Abstract
Calmodulins (CaMs), which are important calcium-binding proteins, play critical roles in plant stress responses. However, limited information is available regarding the biological functions of CaMs under drought stress. In this study, we identified and isolated a CaM gene, ZmCaM2, from maize (Zea mays L.) in length and encodes a 184-amino acid protein containing four EF-hand domains capable of specifically binding calcium ions (Ca2+). Subcellular localization analysis revealed that ZmCaM2 is localized to the nucleus and membrane. Functional characterization indicated that ZmCaM2 negatively regulates drought tolerance in maize by increasing malondialdehyde (MDA) and reactive oxygen species (ROS) content while decreasing antioxidant enzyme activity, proline (Pro) content, abscisic acid (ABA) content and relative water content (RWC). Moreover, ZmCaM2 reduced maize sensitivity to ABA treatment, suggesting that ZmCaM2 negatively regulates the drought tolerance of maize by relying on the ABA pathway. These findings provide new insights into the functional role of ZmCaM2 and may facilitate the development of drought-resistant maize cultivars.
1. Introduction
With global warming and the increasing frequency of extreme weather events, drought stress has become a major factor limiting crop growth, development, and production [1,2]. For instance, severe drought from 1980 to 2015 reduced the yields of wheat and maize by 21% and 40%, respectively [3]. Drought stress caused approximately USD 30 billion in agricultural losses over the past two decades [4]. Thus, enhancing crop drought tolerance is a crucial goal for ensuring global food security. Zea maize (Zea mays L.), one of the world’s most important crops, serves as a key source of food, animal feed, and industrial raw materials. Its growth and development are severely affected by drought stress, resulting in yield losses of 30–90% [5]. Therefore, applying molecular breeding strategies to enhance drought tolerance is essential for sustainable maize production.
Calcium ions (Ca2+), as important second messengers, play vital roles in regulating various abiotic stress responses and in plant growth and development [6,7]. External stimuli can induce rapid changes in cytosolic Ca2+ concentration. Ca2+ binds to Ca2+-binding proteins (also known as Ca2+ sensors), thereby mediating signal transduction and triggering specific cellular responses [8]. Ca2+ sensors can be divided into several groups, such as calmodulin (CaM), calmodulin-like proteins (CMLs), calcium-dependent protein kinases (CDPKs), calcineurin B-like proteins (CBLs), and calcium and calmodulin-dependent protein kinases (CCaMKs) [9,10,11]. CDPKs and CCaMKs act as sensor–responder proteins, while CaMs, CMLs, and CBLs serve as sensor–relay proteins [12,13]. CaMs lack additional functional domains and inherent enzymatic activities [14]. CaMs have been extensively identified in multiple plant species. Specifically, 7 CaMs exist in Arabidopsis [15], 5 in Oryza sativa [16], 25 in Brassica napus [17], 4 in Chrysanthemum seticuspe [18], 5 in barley [19], 18 in wheat [20], and 3 in Vitis vinifera [21]. These proteins are highly conserved and represent some of the most ubiquitous Ca2+ sensors in eukaryotic organisms. Structurally, each CaM contains four canonical EF-hand motifs [22].
Growing evidence suggests that CaMs play essential roles in regulating plant growth and development, as well as in orchestrating responses to diverse abiotic and biotic stresses. The overexpression of GmCaM4 enhances salt tolerance via increased DNA-binding activity of MYB [23]. Overexpression of GmCaM4 improves soybean resistance to pathogens through activation of pathogenesis-related genes [24]. Similarly, overexpression of OsCaM1-1 in rice enhances tolerance to salt and heat stress [25,26]. The OsCaM1 mediated CCaMK-MKK1/6 cascade plays a positive role in regulating lateral root growth in rice [27]. In Arabidopsis, AtCaM1 and AtCaM4 increase salt tolerance by promoting nitric oxide (NO) accumulation [28]. AtCaM1, as a positive regulator, participates in leaf senescence, abscisic acid (ABA) signaling, and reactive oxygen species (ROS) accumulation [29]. AtCaM4 interacts with PATL1 and reduces freezing tolerance in Arabidopsis by modulating the expression of KIN1, COR15b, and COR8.6 [30,31]. Furthermore, AtCaM4 negatively regulates ROS accumulation in the wound-signaling pathway through activation of AtMPK8 [30]. AtCaM3 regulates the heat shock signaling pathway, and its overexpression enhances thermotolerance in Arabidopsis [32]. The overexpression of CaMs enhanced the plant’s tolerance to low temperatures [33]. Additionally, Ca2+/CaM2 interacts with CYCLIC NUCLEOTIDE GATED CHANNEL 15 (CNGC15s) to maintain calcium oscillatory signaling and promote root nodule symbiosis in Medicago truncatula [34]. CsCaM3 enhances cucumber tolerance to high-temperature stress by increasing the activity of antioxidant enzymes superoxide dismutase (SOD) and peroxidase (POD), and regulating the expression of genes involved in the chlorophyll biosynthesis and ABA signaling pathways [35]. Overexpression of StCaM2 reduces the accumulation of ROS in tobacco, thereby enhancing its tolerance to drought and salt stress [36]. HvCaM1 silencing lines exhibit increased tolerance to salt stress [37].
The plant hormone ABA plays a central role in regulating plant responses to abiotic and biotic stresses [38,39]. Accumulating evidence indicates that Ca2+ sensors are integral components of the ABA signaling pathway [40]. CML37, CML38, and CML39 are transcriptionally induced by salt, drought, and ABA [41]. AtCML9 negatively regulates salt tolerance through the ABA-mediated signaling pathway [42]. AtCML20 functions as a negative regulator of drought tolerance through ABA-dependent pathways [43]. EcCaM overexpressing Arabidopsis exhibits hypersensitivity to ABA during seed germination and enhanced tolerance to drought and salinity stress [44]. Collectively, Ca2+ and ABA signaling constitute an intricate regulatory network controlling plant responses to abiotic stress.
Previously, we identified CaM genes in maize and observed that ZmCaM2 can respond to drought stress based on transcriptome analysis [45]. However, the function of ZmCaM2 in response to drought stress remains unclear. In this work, we conducted a detailed characterization of ZmCaM2, including subcellular localization and Ca2+-binding analyses, and elucidated its function in maize during drought stress. These findings lay the groundwork for the application of molecular breeding strategies to develop drought-tolerant maize cultivars.
2. Results
2.1. Cloning and Analysis of the ZmCaM2 Gene
A 552 bp coding sequence (CDS) region was cloned from the maize inbred line B73 and designated as ZmCaM2 (Zm00001d043144). ZmCaM2 encodes a protein of 184 amino acids, with a molecular weight of 20.47 kDa and a theoretical isoelectric point (pI) of 4.84. The gene is located on chromosome 3 and contains no predicted signal peptide. The negative grand average of hydropathicity (GRAVY) value (−0.743) suggests that ZmCaM2 is a hydrophilic protein. Multiple sequence alignment showed that ZmCaM2 contains four conserved EF-hand domains. Phylogenetic analysis indicated that ZmCaM2 is most closely related to OsCaM3 (Figure 1a,b).
