Abstract
High nature value (HNV) grasslands in mountain areas are important ecosystems for biodiversity maintenance and offer a multitude of ecosystem services, but they are constantly threatened by abandonment or intensive fertilization. The aim of this study was to assess the effects of organic and mineral fertilization, under mulching and abandonment scenarios, on the floristic composition and diversity of Nardus stricta-dominated grasslands located in the North-Eastern Carpathians (Romania). The field experiment included 11 variants (control, low, moderate, and high inputs), analyzed as communities with cluster, ordinations, indicator species, and α indices. The results showed a clear separation of communities along the input gradient, from the oligotrophic grassland dominated by Nardus stricta (control variant) to mesotrophic/eutrophic communities dominated by Dactylis glomerata, Festuca pratensis, and Trifolium pratense at moderate and high inputs. Moderate fertilization (10–20 t ha−1 manure; N50P50K50–N100P100K100) maximized species richness (37–38 species), Shannon diversity (H′ = 2.5–2.6), and evenness (E = 0.70–0.75). High inputs reduced diversity and favored competitive grasses. Indicator species analysis highlighted a multitude of species that show the plant communities’ response to adaptive management. Moderate fertilization provides a viable trade-off between productivity and biodiversity, while abandonment or overfertilization accelerates biodiversity loss.
1. Introduction
High nature value (HNV) grasslands are mountain ecosystems of strategic importance for biodiversity conservation, providing ecosystem services and maintaining traditional cultural landscapes in Europe [1,2]. They provide essential support, supply, regulation, and cultural services, contributing to carbon sequestration, water retention, fire risk reduction, and the maintenance of natural and cultural heritage [3]. These semi-natural agricultural systems were formed and maintained through extensive management, especially mowing and grazing, and are characterized by high floristic diversity and the presence of indicator species adapted to oligotrophic and stressful conditions [4,5,6]. In the establishment of mountain grasslands management, the soil–plant relationship plays an essential role, with indicator species providing rapid information on the agrochemical conditions of the soil and on the state of fertility [7,8].
The classification and assessment of HNV grasslands is an essential basis for choosing adapted management techniques and for understanding their role in cultural landscapes [9]. The pressures are exacerbated by difficulties in implementing EU conservation policies, due to limited collaboration between environmental and agricultural institutions, leading to an unfavorable conservation status for species-rich semi-natural habitats [10]. Moderate fertilization can stimulate species productivity and coexistence, reducing competitive dominance—a mechanism similar to that suggested by the intermediate disturbance hypothesis [11,12,13,14].
In the last decades, changes in agricultural practices—the intensification of mineral fertilization or the abandonment of traditional management—have led to major transformations in the structure and functioning of mountain grasslands [15,16,17,18]. The abandonment determines secondary successions and major structural changes, with the degradation of agronomic and ecological value [19]. Different studies propose alternative technologies that are economically viable in mountainous areas, such as mulching combined with organic fertilization, which maintain oligotrophic grasslands at an appropriate level of diversity, with minimal changes in vegetation composition [20,21,22]. Several conceptual approaches propose the segmentation of microbial functional niche to assess alterations caused by fertilization, extracting sensitive and stable guilds, which support sustainable management policies in HNV grasslands [23,24]. Another approach can analyze the nitrogen use efficiency (NUE), which is used in the design of sustainable temporary grassland systems, as an indicator for optimizing fertilization and reducing nutrient losses [25].
Romania has some of the largest areas of HNV grasslands in the European Union, especially in the Western and Southern Carpathians, where they amount to over 2 million hectares [6,26,27]. These ecosystems are valued both for their floristic biodiversity and for their cultural and economic value [28,29]. Recent studies in the Apuseni Natural Park (Romania) show that permanent grasslands in habitat 6520 have an average productivity of 5.2 t ha−1 green mass and a pastoral value of 35 [30], values that highlight the importance of these ecosystems for maintaining biodiversity and providing fodder resources. Although HNV grasslands support medicinal plants, such as Arnica montana, which provide high economic and cultural value, they are constantly threatened by intensive agricultural practices, requiring sustainable conservation strategies [31]. However, maintaining the balance between productivity, biodiversity conservation, and sustainable resource use remains a major challenge for current and future agricultural and environmental policies [32,33]. In this sense, European agricultural policies promote agri-environmental schemes and compensatory measures that support biodiversity, but abandonment and intensification continue to pose significant threats [6,34,35].
In the Apuseni Mountains (Romania), population exodus has accentuated the abandonment of grasslands, leading to the transformation of traditional cultural landscapes and the reduction in biodiversity [36], which highlights the need to restore traditional agricultural practices [37]. Long-term studies conducted in this area have demonstrated that constant application of manure has a significant impact on the floristic composition and productivity of Festuca rubra grasslands [38,39]. In contrast, intensive fertilization leads to the simplification of phytocoenoses and the dominance of competitive grasses and perennial legumes, while the lack of input favors oligotrophic species and reduces forage value [40,41,42,43]. Furthermore, long-term fertilization reorients mycorrhizal colonization strategies in the roots of dominant species (e.g., Agrostis capillaris), favoring storage structures over transfer ones, which may affect nutrient cycling in HNV ecosystems [44]. The assessment of indicator species for soil agrochemical characterization has also been used in several studies conducted in Apuseni Natural Park [7], providing a useful framework for monitoring and adaptive management of type 6520 habitats. These results can inform conservation policies, where, according to comparative studies, institutional collaboration between conservation and agriculture is essential for maintaining the favorable status of species-rich grasslands, integrating socio-cultural aspects [45].
In this context, the objective of the study was to evaluate the solidity of the hypothesis that different management scenarios will affect the stability of the plant community in a Nardus stricta HNV grassland. For this, a multiple-treatment experiment was designed to analyze the effects of organic and mineral fertilization, combined with mulching and abandonment, on the diversity and structure of species. A complex system of data analysis was carried out to identify the most suitable test that answers one or several research questions. (a) Is the application of combined management scenarios visible in the plant communities? (b) Are diversity indices sensitive enough to show the changes due to applied management? (c) Is there a visible shift from an oligotrophic status to a meso- or eutrophicated one? By applying multivariate methods and indicator species analysis, the research aims to highlight the mechanisms by which the intensity and type of input influence the dynamics of mountain phytocoenoses and to provide recommendations for an adaptive and sustainable management of these ecosystems.
2. Results
2.1. The Influence of the Management Scenario on the Grassland Community
The cluster analysis highlighted the vegetation classification and clearly reflected the ecological gradient induced by the different management practices and fertilization rates (Figure 1). Four distinct clusters were identified: Group 1—oligotrophic grasslands dominated by Nardus stricta, typical of nutrient-poor conditions and extensive management; Group 2—transitional communities of Nardus stricta–Festuca rubra, reflecting a moderate shift in floristic composition at slightly increased inputs; Group 3—Festuca rubra–Nardus stricta communities co-dominant with Agrostis capillaris; and Group 4—mesotrophic grasslands of Agrostis capillaris–Festuca rubra, associated with high fertilization levels. The formation of these clusters demonstrates that fertilizers produce major changes in the floristic structure of HNV grasslands, separating two large ecological groups: one corresponding to treatments with low or absent input (T1–T3) and another associated with intensive organic or mineral fertilization (T4–T11). Each management regime thus generated a specific type of grassland, highlighting the close relationship between the level of inputs and the composition of the communities.
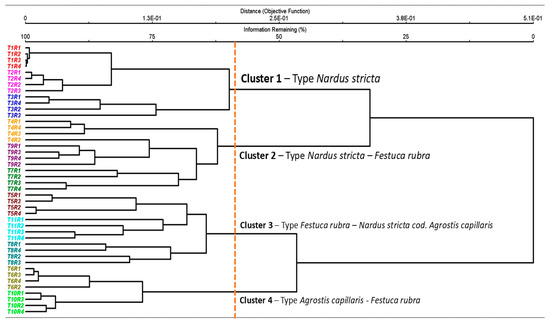
Figure 1.
Floristic classification of the vegetation and the changes of grassland type based on cluster analysis. Legend: T1—unfertilized control; T2—abandonment; T3—mulching; T4—50 kg N ha−1, 50 kg P2O5 ha−1, and 50 kg K2O ha−1 mineral fertilization; T5—100 kg N ha−1, 100 kg P2O5 ha−1, and 100 kg K2O ha−1 mineral fertilization; T6—150 kg N ha−1, 150 kg P2O5 ha−1, and 150 kg K2O ha−1 mineral fertilization; T7—10 t ha−1 cattle manure applied annually; T8—20 t ha−1 cattle manure applied annually; T9—20 t ha−1 cattle manure applied every two years; T10—30 t ha−1 cattle manure applied annually; T11—30 t ha−1 cattle manure applied every two years; R1–R4—replications. The dendrogram cutoff line was set at ~60% remaining information, resulting in four ecologically interpretable clusters.
2.2. Plant Community Patterns Explored Through Principal Coordinates Analysis (PCoA)
To more clearly visualize the relationships between treatments and floristic communities, we used PCoA analysis, which highlighted the distribution of phytocoenoses along the trophic gradient generated by fertilization, which is represented by Axis 1 (Figure 2, Table 1).
