wounding in Populus tremula. Plant Ecol. 2018, 219, 1021–1028. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Okereke, C.N.; Jiang, Y.; Talts, E.; Kaurilind, E.; Niinemets, Ü. Wounding-induced VOC emissions in five tropical agricultural species. Molecules 2021, 26, 2602. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Direct fungicidal activities of C6-aldehydes are important constituents for defense responses in Arabidopsis against Botrytis cinerea. Phytochemistry 2008, 69, 2127–2132. [Google Scholar] [CrossRef]
- Kanagendran, A.; Pazouki, L.; Bichele, R.; Külheim, C.; Niinemets, Ü. Temporal regulation of terpene synthase gene expression in Eucalyptus globulus leaves upon ozone and wounding stresses: Relationships with stomatal ozone uptake and emission responses. Environ. Exp. Bot. 2018, 155, 552–565. [Google Scholar] [CrossRef]
- Kanagendran, A.; Pazouki, L.; Niinemets, Ü. Differential regulation of volatile emission from Eucalyptus globulus leaves upon single and combined ozone and wounding treatments through recovery and relationships with ozone uptake. Environ. Exp. Bot. 2018, 145, 21–38. [Google Scholar] [CrossRef]
- Arimura, G.i.; Ozawa, R.; Kugimiya, S.; Takabayashi, J.; Bohlmann, J.r. Herbivore-induced defense response in a model legume. Two-spotted spider mites induce emission of (E)-β-ocimene and transcript accumulation of (E)-β-ocimene synthase in Lotus japonicus. Plant Physiol. 2004, 135, 1976–1983. [Google Scholar] [CrossRef] [PubMed]
- Brilli, F.; Ruuskanen, T.M.; Schnitzhofer, R.; Müller, M.; Breitenlechner, M.; Bittner, V.; Wohlfahrt, G.; Loreto, F.; Hansel, A. Detection of plant volatiles after leaf wounding and darkening by proton transfer reaction “time-of-flight” mass spectrometry (PTR-TOF). PLoS ONE 2011, 6, e20419. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Staudt, M. BVOCs and global change. Trends Plant Sci. 2010, 15, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Männistö, E.; Ylänne, H.; Kokkonen, N.; Korrensalo, A.; Laine, A.M.; Yli-Pirilä, P.; Keinänen, M.; Tuittila, E.S. Impact of severe drought on biogenic volatile organic compounds emissions from Sphagnum mosses in boreal peatlands. Sci. Total Environ. 2024, 951, 175738. [Google Scholar] [CrossRef]
- Alamgir, A. Pharmacognostical Botany: Classification of medicinal and aromatic plants (MAPs), botanical taxonomy, morphology, and anatomy of drug plants. In Therapeutic Use of Medicinal Plants and Their Extracts: Volume 1: Pharmacognosy; Springer: Cham, Switzerland, 2017; pp. 177–293. [Google Scholar]
- Huang, X.; Jiang, H.; Hao, G. Direct HPLC detection of benzodilactones and quinones in glands of Lysimachia fordiana. Fitoterapia 2009, 80, 173–176. [Google Scholar] [CrossRef]
- Lersten, N.R.; Czlapinski, A.R.; Curtis, J.D.; Freckmann, R.; Horner, H.T. Oil bodies in leaf mesophyll cells of angiosperms: Overview and a selected survey. Am. J. Bot. 2006, 93, 1731–1739. [Google Scholar] [CrossRef]
- Tang, B.; Han, M.; Xu, Q.; Jin, J. Leaf cuticle microstructure of Machilus maomingensis sp. nov. (Lauraceae) from the Eocene of the Maoming Basin, South China. Acta Geol. Sin.-Engl. Ed. 2016, 90, 1561–1571. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, X.; Lai, Y.; Li, X.; Zhang, D.; Chen, Y. Efficient extraction of bioenergy from Cinnamomum camphora leaves. Front. Energy Res. 2020, 8, 90. [Google Scholar] [CrossRef]
- Shang, A.; Gan, R.; Zhang, J.; Xu, X.; Luo, M.; Liu, H.; Li, H. Optimization and characterization of microwave-assisted hydro-distillation extraction of essential oils from Cinnamomum camphora leaf and recovery of polyphenols from extract fluid. Molecules 2020, 25, 3213. [Google Scholar] [CrossRef]