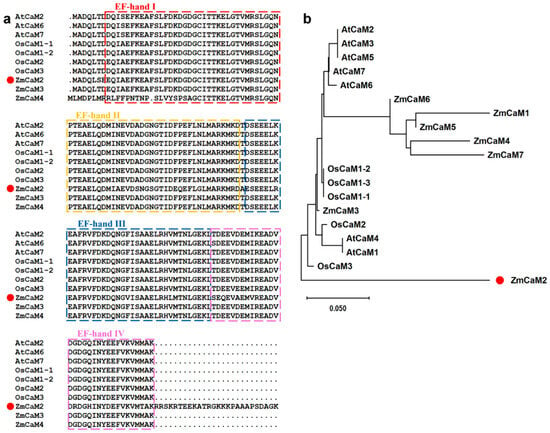
Figure 1.
Multiple sequence alignment and phylogenetic analysis of ZmCaM2. (a) Alignment of amino acid sequences of the ZmCaM2 and other CaM proteins from Arabidopsis and rice, which the EF-hand motifs are underlined; (b) Phylogenetic tree analysis of ZmCaM2 and other orthologs in different species. The accession numbers are as follows: AtCaM1 (AT5G37780), AtCaM2 (At2g27030), AtCaM3 (At3g56800), AtCaM5 (At2g41110), AtCaM6 (At5g21274), AtCaM7 (At3g43810), OsCaM1-1 (LOC_Os03g20370), OsCaM1-2 (LOC_Os07g48780), OsCaM1-3(LOC_Os01g16240), OsCaM2 (LOC_Os05g41210), OsCaM3 (LOC-_Os01g17190), ZmCaM1 (Zm00001d028948), ZmCaM2 (Zm00001d043144), ZmCaM2-1 (Zm00001d040323), ZmCaM3 (Zm00001d038543), ZmCaM4 (Zm00001d038545), ZmCaM5 (Zm00001d022546), ZmCaM6(Zm00001d008278), ZmCaM7 (Zm00001d047597).
2.2. ZmCaM2 Is Involved in the Response to PEG and ABA Induced Stress
To verify the involvement of ZmCaM2 in polyethylene glycol (PEG) and ABA-induced stress responses, uniformly grown maize seedlings at the third-leaf stage (V3 stage) were treated with 20% PEG 6000 (irrigation) or 200 µM ABA (foliar application). The transcriptional level of ZmCaM2 in maize leaves was quantified by quantitative real-time polymerase chain reaction (qRT-PCR) under treatment with 20% PEG6000 and 200 µM ABA. Under 20% PEG6000-simulated drought stress, ZmCaM2 expression remained slightly above the untreated control during the early stages, increased markedly after 12 h, and reached a maximum (5.2-fold) at 24 h (Figure 2a). Treatment with 200 µM ABA significantly induced ZmCaM2 expression, which peaked at 6 h (5.1-fold) (Figure 2b). These findings suggest that ZmCaM2 plays a critical role in regulating plant tolerance to drought stress and in the response to the ABA signaling pathway.
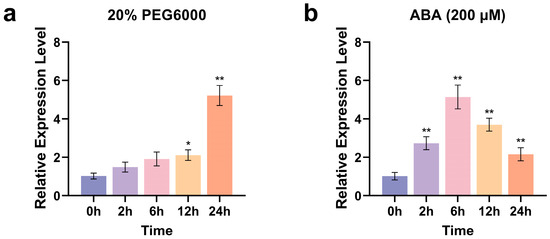
Figure 2.
Expression patterns of ZmCaM2 in response to 20% PEG6000 and 200 µM ABA treatments. (a,b) The relative expression levels were analyzed using the 2−ΔΔCT method. The 0 h time point was set as the control. Statistical significance compared with 0 h was determined using one-way ANOVA (* p < 0.05, ** p < 0.01). The data are presented as means ± standard deviation (SD). Bars indicate the standard error of the mean. The experiment was performed using three biological replicates from three independent plants.
2.3. ZmCaM2 Is Localized to the Nucleus and Plasma Membrane and Can Bind Ca2+
For investigating the subcellular localization of the ZmCaM2, the pCAMBIA1302-ZmCaM2-green fluorescent protein (GFP) (35S::ZmCaM2-GFP) recombinant vector was introduced into Nicotiana benthamiana leaves. The pCAMBIA1302-GFP (35S::GFP) vector was used as a control. As depicted in Figure 3b, the 35S::GFP protein was expressed in both the nucleus and the cell membrane. Similarly, 35S::ZmCaM2-GFP was localized to both the nucleus and the membrane.
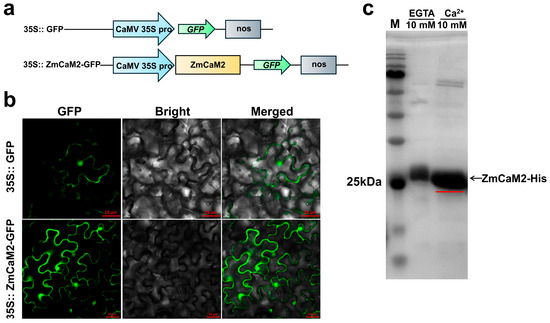
Figure 3.
Subcellular localization of ZmCaM2 and Ca2+ binding properties. (a) Schematic diagram of pCAMBIA1302-GFP (35S::GFP) and pCAMBIA1302-ZmCaM2-GFP (35S:: ZmCaM2-GFP); (b) Subcellular Localization of the 35S:: ZmCaM2-GFP recombinant protein. The pCAMBIA1302-GFP vector was utilized as a control. Scale bars = 25 µm; (c) Ca2+ binding assay of ZmCaM2-His recombinant protein. The red line indicates the migration rate of the ZmCaM2-His recombinant protein.
In order to verify whether ZmCaM2 has the ability to bind to Ca2+, the recombinant protein ZmCaM2-His was purified and then added to either 10 mM CaCl2 or 10 mM EGTA. As depicted in Figure 3c, the migration rate of the recombinant protein ZmCaM2-His was slower in the 10 mM EGTA solution compared to that in the Ca2+ solution. These results were consistent with the report by Garrigos, M. et al. (1991) [46], suggesting that ZmCaM2 is able to bind to Ca2+ in vitro.
2.4. ZmCaM2 Negatively Regulates Drought Tolerance in Maize
To validate the function of ZmCaM2 in maize, we generated ZmCaM2 overexpressing lines (OE1 and OE2) and CRISPR/Cas9-ZmCaM2 mutant lines (C1 and C2). The T3 generation overexpressing lines were confirmed by bar strips and qRT-PCR (Figure S1), while the T3 generation CRISPR/Cas9-ZmCaM2 mutants were verified by sequencing (Figure S2).