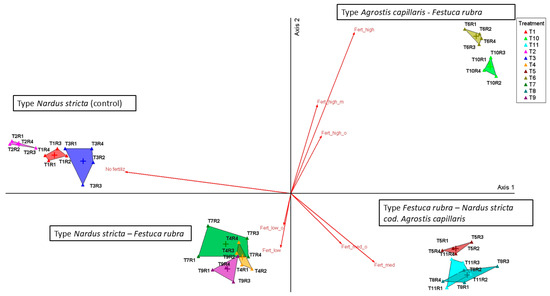
Figure 2.
Principal Coordinates Analysis (PCoA) of grassland communities under different fertilization regimes. Legend: T1—unfertilized control (martor); T2—abandonment; T3—mulching; T4—50 kg N ha−1, 50 kg P2O5 ha−1, and 50 kg K2O ha−1 mineral fertilization; T5—100 kg N ha−1, 100 kg P2O5 ha−1, and 100 kg K2O ha−1 mineral fertilization; T6—150 kg N ha−1, 150 kg P2O5 ha−1, and 150 kg K2O ha−1 mineral fertilization; T7—10 t ha−1 cattle manure applied annually; T8—20 t ha−1 cattle manure applied annually; T9—20 t ha−1 cattle manure applied every two years; T10—30 t ha−1 cattle manure applied annually; T11—30 t ha−1 cattle manure applied every two years; R1–R4—replications. Vectors indicate the fertilization gradient: zero input (no fertilizers applied: T1–T3), low input (reduced doses: T4, T7), medium input (moderate doses: T5, T8, T9, T11), and high input (intensive fertilization: T6, T10), with separation between organic (o) and mineral (m) treatments.

Table 1.
Correlation of experimental factors with the ordination axes (PCoA).
On this axis, treatments without fertilization (T1–T3) were negatively correlated (r = −0.825, p < 0.001) and clustered on the left side of the ordinate, while treatments with medium and high inputs (T5–T11) were positively correlated and clustered on the right side. Axis 2 explained only 11.1% of the variation and had low biological relevance. It partially separated variants with medium input from those with high input. Overall, PCoA confirms that fertilization intensity, and not type (organic vs. mineral), explains most of the variation in floristic composition.
The vectors corresponding to the type of fertilizer (organic vs. mineral) had close values and similar directions, indicating the lack of significant differences between the two types of input at the same intensity level. Thus, the variability of the floristic composition is mainly explained by the intensity of fertilization and not by its type.
The Multi-Response Permutation Procedure (MRPP) analysis was applied to test the robustness of group separation based on floristic analysis and to quantify the distance between the identified grassland types. The groups without input (T1–T3) were significantly separated from all fertilized treatments (p < 0.01), but the differences between treatments with close doses, either organic or mineral, were not statistically significant (Table 2). The greatest difference was observed between the zero-input group (T1–T3) and the high-input group (T6, T10). Intermediate values of the A index were obtained for the zero-input vs. medium-input and low-input vs. high-input comparisons. The lowest values were recorded between medium input and high input, suggesting a partial convergence of the communities under high fertilization.
Table 2.
MRPP pairwise comparisons of floristic composition between the four grassland groups.
2.3. The Comparative Analysis of Community Composition and Species Projection Along Fertilization Gradient
To visualize how these correlations are reflected in the community structure, the distribution of species and treatments was graphically represented by PCoA analysis, where the vectors indicate the direction and amplitude of the species response to the fertilization gradient (Figure 3). These differences confirm the clear separation between the grassland groups previously identified by cluster analysis and MRPP tests. The PCoA showed a clear separation between the treatments with low inputs (T1–T3) and those with medium and high levels of mineral and organic fertilization (T4–T11), arranged along Axis 1. This axis explained most of the variation (87.5%) and represented the main trophic gradient. On the negative side of Axis 1 are phytocenoses dominated by Nardus stricta, Vaccinium myrtillus, and Luzula multiflora, indicating oligotrophic grasslands characteristic of treatments without inputs. On the positive side, medium and high fertilization caused a shift of communities towards competitive mesotrophic species, such as Dactylis glomerata, Festuca pratensis, and Phleum pratense. Axis 2, although explaining only 11.1% of the variation, highlighted the differentiation of leguminous species (Trifolium pratense, Trifolium repens) associated with moderate organic inputs. This distribution confirms the trends highlighted by the cluster and MRPP, suggesting that the application of inputs not only changes the dominance of species but also the typology of phytocenoses in the long term.
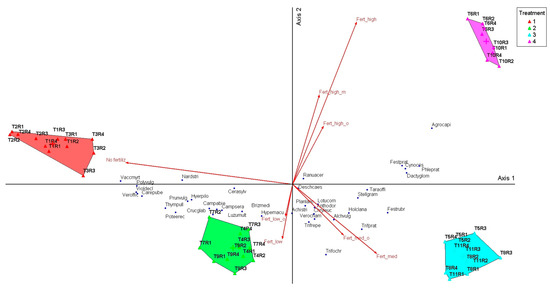
Figure 3.
Principal Coordinates Analysis (PCoA) of floristic composition in relation to intensity of treatments. Legend: No fertiliz—no fertilization; T1—unfertilized control; T2—abandonment; No fertiliz T3—mulching; Fert_low T4—50 kg N ha−1, 50 kg P2O5 ha−1, and 50 kg K2O ha−1 mineral fertilization; Fert_medi T5—100 kg N ha−1, 100 kg P2O5 ha−1, and 100 kg K2O ha−1 mineral fertilization; Fert_hight T6—150 kg N ha−1, 150 kg P2O5 ha−1, and 150 kg K2O ha−1 mineral fertilization; Fert_low_o T7—10 t ha−1 cattle manure annually; Fert_medi_o T8—20 t ha−1 cattle manure annually; Fert_medi_o T9—20 t ha−1 cattle manure applied every two years; Fert_hight_o T10—30 t ha−1 cattle manure annually; Fert_medi_o T11—30 t ha−1 cattle manure applied every two years. Species abbreviations: Agrocapi—Agrostis capillaris L.; Alchvulg—Alchemilla vulgaris L.; Achidist—Achillea distants Waldst. & Kit. ex Willd.; Anthodor—Anthoxanthum odoratum L.; Brizmedi—Briza media L., Campabie—Campanula abietina Griseb. & Schenk.; Campsera-Campanula serata Kit. ex Schult.; Carepube—Carex pubescens L.; Cerasyiv—Cerastium sylvaticum L.; Cynocysi—Cynosurus cristatus L.; Crucglab- Cruciata glabra L.; Chryleuc—Leucanthemum vulgare Lam., Dactglom—Dactylis glomerata L.; Deschcaes—Deschampsia caespitosa L.; Festprat—Festuca pratensis Huds.; Festrubr—Festuca rubra L.; Hierpilo—Hieracium pilosela L.; Hypemacu—Hypericum maculatum Crantz; Holclana—Holcus lanatus L.; Luzumult—Luzula multiflora (Ehrh.) Lej.; Lotuscor—Lotus corniculatus L.; Nardstri-Nardus stricta L.; Phleumprat—Phleum pratense L.; Planlanc—Plantago lanceolata L.; Prunvulg.—Prunella vulgaris L.; Poteerec—Potentilla erecta L.; Polyvulg—Polygala vulgaris L.; Poterect—Potentilla erecta (L.) Raeusch.; Ranuacer—Ranunculus acer L.; Stellgram—Stellaria graminea L.; Taraoffi—Taraxacum officinale Weber ex F.H.Wigg.; Trifprat—Trifolium pratense L.; Trifrepe—Trifolium repens L.; Trifochr—Trifolium ochroleucon L.; Thympule—Thymus pulegiodes L.; Verooffi—Veronica officinalis L.; Verocham—Veronica chamaedrys L.; Vaccmyrt—Vaccinium myrtillus L.; Violdecl—Viola declinata L.
To further explore the contribution of individual species to the separation of phytocenoses highlighted in the PCoA analysis, the correlations between species abundance and the two ordination axes are presented in Table 3.
Table 3.
Species projection on PCoA and proximity to fertilization gradient.
The coefficient values show which species were favored by the lack of fertilization and which respond positively to moderate or high nutrient inputs. The correlations between ordination scores and the abundance of dominant species were analyzed, showing that species characteristic of oligotrophic grasslands, such as Nardus stricta, Vaccinium myrtillus, Luzula multiflora, and Potentilla erecta, were negatively correlated with Axis 1. This position on the ordination indicates that their ecological optimum is associated with the absence of fertilization or with very low inputs. Festuca rubra and Carex pallescens also showed the same trend, confirming their vulnerability to management intensification. In contrast, mesotrophic and competitive species had positive and significant correlations with Axis 1, among which Dactylis glomerata, Festuca pratensis, Phleum pratense, and Agrostis capillaris stand out (all p < 0.001), demonstrating an increase in their abundance under medium and high fertilization conditions. Axis 2, although explaining a smaller proportion of the variation, highlighted a distinct behavior of leguminous species. Trifolium pratense and Trifolium repens were negatively and significantly correlated with this axis, showing that their response was influenced by the application of moderate organic inputs. Other species, such as Briza media and Veronica chamaedrys, also presented significant correlations with Axis 2, suggesting the role of secondary ecological factors in structuring communities.
2.4. The Indicator Species Analysis (ISA) on Plant Communities Shaped by Management Scenarios
To identify the species responsible for these differences, we used ISA analysis. Analysis of species indicator values for the four phytosociological groups (Table 4) highlighted floristic differentiation along the management and fertilization gradient. Four distinct groups were identified and profiled through this method.
