

Combined Impact of Canada Goldenrod Invasion and Soil Microplastic Contamination on Seed Germination and Root Development of Wheat: Evaluating the Legacy of Toxicity


 ,
,  ,
,  ,
,  , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Design
2.2. Seed Germination Rate and Plant Phenotypic Properties Measurement
2.3. Seedling Vigor Index
2.4. Statistical Analysis
3. Results
3.1. The Individual and Combined Effect of S. canadensis Invasion Legacy Effects and Soil Microplastic Contamination on Wheat Seed Germination
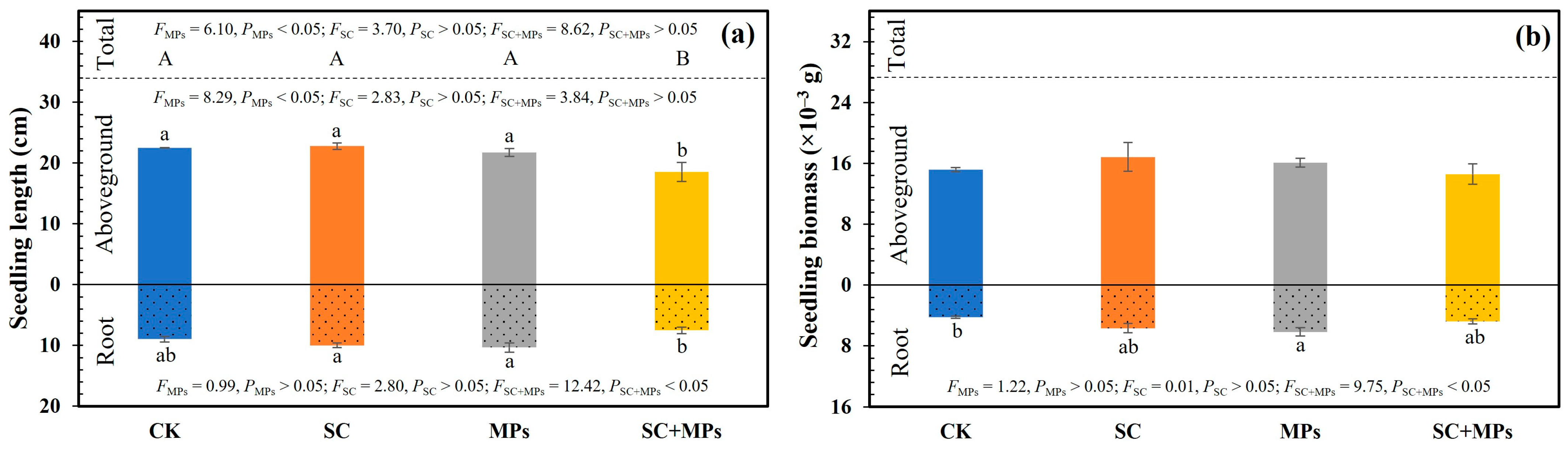
3.2. The Individual and Combined Effect of S. canadensis Invasion Legacy Effects and Soil Microplastic Contamination on the Development of Wheat Seedlings
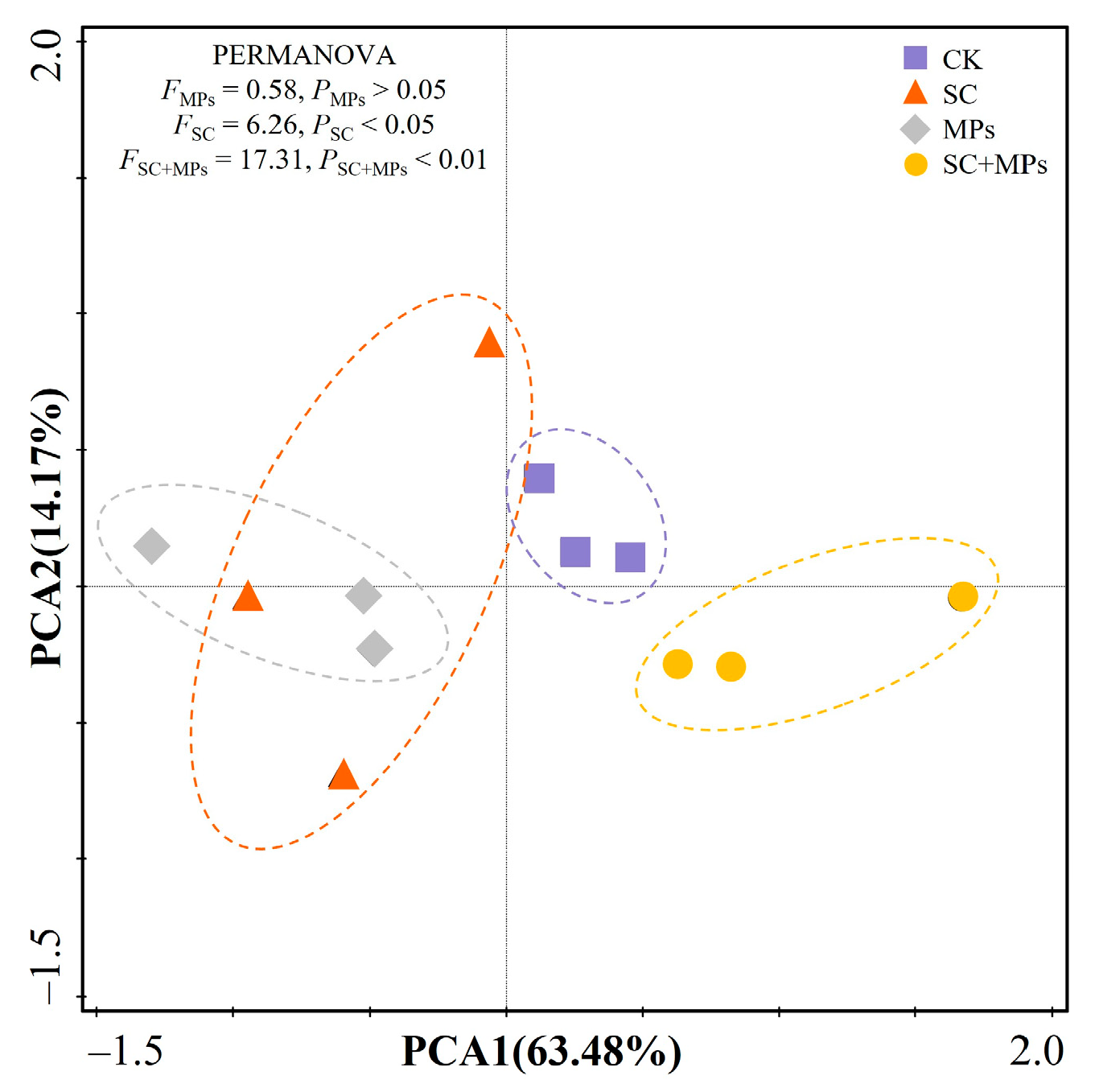
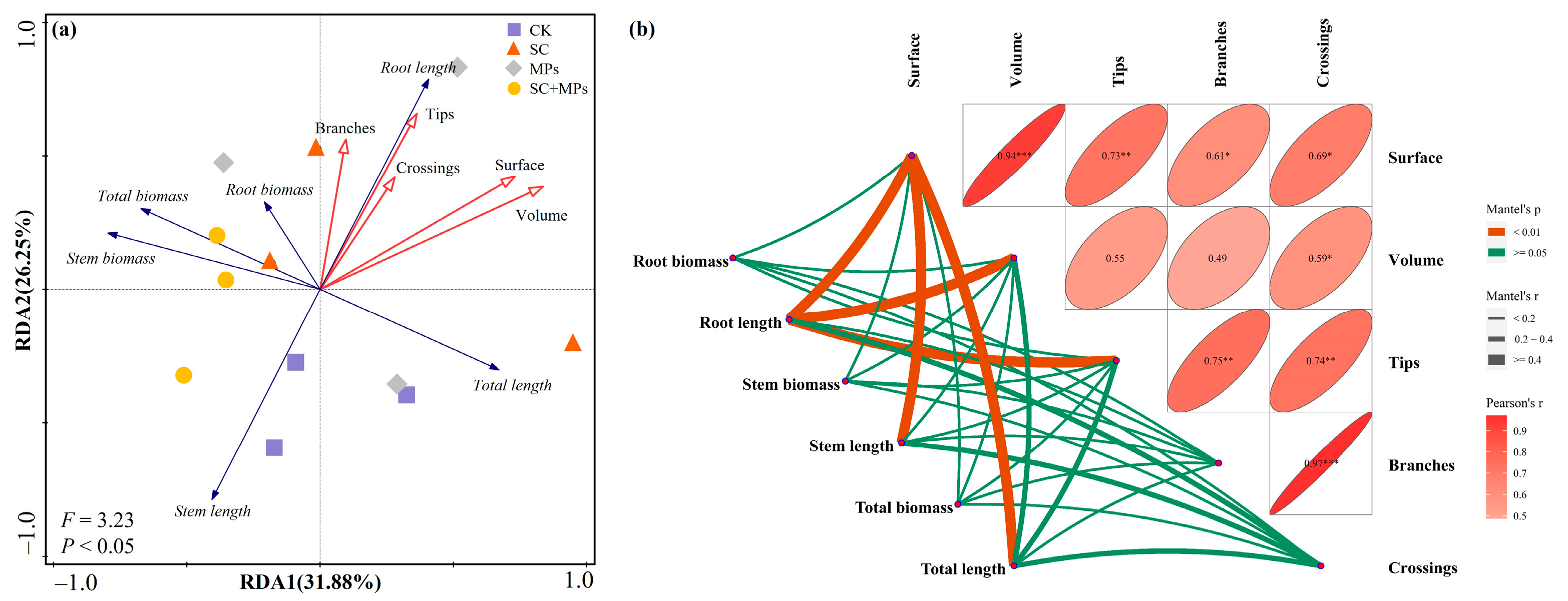
3.3. Effects of Plant Phenotypes on the Length and Biomass of Seedling and Predictors of SVI
4. Discussion
4.1. Impact of Legacy Effects of S. canadensis Invasion and Soil Microplastic Contaminants on Wheat Seed Germination
4.2. Individual Effects of S. canadensis Invasion Legacy Effects or Soil Microplastic Contaminants on Wheat Seedlings
4.3. Interaction Effects of S. canadensis Invasion Legacy Effects and Soil Microplastic Contaminants on Wheat Seedlings
4.4. Implication for Agricultural Sustainability and Management
4.5. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef] [PubMed]
- de Souza Machado, A.A.; Kloas, W.; Zarfl, C.; Hempel, S.; Rillig, M.C. Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; An, S.; Kim, L.; Byeon, Y.M.; Lee, J.; Choi, M.J.; An, Y.J. Translocation and Chronic Effects of Microplastics on Pea Plants (Pisum sativum) in Copper-Contaminated Soil. J. Hazard. Mater. 2022, 436, 129194. [Google Scholar] [CrossRef] [PubMed]
- Tariq, M.; Iqbal, B.; Khan, I.; Khan, A.R.; Jho, E.H.; Salam, A.; Zhou, H.; Zhao, X.; Li, G.; Du, D. Microplastic contamination in the agricultural soil: Mitigation strategies, heavy metals contamination, and impact on human health—A review. Plant Cell Rep. 2024, 43, 65. [Google Scholar] [CrossRef]
- Kida, M.; Ziembowicz, S.; Koszelnik, P. Decomposition of microplastics: Emission of harmful substances and greenhouse gases in the environment. J. Environ. Chem. Eng. 2023, 11, 109047. [Google Scholar] [CrossRef]