For drought tolerance evaluation, wild-type (WT) B104, overexpressing lines, and CRISPR/Cas9-ZmCaM2 mutant lines were subjected to drought stress by withholding water for 10 days (d), followed by re-watering for 3 d. Following treatment, the overexpressing lines exhibited more severe wilting than WT, whereas the mutant lines showed less wilting (Figure 4a). The dry weight of the OE1 and OE2 lines was significantly lower than that of the WT, C1, and C2 strains, whereas the dry weight of the C1 and C2 lines was significantly higher than that of the WT (Figure 4b). After re-watering, the survival rates of OE1 and OE2 were 31% and 33%, respectively. WT, C1, and C2 showed survival rates of 52%, 75%, and 76%, respectively (Figure 4c). These results indicate that ZmCaM2 negatively regulates drought tolerance in maize.
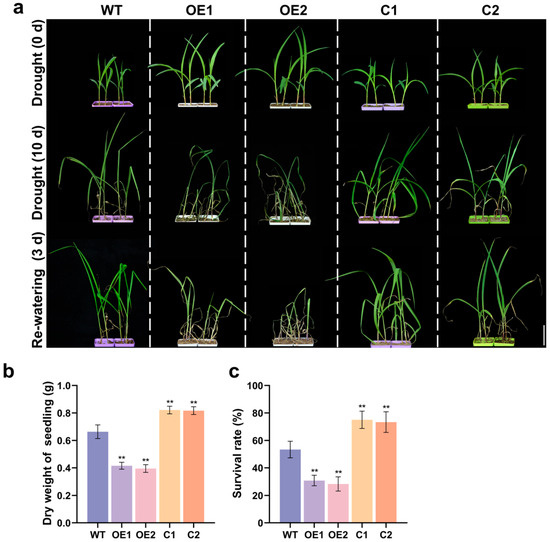
Figure 4.
(a) Images of drought tolerance evaluation in WT, ZmCaM2 overexpressing lines (OE1 and OE2), and CRISPR/Cas9-ZmCaM2 mutant lines (C1 and C2) under drought treatment. Plants were photographed at 0 d, after 10 d of drought treatment (withholding water), and after 3 d of rewatering. Bar = 5 cm; (b) After 10 d of drought stress, whole-plant dry weight was measured in WT, ZmCaM2 overexpressing, and CRISPR/Cas9-ZmCaM2 mutant lines; (c) Survival rates of WT, overexpressing lines, and mutant lines after 3 d of rewatering. Each biological replicate contained 20 plants, and the survival rate of each replicate was calculated as the mean of 20 plants. WT was used as the control. The data are presented as means ± standard deviation (SD). Bars indicate the standard error of the mean. Statistical significance was determined using one-way ANOVA (** p < 0.01). The photographs show representative plants.
2.5. ZmCaM2 Decreases Maize Drought Tolerance by Affecting Antioxidant Enzyme Activity
To further investigate the physiological role of ZmCaM2 under drought stress, the activities of SOD and POD, malondialdehyde (MDA) content, proline (Pro) content, ABA content, ROS content, and relative water content (RWC) were assessed in ZmCaM2 overexpressing lines, CRISPR/Cas9-ZmCaM2 mutant lines, and WT plants. Under normal conditions, no significant differences were observed in SOD and POD activity, MDA content, Pro content, ABA content, ROS content and RWC among the three genotypes (Figure 5a–g). After 10 d of drought treatment, SOD and POD activities in the OE1 and OE2 lines were lower than in the WT, whereas in the C1 and C2 lines, they were significantly higher than in the WT (Figure 5a,b). In the OE1 and OE2 lines, MDA and ROS content was significantly higher than in the WT lines. In contrast, MDA and ROS content in the C1 and C2 lines was significantly lower than in the WT lines (Figure 5c,f). As shown in Figure 5d,e, Pro and ABA content in the OE1 and OE2 lines was significantly lower than in the WT and C1 and C2 lines. Furthermore, Pro and ABA content in the C1 and C2 lines was significantly higher than in the WT lines. After drought treatment, RWC decreased across all maize lines. The OE1 and OE2 lines showed significantly lower RWC than the WT and C1 and C2 lines, whereas the C1 and C2 lines exhibited significantly higher RWC than the WT lines (Figure 5g). The level of relative water content serves as an indicator of a crop’s resistance to drought stress, with crops exhibiting higher relative water content demonstrating greater tolerance to such conditions. These findings suggest that ZmCaM2 negatively regulates maize tolerance to drought stress by increasing MDA and ROS accumulation, reducing antioxidant enzyme activity, lowering ABA content, and decreasing RWC.
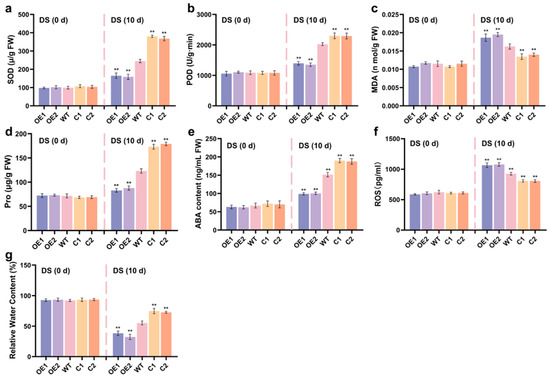
Figure 5.
The physiological indices of the ZmCaM2 overexpressing lines, CRISPR/Cas9-ZmCaM2 mutant lines, and WT were investigated under drought stress conditions. (a) SOD activity; (b) POD activity; (c) MDA content; (d) Pro content; (e) ABA content; (f) ROS content; (g) RWC. Each experiment was performed with three biological replicates. The data are presented as means ± standard deviation (SD). The significance analysis compared with WT was conducted using one-way analysis of variance (ANOVA) (** p < 0.01). WT is set as the control. Bars indicate the standard error of the mean. The experiment was performed using three biological replicates from three independent plants. DS: drought stress.
2.6. ZmCaM2 Negatively Regulates ABA Signal Transduction Pathway
To investigate the role of ZmCaM2 in ABA signaling, WT, ZmCaM2 overexpressing, and CRISPR/Cas9-ZmCaM2 mutant seedlings were treated with 0 µM ABA or 15 µM ABA. The primary root length was used to assess the sensitivity of the three strains to ABA. The primary root length was measured on the tenth day. The results indicate that, compared with WT lines, C1 and C2 lines showed shorter primary root length under ABA treatment, whereas OE1 and OE2 lines had longer primary root length (Figure 6a,b). To further validate the ZmCaM2 response to the ABA signaling pathway, we performed germination assays on WT seeds, ZmCaM2 overexpressing seeds, and CRISPR/Cas9-ZmCaM2 mutant seeds treated with 0 μM ABA or 10 μM ABA. The primary root length was measured on the fourth day. Results indicate that the C1 and C2 lines had significantly greater shorter primary root length than the WT lines under ABA treatment, while the OE1 and OE2 lines had significantly longer primary root length than the WT lines (Figure 6c,d). These results indicate that the overexpression of ZmCaM2 reduces maize sensitivity to ABA stress.