Table 4.
Indicator value of plant species under applied treatments.
Group 1 (Nardus stricta grasslands) is characterized by strong indicators with high significance: Nardus stricta, Campanula abietina, Cerastium sylvaticum, Cruciata glabra, Polygala vulgaris, Viola declinata, Veronica officinalis, and Vaccinium myrtillus.
Group 2 (transitional communities) is associated with grasses (Anthoxanthum odoratum, Briza media, Cynosurus cristatus) and legumes (Lotus corniculatus, Trifolium ochroleucon, Trifolium repens).
Group 3 (Festuca rubra–Agrostis capillaris co-dominance) is marked by species tolerant to moderate inputs: Festuca rubra, Holcus lanatus, Trifolium pratense, Alchemilla vulgaris, Centaurea pseudophrygia, and Stellaria graminea.
Group 4 (competitive mesotrophs) has indicators such as Agrostis capillaris, Dactylis glomerata, Festuca pratensis, Phleum pratense but also generalist species (Taraxacum officinale, Veronica chamaedrys).
Thus, the indicator species analysis (ISA) confirmed that each phytosociological group has a distinct set of characteristic species, which validates the separation of clusters and the ordination gradient obtained by PCoA.
2.5. The Impact of Management Scenarios on Diversity Indices
To evaluate the effects of fertilization on α diversity, four biodiversity indices (S, H′, E, D) were calculated, and the results are presented in Table 5. The overall assessment of biodiversity indices highlights a significant effect of fertilization treatments on the structure of phytocenoses.
Table 5.
The influence of fertilization treatments on biodiversity indices in grasslands.
Species richness (S) had the highest values in the variants with moderate inputs (T4—37 spp., T7—38 spp., T9—37 spp.), confirming the positive effect of moderate organic or mineral fertilization on diversity. The lowest values were recorded in the treatments with high doses of fertilizers (T6—22.5 spp., T10—22.8 spp.), where the competitive pressure of dominant species led to a reduction in the total number of species.
The Shannon index (H′) increased significantly in the fertilized treatments (T4–T9, values > 2.4) compared to the control (T1—1.54) and the abandonment variant (T2—1.30). This shows a more balanced distribution of species and a higher diversity under moderate fertilization conditions. In contrast, in very high doses of inputs (T6, T10), although the Shannon index remains relatively high (>2.1), diversity tends to be supported by a few dominant species.
Species evenness (E) followed the same trend: low values in the control and abandonment (0.43 and 0.38) compared to the highest values in the fertilized treatments (0.70–0.75). This indicates that fertilization favors a more uniform distribution of species abundance, up to an input threshold, after which the communities become unbalanced again.
The Simpson index (D) reflected the same dynamics, with higher values in fertilized treatments (0.83–0.86) compared to the control and abandonment (0.51, 0.42, respectively).
It is observed that moderate fertilization (T4, T7, T9) maximizes α diversity, which confirms the hypothesis that intermediate inputs favor species coexistence by reducing competitive imbalances. These results support the intermediate disturbance hypothesis, indicating that moderate fertilization maximizes species coexistence, while both the absence of inputs and excessive fertilization lead to a reduction in diversity.
3. Discussion
3.1. The Changes of Different Management Scenarios in the Assemblage of Grassland Communities
The long-term application of fertilizer on grasslands represents a disturbance for plant communities, which is visible in the changes of species richness, shifts between dominant species, and community simplification. All these changes can be used as predictors in the intermediate disturbance hypotheses and provide predictability of succession and change. The application of organic and mineral fertilizers determined a clear difference in the evolution of phytocoenoses, confirming the hypothesis that management intensity is the main factor modulating the structure and diversity of grasslands [46,47]. In unfertilized plots (T1), the persistence of a diverse plant community dominated by Nardus stricta indicates that nutrient limitation acts as a stabilizing ecological filter, maintaining oligotrophic conditions and preventing the dominance of competitive species. This supports the view that low-input systems preserve high functional redundancy, ensuring ecological resilience under fluctuating climatic conditions. Comparable findings from mountain grasslands in Romania suggest that the maintenance of traditional management or periodic mowing is essential to sustain this equilibrium. [48].
The decrease in species richness and evenness observed under abandonment (T2) suggests that the cessation of management triggers directional succession toward ruderal and nitrophilous assemblages. In the absence of disturbance, nutrient accumulation and canopy closure promote the dominance of tall grasses and competitive forbs, suppressing stress-tolerant oligotrophic species. These dynamics highlight that complete abandonment, although often perceived as beneficial for conservation, can paradoxically reduce biodiversity in high-altitude grasslands by disrupting the disturbance–productivity balance that maintains species coexistence. [49]. The reduction in specific richness observed in the case of abandonment was previously reported by [19], who showed that after 6–10 years of non-use, diversity and forage value decrease, being replaced by low-value species, such as Nardus stricta. In contrast, mulched variants (T3) maintained high diversity, indicating that this type of extensive management can substitute traditional mowing and prevent the loss of valuable species [21,22].
Mineral fertilization (T4–T6) induced contrasting ecological responses along the fertilizer gradient. At moderate doses, the balance between nutrient enrichment and competitive pressure favored the coexistence of stress-tolerant and fast-growing species, resulting in the highest levels of diversity. This response illustrates the intermediate disturbance hypothesis, where moderate disturbance intensity maintains coexistence by preventing the dominance of a few competitive taxa and sustaining structural heterogeneity [11,50]. Conversely, excessive nutrient inputs triggered a shift toward communities dominated by tall nitrophilous grasses and competitive legumes, reducing species richness and evenness. Such patterns, also reported for other high nature value grasslands, suggest that high fertilization disrupts resource partitioning and light availability, leading to competitive exclusion and loss of ecological resilience. [20,41,42,51,52]. Similar studies from the Apuseni Mountains (Romania), using NIR techniques, confirm the decrease in protein content and digestibility under intensive NPK, linked to the reduction in Fabaceae, and recommend moderate doses or organic fertilization to maintain biodiversity and forage quality [53], which highlights the importance of integrated management in HNV.
Moderate organic fertilization (10–20 t ha−1 of cattle manure) promoted the recovery of perennial legumes, such as Trifolium pratense and Lotus corniculatus, enhancing both floristic diversity and forage quality. This response reflects the dual ecological function of organic amendments, which gradually release nutrients and improve soil microbial activity without disrupting the oligotrophic balance of mountain grasslands. The slow nutrient mineralization and increase in soil organic matter create favorable microsites for legume establishment, supporting nitrogen fixation and stabilizing the plant community. Similar effects have been reported in other mountain ecosystems, where moderate organic inputs sustain long-term productivity while preserving species coexistence [6,54]. Our results are comparable to long-term studies carried out on Festuca rubra meadows in the Apuseni Mountains, where it was observed that the dose of 10 t ha−1 contributed to balancing the floristic composition and increasing the Shannon index, suggesting that moderate organic fertilization can support both productivity and maintain diversity [38]. At the same time, periodic fertilization with 20 t ha−1 every two years (T9) allowed maintaining a balance between productivity and biodiversity, the results being similar to those reported by [6] and [55] for HNV grasslands in the Romanian Carpathians.
At high manure doses (T10, T11), diversity was again restricted, with values close to those obtained for intensive mineral fertilization. These results indicate that both zero and excessive inputs lead to biodiversity loss, while moderate doses provide a good compromise between productivity and species conservation [28,56,57]. Our results confirm that low NPK doses do not produce major structural changes, in line with 6-year studies in the Apuseni Mountains, where it was shown that low fertilization does not change grassland type but influences species diversity [58].
The nonlinear response of diversity to the fertilization gradient can be explained by ecological mechanisms related to asymmetric competition for light and nutrients. At moderate doses, increased resource availability reduces the advantage of dominant species and allows stress-tolerant species to coexist, which maximizes diversity. In contrast, at high inputs, excessive fertilization favors nitrophilic species and competitive grasses, which, by increasing biomass and shading the lower layer, gradually exclude oligotrophic species adapted to poor soils. This transition from balanced communities to those dominated by a few opportunistic species explains the sharp decline in Shannon and Simpson indices at high fertilization rates.
3.2. Plant Specificity to Applied Treatments
The results of ISA show that each identified phytocoenotic group has a distinct set of species with diagnostic value. Oligotrophic grasslands are characterized by Nardus stricta, Vaccinium myrtillus, and Potentilla erecta, species typical for acidic soils and restrictive conditions [5], which indicate a native adaptation to reduced nutrient conditions. Several recent studies confirm that Nardus stricta is a characteristic species for oligotrophic grasslands on acidic soils, such as the case of reserves in Germany, where typical acidophilic species decrease with the improvement of soil acidity [59]. The presence and the coverage of this species can be used to indicate the shifts from acidic to neutral or alkaline conditions. Vaccinium myrtillus also thrives in restricted acidophilic environments, such as degraded grasslands [60,61]. The case study of Nardus stricta grasslands (6230) provides further evidence for the role of these species as ecological indicators in oligotrophic communities [8,62]. In contrast to oligotrophic grasslands, mesotrophic and eutrophic communities were dominated by species with higher nutritional requirements, such as Dactylis glomerata, Festuca pratensis, and Trifolium pratense. These species are recognized as competitive under moderate and intense fertilization conditions, where biomass accumulation and the reduction in species sensitive to low nutrient inputs are favored. This opposition between species characteristic of oligotrophic environments and those associated with mesotrophic/eutrophic communities highlights the impact of the fertilization gradient on the structure of mountain vegetation. Similar results have been reported in recent studies on mixtures of grasses and legumes fertilized with mineral and organic fertilizers, where Dactylis glomerata and Trifolium pratense became dominant [63,64]. Parallel findings have also been presented for high nature value mountain grasslands in the Apuseni Mountains, where increased fertilization levels have led to an increased role for mesotrophic and eutrophic species [6]. Such results confirm the role of indicator species as essential tools for diagnosing the agrochemical status of the soil and for underpinning adaptive management strategies for mountain grasslands [7,8,40].