- Peng, F.; Guo, H.; Hao, M.; Guo, J.; Yang, Y.; Tan, P. The ultrastructure characteristics of secretory cavities associated with the secretory products of Ginkgo biloba. Am. J. Plant Sci. 2011, 3, 102–109. [Google Scholar] [CrossRef]
- Ali, J.K.; Sosa, A.A. Anatomical study of some characters in certain species of genus Jasminum L. growing in Iraq. Int. J. Sci. Res. 2015, 5, 1137–1140. [Google Scholar]
- Kröber, W.; Heklau, H.; Bruelheide, H. Leaf morphology of 40 evergreen and deciduous broadleaved subtropical tree species and relationships to functional ecophysiological traits. Plant Biol. 2015, 17, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Xia, N. Leaf epidermal features of Lithocarpus (Fagaceae) from China and their systematic significance. Bot. J. Linn. Soc. 2012, 168, 216–228. [Google Scholar] [CrossRef]
- Marinho, C.R.; Zacaro, A.A.; Ventrella, M.C. Secretory cells in Piper umbellatum (Piperaceae) leaves: A new example for the development of idioblasts. Flora 2011, 206, 1052–1062. [Google Scholar] [CrossRef]
- Turner, G.W. A brief history of the lysigenous gland hypothesis. Bot. Rev. 1999, 65, 76–88. [Google Scholar] [CrossRef]
- Deng, M.; Hipp, A.; Song, Y.; Li, Q.; Coombes, A.; Cotton, A. Leaf epidermal features of Quercus subgenus Cyclobalanopsis (Fagaceae) and their systematic significance. Bot. J. Linn. Soc. 2014, 176, 224–259. [Google Scholar] [CrossRef]
- Sun, Z.; Niinemets, Ü.; Hüve, K.; Rasulov, B.; Noe, S.M. Elevated atmospheric CO2 concentration leads to increased whole-plant isoprene emission in hybrid aspen (Populus tremula × Populus tremuloides). New Phytol. 2013, 198, 788–800. [Google Scholar] [CrossRef]
- Velikova, V.; Loreto, F.; Tsonev, T.; Brilli, F.; Edreva, A. Isoprene prevents the negative consequences of high temperature stress in Platanus orientalis leaves. Funct. Plant Biol. 2006, 33, 931–940. [Google Scholar] [CrossRef]
- Velikova, V.; Sharkey, T.D.; Loreto, F. Increased Thermostability of Thylakoid Membranes in Isoprene-Emitting Leaves Probed with Three Biophysical Techniques. Plant Physiol. 2012, 157, 905–916. [Google Scholar] [CrossRef]
- Singsaas, E.L.; Lerdau, M.; Winter, K.; Sharkey, T.D. Isoprene increases thermotolerance of isoprene-emitting species. Plant Physiol. 1997, 115, 1413–1420. [Google Scholar] [CrossRef]
- Siwko, M.E.; Marrink, S.J.; de Vries, A.H.; Kozubek, A.; Uiterkamp, A.J.S.; Mark, A.E. Does isoprene protect plant membranes from thermal shock? A molecular dynamics study. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Mannozzi, M.; Maris, C.; Nascetti, P.; Ferranti, F.; Pasqualini, S. Ozone quenching properties of isoprene and its antioxidant role in leaves. Plant Physiol. 2001, 126, 993–1000. [Google Scholar] [CrossRef]
- Affek, H.P.; Yakir, D. Protection by isoprene against singlet oxygen in leaves. Plant Physiol. 2002, 129, 269–277. [Google Scholar] [CrossRef]
- Possell, M.; Loreto, F. The role of volatile organic compounds in plant resistance to abiotic stresses: Responses and mechanisms. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 209–235. [Google Scholar]