- Khan, A.; Jie, Z.; Wang, J.; Nepal, J.; Ullah, N.; Zhao, Z.Y.; Wang, P.Y.; Ahmad, W.; Khan, A.; Wang, W. Ecological risks of microplastics contamination with green solutions and future perspectives. Sci. Total Environ. 2023, 899, 165688. [Google Scholar] [CrossRef] [PubMed]
- Khalid, N.; Aqeel, M.; Noman, A. Microplastics could be a threat to plants in terrestrial systems directly or indirectly. Environ. Pollut. 2020, 267, 115653. [Google Scholar] [CrossRef] [PubMed]
- Javed, Q.; Sun, J.; Rutherford, S.; Li, J.; Iqbal, B.; Xiang, Y.; Ren, G.; He, F.; Pan, l.; Bo, Y.; et al. Soil pollution and the invasion of congener Sphagneticola in crop lands. J. Environ. Manag. 2023, 340, 118013. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Khan, A.; Ullah, A.; Ul Haq, S.I.; Hassan, N.; Iqbal, B.; Ahmad, N.; Mahmoud, E.A.; Elansary, H.O. Insights concerning advancing the agroecological sustainability of salinity tolerance through proteomics profiling of hexaploid wheat (Triticum aestivum L.). S. Afr. J. Bot. 2023, 158, 142–148. [Google Scholar] [CrossRef]
- Colzi, I.; Renna, L.; Bianchi, E.; Castellani, M.B.; Coppi, A.; Pignattelli, S.; Loppi, S.; Gonnelli, C. Impact of microplastics on growth photosynthesis and essential elements in Cucurbita pepo L. J. Hazard. Mater. 2022, 423, 127238. [Google Scholar] [CrossRef]
- Zhang, Z.; Cui, Q.; Chen, L.; Zhu, X.; Zhao, S.; Duan, C.; Zhang, X.; Song, D.; Fang, L. A critical review of microplastics in the soil-plant system: Distribution, uptake, phytotoxicity and prevention. J. Hazard. Mater. 2022, 424, 127750. [Google Scholar] [CrossRef] [PubMed]
- de Souza Machado, A.A.; Lau, C.W.; Kloas, W.; Bergmann, J.; Bachelier, J.B.; Faltin, E.; Becker, R.; Görlich, A.S.; Rillig, M.C. Microplastics Can Change Soil Properties and Affect Plant Performance. Environ. Sci. Technol. 2019, 53, 6044–6052. [Google Scholar] [CrossRef] [PubMed]
- Lozano, Y.M.; Rillig, M.C. Effects of microplastic fibers and drought on plant communities. Environ. Sci. Technol. 2020, 54, 6166–6173. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Rillig, M.C. Legacy effect of microplastics on plant–soil feedbacks. Front. Plant Sci. 2022, 13, 965576. [Google Scholar] [CrossRef]