Overall, the results demonstrate that Nardus stricta meadows respond differently to the intensity and type of fertilization, but the general trend remains the same: extensive or moderate management preserves diversity, while excessive intensification leads to simplification of the phytocoenosis. These findings support the recommendations for adaptive management and sustainable use of mountain resources [59,65,66,67]. Changes generated by fertilization are subsequently amplified or attenuated by climate variability, which complicates predictions of the long-term dynamics of these ecosystems. Although fertilization treatments explain a large part of the observed variation in vegetation composition, these changes cannot be interpreted as isolated cases. During the experimental period (2020–2024), the average annual temperature in the study area ranged between 5.9 and 6.8 °C, and the total amount of precipitation between 620 and 699 mm, with a pronounced minimum in 2024. These moderate but significant climatic variations in the mountain context may partly explain the annual fluctuations in diversity indices, particularly the slight reduction observed in the last year, when the precipitation deficit accentuated water stress and interspecific competition. Thus, the effects of fertilization on community structure cannot be analyzed in isolation but in correlation with the thermal and water regime, which modulates the response of oligotrophic and mesotrophic species in mountain conditions. Interannual climatic fluctuations (temperature, water balance) can amplify or attenuate the effects of management, generating both directional successions and annual variations in the structure of phytocoenoses. This aspect is particularly relevant for mountain grasslands, where the interaction between anthropogenic and climatic factors determines the dynamics of plant communities. Studies in the Apuseni Mountains show that these changes are magnified by climatic fluctuations, which induce directional successions in initial periods and annual fluctuations related to temperature and water balance, affecting the vegetation composition in similar grasslands [68]. Climate fluctuations can increase the speed of vegetation changes in mountain grasslands—different studies show directional successions towards species adapted to warmer and drier conditions [69] and changes in floral composition under warming conditions [70]. These results indicate a background of a high global sensitivity of vegetation indicators to temperature and precipitation (global meta-analysis) and dramatic examples, such as the effects of the extreme drought of 2022 [71]. The context demands the necessity to integrate climate monitoring into HNV management in order to distinguish between human-induced and natural changes—as demonstrated by integrated monitoring programs for HNV [72], the German standardized observation system [73], and the international GLORIA network dedicated to alpine eco-vegetation [74].
3.3. Species Structure and Diversity Under the Input Gradients
The results of our study demonstrate that fertilization intensity plays a central role in shaping the structure and diversity of high nature value (HNV) grasslands. There is a clear separation of grassland communities along the input gradient, from unfertilized oligotrophic grasslands to intensively fertilized mesotrophic systems. This pattern highlights the sensitivity of oligotrophic grasslands, dominated by Nardus stricta and associated stress-tolerant species, to increased nutrient availability. Also, intensive fertilization reduces diversity and favors competitive species, potentially increasing the risk of toxic species in grasslands [75]. To prevent animal poisoning, technologies such as NIR hyperspectral imaging can discriminate species and botanical families, allowing the identification of toxic species in grasslands similar to those in the Apuseni Mountains [76]. This highlights the need for advanced monitoring in sustainable HNV management. Our results are consistent with previous reports showing that moderate fertilization can improve species coexistence by balancing competitive dynamics, while both the absence of inputs and excessive fertilization reduce biodiversity [6,46,57,77,78]. Intermediate input treatments (T4, T7, T9) maximized species richness and Shannon diversity, supporting the intermediate disturbance hypothesis in grassland ecosystems [11,12,50].
Overall, the results highlight that reduced or absent inputs favor oligotrophic and stress-tolerant species [5], thus maintaining the traditional character and high biodiversity of mountain meadows. In contrast, intense fertilization causes a transition towards communities dominated by competitive grasses and perennial legumes, with a visible reduction in floristic diversity. These findings are in agreement with both national studies [6,40,46,79] and recent international research showing that excessive mineral fertilization increases biomass but significantly reduces species richness and community evenness [47,57]. Also, the global analysis carried out by [69] confirms that climatic fluctuations can amplify these effects, accelerating the succession towards simpler mesotrophic communities and reducing the stability of ecosystems. Thus, the transition from oligotrophic to mesotrophic systems, accompanied by gradual losses of diversity, reflects a general pattern recently reported for mountain ecosystems in Europe [26,33,70].
For high natural value grasslands, the application of moderate fertilization represents an optimal solution for maintaining the balance between productivity and biodiversity and sustainable long-term management. Extreme management and high-input fertilization or abandonment, the marginal effects of the total lack of inputs to excessive fertilization, lead to the simplification of plant communities. Both these extremes have a great magnitude impact and lead to biodiversity losses and dominance shifting and produce a high instability of community assemblage. This opposition between the species characteristic of oligotrophic environments (Nardus stricta, Vaccinium myrtillus, Potentilla erecta) and those associated with mesotrophic and eutrophic communities (Dactylis glomerata, Festuca pratensis, Trifolium pratense) highlights the importance of the fertilization gradient as a determining factor in the structure and dynamics of mountain grasslands. Maintaining the balance between productivity and diversity depends on adaptive management, which integrates both agricultural pressure and climatic variability, which is the only viable option for the conservation of biodiversity and the sustainable use of resources in the mountain area.
The study demonstrates that fertilization intensity is the main determinant of the structure and diversity of high nature value (HNV) grasslands. Potential limitations of this are associated with the short period of experimentation (only 4 years), which offers a a good image of the potential change trends. The continuity in experimentation will offer a complex image of both the successional trajectories of vegetation and the resilience potential of plant communities. The study area limits the application of the results to similar types of grasslands, but it needs to be replicated in other climatic conditions to assess the combined and unique impact of treatments and climate. Interdisciplinarity is a good solution to connect the changes in plant communities and species dynamics with soil characteristics and evolution due to the application of treatments.
At the applied level, a clear definition of oligotrophic grasslands is essential for guiding sustainable management strategies and for maintaining the HNV character of these ecosystems. The results show that moderate input can represent a viable compromise between productivity and biodiversity conservation, maintaining the HNV character of grasslands. The identification of species with an indicator value for each level of intensification provides a useful tool for monitoring changes and implementing adaptive management strategies.
4. Materials and Methods
4.1. Soil and Climatic Conditions
The experiment was conducted during the 2020–2024 period at a permanent grassland derived from Nardus stricta L. on the Plaiu Sarului (47030’42’’ N, 25037’95’’ E) in the region of the North-Eastern Carpathians (Dorna Depression) at 845 m above sea level. The type of soil within the experimental field is represented by luvisol, in which a well-developed thatch of dead and living stems, leaves, and roots with a thickness of 8–10 cm is noticeable [80]. Mineral soil horizon of moderate humus accumulation (2.43%) below thatch layer is light in color with moderately acidic reaction (pH = 5.4) and medium texture (content of clay 22,3%). The eluvial horizon, 42 cm thick, is weakly skeletal with medium texture (clay = 19.5%) and is moderately acidic (pH = 5.1). The value of the cation-exchange capacity from the humiferous horizon (Ao) is 11.7 meq 100−1 g of soil, and the degree of base saturation is 64,3%. The humus accumulation horizon is moderately supplied with nitrogen (0.952%) and potassium (257 ppm) and poorly supplied with mobile phosphorus (12.4 ppm).
The climatic conditions in the area are characterized by an average temperature of 6.3 °C and 675 mm of total annual precipitation. In the Plaiu Sarului area, the amounts of atmospheric precipitation are characteristic of depressional mountain areas. Thus, the months with the lowest precipitation are January, February, March, and December (32.4 mm to 38.9 mm), and the months with the highest precipitation are obviously the summer months of May, June, July, and August (85.4 mm to 118.2 mm). The relative humidity of the air has high values throughout the year of 80–92%, with an average value of 85.2%. The precipitation recorded during the experimental period was 698.9 mm in 2020, 677.2 mm in 2021, 695.5 mm in 2022, 680.1 mm in 2023, and 620.6 mm in 2024. The average annual temperatures were 6.8 °C in 2020, 5.9 °C in 2021, 6.3 °C in 2022, 6.6 °C in 2023, and 6.7 °C in 2024.
4.2. Experimental Design
A monofactorial experiment with 11 treatments was organized according to the randomized block method in four replicates.
This classification reflects the reference to the thresholds established by the Nitrates Directive (170 kg N ha−1year−1), as well as the ecological response of the vegetation observed in the field [81, 82].