- Sun, Z.; Copolovici, L.; Niinemets, Ü. Can the capacity for isoprene emission acclimate to environmental modifications during autumn senescence in temperate deciduous tree species Populus tremula? J. Plant Res. 2012, 125, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Loreto, F. Water stress, temperature, and light effects on isoprene emission and photosynthesis of Kudzu leaves. Oecologia 1993, 95, 328–333. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Sun, Z.; Talts, E. Controls of the quantum yield and saturation light of isoprene emission in different-aged aspen leaves. Plant Cell Environ. 2015, 38, 2707–2720. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H. Non-mevalonate isoprenoid biosynthesis: Enzymes, genes and inhibitors. Biochem. Soc. Trans. 2000, 28, 785–789. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Yeh, S. Isoprene emission from plants. Annu. Rev. Plant Biol. 2001, 52, 407–436. [Google Scholar] [CrossRef]
- Velikova, V.; Pinelli, P.; Pasqualini, S.; Reale, L.; Ferranti, F.; Loreto, F. Isoprene decreases the concentration of nitric oxide in leaves exposed to elevated ozone. New Phytol. 2005, 166, 419–426. [Google Scholar] [CrossRef]
- Niinemets, Ü. Mild versus severe stress and BVOCs: Thresholds, priming and consequences. Trends Plant Sci. 2010, 15, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Shen, Y.; Niinemets, Ü. Responses of isoprene emission and photochemical efficiency to severe drought combined with prolonged hot weather in hybrid Populus. J. Exp. Bot. 2020, 71, 7364–7381. [Google Scholar] [CrossRef] [PubMed]
- Oku, H.; Mutanda, I.; Inafuku, M. Molecular characteristics of isoprene synthase and its control effects on isoprene emissions from tropical trees. J. Plant Res. 2023, 136, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.; Frank, T.D.; DeLucia, E.H. A method for quantitative analysis of spatially variable physiological processes across leaf surfaces. Photosynth. Res. 2006, 90, 161–172. [Google Scholar] [CrossRef]
- Tritsch, D.; Hemmerlin, A.; Bach, T.J.; Rohmer, M. Plant isoprenoid biosynthesis via the MEP pathway: In vivo IPP/DMAPP ratio produced by (E)-4-hydroxy-3-methylbut-2-enyl diphosphate reductase in tobacco BY-2 cell cultures. FEBS Lett. 2010, 584, 129–134. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Harrison, S.P.; Morfopoulos, C.; Dani, K.S.; Prentice, I.C.; Arneth, A.; Atwell, B.J.; Barkley, M.P.; Leishman, M.R.; Loreto, F.; Medlyn, B.E. Volatile isoprenoid emissions from plastid to planet. New Phytol. 2013, 197, 49–57. [Google Scholar] [CrossRef]
- Joó, É.; Dewulf, J.; Amelynck, C.; Schoon, N.; Pokorska, O.; Šimpraga, M.; Steppe, K.; Aubinet, M.; Van Langenhove, H. Constitutive versus heat and biotic stress induced BVOC emissions in Pseudotsuga menziesii. Atmos. Environ. 2011, 45, 3655–3662. [Google Scholar] [CrossRef]
- Peer, W.A.; Langenheim, J.H. Influence of phytochrome on leaf monoterpene variation in Satureja douglasii. Biochem. Syst. Ecol. 1998, 26, 25–34. [Google Scholar] [CrossRef]
- Loreto, F.; Nascetti, P.; Graverini, A.; Mannozzi, M. Emission and content of monoterpenes in intact and wounded needles of the Mediterranean pine, Pinus pinea. Funct. Ecol. 2000, 14, 589–595. [Google Scholar] [CrossRef]
- Zabaras, D.; Spooner-Hart, R.; Wyllie, S.G. Effects of mechanical wounding on concentration and composition of essential oil from Melaleuca alternifolia leaves. Biochem. Syst. Ecol. 2002, 30, 399–412. [Google Scholar] [CrossRef]
- Kask, K.; Kaurilind, E.; Talts, E.; Kännaste, A.; Niinemets, Ü. Combined acute ozone and water stress alters the quantitative relationships between O3 uptake, photosynthetic characteristics and volatile emissions in Brassica nigra. Molecules 2021, 26, 3114. [Google Scholar] [CrossRef]