- Wang, S.; Wei, M.; Wu, B.; Cheng, H.; Wang, C. Combined nitrogen deposition and Cd stress antagonistically affect the allelopathy of invasive alien species Canada goldenrod on the cultivated crop lettuce. Sci. Hortic. 2020, 261, 108955. [Google Scholar] [CrossRef]
- Liu, M.T.; Lu, S.B.; Song, Y.; Lei, L.L.; Hu, J.N.; Lv, W.W.; Zhou, W.Z.; Cao, C.J.; Shi, H.H.; Yang, X.F. Microplastic and mesoplastic pollution in farmland soils in suburbs of Shanghai, China. Environ. Pollut. 2018, 242, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhao, Y.; Padilla-Zakour, O.; Yang, G. Polyphenols, antioxidant and antimicrobial activities of leaf and bark extracts of Solidago canadensis L. Ind. Crops Prod. 2015, 74, 803–809. [Google Scholar] [CrossRef]
- Zhang, Z.; Bhowmik, P.C.; Suseela, V. Effect of soil carbon amendments in reversing the legacy effect of plant invasion. J. Appl. Ecol. 2021, 58, 181–191. [Google Scholar] [CrossRef]
- Suzuki, K.; Tungalag, R.; Narantsetseg, A.; Tsendeekhuu, T.; Shinoda, M.; Yamanaka, N.; Kamijo, T. Composition, distribution and environmental drivers of Mongolian rangeland plant communities. J. Plant Ecol. 2023, 16, rtac100. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, Y.; Wu, B.; Wang, S.; Wei, M.; Du, D.; Wang, C. Allelopathy of three Compositae invasive alien species on indigenous Lactuca sativa L. enhanced under Cu and Pb pollution. Sci. Hortic. 2020, 267, 109323. [Google Scholar] [CrossRef]
- Adomako, M.O.; Ning, L.; Tang, M.; Du, D.L.; van Kleunen, M.; Yu, F.H. Diversity- and density-mediated allelopathic effects of resident plant communities on invasion by an exotic plant. Plant Soil 2019, 440, 581–592. [Google Scholar] [CrossRef]
- Hannula, S.E.; Heinen, R.; Huberty, M.; Steinauer, K.; Long, J.R.D.; Jongen, R.; Bezemer, T.M. Persistence of plant-mediated microbial soil legacy effects in soil and inside roots. Nat. Commun. 2021, 12, 5686. [Google Scholar] [CrossRef] [PubMed]
- Lacalle, R.G.; Becerril, J.M.; Garbisu, C. Biological methods of polluted soil remediation for an effective economically-optimal recovery of soil health and ecosystem services. J. Environ. Sci. Public Health 2020, 4, 112–133. [Google Scholar] [CrossRef]
