

Identification and Genetic Analysis of Downy Mildew Resistance in Intraspecific Hybrids of Vitis vinifera L.


Abstract
1. Introduction
2. Results
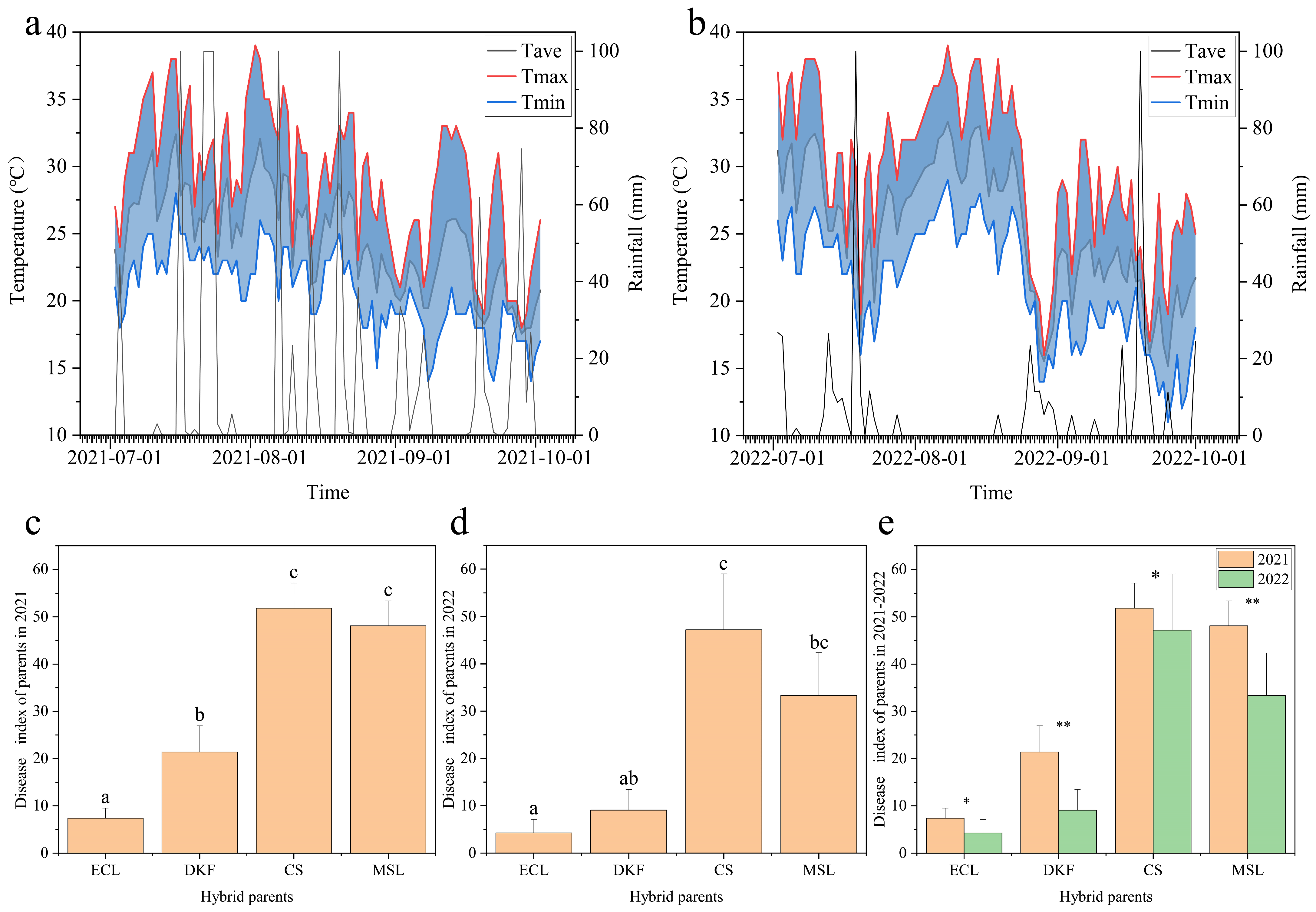
2.1. Identification of Disease Resistance of Hybrid Parents
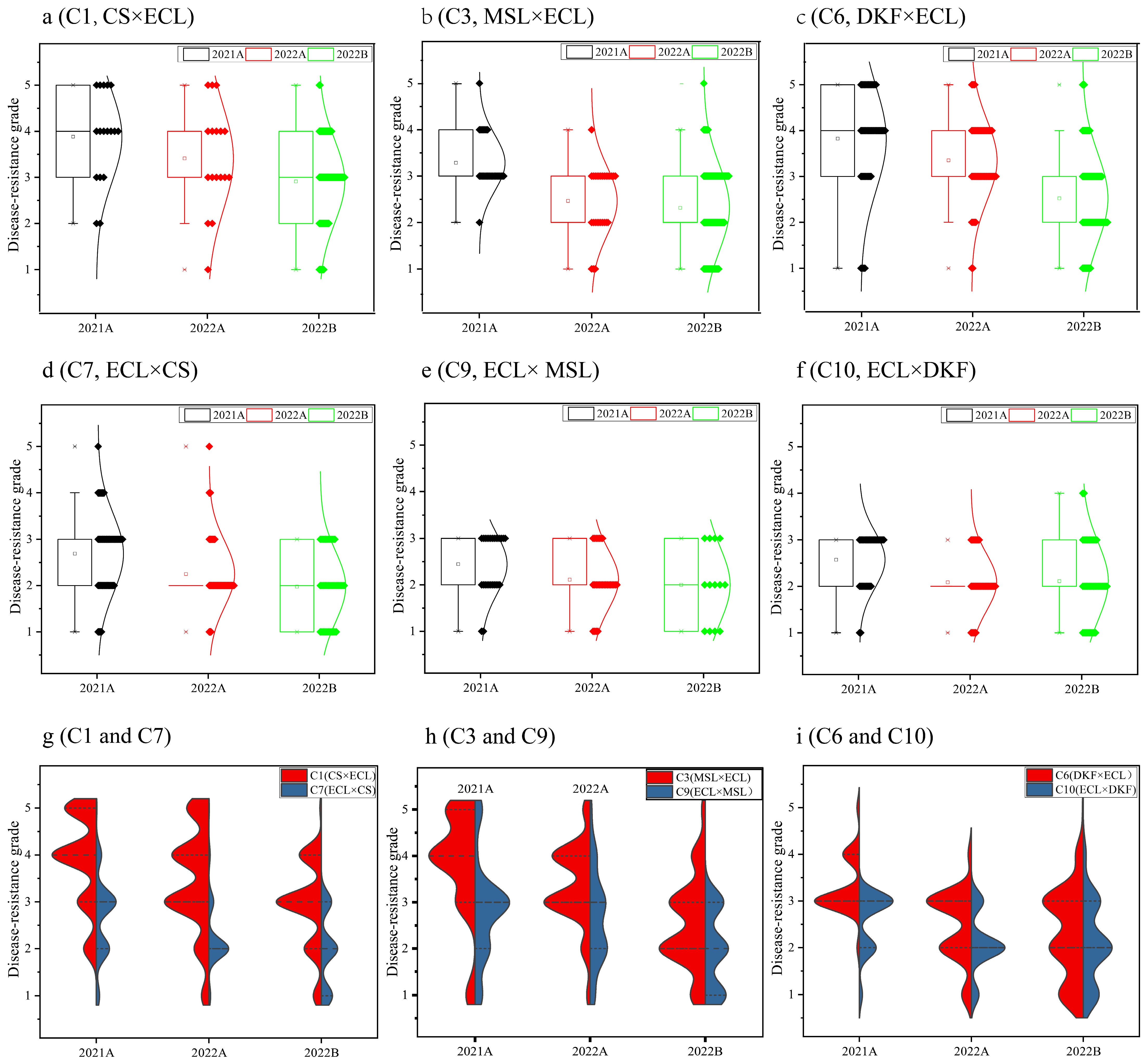
2.2. Identification and Genetic Analysis of Disease Resistance in V. vinifera Hybrids
2.3. Regression Analysis and Comprehensive Evaluation of Disease Resistance Traits
3. Discussion
3.1. Evaluation of Downy Mildew Resistance in V. vinifera
3.2. Genetic Characteristics of Disease Resistance in V. vinifera Hybrid Progeny
3.3. Relationship Between Disease Resistance Traits