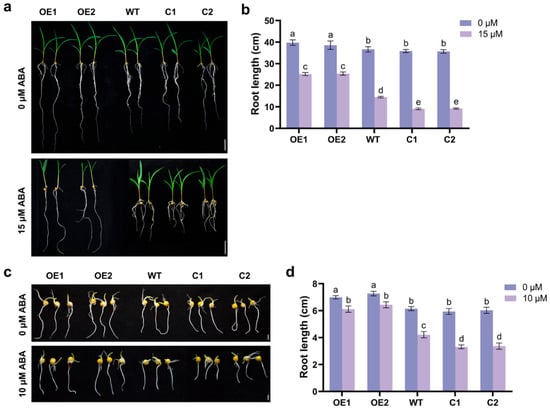
Figure 6.
Physiological and molecular responses of maize to ABA in ZmCaM2 lines (a) The phenotype of WT, ZmCaM2 overexpressing lines (OE1 and OE2), and CRISPR/Cas9-ZmCaM2 mutant lines (C1 and C2) grown in Hoagland solution with 0 µM ABA or 15 µM ABA. Scale bars = 5 cm. (b) The primary root length of WT, ZmCaM2 overexpressing lines, and CRISPR/Cas9-ZmCaM2 mutant lines were analyzed under 0 µM ABA or 15 µM ABA treatment. (c) Seed germination phenotypes of ZmCaM2 overexpressing lines, CRISPR/Cas9-ZmCaM2 mutant lines, and WT plants in 0 µM ABA or 10 µM ABA. Scale bar = 1 cm. (d) The primary root length of WT, ZmCaM2 overexpressing lines, and CRISPR/Cas9-ZmCaM2 mutant lines were analyzed under 0 µM ABA or 10 µM ABA treatment. Each experiment was performed with three biological replicates. The data are presented as means ± standard deviation (SD). The significance analysis was performed using two-way ANOVA (different lowercase letters indicate a difference at the 0.01 level p < 0.01). Bars indicate the standard error of the mean. The experiment was performed using three biological replicates from three independent plants.
Meanwhile, the expression levels of ABA-responsive genes (ZmRAB18 and ZmABF2) [47] were evaluated in the WT, ZmCaM2 overexpressing lines, and CRISPR/Cas9-ZmCaM2 mutant lines under 15 µM ABA treatment at 0 h and 6 h, respectively. The transcript levels of ZmRAB18 and ZmABF2 were not significantly different among the WT, OE1 and OE2, and the C1 and C2 lines under ABA treatment for 0 h (Figure 7a,b). However, the transcript levels of ZmRAB18 and ZmABF2 were significantly higher in the C1 and C2 lines compared to that of the WT after ABA treatment for 6 h, and the OE1 and OE2 lines exhibited opposite results (Figure 7a,b).
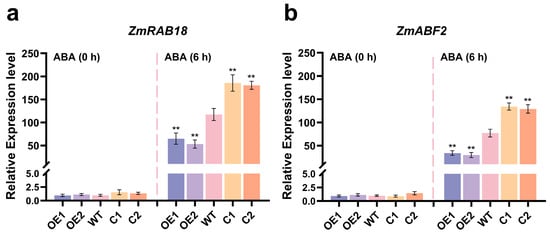
Figure 7.
The relative expression levels of ABA-responsive genes, including (a) ZmRAB18 (GRMZM2G098750) and (b) ZmABF2 (GRMZM2G479760), were analyzed in WT, ZmCaM2-overexpressing lines (OE1 and OE2) and CRISPR/Cas9-ZmCaM2 mutant lines (C1 and C2). (a,b) The expression levels of ABA-responsive genes ZmRAB18 (GRMZM2G098750) and ZmABF2 (GRMZM2G479760) under 15 µM ABA treatment at 0 h and 6 h, respectively. Expression level of WT at 0 h was set to 1.0. Relative expression levels were calculated using the 2−△△CT method. The data were the average (±SD) of three independent experiments. The significance analysis compared with WT was performed using one-way ANOVA (** p < 0.01). Bars indicate standard error of the mean.
3. Discussion
Drought constitutes a major abiotic stress in agriculture, often leading to decreased crop productivity or total crop failure, thereby posing substantial challenges to farmers’ livelihoods [48,49]. Exploring drought tolerance genes and understanding their functions are critical for improving maize drought tolerance. Notably, CaMs play essential roles in regulating physiological responses to drought stress in plants [48]. However, the function of CaMs in maize remains poorly understood. In this study, we isolated a CaM gene from maize, designated ZmCaM2. Evolutionary analysis revealed high homology between ZmCaM2 and OsCaM2, and sequence alignment showed that ZmCaM2 contains four conserved EF-hand domains (Figure 1). Moreover, ZmCaM2 specifically binds Ca2+ and is localized in the nucleus and membrane (Figure 3). Functional analysis indicates that overexpression of ZmCaM2 reduces maize tolerance to drought stress, whereas functional loss of ZmCaM2 enhances maize tolerance to drought stress (Figure 4). Further analysis indicated that ZmCaM2 negatively regulates drought tolerance by modulating multiple physiological parameters and the ABA signaling pathway (Figure 5–7). These results offer valuable insights into the role of ZmCaM2 in modulating maize drought tolerance via calcium ion binding and the ABA signaling pathway.
Numerous Ca2+ binding proteins have been demonstrated to be localized in the cytoplasm, vacuole, cell membrane or the nucleus [50]. In this study, we demonstrated that ZmCaM2 specifically binds to Ca2+ and localizes to both the nucleus and cell membrane (Figure 3). Therefore, ZmCaM2 may directly or indirectly regulate the expression of downstream target genes by binding calcium ions, thereby reducing antioxidant enzyme activity and ABA content while increasing ROS levels, thus diminishing maize tolerance to drought stress.
Several lines of evidence indicate that CaMs play critical roles in mediating responses to drought stress. For example, OsMSR2 positively regulates drought and salt tolerance [51]. Overexpression of TaCAM2-D in Arabidopsis enhances tolerance to both drought and salt stress [52]. Plants overexpressing EcCaM show enhanced tolerance to drought stress [44]. MtCaMP1 improves drought tolerance in plants by reducing the accumulation of H2O2 and MDA [53]. In contrast, ZmCaM2 negatively regulates drought tolerance in maize (Figure 4), suggesting that it functions as a negative regulator of the drought stress response.
ROS production is essential for protecting plants from various abiotic stresses; however, excessive ROS accumulation can lead to cellular damage [54]. Plants have evolved diverse enzymatic and non-enzymatic antioxidants to scavenge excess ROS and maintain ROS homeostasis [55]. Extensive studies have suggested that CaMs are involved in modulating ROS-related signal transduction [56]. Overexpression of MsCML46 enhances tolerance to freezing, drought, and salt stresses in tobacco by enhancing antioxidant enzyme activities, and reducing ROS accumulation [57]. Plants overexpressing ShCML44 show higher antioxidant enzyme activities and lower ROS levels, resulting in enhanced tolerance to abiotic stresses [58]. Following 10 d of drought stress treatment, ZmCaM2 overexpressing lines showed significantly lower SOD and POD activities, ABA and Pro contents, and RWC, while MDA and ROS content was significantly elevated. In contrast, the CRISPR/Cas9-ZmCaM2 mutant line exhibited enhanced SOD and POD activities, increased ABA and Pro contents, and reduced MDA and ROS accumulation (Figure 5). These results suggest that ZmCaM2 functions as a negative regulator of drought tolerance via modulation of ROS homeostasis.