The research covered four blocks, and the experiment had the following treatments:
- •
- T1—unfertilized (control);
- •
- T 2—abandonment (unharvested or non-grazing);
- •
- T3—mulching (cut and leave the biomass on site);
- •
- T4—fertilization with 50 kg N ha−1, 50 kg P2O5 ha−1, and 50 kg K2O ha−1 applied annually;
- •
- T5—fertilization with 100 kg N ha−1, 100 kg P2O5 ha−1, and 100 kg K2O ha−1 annually;
- •
- T6—fertilization with 150 kg N ha−1, 150 kg P2O5 ha−1, and 150 kg K2O ha−1 annually;
- •
- T7—10 t ha−1 cattle manure annually;
- •
- T8—20 t ha−1 cattle manure annually;
- •
- T9—20 t ha−1 cattle manure at 2 years;
- •
- T10—30 t ha−1 cattle manure annually;
- •
- T11—30 t ha−1 cattle manure at 2 years.
Two types of fertilizers were used: an organic one represented by well-fermented cattle manure (older than two years) and a mineral one represented by a complex fertilizer with nitrogen, phosphorus, and potassium at a ratio of 20:20:20 (N:P2O5:K2O). The cattle manure had 16% dry matter and the following chemical composition: N—0.445%, P2O5—0.212%, and K2O—0.695%. Fertilizers were applied manually in the same period, in early spring, before the start of active vegetation growth, respectively.
4.3. Vegetation Survey
The floristic studies were performed according to a modified Braun–Blanquét method (Figure 4, Table 6), with smaller intervals adapted for a better and more realistic vegetation assessment [81]. Floristic studies were performed in 2024, at the beginning of July, when the grasses were in the phenological flowering phase. The optimal time for determining the floristic composition is in July, when the grasses are in bloom, which ensures the correctness of the composition of the floristic survey, as shown by other studies conducted on semi-natural grasslands [82,83]. Mowing was performed in the experimental field once per year at a height of 4 cm in the middle of July.
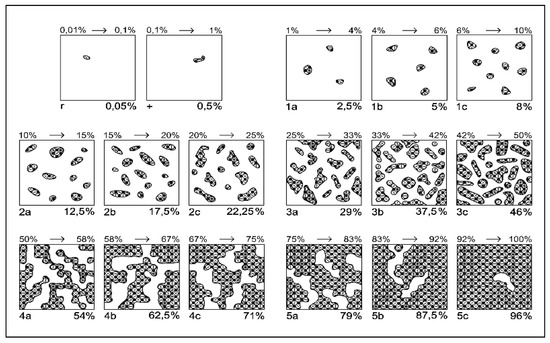
Figure 4.
Modified Braun–Blanquét scale for grasslands, based on species [81]. Legend: 1 to 5 indicates the class of coverage; a,b,c indicate the sub-note of each class.
Table 6.
Modified Braun–Blanquét scale for assessing the abundance–dominance of plant species, based on classes and sub-classes [81].
4.4. Data Analysis
In order to completely explore the data and to analyze the qualitative effect of treatments, we have used four categories of treatments: zero input, low input, medium input, and high input. This approach can be used to additionally quantify the effect of fertilizer as reported by the applied quantity. To integrate the experimental treatments (T1–T11) into the multivariate analyses, they were coded according to the level of nutrient inputs and the type of fertilizer: “Zero-input” category, characterized by the lack of nutrient input (T1–T3); low-input category: T4 and T7; medium-input category: T5, T8, T9, and T11; and high-input category: T6 and T10. The differentiation between mineral and organic fertilization was introduced by additional binary variables (e.g., Low_o, Low_m, Med_o, Med_m), allowing the separate testing of the effects of the type of fertilizer on the community structure. The procedure for classifying the experimental treatments by input categories (“Zero-input”, “Low-input”, “Medium-input”, “High-input”) follows common practices in the experimental ecology literature, where treatments with different levels of fertilization (or nutrient input) are compared for effects on plant diversity and productivity [84,85].
Data analysis was performed with PC-ORD 7 software [86,87]. For cluster analysis, the Sørensen similarity index (Bray–Curtis) and the UPGMA (group average linkage) method were used. Both the index and the method were applied due to their ability to show a clear delimitation between ecological and phytosociological groups from the analyzed plant communities [88,89,90]. The cutoff level of the dendrogram was set at approximately 60% of the remaining information, which allowed obtaining clusters with ecological and phytosociological relevance. The choice of this threshold aligns with the methodologies used in recent analyses, where UPGMA cutoff values based on Sørensen/Bray–Curtis were used to obtain ecologically meaningful groupings [14,90].
Ordination analysis was performed by PCoA (Principal Coordinates Analysis) using the Sørensen (Bray–Curtis) distance, a method frequently applied in ecology to compare plant communities based on floristic composition. The choice of PCoA was based on its advantages over iterative methods, such as NMDS, as it provides a unique and reproducible solution, with direct interpretation of distances between samples. In addition, the Bray–Curtis distance is considered one of the most robust dissimilarity measures for vegetation data, being widely used in recent studies of fertilization and management of grasslands and soils [91,92,93]. For each experimental variant, vectors corresponding to fertilization levels (zero, low, medium, high) and input type (organic or mineral) were drawn. The vectors were normalized, and their significance was tested by permutations (n = 999). PC-ORD offers ordination options based on Bray–Curtis distances, overlays or joint plots, and randomization tests, which makes the choice to use vectors corresponding to fertilization levels consistent with good methodological practices [87,94]. The first two ordination axes were retained for interpretation, as they together explained over 95% of the variation in floristic composition.
To test the differences between the groups identified by cluster and ordination analysis, MRPP (Multi-Response Permutation Procedure) analysis was applied. Statistical significance was assessed at the α = 0.05 level. Additionally, indicator species analysis (ISA) [95] was used to identify the species characteristic of each grassland group. For each experimental treatment, α diversity indices were calculated—species richness (S), Shannon–Wiener index (H′), species evenness (E), and Simpson index (D). These indices are widely used in grassland ecology studies, being considered robust tools for assessing the diversity and balance of plant communities [51,96,97,98]. The formulas for these indices are standard in community ecology [99] and were calculated based on relative abundance data. Differences between treatments were tested by one-way ANOVA using SPSS version 29 Academic and scored for significant differences by the LSD test (p < 0.05).
5. Conclusions
A clear separation of communities along the input gradient, from oligotrophic grasslands dominated by stress-tolerant species to mesotrophic systems characterized by competitive grasses, was recorded.
Moderate fertilization maximizes species richness and floristic diversity by maintaining a balance between competing species and those adapted to restrictive conditions.
Both the abandonment and the application of excessive input cause a reduction in biodiversity, favoring the dominance of a few tolerant or competitive species.
Each management scenario presents a list of indicator species associated with oligotrophic, mesotrophic, and eutrophic communities.
The controlled application of organic and mineral fertilizers, below critical thresholds, emerges as an effective solution for the sustainable use of mountain plant resources.