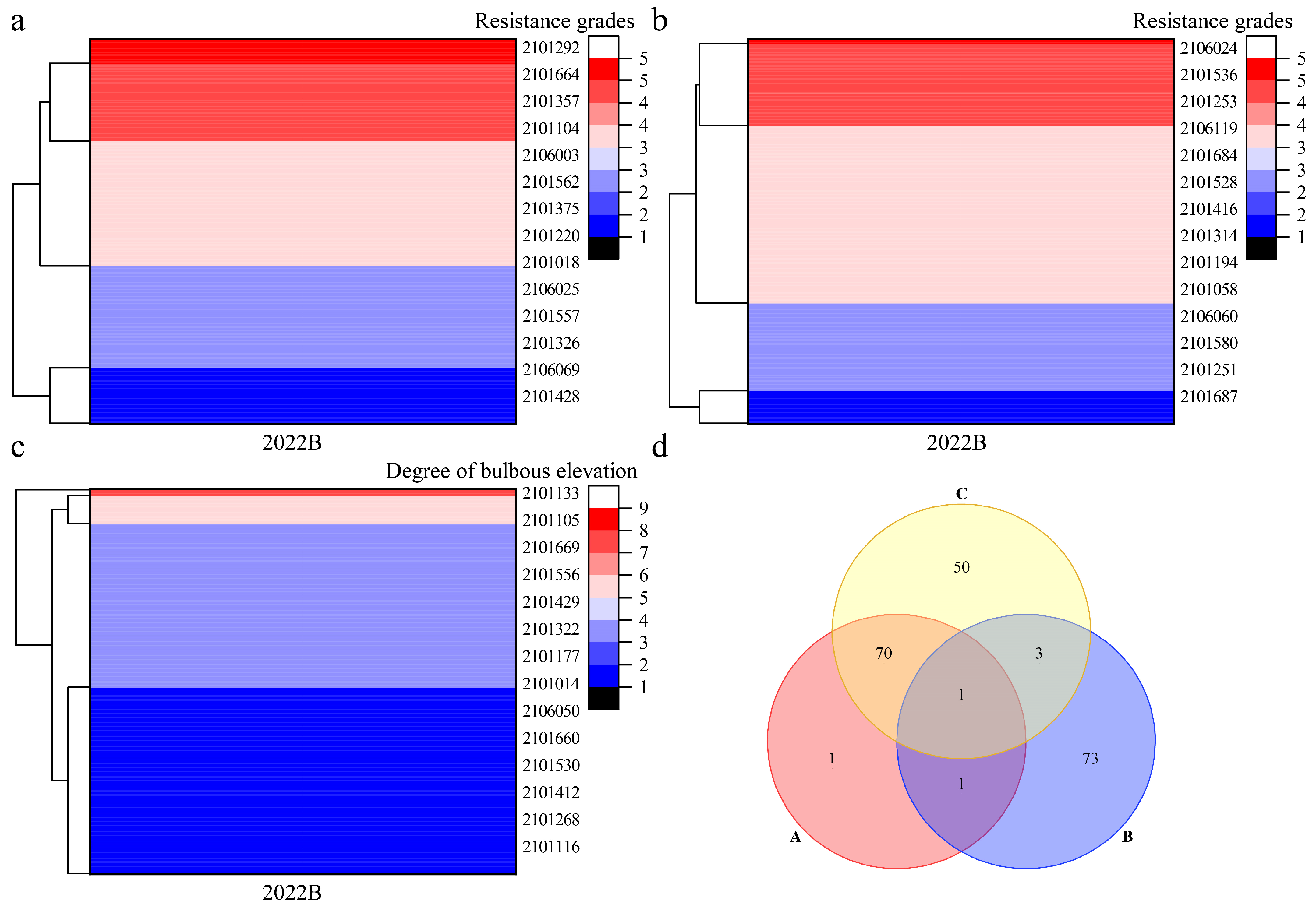
3.4. Screening of Hybrid Progeny with High Downy Mildew Resistance
4. Materials and Methods
4.1. Materials
4.2. Investigation of the Degree of Leaf Bulbous Elevation
4.3. Identification and Grading of Downy Mildew
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Gil-Muñoz, R.; Moreno-Olivares, J.D.; Paladines-Quezada, D.F.; Bleda-Sánchez, J.A.; Cebrían-Pérez, A.; Giménez-Bañón, M.J.; Fernández-Fernández, J.I. Characterization of Anthocyanins From Intraspecific Crosses of Monastrell With Other Premium Varieties. Front. Nutr. 2021, 8, 664515. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Mechanism of Kaolin Particle Film Regulating Grape Leaves Againet Downy Mildew. Ph.D. Thesis, Northwest A&F University, Yangling, China, 2022. [Google Scholar]
- Wang, Y.; Xue, T.; Han, X.; Guan, L.; Zhang, L.; Wang, H.; Li, H. Kaolin Particle Film Affects Grapevine Berry Quality in cv. Meili in Humid Climate Conditions. HortScience 2020, 55, 1987–2000. [Google Scholar] [CrossRef]
- Nenko, N.; Ilyina, I.; Kiseleva, G.; Shalyaho, T. Study of grape resistance to stressors of the winter period in the south of Russia. Agric. Food 2016, 4, 260–266. Available online: https://www.scientific-publications.net/en/article/1001040/ (accessed on 1 March 2025).