- Goodger, J.Q.; Senaratne, S.L.; Nicolle, D.; Woodrow, I.E. Differential metabolic specialization of foliar oil glands in Eucalyptus brevistylis Brooker (Myrtaceae). Tree Physiol. 2018, 38, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Goodger, J.Q.; Seneratne, S.L.; Nicolle, D.; Woodrow, I.E. Foliar essential oil glands of Eucalyptus subgenus Eucalyptus (Myrtaceae) are a rich source of flavonoids and related non-volatile constituents. PLoS ONE 2016, 11, e0151432. [Google Scholar]
- Ekanayaka, E.P.; Li, C.; Jones, A.D. Sesquiterpenoid glycosides from glandular trichomes of the wild tomato relative Solanum habrochaites. Phytochemistry 2014, 98, 223–231. [Google Scholar] [CrossRef]
- Paré, P.W.; Tumlinson, J.H. Plant volatiles as a defense against insect herbivores. Plant Physiol. 1999, 121, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Piesik, D.; Łyszczarz, A.; Tabaka, P.; Lamparski, R.; Bocianowski, J.; Delaney, K. Volatile induction of three cereals: Influence of mechanical injury and insect herbivory on injured plants and neighbouring uninjured plants. Ann. Appl. Biol. 2010, 157, 425–434. [Google Scholar] [CrossRef]
- Kopaczyk, J.M.; Warguła, J.; Jelonek, T. The variability of terpenes in conifers under developmental and environmental stimuli. Environ. Exp. Bot. 2020, 180, 104197. [Google Scholar] [CrossRef]
- Grote, R.; Monson, R.K.; Niinemets, Ü. Leaf-level models of constitutive and stress-driven volatile organic compound emissions. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 315–355. [Google Scholar]
- Monson, R.K. Metabolic and gene expression controls on the production of biogenic volatile organic compounds. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 153–179. [Google Scholar]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a key floral and foliar volatile involved in multiple interactions between plants and other organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef]
- Smith, W.; Shivaji, R.; Williams, W.; Luthe, D.; Sandoya, G.; Smith, C.; Sparks, D.; Brown, A. A maize line resistant to herbivory constitutively releases (E)-β-caryophyllene. J. Econ. Entomol. 2012, 105, 120–128. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Nerg, A.M.; Blande, J.D. Multitrophic signalling in polluted atmospheres. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 285–314. [Google Scholar]
- Blande, J.D.; Holopainen, J.K.; Niinemets, Ü. Plant volatiles in polluted atmospheres: Stress responses and signal degradation. Plant Cell Environ. 2014, 37, 1892–1904. [Google Scholar] [CrossRef]
- Wang, F.; Park, Y.L.; Gutensohn, M. Glandular trichome-derived mono-and sesquiterpenes of tomato have contrasting roles in the interaction with the potato aphid Macrosiphum euphorbiae. J. Chem. Ecol. 2021, 47, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.; Cui, Z.; Shu, B.; Zhao, S.; Yu, X. Histochemistry and cell wall specialization of oil cells related to the essential oil accumulation in the bark of Cinnamomum cassia Presl. (Lauraceae). Plant Prod. Sci. 2012, 15, 1–9. [Google Scholar] [CrossRef]
- Barbosa, L.C.A.; Filomeno, C.A.; Teixeira, R.R. Chemical variability and biological activities of Eucalyptus spp. essential oils. Molecules 2016, 21, 1671. [Google Scholar] [CrossRef]
- Serrato-Valenti, G.; Bisio, A.; Cornara, L.; Ciarallo, G. Structural and histochemical investigation of the glandular trichomes of Salvia aurea L. leaves, and chemical analysis of the essential oil. Ann. Bot. 1997, 79, 329–336. [Google Scholar] [CrossRef]