- Westerveld, S.; McKeown, A.; McDonald, M.R. (219) Nitrogen Relationships to Yield and Quality of Carrots Grown on Mineral and Organic Soil in Ontario. HortScience 2005, 40, 1073D. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, H.; Wang, C.; Cheng, J.; Qiang, S. A comparative study reveals the key biological traits causing bioinvasion differences among four alien species of genus Veronica in China. J. Plant Ecol. 2023, 16, rtac068. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Y.; Xiao, M.; Zhao, C.; Yao, H. A meta-analysis of film mulching cultivation effects on soil organic carbon and soil greenhouse gas fluxes. Catena 2021, 206, 105483. [Google Scholar] [CrossRef]
- Lee, T.Y.; Kim, L.; Kim, D.; An, S.; An, Y.J. Microplastics from shoe sole fragments cause oxidative stress in a plant (Vigna radiata) and impair soil environment. J. Hazard. Mater. 2022, 429, 128306. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, A.A.; Farooq, M.; Nawaz, A.; Yadav, L.; Chauhan, B.S.; Adkins, S. Impact of invasive plant species on the livelihoods of farming households: Evidence from Parthenium hysterophorus invasion in rural Punjab, Pakistan. Biol. Invasions 2019, 21, 3285–3304. [Google Scholar] [CrossRef]
- Li, B.; Ge, T.; Hill, P.W.; Jones, D.L.; Zhu, Z.; Zhran, M.; Wu, J. Experimental strategies to measure the microbial uptake and mineralization kinetics of dissolved organic carbon in soil. Soil Ecol. Lett. 2020, 2, 180–187. [Google Scholar] [CrossRef]
- Guo, M.; Zhao, F.; Tian, L.; Ni, K.; Lu, Y.; Borah, P. Effects of polystyrene microplastics on the seed germination of herbaceous ornamental plants. Sci. Total Environ. 2022, 809, 151100. [Google Scholar] [CrossRef] [PubMed]
- Bosker, T.; Bouwman, L.J.; Brun, N.R.; Behrens, P.; Vijver, M.G. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum. Chemosphere 2019, 226, 774–781. [Google Scholar] [CrossRef]
- Lian, J.; Wu, J.; Zeb, A.; Zheng, S.; Ma, T.; Peng, F.; Tang, J.; Liu, W. Do polystyrene nanoplastics affect the toxicity of cadmium to wheat (Triticum aestivum L.)? Environ. Pollut. 2020, 263, 114498. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jiang, K.; Zhou, J.; Wu, B. Solidago canadensis invasion affects soil N-fixing bacterial communities in heterogeneous landscapes in urban ecosystems in East China. Sci. Total Environ. 2018, 631, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Tang, S.C.; Pan, Y.M.; Wei, C.Q.; Lü, S.H. Increased precipitation magnifies the effects of N addition on performance of invasive plants in subtropical native communities. J. Plant Ecol. 2022, 15, 473–484. [Google Scholar] [CrossRef]