- Atucha, A.; Hedtcke, J.; Workmaster, B. Evaluation of Cold-climate interspecific Hybrid Wine Grape Cultivars for the Upper Midwest. J. Am. Pomol. Soc. 2018, 72, 80–93. [Google Scholar]
- Nenko, N.; Ilyina, I.; Kiseleva, G.; Yablonskaya, E. Low-Temperature Stress Tolerance of Grapevine Varieties of Different Ecological and Geographical Origin. Proc. Latv. Acad. Sciences. Sect. B Nat. Exact Appl. Sci. 2018, 73, 56–65. [Google Scholar] [CrossRef]
- Schrader, J.A.; Cochran, D.R.; Domoto, P.A.; Nonnecke, G.R. Phenology and Winter Hardiness of Cold-climate Grape Cultivars and Advanced Selections in Iowa Climate. HortTechnology 2019, 29, 906–922. [Google Scholar] [CrossRef]
- Egorov, E.A. Grape breeding is a key link in the development of the grapes and wine-making industry. Vavilovskii Zhurnal Genet. I Sel. 2021, 25, 408–413. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.-S.; Yang, X.-Q.; Wang, Z.-Y.; Ma, L.; Tu, H.-X.; Ma, Y.; Zhou, J.-T.; Zhang, J.; Wang, H.; et al. Inheritance analysis of fruit-related traits in Chinese cherry [Cerasus pseudocerasus (Lindl.) G.Don] breeding progenies. Sci. Hortic. 2023, 307, 111519. [Google Scholar] [CrossRef]
- Park, Y.-S.; Heo, J.-Y.; Bang, S.-B. ‘Black Eye’: A Unique Korean Grape Variety Developed from a Cross between ‘Muscat Bailey (V. labrusca × V. vinifera) A’ and Korean Wild Grape (V. amurensis). HortScience 2017, 52, 786–788. [Google Scholar] [CrossRef]
- Teissedre, P.-L. Composition of grape and wine from resistant vines varieties. OENO One 2018, 52, 211–217. [Google Scholar] [CrossRef]
- Tetali, S.; Karkamkar, S.; Phalake, S. Grape Breeding for powdery mildew resistance. Indian J. Hortic. 2018, 75, 541. [Google Scholar] [CrossRef]
- Salmon, J.-M.; Ojeda, H.; Escudier, J.-L. Disease resistant grapevine varieties and quality: The case of Bouquet varieties. OENO One 2018, 52, 225–230. [Google Scholar] [CrossRef]
- Clark, M.; Hemstad, P.; Luby, J. ‘Itasca’ Grapevine, a New Cold-hardy Hybrid for White Wine Production. HortScience 2017, 52, 649–651. [Google Scholar] [CrossRef]
- Manso-Martínez, C.; Sáenz-Navajas, M.-P.; Hernández, M.; Menéndez, C. Sensory profiling and quality assessment of wines derived from Graciano × Tempranillo selections. LWT-Food Sci. Technol. 2020, 127, 109394. [Google Scholar] [CrossRef]
- Yang, Y.; Cuenca, J.; Wang, N.; Liang, Z.; Sun, H.; Gutierrez, B.; Xi, X.; Arro, J.; Wang, Y.; Fan, P.; et al. A key ‘foxy’ aroma gene is regulated by homology-induced promoter indels in the iconic juice grape ‘Concord’. Hortic. Res. 2020, 7, 67. [Google Scholar] [CrossRef]
- Bowers, J.E.; Meredith, C.P. The parentage of a classic wine grape, Cabernet Sauvignon. Nat. Genet. 1997, 16, 84–87. [Google Scholar] [CrossRef]