Author Contributions
Conceptualization, C.S. (Costel Samuil) and V.V.; methodology, C.S. (Costel Samuil) and V.V.; software, A.I.N. and B.G.; validation, C.S. (Culiță Sîrbu), A.I.N., and B.G.; formal analysis, C.S. (Culiță Sîrbu) and A.I.N.; writing—original draft preparation, C.S. (Costel Samuil) and V.V.; writing—review and editing, C.S. (Costel Samuil), V.V., and A.I.N.; supervision, C.S. (Culiță Sîrbu), A.I.N., and B.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Ion Ionescu de la Brad, Iasi University of Life Sciences.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Buchmann, N.; Fuchs, K.; Feigenwinter, I.; Gilgen, A.K. Multifunctionality of Permanent Grasslands: Ecosystem Services and Resilience to Climate Change. In Grassland Science in Europe; European Grassland Federation: Zurich, Switzerland, 2019; Volume 24, pp. 19–26. [Google Scholar]
- Lomba, A.; McCracken, D.; Herzon, I. High Nature Value Farming Systems in Europe. Ecol. Soc. 2023, 28, 20. [Google Scholar] [CrossRef]
- Liu, S.; Ward, S.E.; Wilby, A.; Manning, P.; Gong, M.; Davies, J.; Killick, R.; Quinton, J.N.; Bardgett, R.D. Multiple Targeted Grassland Restoration Interventions Enhance Ecosystem Service Multifunctionality. Nat. Commun. 2025, 16, 3971. [Google Scholar] [CrossRef]
- Jakobsson, S.; Envall, I.; Bengtsson, J.; Rundlöf, M.; Svensson, M.; Åberg, C.; Lindborg, R. Effects on Biodiversity in Semi-Natural Pastures of Giving the Grazing Animals Access to Additional Nutrient Sources: A Systematic Review. Environ. Evid. 2024, 13, 18. [Google Scholar] [CrossRef]
- Sângeorzan, D.; Rotar, I.; Păcurar, F.; Vaida, I.; Suteu, A.; Deac, V. The definition of oligotrophic grasslands. Rom. J. Grassl. Forage Crops 2018, 17, 33–43. [Google Scholar]
- Vaida, I.; Păcurar, F.; Rotar, I.; Tomoş, L.; Stoian, V. Changes in Diversity Due to Long-Term Management in a High Natural Value Grassland. Plants 2021, 10, 739. [Google Scholar] [CrossRef]
- Maruşca, T.; Păcurar, F.S.; Taulescu, E.; Vaida, I.; Nicola, N.; Scrob, N.; Dragoș, M.M. Indicative species for the agrochemical properties of mountain grasslands soil from the Apuseni Natural Park (ROSci 0002). Rom. J. Grassl. Forage Crops. 2022, 31, 3971. [Google Scholar]
- Păcurar, F. Specii Indicator Pentru Evaluarea şi Elaborarea Managementului Sistemelor de Pajişti cu Înaltă Valoare Naturală (HNV); Casa Cărţii de Ştiinţă: Cluj-Napoca, Romania, 2020. [Google Scholar]
- Rușdea, E.; Reif, A.; Höchtl, F.; Păcurar, F.; Rotar, I.; Stoie, A.; Dahlström, A.; Svensson, R.; Aronsson, M. Grassland Vegetation and Management-on the Interface between Science and Education. Bull. Univ. Agr. Sci. Vet. Med. Cluj-Napoca Agric. 2011, 68, 12–20. [Google Scholar] [CrossRef]
- Balazsi, A.; Rotar, I.; Păcurar, F.; Vidican, R.; Pleșa, A.; Gliga, A.; Mălinaș, A. Mulching and mulching with organic fertilizing as an alternative way to conserve the oligotrophy grasslands’ phytodiversity and maintain their productivity in Apuseni Mountains. Rom. J. Grassl. Forage Crops. 2014, 9, 7. [Google Scholar]
- Humbert, J.Y.; Dwyer, M.J.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Change Biol. 2016, 22, 110–120. [Google Scholar] [CrossRef]
- Tilman, D. Secondary Succession and the Pattern of Plant Dominance along Experimental Nitrogen Gradients. Ecol. Monogr. 1987, 57, 189–214. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U. Disturbance Effects on Species Diversity and Functional Diversity in Riparian and Upland Plant Com-Munities. Ecology 2010, 91, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Jiao, F.; Li, Y.H.; Kallenbach, R.L. Anthropogenic disturbances are key to maintaining the biodiversity of grasslands. Sci. Rep. 2016, 6, 22132. [Google Scholar] [CrossRef]
- Diviaková, A.; Ollerová, H.; Stašiov, S.; Veverková, D.; Novikmec, M. Plant Functional Structure Varies across Different Management Regimes in Submontane Meadows. Nat. Conserv. 2024, 56, 181–200. [Google Scholar] [CrossRef]
- Fernández-Guisuraga, J.M.; Fernández-García, V.; Tárrega, R.; Marcos, E.; Valbuena, L.; Pinto, R.; Monte, P.; Beltrán, D.; Huerta, S.; Calvo, L. Transhumant Sheep Grazing Enhances Ecosystem Multifunctionality in Productive Mountain Grasslands: A Case Study in the Cantabrian Mountains. Front. Ecol. Evol. 2022, 10, 861611. [Google Scholar] [CrossRef]
- Gaga, I.; Rotar, I.; Păcurar, F.S.; Plesa, A.D.; Vaida, I. Study of Grassland Types from the Agricultural Research-Development Station (Ards) Turda. Rom. J. Grassl. Forage Crops 2020, 21, 19. [Google Scholar]
- Vaida, I.; Rotar, I.; Păcurar, F.; Vidican, R.; Pleşa, A.; Mălinaş, A.; Stoian, V. Impact on the Abandonment of Semi-Natural Grasslands from Apuseni Mountains. Bull. Univ. Agr. Sci. Vet. Med. Cluj-Napoca Agric. 2016, 73, 323–331. [Google Scholar] [CrossRef][Green Version]
- Păcurar, F.; Rotar, I.; Pleșa, A.; Balázsi, Á.; Vidican, R. Study of the Floristic Composition of Certain Secondary Grasslands in Different Successional Stages as a Result of Abandonment. Rom. J. Grassl. Forage Crops 2015, 72, 11165. [Google Scholar] [CrossRef][Green Version]
- Păcurar, F.; Balázsi, Á.; Rotar, I.; Vaida, I.; Reif, A.; Vidican, R.; Ruşdea, E.; Stoian, V.; Sângeorzan, D. Technologies Used for Maintaining Oligotrophic Grasslands and Their Biodiversity in a Mountain Landscape. Rom. Biotechnol. Lett. 2018, 23, 13614–13623. [Google Scholar] [CrossRef]
- Rotar, I.; Păcurar, F.; Vaida, I.; Nicola, N.; Pleșa, A. The Effect of Mulching on a Grasslands in the Apuseni Mountains. Rom. J. Grassl. Forage Crops 2020, 25, 1128–1135. [Google Scholar]
- Zarzycki, J.; Józefowska, A.; Kopeć, M. Can Mulching or Composting Be Applied to Maintain Semi-Natural Grassland Managed for Biodiversity? J. Nat. Conserv. 2024, 78, 126584. [Google Scholar] [CrossRef]
- Stoian, V.; Vidican, R.; Păcurar, F.; Corcoz, L.; Pop-Moldovan, V.; Vaida, I.; Vâtcă, S.-D.; Stoian, V.A.; Plesa, A. Exploration of Soil Functional Microbiomes—A Concept Proposal for Long-Term Fertilized Grasslands. Plants 2022, 11, 1253. [Google Scholar] [CrossRef] [PubMed]
- Corcoz, L.; Păcurar, F.; Pop-Moldovan, V.; Vaida, I.; Stoian, V.; Vidican, R. Mycorrhizal Patterns in the Roots of Dominant Festuca Rubra in a High-Natural-Value Grassland. Plants 2021, 11, 112. [Google Scholar] [CrossRef]
- Mălinas, A.; Rotar, I.; Vidican, R.; Iuga, V.; Păcurar, F.; Mălinas, C.; Moldovan, C. Designing a Sustainable Temporary Grassland System by Monitoring Nitrogen Use Efficiency. Agronomy 2020, 10, 149. [Google Scholar] [CrossRef]
- Sângeorzan, D.D.; Păcurar, F.; Reif, A.; Weinacker, H.; Ruşdea, E.; Vaida, I.; Rotar, I. Detection and Quantification of Arnica Montana L. Inflorescences in Grassland Ecosystems Using Convolutional Neural Networks and Drone-Based Remote Sensing. Remote Sens. 2024, 16, 2012. [Google Scholar] [CrossRef]
- Brinkmann, K.; Păcurar, F.; Rotar, I.; Rușdea, E.; Auch, E.; Reif, A. The Grasslands of the Apuseni Mountains, Romania. In Grasslands in Europe of High Nature Value; KNNV Publishing: Wageningen, The Netherlands, 2009; pp. 9–20. ISBN 9789050113168. [Google Scholar]
- Janišová, M.; Škodová, I.; Magnes, M.; Iuga, A.; Biro, A.-S.; Ivașcu, C.M.; Ďuricová, V.; Buzhdygan, O.Y. Role of Livestock and Traditional Management Practices in Maintaining High Nature Value Grasslands. Biol. Conserv. 2025, 309, 111301. [Google Scholar] [CrossRef]
- Reif, A.; Auch, E.; Bühler, J.; Brinkmann, K.; Goia, A.I.; Pacurar, F.; Rusdea, E. Landschaft Und Landnutzung Im Apusenigebirge Rumäniens. Carinthia II 2005, 195, 161–201. [Google Scholar]
- Marușca, T.; Păcurar, F.S.; Scrob, N.; Vaida, I.; Nicola, N.; Taulescu, E.; Dragoș, M.; Lukács, Z. Contributions to the Assessment of Grasslands Productivity of the Apuseni Natural Park (ROSci 0002). Rom. J. Grassl. Forage Crops 2021, 24, 23. [Google Scholar]
- Michler, B.; Rotar, I.; Păcurar, F. Biodiversity and Conservation of Medicinal Plants: A Case Study in the Apuseni Mountains in Romania. Bull. USAMV-CN 2006, 62, 86–87. [Google Scholar]