- Stefanello, M.É.A.; Pascoal, A.C.; Salvador, M.J. Essential oils from neotropical Myrtaceae: Chemical diversity and biological properties. Chem. Biodivers. 2011, 8, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Kännaste, A.; Copolovici, L.; Niinemets, Ü. Gas chromatography–mass spectrometry method for determination of biogenic volatile organic compounds emitted by plants. In Plant Isoprenoids: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2014; pp. 161–169. [Google Scholar]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Nagesh, K.M.; Piovene, C. Emission and function of volatile organic compounds in response to abiotic stress. In Abiotic Stress in Plants: Mechanisms and Adaptations; IntechOpen: London, UK, 2011; 367p. [Google Scholar]
- Liu, B.; Kaurilind, E.; Jiang, Y.; Niinemets, Ü. Methyl salicylate differently affects benzenoid and terpenoid volatile emissions in Betula pendula. Tree Physiol. 2018, 38, 1513–1525. [Google Scholar] [CrossRef]
- Nagel, J.; Culley, L.K.; Lu, Y.; Liu, E.; Matthews, P.D.; Stevens, J.F.; Page, J.E. EST analysis of hop glandular trichomes identifies an O-methyltransferase that catalyzes the biosynthesis of xanthohumol. Plant Cell 2008, 20, 186–200. [Google Scholar] [CrossRef]
- Zager, J.J.; Lange, B.M. Assessing flux distribution associated with metabolic specialization of glandular trichomes. Trends Plant Sci. 2018, 23, 638–647. [Google Scholar] [CrossRef]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.; Arora, S. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Okereke, C.N.; Liu, B.; Kaurilind, E.; Niinemets, Ü. Heat stress resistance drives coordination of emissions of suites of volatiles after severe heat stress and during recovery in five tropical crops. Environ. Exp. Bot. 2021, 184, 104375. [Google Scholar] [CrossRef]
- Nagalingam, S.; Seco, R.; Kim, S.; Guenther, A. Heat stress strongly induces monoterpene emissions in some plants with specialized terpenoid storage structures. Agric. For. Meteorol. 2023, 333, 109400. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Onoda, Y.; Wright, I.J.; Evans, J.R.; Hikosaka, K.; Kitajima, K.; Niinemets, Ü.; Poorter, H.; Tosens, T.; Westoby, M. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytol. 2017, 214, 1447–1463. [Google Scholar] [CrossRef]
- Tilney, P.M.; Nel, M.; van Wyk, A.E. Foliar secretory structures in Melia azedarach (Meliaceae), a widely cultivated and often invasive tree. N. Z. J. Bot. 2018, 56, 198–215. [Google Scholar] [CrossRef]

| Number | Family | Species | Presence of Terpene-Storage Structures with Reference | Plant Functional Type | Species Ecological Characteristics |
|---|---|---|---|---|---|
| 1 | Aceraceae | Acer buergerianum | No [41] | Deciduous | HT; CT |
| 2 | Aceraceae | Acer cinnamomifolium | No [41] | Deciduous | HT; LT |
| 3 | Aceraceae | Acer henryi | No [41] | Deciduous | HT; CT |
| 4 | Aceraceae | Acer palmatum | No [41] | Deciduous | HT; CT; ST |
| 5 | Moraceae | Broussonetia papyrifera | No | Deciduous | LT |
| 6 | Theaceae | Camellia japonica | No | Evergreen | HT; ST |
| 7 | Nyssaceae | Camptotheca acuminata | No [42] | Deciduous | LT |
| 8 | Fagaceae | Castanopsis sclerophylla | No | Evergreen | LT |
| 9 | Calycanthacecae | Chimonanthus praecox | Oil cells [43] | Deciduous | CT; LT |
| 10 | Lauraceae | Cinnamomum camphora | Oil cells [44,45,46] and glandular trichomes [45,46] | Evergreen | HT; ST; LT |
| 11 | Lauraceae | Cinnamomum japonicum | Oil cells [44,45] | Evergreen | HT; LT |
| 12 | Fagaceae | Cyclobalanopsis glauca | No | Evergreen | CT; ST |
| 13 | Fagaceae | Cyclobalanopsis myrsinifolia | No | Evergreen | CT; ST |