- Yuan, Y.G.; Jin, H.F.; Li, J.M. Effects of latitude and soil microbes on the resistance of invasive Solidago canadensis to its co-evolved insect herbivore Corythucha marmorata. J. Plant Ecol. 2022, 15, 549–560. [Google Scholar] [CrossRef]
- Li, G.; Cui, X.; Tariq, M.; Khan, I.; Khan, A.R.; Obaid, S.A.; Ansari, M.J.; Zhou, H.; Iqbal, B.; Zhao, X. Microplastic and cadmium contamination: Impact on the soil by inhibiting the growth of pak choi (Brassica rapa subsp. chinensis). Process Saf. Environ. Prot. 2024, 189, 714–727. [Google Scholar] [CrossRef]
- Pidatala, V.R.; Li, K.F.; Sarkar, D.; Ramakrishna, W.; Datta, R. Identification of biochemical pathways associated with lead tolerance and detoxification in Chrysopogon zizanioides L. Nash (Vetiver) by Metabolic profiling. Environ. Sci. Technol. 2016, 50, 2530–2537. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2021, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Feng, X.; Liu, Y.; Cui, W.; Sun, Y.; Zhang, S.; Wang, F. Effects of microplastics and carbon nanotubes on soil geochemical properties and bacterial communities. J. Hazard. Mater. 2022, 433, 128826. [Google Scholar] [CrossRef]
- Pan, L.; He, F.; Liang, Q.; Bo, Y.; Lin, X.; Javed, Q.; Ullah, M.S.; Sun, J. Allelopathic Effects of caffeic acid and its derivatives on seed germination and growth competitiveness of native plants (Lantana indica) and invasive plants (Solidago canadensis). Agriculture 2023, 13, 1719. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Lehnert, T.; Linck, L.T.; Lehmann, A.; Rillig, M.C. Microplastic shape polymer type and concentration affect soil properties and plant biomass. Front. Plant Sci. 2021, 12, 616645. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Y.; Xiang, L.; Redmile-Gordon, M.; Gu, C.; Yang, X.; Jiang, X.; Barceló, D. Perspectives on ecological risks of microplastics and phthalate acid esters in crop production systems. Soil Ecol. Lett. 2022, 4, 97–108. [Google Scholar] [CrossRef]