- Hagemann, N.; Gerling, C.; Hölting, L.; Kernecker, M.; Markova-Nenova, N.N.; Wätzold, F.; Wendler, J.; Cord, A.F. Improving Re-Sult-Based Schemes for Nature Conservation in Agricultural Landscapes—Challenges and Best Practices from Selected European Countries. Reg. Environ. Change 2025, 25, 12. [Google Scholar] [CrossRef]
- Păcurar, F.; Reif, A.; Rusḑea, E. Conservation of Oligotrophic Grassland of High Nature Value (HNV) through Sustainable Use of Arnica Montana in the Apuseni Mountains, Romania. In Medicinal Agroecology; CRC Press Boca Raton: London, New York, 2023; pp. 177–201. [Google Scholar]
- Elliott, J.; Tindale, S.; Outhwaite, S.; Nicholson, F.; Newell-Price, P.; Sari, N.H.; Hunter, E.; Sánchez-Zamora, P.; Jin, S.; Gallardo-Cobos, R.; et al. European Permanent Grasslands: A Systematic Review of Economic Drivers of Change, Including a Detailed Analysis of the Czech Republic. Sustainability 2024, 13, 116. [Google Scholar] [CrossRef]
- Sattler, C.; Schrader, J.; Hüttner, M.L.; Henle, K. Effects of Management, Habitat and Landscape Characteristics on Biodiversity of Orchard Meadows in Central Europe: A Brief Review. Nat. Conserv. 2024, 55, 103–134. [Google Scholar] [CrossRef]
- Pleșa, A.; Rotar, I.; Păcurar, F.; Vidican, R.; Balaszi, A. About Arnica Montana Grasslands and Their Context. Bull. USAMV Agr. 2014, 71, 282–285. [Google Scholar] [CrossRef][Green Version]
- Reif, A.; Ruşdea, E.; Păcurar, F.; Rotar, I.; Brinkmann, K. A Traditional Cultural Landscape in Transformation. Mt. Res. Dev. 2008, 28, 18–22. [Google Scholar] [CrossRef]
- Vaida, I.; Rotar, I.; Păcurar, F. The Cumulative Effect of Manure on a Festuca Rubra Grasslands for 15 Years. Bull. Univ. Agr. Sci. Vet. Med. Cluj-Napoca Agric. 2017, 74, 126. [Google Scholar] [CrossRef][Green Version]
- Pleșa, A.; Rotar, I.; Păcurar, F.; Vidican, R.; Balázsi, A.; Stoian, V. Organic Fertilization on Mountain Grassland. In Grassland and Forages in High Output Dairy Farming Systems; Hopkins, A., Collins, R.P., Fraser, M.D., King, V.R., Lloyd, D.C., Moorby, J.M., Robson, P.R.H., Eds.; Grassland Science in Europe; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; Volume 20, p. 289. [Google Scholar]
- Rotar, I.; Vaida, I.; Păcurar, F. Species with Indicative Values for the Management of the Mountain Grasslands. Rom. Agric. Res. 2020, 37, 189–196. [Google Scholar] [CrossRef]
- De Schrijver, A.; De Frenne, P.; Ampoorter, E.; Van Nevel, L.; Demey, A.; Wuyts, K.; Verheyen, K. Cumulative nitrogen input drives species loss in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2011, 20, 803–816. [Google Scholar] [CrossRef]
- Soons, M.B.; Hefting, M.M.; Dorland, E.; Lamers, L.P.M.; Versteeg, C.; Bobbink, R. Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biol. Conserv. 2017, 212, 390–397. [Google Scholar] [CrossRef]
- Vîntu, V.; Samuil, C.; Sirbu, C.; Popovici, C.I.; Stavarache, M. Sustainable Management of Nardus stricta L. Grasslands in Romania’s Carpathians. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 142–145. [Google Scholar] [CrossRef][Green Version]
- Corcoz, L.; Păcurar, F.; Pop-Moldovan, V.; Vaida, I.; Pleșa, A.; Stoian, V.; Vidican, R. Long-Term Fertilization Alters Mycorrhizal Colonization Strategy in the Roots of Agrostis capillaris. Agriculture 2022, 12, 847. [Google Scholar] [CrossRef]
- Balazsi, A.; Păcurar, F.; Mihu-Pintilie, A.; Konold, W. How Do Public Institutions on Nature Conservation and Agriculture Contribute to the Conservation of Species-Rich Hay Meadows? Int. J. Conserv. Sci. 2018, 9, 549–564. [Google Scholar]
- Gaga, I.; Păcurar, F.; Vaida, I.; Plesa, A.; Rotar, I. Responses of Diversity and Productivity to Organo-Mineral Fertilizer Inputs in a High-Natural-Value Grassland, Transylvanian Plain, Romania. Plants 2022, 11, 1975. [Google Scholar] [CrossRef]
- Shi, T.S.; Collins, S.L.; Yu, K.; Peñuelas, J.; Sardans, J.; Li, H.; Ye, J.S. A Global Meta-Analysis on the Effects of Organic and Inorganic Fertilization on Grasslands and Croplands. Nat. Commun. 2024, 15, 3411. [Google Scholar] [CrossRef]
- Titěra, J.; Pavlů, V.V.; Pavlů, L.; Hejcman, M.; Gaisler, J.; Schellberg, J. Response of Grassland Vegetation Composition to Different Fertilizer Treatments Recorded over Ten Years Following 64 Years of Fertilizer Applications in the Rengen Grassland Experiment. Appl. Veg. Sci. 2020, 23, 417–427. [Google Scholar] [CrossRef]
- Melts, I.; Lanno, K.; Sammul, M.; Uchida, K.; Heinsoo, K.; Kull, T.; Laanisto, L. Fertilising Semi-Natural Grasslands May Cause Long-Term Negative Effects on Both Biodiversity and Ecosystem Stability. J. Appl. Ecol. 2018, 55, 1851–1864. [Google Scholar] [CrossRef]
- Schaub, S.; Finger, R.; Leiber, F.; Probst, S.; Kreuzer, M.; Weigelt, A.; Buchmann, N.; Scherer-Lorenzen, M. Plant Diversity Effects on Forage Quality, Yield and Revenues of Semi-Natural Grasslands. Nat. Commun. 2020, 11, 768. [Google Scholar] [CrossRef] [PubMed]
- Hautier, Y.; Zhang, P.; Loreau, M.; Wilcox, K.R.; Seabloom, E.W.; Borer, E.T.; Byrnes, J.E.; Koerner, S.E.; Komatsu, K.J.; Lefcheck, J.S.; et al. General Destabilizing Effects of Eutrophication on Grassland Productivity at Multiple Spatial Scales. Nat. Commun. 2020, 11, 5375. [Google Scholar] [CrossRef] [PubMed]
- Gârda, N.T.; Păcurar, F.; Rotar, I.; Bogdan, A. The floristic and stational characterisation of the grassland subtype Agrostis capillaris+ Trisetum flavescens from Apuseni Mountains, Romania. Res. J. Agric. Sci. 2010, 42, 454–457. [Google Scholar]
- Dale, L.M.; Thewis, A.; Rotar, I.; Boudry, C.; Păcurar, F.S.; Lecler, B.; Agneessens, R.; Dardenne, P.; Baeten, V. Fertilization Effects on the Chemical Composition and in Vitro Organic Matter Digestibility of Semi-Natural Meadows as Predicted by NIR Spectrometry. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 58–64. [Google Scholar] [CrossRef]
- Isselstein, J.; Jeangros, B.; Pavlu, V. Agronomic Aspects of Biodiversity Targeted Management of Temperate Grasslands in Europe—A Review. Agron. Res. 2005, 3, 139–151. [Google Scholar]
- Weigelt, A.; Weisser, W.W.; Buchmann, N.; Scherer-Lorenzen, M. Biodiversity for multifunctional grasslands: Equal productivity in high-diversity low-input and low-diversity high-input systems. Biogeosciences 2009, 6, 1695–1706. [Google Scholar]
- Rotar, I.; Păcurar, F.; Vidican, R.; Bogdan, A. Impact of grassland management on occurrence of Arnica montana L. Grassland—A European Resource? Grassl. Sci. Eur. 2012, 17, 701–806. [Google Scholar]
- Wan, L.; Liu, G.; Su, X. Organic Fertilization Balances Biodiversity Maintenance, Grass Production, Soil Storage, Nutrient Cycling and Greenhouse Gas Emissions for Sustainable Grassland Development in China: A Meta-Analysis. Agric. Ecosyst. Environ. 2025, 381, 109473. [Google Scholar] [CrossRef]
- Rotar, I.; Păcurar, F.; Pleșa, A.; Balázsi, Á. Low mineral fertilization on grassland after 6 years. Lucr. Ştiinţifice Ser. Agron. 2015, 58, 51–54. [Google Scholar]
- Peppler-Lisbach, C.; Kratochwil, A.; Mazalla, L.; Rosenthal, G.; Schwabe, A.; Schwane, J.; Stanik, N. Synopsis of Nardus Grassland Resurveys Across Germany: Is Eutrophication Driven by a Recovery of Soil pH After Acidification? J. Veg. Sci. 2025, 36, 70040. [Google Scholar] [CrossRef]
- Sherstiuk, M.; Skliar, V.; Kašpar, J.; Mohammadi, Z. Size and Vitality Characteristics of Bilberry (Vaccinium myrtillus L.) Popula-Tions in the Central Bohemian Region of the Czech Republic: A Case Study of Non-Timber Forest Products. Glob. Ecol. Conserv. 2024, 56, 03295. [Google Scholar]
- Hegland, S.J.; Gillespie, M.A. Vaccinium Dwarf Shrubs Responses to Experimental Warming and Herbivory Resistance Treatment Are Species-and Context Dependent. Front. Ecol. Evol. 2024, 12, 1347837. [Google Scholar] [CrossRef]
- Dalle Fratte, M.; Cerabolini, B.E. Extending the Interpretation of Natura 2000 Habitat Types beyond Their Definition Can Bias Their Conservation Status Assessment: An Example with Species-Rich Nardus Grasslands (6230). Ecol. Indic. 2023, 156, 111113. [Google Scholar] [CrossRef]
- Borawska-Jarmułowicz, B.; Mastalerczuk, G.; Janicka, M.; Wróbel, B. Effect of Silicon-Containing Fertilizers on the Nutritional Value of Grass–Legume Mixtures on Temporary Grasslands. Agriculture 2022, 12, 145. [Google Scholar] [CrossRef]