The plant hormone ABA is a key signaling molecule that mediates plant responses to various stresses [59]. Many pieces of evidence demonstrated that Ca2+ sensors participate in ABA-mediated signaling pathway [40]. Overexpression of OsMSR2 in Arabidopsis heightened ABA sensitivity and conferred enhanced tolerance to both high salinity and drought stress [51]. AtCML24 was found to be involved in ABA signaling pathway [60]. AtCML42 mutants show enhanced tolerance to drought stress through ABA-dependent signaling pathway [61]. SlCML39 negatively regulates plant tolerance to high temperature stress in an ABA-dependent manner [62]. Under ABA treatment, the primary root length of ZmCaM2 overexpressing lines was significantly longer than that of the WT and CRISPR/Cas9-ZmCaM2 mutant lines, both at the seedling stage and during germination (Figure 6). Therefore, ZmCaM2 mediates the maize response to drought stress via the ABA signaling pathway. In contrast, our previous research demonstrated that homologous ZmCaM2-1 gene does not mediate any response via the ABA signaling pathway [63], suggesting that different members of the CaM gene family fulfill distinct roles in maize. Further exploration of the functions of CaM in maize is essential.
4. Materials and Methods
4.1. Plant Materials, Growing Conditions, and Treatments
The maize inbred lines B73 and B104 were provided by the Maize Breeding Innovation Team of Jilin Agricultural University. Seeds were sown in germination boxes and cultivated under controlled conditions at 28 °C with a 16 h light/8 h dark photoperiod until the V3 stage, with soil moisture maintained at 80% of field capacity. At the V3 stage, seedlings were subjected to drought and hormonal treatments in the cultivation trays, including irrigation with 20% PEG6000 until the soil reaches saturation, foliar spraying with 200 µM ABA and place the cultivation trays in a sealed incubator, or watering as a control. Leaf samples (one-quarter sections from equivalent regions showing uniform growth) were collected from three independent plants at 0, 2, 6, 12, and 24 h post-treatment and stored at −80 °C. The expression levels of ZmCaM2 were analyzed using qRT-PCR, and all primers used are listed in Table S1.
4.2. RNA Extraction and qRT-PCR
A total of 1 µg RNA was extracted using Trizol reagent (Tiangen, Beijing, China) and subsequently reverse-transcribed into cDNA using a TOYOBO reverse transcription kit (TOYOBO, Shanghai, China). qRT-PCR was performed on a QuantStudio 3 instrument (Thermo, Waltham, MA, USA), with ZmActin (GRMZM2G126010) serving as the internal control. Relative gene expression was calculated using the 2−ΔΔCT method [64]. Each experiment included three independent biological replicates. All primer sequences are listed in Table S1.
4.3. ZmCaM2 Cloning and Sequence Analysis
The full-length sequence of ZmCaM2 was cloned from leaves of the inbred maize line B73 using reverse transcription PCR (RT-PCR). Primer sequences are listed in Table S1. Homologous amino acid sequences of ZmCaM2 were identified by BLAST (v2.17.0) searches against the NCBI database, and phylogenetic analysis was performed using MEGA 7.0. Molecular weight (MW), pI, and GRAVY of ZmCaM2 were predicted using the ExPASy ProtParam tool (http://web.expasy.org/protparam/, accessed on 3 October 2025).
4.4. Subcellular Localization
The full-length CDS of ZmCaM2 was inserted into the pCAMBIA1302 vector in-frame with GFP using the Seamless Cloning Kit (Beyotime, Shanghai, China), with primer sequences provided in Table S1. Agrobacterium-mediated transformation was used to introduce either the pCAMBIA1302-GFP (35S::GFP) plasmid or the pCAMBIA1302-ZmCaM2-GFP (35S::ZmCaM2-GFP) construct into Nicotiana benthamiana plants. Following transformation, plants were maintained in the dark at 22 °C for 16–24 h. GFP fluorescence was visualized using a confocal laser scanning microscope (Leica, Frankfurt, Germany) with excitation at 488 nm.
4.5. Prokaryotic Expression Analysis of ZmCaM2 and Ca2+ Binding Assay
The CDS region of ZmCaM2 was cloned into the PET-29b vector via seamless cloning, with all primer sequences provided in Table S1. The recombinant plasmid was subsequently transformed into the E. coli BL21 (DE3) prokaryotic expression strain. Recombinant protein was purified using a His-tag Protein Purification Kit (LABLEAD, Beijing, China). For the Ca2+-binding assay, purified protein was incubated with either 10 mM CaCl2 or 10 mM EGTA.
4.6. Transgenic Plant Generation and Drought Tolerance Identification
The CDS of ZmCaM2, containing BamH I and Spe I restriction sites, was cloned into the pCAMBIA3301-UBI vector using T4 DNA ligase (TaKaRa, Beijing, China) to generate the pCAMBIA3301-UBI-ZmCaM2 recombinant plasmid. This plasmid was introduced into the inbred maize line B104 via Agrobacterium-mediated transformation, and T3 transgenic plants were identified by BAR strip assay and qRT-PCR.
The CRISPR/Cas9-ZmCaM2 mutant lines were generated through targeted knockout. A 20 bp sgRNA was inserted into the pegCas9PUB-B vector, which was subsequently introduced into B104 maize via Agrobacterium-mediated transformation. Homozygous mutant lines were confirmed by sequencing.
For drought tolerance evaluation, to ensure uniform soil moisture, we placed the WT B104, ZmCaM2 overexpression (OE1 and OE2), and CRISPR/Cas9-ZmCaM2 (C1 and C2) maize lines from the V3 stage into planting trays, maintaining soil moisture at 80% of field capacity. Subsequently, the V3 stage seedlings of ZmCaM2 overexpressing lines, CRISPR/Cas9-ZmCaM2 mutants, and WT lines were subjected to drought stress by withholding water for 10 d until the soil moisture reached 35% of field capacity. The seedlings from WT, ZmCaM2 overexpressing lines, and CRISPR/Cas9-ZmCaM2 mutant lines were selected, washed, and dried at 105 °C until they reached a constant weight, after which they were weighed. Subsequently, continue watering for 3 d to maintain soil moisture at 80%. Drought tolerance images of WT B104, ZmCaM2 overexpressing lines, and CRISPR/Cas9-ZmCaM2 lines were captured at the V3 stage, on days 10 of drought treatment and days 3 of rewatering with a Nikon D7000 camera (Nikon, Tokyo, Japan).