- Li, G.; Tang, Y.; Son, Y.; Zhao, X.; Iqbal, B.; Khan, K.Y.; Guo, R.; Yin, W.; Zhao, X.; Du, D. Divergent responses in microbial metabolic limitations and carbon use efficiency to variably sized polystyrene microplastics in soil. Land Degrad. Dev. 2024, 35, 2658–2671. [Google Scholar] [CrossRef]
- Liao, Y.C.; Nazygul, J.; Li, M.; Wang, X.L.; Jiang, L.J. Effects of microplastics on the growth physiology and biochemical characteristics of wheat (Triticum aestivum). Acta Sci. Circumstantiae 2019, 40, 4661–4667. [Google Scholar] [CrossRef]
- Qi, Y.; Yang, X.; Pelaez, A.M.; Lwanga, E.H.; Beriot, N.; Gertsen, H.; Garbeva, P.; Geissen, V. Macro- and micro-plastics in soil-plant system: Effects of plastic mulch film residues on wheat (Triticum aestivum) growth. Sci. Total Environ. 2018, 645, 1048–1056. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.; Iavicoli, I.; Calabrese, E.J. The two faces of nanomaterials: A quantification of hormesis in algae and plants. Environ. Int. 2019, 131, 105044. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Liang, J.; Zeng, G.; Li, X.; Chen, M.; Jiang, L.; Xing, W.; Tang, N. Responses of seeds of typical Brassica crops to tetracycline stress: Sensitivity difference and source analysis. Ecotoxicol. Environ. Saf. 2019, 184, 109597. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hou, H.; Liu, Y.; Yin, S.; Bian, S.; Liang, S.; Wan, C.; Yuan, S.; Xiao, K.; Liu, B.; et al. Microplastics affect rice (Oryza sativa L.) quality by interfering metabolite accumulation and energy expenditure pathways: A field study. J. Hazard. Mater. 2022, 422, 126834. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Pei, L.; Zhao, Y.; Shan, J.; Zheng, X.; Xu, G.; Sun, Y.; Wang, F. Effects of microplastics and nitrogen deposition on soil multifunctionality particularly C and N cycling. J. Hazard. Mater. 2023, 451, 131152. [Google Scholar] [CrossRef]
- Zang, H.; Zhou, J.; Marshall, M.R.; Chadwick, D.R.; Wen, Y.; Jones, D.L. Microplastics in the agroecosystem: Are they an emerging threat to the plant-soil system. Soil Biol. Biochem. 2020, 148, 107926. [Google Scholar] [CrossRef]