- Călina, J.; Călina, A.; Iancu, T.; Miluț, M.; Croitoru, A.C. Research on the Influence of Fertilization System on the Production and Sustainability of Temporary Grasslands from Romania. Agronomy 2022, 12, 2979. [Google Scholar] [CrossRef]
- Bobbink, R. Review and Revision of Empirical Critical Loads and Dose-Response Relationships. In UNECE Workshop on the Review and Revision of Empirical Critical Loads and Dose-Response Relationships; RIVM: Bilthoven, The Netherlands, 2011. [Google Scholar]
- Nazare, A.I.; Stavarache, M.; Samuil, C.; Vîntu, V. The Improvement of Nardus Stricta L. Permanent Meadow from the Dorna Depression through Mineral and Organic Fertilization. Sci. Pap. Ser. A. Agron. 2024, 67, 294–301. [Google Scholar]
- Zornić, V.; Petrović, M.; Babić, S.; Lazarević, Đ.; Đurović, V.; Sokolović, D.; Tomić, D. NPK fertilizer addition effect on “Nardus stricta“ type grassland in Kopaonik Mountine. In Proceedings of the 1st International Symposium on Biotechnology (XXVIII Savetovanje o Biotehnologiji sa Međunarodnim Učešćem), Kopaonik Mountain, Serbia, 17–18 March 2023. [Google Scholar]
- Păcurar, F.; Rotar, I.; Reif, A.; Vidican, R.; Stoian, V.; Gärtner, S.M.; Allen, R.B. Impact of Climate on Vegetation Change in a Mountain Grassland—Succession and Fluctuation. Not. Bot. Horti Agrobot. Cluj-Napoca 2014, 42, 347–356. [Google Scholar] [CrossRef]
- Zhu, K.; Song, Y.; Lesage, J.C.; Luong, J.C.; Bartolome, J.W.; Chiariello, N.R.; Dudney, J.; Field, C.B.; Hallett, L.M.; Hammond, M.; et al. Rapid Shifts in Grassland Communities Driven by Climate Change. Nat. Ecol. Evol. 2024, 8, 2252–2264. [Google Scholar] [CrossRef]
- Movedi, E.; Bocchi, S.; Paleari, L.; Vesely, F.M.; Vagge, I.; Confalonieri, R. Impacts of Climate Change on Semi-Natural Alpine Pastures Productivity and Floristic Composition. Reg. Environ. Change 2023, 23, 159. [Google Scholar] [CrossRef]
- Straffelini, E.; Luo, J.; Tarolli, P. Climate Change Is Threatening Mountain Grasslands and Their Cultural Ecosystem Services. Catena 2024, 237, 107802. [Google Scholar] [CrossRef]
- Poláková, J.; Maroušková, A.; Holec, J.; Kolářová, M.; Janků, J. Changes in grassland area in lowlands and marginal uplands: Medium-term differences and potential for carbon farming. Soil Water Res. 2023, 18, 245. [Google Scholar] [CrossRef]
- Hünig, C.; Benzler, A. The Monitoring of Agricultural Land with High Natural Value in Germany; BfN-Skripten 196; Bundesamt für Naturschutz: Bonn, Germany, 2017. [Google Scholar]
- Butz, R.J.; Dennis, A.; Millar, C.I.; Westfall, R.D. Global Observation Research Initiative in Alpine Environments (GLORIA): Results from Four Target Regions in California. In AGU Fall Meeting Abstracts; Abstract B21-0705; American Geophysical Union: San Francisco, CA, USA, 2008; Volume 2008. [Google Scholar]
- Socher, S.A.; Prati, D.; Boch, S.; Müller, J.; Baumbach, H.; Gockel, S.; Hemp, A.; Schöning, I.; Wells, K.; Buscot, F.; et al. Interacting Effects of Fertilization, Mowing and Grazing on Plant Species Diversity of 1500 Grasslands in Germany Differ between Regions. Basic Appl. Ecol. 2013, 14, 126–136. [Google Scholar] [CrossRef]
- Dale, L.M.; Thewis, A.; Boudry, C.; Rotar, I.; Păcurar, F.S.; Abbas, O.; Dardenne, P.; Baeten, V.; Pfister, J.; Fernández Pierna, J.A. Discrimination of Grassland Species and Their Classification in Botanical Families by Laboratory Scale NIR Hyperspectral Imaging: Preliminary Results. Talanta 2013, 116, 149–154. [Google Scholar] [CrossRef]
- Kagan, K.; Jonak, K.; Wolińska, A. The Impact of Reduced N Fertilization Rates According to the “Farm to Fork” Strategy on the Environment and Human Health. Appl. Sci. 2024, 14, 10726. [Google Scholar] [CrossRef]
- Villa-Galaviz, E.; Smart, S.M.; Ward, S.E.; Fraser, M.D.; Memmott, J. Fertilization Using Manure Minimizes the Trade-Offs between Biodiversity and Forage Production in Agri-Environment Scheme Grasslands. PLoS ONE 2023, 18, e0290843. [Google Scholar] [CrossRef]
- Păcurar, F.; Rotar, I.; Morea, A.; Stoie, A.; Vaida, I. Management of High Nature Value Grasslands in the Apuseni Mountains. Rom. Biotechnol. Lett. 2012, 17, 7164–7174. [Google Scholar]
- Stănilă, A.L.; Dumitru, M. Soils Zones in Romania and Pedogenetic Processes. Agric. Agric. Sci. Procedia 2016, 10, 135–139. [Google Scholar] [CrossRef][Green Version]
- Păcurar, F.; Rotar, I. Metode de Studiu și Interpretare a Vegetaţiei Pajiştilor; Risoprint: Cluj-Napoca, Romania, 2014; ISBN 978-973-53-1271-8. [Google Scholar]
- Fartyal, A.; Chaturvedi, R.K.; Bargali, S.S.; Bargali, K. The Relationship between Phenological Characteristics and Life Forms within Temperate Semi-Natural Grassland Ecosystems in the Central Himalaya Region of India. Plants 2025, 14, 835. [Google Scholar] [CrossRef] [PubMed]
- Andreatta, D.; Bachofen, C.; Dalponte, M.; Klaus, V.H.; Buchmann, N. Extracting Flowering Phenology from Grassland Species Mixtures Using Time-Lapse Cameras. Remote Sens. Environ. 2023, 298, 113835. [Google Scholar] [CrossRef]
- Li, X.; Pearson, D.E.; Ortega, Y.K.; Jiang, L.; Wang, S.; Gao, Q.; Wang, D.; Hautier, Y.; Zhong, Z. Nitrogen Inputs Suppress Plant Diversity by Overriding Consumer Control. Nat. Commun. 2025, 16, 5855. [Google Scholar] [CrossRef] [PubMed]
- Caldararu, S.; Rolo, V.; Stocker, B.D.; Gimeno, T.E.; Nair, R. Ideas and Perspectives: Beyond Model Evaluation–Combining Experi-Ments and Models to Advance Terrestrial Ecosystem Science. Biogeosciences 2023, 20, 3637–3649. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- McCune, B.; Mefford, M. PC-ORD: Multivariate Analysis of Ecological Data, version 6; MjM Software: Gleneden Beach, OR, USA, 2011. [Google Scholar]
- Hao, Z.; Zhang, J.; Song, B. Assessing Biological Dissimilarities between Five Forest Plots in China Using Species Abun-Dance and Phylogenetic Information. For. Ecosyst. 2019, 6, 47. [Google Scholar] [CrossRef]
- Campos, R.; Fontana, V.; Losapio, G.; Moretti, M. How Cities Impact Diversity and Ecological Networks: A Functional and Phylogenetic Perspective. Urban Ecosyst. 2024, 27, 1359–1373. [Google Scholar] [CrossRef]
- Rocha-Filho, L.C.; Santos, J.L.; Pereira, B.A. The Initial Impact of a Hydroelectric Reservoir on Tree Community Structure. Forests 2025, 16, 1236. [Google Scholar] [CrossRef]
- Sun, Y.; Tao, C.; Deng, X.; Liu, H.; Shen, Z.; Liu, Y.; Li, R.; Shen, Q.; Geisen, S. Organic Fertilization Enhances the Resistance and Resilience of Soil Microbial Communities under Extreme Drought. J. Adv. Res. 2023, 47, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jaunatre, R.; Buisson, E.; Leborgne, E.; Dutoit, T. Soil Fertility and Landscape Surrounding Former Arable Fields Drive the Ecological Resilience of Mediterranean Dry Grassland Plant Communities. Front. Ecol. Evol. 2023, 11, 1210591. [Google Scholar] [CrossRef]
- Manu, M.; Băncilă, R.I.; Onete, M. Soil Fauna-Indicators of Ungrazed Versus Grazed Grassland Ecosystems in Romania. Diversity 2025, 17, 323. [Google Scholar] [CrossRef]
- Peck, J. Multivariate Analysis for Community Ecologists: Step by Step Using PC-ORD; MjM Software Design: Gleneden Beach, OR, USA, 2010; ISBN 978-0972129018. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Yan, H.; Li, F.; Liu, G. Diminishing Influence of Negative Relationship between Species Richness and Evenness on the Modeling of Grassland α-Diversity Metrics. Front. Ecol. Evol. 2023, 11, 1108739. [Google Scholar] [CrossRef]
- Sang, Y.; Gu, H.; Meng, Q.; Men, X.; Sheng, J.; Li, N.; Wang, Z. An Evaluation of the Performance of Remote Sensing Indices as an Indication of Spatial Variability and Vegetation Diversity in Alpine Grassland. Remote Sens. 2024, 16, 4726. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Oduor, A.M.; Kleunen, M.; Liu, Y. Diversity and Productivity of a Natural Grassland Decline with the Number of Global Change Factors. J. Plant Ecol. 2025, 17, 112–124. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; ISBN 978-0-632-05633-0. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).