| 14 | Elaeocarpaceae | Elaeocarpus glabripetalus var. glabripetalus | No | Evergreen | LT |
| 15 | Rubiaceae | Gardenia jasminoides | No [41] | Evergreen | HT |
| 16 | Ginkgoaceae | Ginkgo biloba | Resin ducts [47] | Deciduous | LT; CT |
| 17 | Aquifoliaceae | Ilex chinensis | No | Evergreen | HT; CT; ST |
| 18 | Oleaceae | Jasminum mesnyi | Glandular trichomes [48] | Deciduous | HT; ST; LT |
| 19 | Sapindaceae | Koelreuteria bipinnata var. integrifoliola | No [49] | Deciduous | HT; CT; LT |
| 20 | Oleaceae | Ligustrum lucidum | No | Evergreen | HT; CT; ST; LT |
| 21 | Magnoliaceae | Liriodendron chinense | Oil cells [43] | Deciduous | HT; LT |
| 22 | Fagaceae | Lithocarpus harlandii | No [50] | Evergreen | HT; LT |
| 23 | Lauraceae | Machilus leptophylla | Oil cells [44,45] | Evergreen | ST |
| 24 | Lauraceae | Machilus pauhoi | Oil cells [44,46] | Evergreen | ST |
| 25 | Lauraceae | Machilus thunbergii | Oil cells [44] | Evergreen | ST |
| 26 | Magnoliaceae | Magnolia denudata | Oil cells [43,44] | Deciduous | HT; LT |
| 27 | Magnoliaceae | Magnolia grandiflora | Oil cells [43,51] | Evergreen | HT; CT; LT |
| 28 | Magnoliaceae | Magnolia liliflora | Oil cells [43,51] | Deciduous | CT; LT |
| 29 | Meliaceae | Melia azedarach | No | Deciduous | HT; LT |
| 30 | Magnoliaceae | Michelia chapensis | Oil cells [43,51] | Evergreen | HT; LT |
| 31 | Magnoliaceae | Michelia figo | Oil cells [43] | Evergreen | HT; LT |
| 32 | Magnoliaceae | Michelia foveolata | Oil cells [43] | Evergreen | HT |
| 33 | Magnoliaceae | Michelia maudiae | Oil cells [43,51] | Evergreen | HT; LT |
| 34 | Oleaceae | Osmanthus fragrans | No [41] | Evergreen | HT; LT |
| 35 | Magnoliaceae | Parakmeria lotungensis | Oil cells [43,51] | Evergreen | HT; LT |
| 36 | Rosaceae | Photinia × fraseri | No | Evergreen | HT; LT |
| 37 | Rosaceae | Photinia serrulata | No [41] | Evergreen | HT; LT |
| 38 | Pittosporaceae | Pittosporum tobira | Oil ducts [52] | Evergreen | LT |
| 39 | Platanaceae | Platanus orientalis | No | Deciduous | HT; CT; LT |
| 40 | Rosaceae | Prunus mume | No [41] | Deciduous | CT; LT |
| 41 | Rosaceae | Pyracantha fortuneana | No | Evergreen | CT; LT |
| 42 | Rosaceae | Quercus serrata var. brevipetiolata | Glandular trichomes [53] | Deciduous | CT; LT |
| 43 | Sapindaceae | Sapindus mukorossi | No | Deciduous | LT |
| 44 | Leguminosae | Sophora japonica | No [41] | Deciduous | HT; LT |
| 45 | Leguminosae | Sophora japonica var. japonica f. pendula | No [41] | Deciduous | HT; LT |
| Compound | Changes in Compounds After Mechanical Injury | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Functional Type | Storage Species | Non-Storage Species | |||||||||
| Increased | New Compound | Reduced | Disappeared | Increased | New Compound | Reduced | Disappeared | ||||
| Monoterpenes | C10H16 | α-pinene | Deciduous Evergreen | 9, 16 *, 18, 21,26, 28, 42 * 10 *, 11 *, 23, 24 *, 30, 31 *, 33 *, 35 *, 38 * | 27, 32 | 3, 4, 7, 29, 40, 43, 45; 14, 17 *, 20 *, 36 | 1; 34 * | 5, 19; 6, 8 *, 12, 13, 22, 37, 41 | |||
| C10H16 | α-terpinolene | Deciduous Evergreen | 9, 28; 10 *, 23, 24, 25, 30, 32 *, 33 * | 38 * | 12 | 1 * | 13 *, 34 * | 8 *, 22 * | |||
| C10H16 | limonene | Deciduous Evergreen | 9 *, 21, 28 *, 42 * 10 *, 23 *, 24, 25 *, 27 *, 30, 32, 33 *, 35 *, 38 * | 11 *, 31 * | 12, 37 | 1 *, 7 *, 43 *, 6 * | 8 *, 13, 34 * | 3 *22 * | |||
| C10H16O | camphor | Deciduous Evergreen | 9 *, 16 *, 18, 26; 10 *, 33 * | 28 *; 11 *, 27 *, 32 *, 35 *, 38 * | 25 | 1, 2, 40, 42, 45; 12, 14, 15, 17, 36, 37 | 3 *, 7, 39 *; 34 * | 4, 5, 29, 43; 6, 13, 22, 41 | |||