- Liu, Y.; Xiao, M.; Shahbaz, M.; Hu, Z.; Zhu, Z.; Lu, S.; Yu, Y.; Yao, H.; Chen, J.; Ge, T. Microplastics in soil can increase nutrient uptake by wheat. J. Hazard. Mater. 2022, 438, 129547. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam Mahadevan, I.; Harsha Vardhan, M.K.; Rejith Kumar, R.S.; Rohinth, M.; Tawfeeq Ahmed, Z.H.; Prakash, P.; Kumar, J.A. Biotechnological interventions for monitoring and mitigating microplastic pollution and development of alternatives to single-use plastics. Environ. Qual. Manag. 2024, 34, e22186. [Google Scholar] [CrossRef]
- Yuan, L.; Zhou, L.; Li, J. Effect of microplastics on the allelopathic effects of native and invasive plants on co-occurring invaders. Front. Plant Sci. 2024, 15, 1425815. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.F.; Cheng, W.C.; Wang, L.; Xie, Y.X.; Qin, P.; Shi, C. Immobilizing lead in aqueous solution and loess soil using microbially induced carbonate/phosphate precipitation (MICP/MIPP) under harsh pH environments. J. Hazard. Mater. 2024, 480, 135884. [Google Scholar] [CrossRef]
- Ma, T.; Parker, T.; Fetcher, N.; Unger, S.L.; Gewirtzman, J.; Moody, M.L.; Tang, J. Leaf and root phenology and biomass of Eriophorum vaginatum in response to warming in the Arctic. J. Plant Ecol. 2022, 15, 1091–1105. [Google Scholar] [CrossRef]
- Zubek, S.; Majewska, M.L.; Kapusta, P.; Stefanowicz, A.M.; Blaszkowski, J.; Rożek, K.; Stanek, M.; Karpowicz, F.; Zalewska-Gałosz, J. Solidago canadensis invasion in abandoned arable fields induces minor changes in soil properties and does not affect the performance of subsequent crops. Land Degrad. Dev. 2019, 31, 334–345. [Google Scholar] [CrossRef]
- Wang, C.; Xiao, H.; Zhao, L.; Liu, J.; Wang, L.; Zang, F.; Shi, Y.; Du, D. The allelopathic effects of invasive plant Solidago canadensis on seed germination and growth of Lactuca sativa enhanced by different types of acid deposition. Ecotoxicology 2016, 25, 555–562. [Google Scholar] [CrossRef]
- Pignattelli, S.; Broccoli, A.; Renzi, M. Physiological responses of garden cress (L. sativum) to different types of microplastics. Sci. Total Environ. 2020, 727, 138609. [Google Scholar] [CrossRef] [PubMed]
- Pignattelli, S.; Broccoli, A.; Piccardo, M.; Felline, S.; Terlizzi, A.; Renzi, M. Short-term physiological and biometrical responses of Lepidium sativum seedlings exposed to PET-made microplastics and acid rain. Ecotoxicol. Environ. Saf. 2021, 208, 111718. [Google Scholar] [CrossRef]