4.7. Physiological Indicators Measurement
Physiological indices of WT, ZmCaM2 overexpressing lines and CRISPR/Cas9-ZmCaM2 mutant lines were measured after 10 d of drought treatment. About 0.5 g of plant leaves were utilized for the determination of RWC, MDA, Pro, ABA and ROS content, and the activities of SOD and POD. RWC of leaves was measured according to the previously described method [65]. MDA content was determined using the thiobarbituric acid-based method [66]. Pro content was determined using the indophenol III colorimetric method [67]. The ABA content was determined using the plant abscisic acid ELISA detection kit (ELK, Wuhan, China), according to the manufacturer’s guidelines. Following the manufacturer’s instructions (KETE, Wenzhou, China), ROS were extracted using the Plant ROS ELISA Kit. ABA and ROS content were measured at 450 nm using a full-wavelength enzyme-labeling apparatus (HBS-ScanY, Shanghai, China). SOD activity was measured using the nitrotetrazolium blue chloride method [68]. POD activity was determined according to the previously described method [69]. The absorbance values of MDA, SOD, and POD were measured using a spectrophotometer (INESA, Shanghai, China). The experiment was performed using three biological replicates from three independent plants.
4.8. ABA Stress Treatment
The seeds of WT, ZmCaM2 overexpressing lines (OE1 and OE2), and CRISPR/Cas9-ZmCaM2 mutant lines (C1 and C2) were sterilized and placed on wet filter paper at 28 °C for 3 d until the taproot reached about 3 cm. The seedlings were transferred into Hoagland solution containing 0 µM or 15 µM ABA for 10 d, and the primary root length was measured and calculated. The experiment was performed with three independent biological repetitions. The three-leaf stage seedlings were treated with 15 µM ABA, and the samples were collected at 0 h and 6 h, respectively.
For the seed germination experiment under ABA stress, we disinfected seeds from the WT, ZmCaM2 overexpressing lines, and CRISPR/Cas9-ZmCaM2 mutant lines, followed by treatment with 0 μM or 10 μM ABA for 4 d, and the primary root length was measured and calculated.
4.9. Statistical Analysis
The statistical experiments were performed with three biological replicates. All data were analyzed using GraphPad Prism 9.0 software. The significant differences between samples were analyzed using one-way ANOVA (* p < 0.05, ** p < 0.01) and bars indicate the standard error of the mean.
5. Conclusions
In our work, we focused on drought tolerance at the seedling stage and found that ZmCaM2 may reduce the tolerance of maize to drought stress by binding Ca2+ and relying on the ABA signaling pathway to suppress antioxidant enzyme activity, decrease RWC and ABA content, and increase ROS accumulation (Figure 8). However, its effects on agronomic traits and yield under field drought conditions, as well as its practical value in breeding, require long-term evaluation in larger populations. To mitigate yield losses under climate change, assessing the potential application of ZmCaM2 in drought- and stress-resilient breeding will remain a major focus of our future research. This line of work has already been incorporated into our genome-assisted breeding strategy.
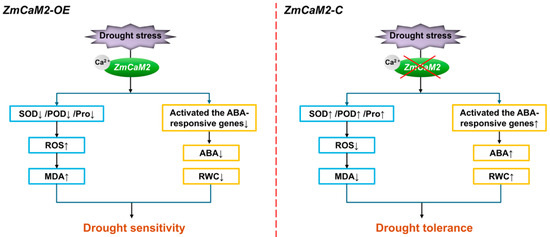
Figure 8.
A proposed model of ZmCaM2 in response to drought stress.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants14233656/s1, Figure S1: Detection of transgenic maize; Figure S2: Detection of mutant maize; Table S1: RT-qPCR primers used in this study.
Author Contributions
Conceptualization, L.J. and Z.Z.; Methodology, L.J. and Z.Z.; Validation, M.L., P.B., H.W. and Z.W. (Zhiqiang Wu); Investigation, M.L., P.B., H.W. and Z.W. (Zhiqiang Wu), X.X.; Resources, L.J., Z.Z., W.Y., J.C. and X.R.; Data curation, M.L., P.B., H.W. and Z.W. (Zhen Wang); Writing—original draft preparation, M.L., P.B., Z.W. (Zhiqiang Wu) and H.W.; Writing—review and editing, L.J., Z.Z., M.L. and P.B.; Visualization, M.L., P.B., H.W. and Z.W. (Zhiqiang Wu).; Supervision, L.J., Z.Z., W.Y., J.C. and X.R.; Project administration, L.J. and Z.Z.; Funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Jilin Provincial Scientific and Technological Development Program (20240602040RC).
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its Supplementary Materials files.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| ABA | Abscisic acid |
| ROS | Reactive oxygen species |
| Ca2+ | Calcium ions |
| CaM | Calmodulin |
| CMLs | Calmodulin-like proteins |
| CDPKs | Calcium-dependent protein kinases |
| CBLs | Calcineurin B-like proteins |
| CCaMKs | Calcium and calmodulin -dependent protein kinases |
| GFP | Green fluorescent protein |
| WT | Wild type |
| h | Hour |
| d | Day |
| V3 | Third-leaf stage |
| CDS | Coding sequence |
| RWC | Relative water content |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| MDA | Malondialdehyde |
| Pro | Proline |
| PI | Isoelectric point |
| GRAVY | Grand Average of Hy-dropathicity |
| MW | Molecular weight |
| RT-PCR | Reverse transcription PCR |
References
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Qin, F. The battle of crops against drought: Genetic dissection and improvement. J. Integr. Plant Biol. 2023, 65, 496–525. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global Synthesis of Drought Effects on Maize and Wheat Production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef]
- Yadava, P.; Abhishek, A.; Singh, R.; Singh, I.; Kaul, T.; Pattanayak, A.; Agrawal, P.K. Advances in Maize Transformation Technologies and Development of Transgenic Maize. Front. Plant Sci. 2017, 7, 1949. [Google Scholar] [CrossRef]
- Sheoran, S.; Kaur, Y.; Kumar, S.; Shukla, S.; Rakshit, S.; Kumar, R. Recent Advances for Drought Stress Tolerance in Maize (Zea mays L.): Present Status and Future Prospects. Front. Plant Sci. 2022, 13, 872566. [Google Scholar] [CrossRef]