| C10H16 | cyclofeuchene | Deciduous Evergreen | 33 *, 38 * | 10 *, 35 * | 23, 24 | 45 *; 8 *, 34 * | 13 * | ||||
| C10H16 | β-phellandrene | Deciduous Evergreen | 9, 28; 23 *, 24, 30 *, 32 *, 33 *, 35 * | 10 *, 11 *, 38 * | |||||||
| C10H16 | α-phellandrene | Deciduous Evergreen | 10 *, 23 *, 24, 30, 32 *, 33 * | 21 *; 11 *, 35 *, 38 * | 1 * | 8 *, 22 * | |||||
| C10H16 | camphene | Deciduous Evergreen | 28; 11 *, 23, 24, 27 *, 30, 32 *, 33 *, 38 * | 21 *; 10 *, 25 *, 31 *, 35 * | 12, 41 | 13 *, 34 * | 8 *, 36 * | ||||
| C10H16 | (E)-β-ocimene | Deciduous Evergreen | 28 *; 10 *, 33 | 9 *, 21 *; 11 *, 23 *, 24 *, 35 *, 38 * | 32 | 36 * | 22 * | 2 *; 8 *, 41 * | |||
| C10H16 | γ-terpinene | Deciduous Evergreen | 9 *, 21 *, 28 *; 10 *, 23, 24, 25 *, 27 *, 30 *, 32, 33 * | 11 *, 35 *, 38 * | |||||||
| C10H16 | pseudolimonene | Deciduous Evergreen | 9, 21 *; 10 *, 23, 24, 27 *, 28, 30, 33 *, 35 * | 11 *, 25 *, 38 * | 1; 34 *, 42 * | 13 | 8 *, 22 * | ||||
| C10H16 | α-terpinene | Deciduous Evergreen | 10 *, 23, 25 *, 27 *, 30, 32, 33 * | 21 *, 28 *; 35 *, 38 * | 13 | 8 * | |||||
| C10H14 | p-cymene | Deciduous Evergreen | 9 *, 28 *; 10 *, 11 *, 23 *, 27, 30, 33 * | 21 *; 25 *, 35 * | 24, 32 | 38 * | 13 *, 34 * | 8 *, 22 * | |||
| C10H16 | α-fenchene | Deciduous Evergreen | 21; 10 *, 23, 30, 32 * | 27 *, 35 *, 38 * | 8 *, 13 *, 34 * | ||||||
| C10H16 | α-thujene | Deciduous Evergreen | 28 *; 30 *, 32, 33 * | 25 | 27 * | ||||||
| C10H18O | eucalyptol | Deciduous Evergreen | 28; 10 *, 30, 32 *, 33 *, 35 * | 21 *; 23 * | |||||||
| C10H16 | 3-carene | Deciduous Evergreen | 21 *; 10 *, 30 *, 35 *, 38 * | ||||||||
| C10H16 | β-fenchene | Deciduous Evergreen | 38 * | 10 *, 23 *, 33 *, 35 * | |||||||
| C10H18O | α-terpineol | Deciduous Evergreen | 28 *; 10 *, 33 *, 35 * | ||||||||
| C10H18O | trans-4-thujanol | Deciduous Evergreen | 10 *, 30 *, 33 *, 35 * | ||||||||
| C10H18O | endo-borneol | Deciduous Evergreen | 10 *, 11 *, 33 *, 35 *, 38 * | ||||||||
| C10H18O | fenchol | Deciduous Evergreen | 33 *, 35 *, 38 * | ||||||||
| Sesquiterpene | C15H24 | α-bulnesene | Deciduous Evergreen | 28; 11, 23, 27, 32 | 38 * | 3 * | |||||
| C15H24 | caryophyllene | Deciduous Evergreen | 11 *, 23 *, 24 *, 27 * | ||||||||
| C15H24 | β-longipinene | Deciduous Evergreen | 28 *; 10 *, 11 *, 23 *, 24 *, 32 *, 35 *, 38 * | 19 *; 34 * | |||||||
| C15H24 | γ-humulene | Deciduous Evergreen | 28 *; 10 *, 11 *, 32 * | ||||||||
| C15H24 | germacrene | Deciduous Evergreen | 28 *; 23 *, 31 * | ||||||||
| C15H24 | cis-muurola-3,5-diene | Deciduous Evergreen | 21 * | ||||||||
| C15H24 | cis-muurola-4(14),5-diene | Deciduous Evergreen | 10 *, 11 *, 23 *, 27 *, 31 *, 38 * | 4 *, 5 *, 44 *, 45 *; 8 *, 22 * | |||||||
| C15H24 | β-maaliene | Deciduous Evergreen | 11 *, 23 *, 24 *, 32 * | ||||||||
| C15H24 | α-muurolene | Deciduous Evergreen | 23 *, 24 *, 32 * | 3 *, 45 * | |||||||
| C15H24 | bicyclogermacrene | Deciduous Evergreen | 10 *, 11 *, 23 *, 27 *, 32 *, 38 * | 3 *, 19 * | |||||||
| C15H24 | β-copaene | Deciduous Evergreen | 11 *, 23 *, 32 *, 35 *, 38 * | 19 *, 45 * | |||||||
| C15H24 | δ-guaiene | Deciduous Evergreen | 38 * | 3 *, 19 * | |||||||
| Aromatic compound | C14H18O2 | 4-cyclopentyl ethylbenzoate | Deciduous Evergreen | 16 *, 18, 42 | 26; 10, 23, 32, 35 * | 9 * 38 * | 6 *, 20, 37 | 17 * | 13 *, 15, 34 * | 4 *; 22 * | |
| C10H12 | 2,5-dimethylstyrene | Deciduous Evergreen | 28; 10 *, 23, 24, 27 *, 30 *, 33 *, 35 *, 38 * | 21 * | 1, 2 * | 3 * 12, 34 * | 8 *, 13 *, 22 * | ||||