- Pignattelli, S.; Broccoli, A.; Piccardo, M.; Terlizzi, A.; Renzi, M. Effects of polyethylene terephthalate (PET) microplastics and acid rain on physiology and growth of Lepidium sativum. Environ. Pollut. 2021, 282, 116997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Luo, X.S.; Xu, J.; Yao, X.; Fan, J.; Mao, Y.; Song, Y.; Yang, J.; Pan, J.; Khattak, W.A. Dry–wet cycle changes the influence of microplastics (MPs) on the antioxidant activity of lettuce and the rhizospheric bacterial community. J. Soils Sediments 2023, 23, 2189–2201. [Google Scholar] [CrossRef]
- Xue, Z.F.; Cheng, W.C.; Rahman, M.M.; Wang, L.; Xie, Y.X. Immobilization of Pb(II) by Bacillus megaterium-based microbial-induced phosphate precipitation (MIPP) considering bacterial phosphorolysis ability and Ca-mediated alleviation of lead toxicity. Environ. Pollut. 2024, 355, 124229. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Tang, Y.; Xie, H.; Iqbal, B.; Wang, Y.; Dong, K.; Zhao, X.; Kim, H.-J.; Du, D.; Xiao, C. Combined Impact of Canada Goldenrod Invasion and Soil Microplastic Contamination on Seed Germination and Root Development of Wheat: Evaluating the Legacy of Toxicity. Plants 2025, 14, 181. https://doi.org/10.3390/plants14020181
Li G, Tang Y, Xie H, Iqbal B, Wang Y, Dong K, Zhao X, Kim H-J, Du D, Xiao C. Combined Impact of Canada Goldenrod Invasion and Soil Microplastic Contamination on Seed Germination and Root Development of Wheat: Evaluating the Legacy of Toxicity. Plants. 2025; 14(2):181. https://doi.org/10.3390/plants14020181
Chicago/Turabian StyleLi, Guanlin, Yi Tang, Hongliang Xie, Babar Iqbal, Yanjiao Wang, Ke Dong, Xin Zhao, Hyun-Jun Kim, Daolin Du, and Chunwang Xiao. 2025. "Combined Impact of Canada Goldenrod Invasion and Soil Microplastic Contamination on Seed Germination and Root Development of Wheat: Evaluating the Legacy of Toxicity" Plants 14, no. 2: 181. https://doi.org/10.3390/plants14020181
APA StyleLi, G., Tang, Y., Xie, H., Iqbal, B., Wang, Y., Dong, K., Zhao, X., Kim, H.-J., Du, D., & Xiao, C. (2025). Combined Impact of Canada Goldenrod Invasion and Soil Microplastic Contamination on Seed Germination and Root Development of Wheat: Evaluating the Legacy of Toxicity. Plants, 14(2), 181. https://doi.org/10.3390/plants14020181