- Yuan, P.; Du, L.; Poovaiah, B.W. Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity. Int. J. Mol. Sci. 2018, 19, 1764. [Google Scholar] [CrossRef] [PubMed]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.P. Calcium signalling in plant biotic interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Shariq Iqbal, M.; Singh, S.P.; Buaboocha, T. Ca2+/Calmodulin Complex Triggers CAMTA Transcriptional Machinery Under Stress in Plants: Signaling Cascade and Molecular Regulation. Front. Plant Sci. 2020, 11, 598327. [Google Scholar] [CrossRef]
- Hamel, L.P.; Sheen, J.; Séguin, A. Ancient signals: Comparative genomics of green plant CDPKs. Trends Plant Sci. 2014, 19, 79–89. [Google Scholar] [CrossRef]
- Wang, J.P.; Munyampundu, J.P.; Xu, Y.P.; Cai, X.Z. Phylogeny of Plant Calcium and Calmodulin-Dependent Protein Kinases (CCaMKs) and Functional Analyses of Tomato CCaMK in Disease Resistance. Front. Plant Sci. 2015, 6, 1075. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Mahiwal, S.; Nambiar, D.M.; Pandey, G.K. CBL-CIPK module-mediated phosphoregulation: Facts and hypothesis. Biochem. J. 2020, 477, 853–871. [Google Scholar] [CrossRef]
- Reddy, A.S.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with stresses: Roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium Sensors as Key Hubs in Plant Responses to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [PubMed]
- McCormack, E.; Tsai, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef]
- Zhu, X.; Dunand, C.; Snedden, W.; Galaud, J.P. CaM and CML emergence in the green lineage. Trends. Plant Sci. 2015, 20, 483–489. [Google Scholar] [CrossRef]
- Boonburapong, B.; Buaboocha, T. Genome-wide identification and analyses of the rice calmodulin and related potential calcium sensor proteins. BMC Plant Biol. 2007, 7, 4. [Google Scholar] [CrossRef][Green Version]
- He, X.; Liu, W.; Li, W.; Liu, Y.; Wang, W.; Xie, P.; Kang, Y.; Liao, L.; Qian, L.; Liu, Z.; et al. Genome-wide identification and expression analysis of CaM/CML genes in Brassica napus under abiotic stress. J. Plant Physiol. 2020, 255, 153251. [Google Scholar] [CrossRef]
- Fu, M.; Wu, C.; Li, X.; Ding, X.; Guo, F. Genome-Wide Identification and Expression Analysis of CsCaM/CML Gene Family in Response to Low-Temperature and Salt Stresses in Chrysanthemum seticuspe. Plants 2022, 11, 1760. [Google Scholar] [CrossRef]
- Cai, K.; Kuang, L.; Yue, W.; Xie, S.; Xia, X.; Zhang, G.; Wang, J. Calmodulin and calmodulin-like gene family in barley: Identification, characterization and expression analyses. Front. Plant Sci. 2022, 13, 964888. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, W.; Liu, L.; Su, Y.; Li, Y.; Jia, W.; Jiao, B.; Wang, J.; Yang, F.; Dong, F.; et al. Genome-wide identification and expression analysis of calmodulin and calmodulin-like genes in wheat (Triticum aestivum L.). Plant Signal Behav. 2022, 17, 2013646. [Google Scholar] [CrossRef] [PubMed]
- Vandelle, E.; Vannozzi, A.; Wong, D.; Danzi, D.; Digby, A.M.; Dal Santo, S.; Astegno, A. Identification, characterization, and expression analysis of calmodulin and calmodulin-like genes in grapevine (Vitis vinifera) reveal likely roles in stress responses. Plant Physiol. Biochem. 2018, 129, 221–237. [Google Scholar] [CrossRef]
- Kim, M.C.; Chung, W.S.; Yun, D.J.; Cho, M.J. Calcium and calmodulin-mediated regulation of gene expression in plants. Mol. Plant 2009, 2, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.H.; Park, C.Y.; Kim, J.C.; Heo, W.D.; Cheong, M.S.; Park, H.C.; Kim, M.C.; Moon, B.C.; Choi, M.S.; Kang, Y.H.; et al. Direct interaction of a divergent CaM isoform and the transcription factor, MYB2, enhances salt tolerance in arabidopsis. J. Biol. Chem. 2005, 280, 3697–3706. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.; El-Habbak, M.H.; Havens, W.M.; Singh, A.; Zheng, D.; Vaughn, L.; Haudenshield, J.S.; Hartman, G.L.; Korban, S.S.; Ghabrial, S.A. Overexpression of GmCaM4 in soybean enhances resistance to pathogens and tolerance to salt stress. Mol. Plant Pathol. 2014, 15, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Saeng-ngam, S.; Takpirom, W.; Buaboocha, T.; Chadchawan, S. The role of the OsCam1-1 salt stress sensor in ABA accumulation and salt tolerance in rice. J. Plant Biol. 2012, 55, 198–208. [Google Scholar] [CrossRef]
- Wu, H.C.; Luo, D.L.; Vignols, F.; Jinn, T.L. Heat shock-induced biphasic Ca(2+) signature and OsCaM1-1 nuclear localization mediate downstream signalling in acquisition of thermotolerance in rice (Oryza sativa L.). Plant Cell Environ. 2012, 35, 1543–1557. [Google Scholar] [CrossRef]
- Yang, J.; Ji, L.; Liu, S.; Jing, P.; Hu, J.; Jin, D.; Wang, L.; Xie, G. The CaM1-associated CCaMK-MKK1/6 cascade positively affects lateral root growth via auxin signaling under salt stress in rice. J. Exp. Bot. 2021, 72, 6611–6627. [Google Scholar] [CrossRef]
- Zhou, S.; Jia, L.; Chu, H.; Wu, D.; Peng, X.; Liu, X.; Zhang, J.; Zhao, J.; Chen, K.; Zhao, L. Arabidopsis CaM1 and CaM4 Promote Nitric Oxide Production and Salt Resistance by Inhibiting S-Nitrosoglutathione Reductase via Direct Binding. PLoS Genet. 2016, 12, e1006255. [Google Scholar] [CrossRef]
- Dai, C.; Lee, Y.; Lee, I.C.; Nam, H.G.; Kwak, J.M. Calmodulin 1 Regulates Senescence and ABA Response in Arabidopsis. Front. Plant Sci. 2018, 9, 803. [Google Scholar] [CrossRef]
- Takahashi, F.; Mizoguchi, T.; Yoshida, R.; Ichimura, K.; Shinozaki, K. Calmodulin-dependent activation of MAP kinase for ROS homeostasis in Arabidopsis. Mol. Cell 2011, 41, 649–660. [Google Scholar] [CrossRef]
- Chu, M.; Li, J.; Zhang, J.; Shen, S.; Li, C.; Gao, Y.; Zhang, S. AtCaM4 interacts with a Sec14-like protein, PATL1, to regulate freezing tolerance in Arabidopsis in a CBF-independent manner. J. Exp. Bot. 2018, 69, 5241–5253. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.; Zhou, S.; Wang, L.; Cheng, Y.; Zhao, L. Nitric oxide functions as a signal and acts upstream of AtCaM3 in thermotolerance in Arabidopsis seedlings. Plant Physiol. 2010, 153, 1895–1906. [Google Scholar] [CrossRef]
- Yang, N.; Peng, C.; Cheng, D.; Huang, Q.; Xu, G.; Gao, F.; Chen, L. The over-expression of calmodulin from Antarctic notothenioid fish increases cold tolerance in tobacco. Gene 2013, 521, 32–37. [Google Scholar] [CrossRef]