| C9H10O2 | 2-phenylethyl formate | Deciduous Evergreen | 25 *, 30 *, 32 * | ||||||||
| C9H12O | 3,4-dimethylbenzyl alcohol | Deciduous Evergreen | 30 *, 33 *, 35 * | ||||||||
| C15H14O2 | 2-phenylethyl benzoate | Deciduous Evergreen | 18 * | 5; 37 * | 29 *, 40 * 12 * | 14 * | |||||
| C13H18O2 | 2,4-butylethylbenzoate | Deciduous Evergreen | 7 *; 8 * | 43 *; 12, 14, 41 | |||||||
| Compound Class | Presence of Storage Structures | Treatment | Isoprene | Monoterpenes | Sesquiterpenes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Deciduous | Deciduous | Evergreen | Deciduous | Evergreen | |||||||
| Intact | Damaged | Intact | Damaged | Intact | Damaged | Intact | Intact | Damaged | |||
| Monoterpenes | Non-storage | Intact | r = 0.656 *, n = 9 | r = 0.805 *, n = 9 | —— | r = 0.265, n = 33 | —— | r = 0.813 **, n = 39 | r = −0.185, n = 18 | n = 3 | n = 3 |
| Damaged | r = −0.237, n = 15 | r = −0.297, n = 15 | —— | —— | —— | —— | n = 3 | —— | —— | ||
| Storage | Intact | n = 3 | n = 3 | —— | r = 0.807 *, n = 18 | —— | r = 0.671 *, n = 36 | r = 0.695 *, n = 9 | r = 0.606 *, n = 27 | r = −0.097, n = 18 | |
| Damaged | n = 3 | n = 3 | —— | —— | —— | —— | n = 6 | r = 0.475, n = 27 | r = −0.350, n = 18 | ||
| Sesquiterpenes | Non-storage | Intact | r = −0.792 *, n = 9 | r = 0.149, n = 9 | r = −0.156, n = 15 | r = −0.240, n = 18 | n = 3 | n = 3 | —— | n = 3 | n = 3 |
| Damaged | —— | —— | —— | —— | —— | —— | n = 3 | —— | —— | ||
| Storage | Intact | —— | —— | —— | —— | r = 0.179, n = 27 | r = 0.224, n = 27 | n = 3 | —— | r = −0.082, n = 18 | |
| Damaged | —— | —— | —— | —— | r = 0.149, n = 15 | r = −0.101, n = 15 | n = 3 | —— | —— | ||
| Aromatic compounds | Non-storage | Intact | n = 3 | n = 3 | r = 0.710 *, n = 18 | r = 0.458 *, n = 21 | r = 0.555 **, n = 36 | r = 0.315, n = 33 | r = −0.751 *, n = 12 | n = 3 | n = 3 |
| Damaged | —— | n = 3 | r = 0.554 *, n = 18 | r = 0.136, n = 21 | r = −0.095, n = 36 | r = −0.276, n = 33 | n = 3 | —— | —— | ||
| Storage | Intact | —— | —— | r = −0.107, n = 15 | r = −0.056, n = 18 | r = 0.287, n = 33 | r = −0.088, n = 33 | n = 6 | r = 0.640 *, n = 24 | r = 0.169, n = 21 | |
| Damaged | —— | —— | r = −0.311, n = 15 | r = −0.170, n = 15 | r = 0.688 **, n = 36 | r = 0.749 **, n = 36 | n = 6 | r = 0.271, n = 27 | r = 0.220, n = 18 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Mao, Y.; Yuan, H.; Guo, M.; Zhou, G.; Niinemets, Ü.; Sun, Z. Impacts of Mechanical Injury on Volatile Emission Rate and Composition in 45 Subtropical Woody Broad-Leaved Storage and Non-Storage Emitters. Plants 2025, 14, 821. https://doi.org/10.3390/plants14050821
Yuan Y, Mao Y, Yuan H, Guo M, Zhou G, Niinemets Ü, Sun Z. Impacts of Mechanical Injury on Volatile Emission Rate and Composition in 45 Subtropical Woody Broad-Leaved Storage and Non-Storage Emitters. Plants. 2025; 14(5):821. https://doi.org/10.3390/plants14050821
Chicago/Turabian StyleYuan, Yali, Yimiao Mao, Hao Yuan, Ming Guo, Guomo Zhou, Ülo Niinemets, and Zhihong Sun. 2025. "Impacts of Mechanical Injury on Volatile Emission Rate and Composition in 45 Subtropical Woody Broad-Leaved Storage and Non-Storage Emitters" Plants 14, no. 5: 821. https://doi.org/10.3390/plants14050821
APA StyleYuan, Y., Mao, Y., Yuan, H., Guo, M., Zhou, G., Niinemets, Ü., & Sun, Z. (2025). Impacts of Mechanical Injury on Volatile Emission Rate and Composition in 45 Subtropical Woody Broad-Leaved Storage and Non-Storage Emitters. Plants, 14(5), 821. https://doi.org/10.3390/plants14050821