- Del Cerro, P.; Cook, N.M.; Huisman, R.; Dangeville, P.; Grubb, L.E.; Marchal, C.; Ho Ching Lam, A.; Charpentier, M. Engineered CaM2 modulates nuclear calcium oscillation and enhances legume root nodule symbiosis. Proc. Natl. Acad. Sci. USA 2022, 119, e2200099119. [Google Scholar] [CrossRef]
- Yu, B.; Yan, S.; Zhou, H.; Dong, R.; Lei, J.; Chen, C.; Cao, B. Overexpression of CsCaM3 improves high temperature tolerance in cucumber. Front. Plant Sci. 2018, 9, 797. [Google Scholar] [CrossRef]
- Raina, M.; Kumar, A.; Yadav, N.; Kumari, S.; Yusuf, M.A.; Mustafiz, A.; Kumar, D. StCaM2, a calcium binding protein, alleviates negative effects of salinity and drought stress in tobacco. Plant Mol. Biol. 2021, 106, 85–108. [Google Scholar] [CrossRef]
- Shen, Q.; Fu, L.; Su, T.; Ye, L.; Huang, L.; Kuang, L.; Wu, L.; Wu, D.; Zhang, G. Calmodulin HvCaM1 negatively regulates salt tolerance via modulation of HvHKT1s and HvCAMTA4. Plant Physiol. 2020, 183, 1650–1662. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in integrating plant responses to drought and salt stresses. Field Crops Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Khan, N.A. Minimising toxicity of cadmium in plants—Role of plant growth regulators. Protoplasma 2015, 252, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Vanderbeld, B.; Snedden, W.A. Developmental and stimulus-induced expression patterns of Arabidopsis calmodulin-like genes CML37, CML38 and CML39. Plant Mol. Biol. 2007, 64, 683–697. [Google Scholar] [CrossRef]
- Magnan, F.; Ranty, B.; Charpenteau, M.; Sotta, B.; Galaud, J.P.; Aldon, D. Mutations in AtCML9, a calmodulin-like protein from Arabidopsis thaliana, alter plant responses to abiotic stress and abscisic acid. Plant J. 2008, 56, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Qiao, Z.; Liu, H.; Acharya, B.R.; Li, C.; Zhang, W. CML20, an Arabidopsis Calmodulin-like Protein, Negatively Regulates Guard Cell ABA Signaling and Drought Stress Tolerance. Front. Plant Sci. 2017, 8, 824. [Google Scholar] [CrossRef] [PubMed]
- Jamra, G.; Agarwal, A.; Singh, N.; Sanyal, S.K.; Kumar, A.; Pandey, G.K. Ectopic expression of finger millet calmodulin confers drought and salinity tolerance in Arabidopsis thaliana. Plant Cell Rep. 2021, 40, 2205–2223. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, L.; Li, J.; Yang, W.; Ci, J.; Ren, X.; Wang, W.; Wang, Y.; Jiang, L.; Yang, W. Identification and expression analysis revealed drought stress-responsive Calmodulin and Calmodulin-like genes in maize. J. Plant Interact. 2022, 17, 450–461. [Google Scholar] [CrossRef]
- Garrigos, M.; Deschamps, S.; Viel, A.; Lund, S.; Champeil, P.; Møller, J.V.; le Maire, M. Detection of Ca(2+)-binding proteins by electrophoretic migration in the presence of Ca2+ combined with 45Ca2+ overlay of protein blots. Anal. Biochem. 1991, 194, 82–88. [Google Scholar] [CrossRef]
- Yan, Z.; Zhang, F.; Mu, C.; Ma, C.; Yao, G.; Sun, Y.; Hou, J.; Leng, B.; Liu, X. The ZmbHLH47-ZmSnRK2.9 Module Promotes Drought Tolerance in Maize. Int. J. Mol. Sci. 2024, 25, 4957. [Google Scholar] [CrossRef]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Perochon, A.; Aldon, D.; Galaud, J.P.; Ranty, B. Calmodulin and calmodulin-like proteins in plant calcium signaling. Biochimie 2011, 93, 2048–2053. [Google Scholar] [CrossRef]
- Xu, G.Y.; Rocha, P.S.; Wang, M.L.; Xu, M.L.; Cui, Y.C.; Li, L.Y.; Zhu, Y.X.; Xia, X. A novel rice calmodulin-like gene, OsMSR2, enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis. Planta 2011, 234, 47–59. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Dong, F.; Zou, J.; Gao, C.; Zhu, Z.; Liu, Y. Multiple roles of wheat calmodulin genes during stress treatment and TaCAM2-D as a positive regulator in response to drought and salt tolerance. Int. J. Biol. Macromol. 2022, 220, 985–997. [Google Scholar] [CrossRef]
- Wang, T.Z.; Zhang, J.L.; Tian, Q.Y.; Zhao, M.G.; Zhang, W.H. A Medicago truncatula EF-hand family gene, MtCaMP1, is involved in drought and salt stress tolerance. PLoS ONE 2013, 8, e58952. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Li, S.; Liu, S.; Zhang, Q.; Cui, M.; Zhao, M.; Li, N.; Wang, S.; Wu, R.; Zhang, L.; Cao, Y.; et al. The interaction of ABA and ROS in plant growth and stress resistances. Front. Plant Sci. 2022, 13, 1050132. [Google Scholar] [CrossRef]
- Sierla, M.; Waszczak, C.; Vahisalu, T.; Kangasjärvi, J. Reactive oxygen species in the regulation of stomatal movements. Plant Physiol. 2016, 171, 1569–1580. [Google Scholar] [CrossRef]
- Du, B.; Chen, N.; Song, L.; Wang, D.; Cai, H.; Yao, L.; Li, X.; Guo, C. Alfalfa (Medicago sativa L.) MsCML46 gene encoding calmodulin-like protein confers tolerance to abiotic stress in tobacco. Plant Cell Rep. 2021, 40, 1907–1922. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Liu, H.; Xing, Y.; Hussain, S.; Ouyang, B.; Zhang, Y.; Li, H.; Ye, Z. Overexpression of calmodulin-like (ShCML44) stress-responsive gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses. Sci. Rep. 2016, 6, 31772. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef]
- Delk, N.A.; Johnson, K.A.; Chowdhury, N.I.; Braam, J. CML24, regulated in expression by diverse stimuli, encodes a potential Ca2+ sensor that functions in responses to abscisic acid, daylength, and ion stress. Plant Physiol. 2005, 139, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Vadassery, J.; Reichelt, M.; Hause, B.; Gershenzon, J.; Boland, W.; Mithöfer, A. CML42-mediated calcium signaling coordinates responses to Spodoptera herbivory and abiotic stresses in Arabidopsis. Plant Physiol. 2012, 159, 1159–1175. [Google Scholar] [CrossRef]
- Ding, H.; Qian, Y.; Fang, Y.; Ji, Y.; Sheng, J.; Ge, C. Characteristics of SlCML39, a Tomato Calmodulin-like Gene, and Its Negative Role in High Temperature Tolerance of Arabidopsis thaliana during Germination and Seedling Growth. Int. J. Mol. Sci. 2021, 22, 11479. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, M.; Wang, H.; Li, M.; Liu, X.; Zang, Z.; Jiang, L. ZmCaM2-1, a Calmodulin Gene, Negatively Regulates Drought Tolerance in Transgenic Arabidopsis Through the ABA-Independent Pathway. Int. J. Mol. Sci. 2025, 26, 2156. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Smart, R.E. Rapid estimates of relative water content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).