- Roach, M.J.; Johnson, D.L.; Bohlmann, J.; van Vuuren, H.J.J.; Jones, S.J.M.; Pretorius, I.S.; Schmidt, S.A.; Borneman, A.R. Population sequencing reveals clonal diversity and ancestral inbreeding in the grapevine cultivar Chardonnay. PLoS Genet. 2018, 14, e1007807. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Gates, M.J.; Lavin, E.H.; Acree, T.E.; Sacks, G.L. Comparison of odor-active compounds in grapes and wines from Vitis vinifera and non-foxy American grape species. J. Agric. Food Chem. 2011, 59, 10657–10664. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhang, L.; Zhao, K.; Niu, R.; Zhai, H.; Zhang, J. VaERD15, a Transcription Factor Gene Associated with Cold-Tolerance in Chinese Wild Vitis amurensis. Front. Plant Sci. 2017, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Zhang, G.; Zhao, F.; Zhu, D.; Fan, X.; Zhang, Z.; Liu, X. VvBAP1 Is Involved in Cold Tolerance in Vitis vinifera L. Front. Plant Sci. 2018, 9, 726. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Z. Minor resistant genes accumulation by replacement in the grape-powdery mildew pathosystem. Acta Bot. Boreali-Occident. Sin. 1995, 15, 120–124. [Google Scholar]
- Wang, Z.-L.; Xue, T.-T.; Gao, F.-F.; Zhang, L.; Han, X.; Wang, Y.; Hui, M.; Wu, D.; Li, H.; Wang, H. Intraspecific recurrent selection in V. vinifera: An effective method for breeding of high quality, disease-, cold-, and drought -resistant grapes. Euphytica 2021, 217, 111. [Google Scholar] [CrossRef]
- Li, Y.; Cui, C.W.; Li, N.N.; Li, Y.S.; Yang, J.M.; Wang, H.; Wang, Y.J. Difference of aromatic components of wine and distilled liquor from Ecolly and Petit Manseng grape. China Brew. 2020, 39, 71–77. [Google Scholar] [CrossRef]
- Li, H. A study on the differences in susceptibility to downy mildew (Plasmopara viticola) among the cultivars of Vitis vinifera. Acta Hortic. Sin. 1988, 15, 23–26. [Google Scholar]
- Wang, Z.; Wang, Y.; Cao, X.; Wu, D.; Hui, M.; Han, X.; Yao, F.; Li, Y.; Li, H.; Wang, H. Screening and Validation of SSR Molecular Markers for Identification of Downy Mildew Resistance in Intraspecific Hybrid F1 Progeny (V. vinifera). Horticulturae 2022, 8, 706. [Google Scholar] [CrossRef]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Francesca, S.; Giosuè, S.; Tubiello, F.; Rossi, V.; Federico, S.; Cynthia, R.; Gullino, M. Downy mildew (Plasmopara viticola) epidemics on grapevine under climate change. Glob. Change Biol. 2006, 12, 1299–1307. [Google Scholar] [CrossRef]
- Williams, M.G.; Magarey, P.A.; Sivasithamparam, K. Effect of temperature and light intensity on early infection behaviour of a Western Australian isolate of Plasmopara viticola, the downy mildew pathogen of grapevine. Australas. Plant Pathol. 2007, 36, 325–331. [Google Scholar] [CrossRef]
- Li, H. Studies on the resistance of grapevine to powdery mildew. Plant Pathol. 1993, 42, 792–796. [Google Scholar] [CrossRef]
- Donald, T.M.; Pellerone, F.; Adam-Blondon, A.F.; Bouquet, A.; Thomas, M.R.; Dry, I.B. Identification of resistance gene analogs linked to a powdery mildew resistance locus in grapevine. Theor. Appl. Genet. 2002, 104, 610–618. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; He, P.; Chen, J.; Lamikanra, O.; Lu, J. Evaluation of foliar resistance to Uncinula necator in Chines wild Vitis species. Vitis-J. Grapevine Res. 2015, 34, 159–164. [Google Scholar]
- Kozma, P., Jr. Winegrape breeding for fungus disease resistance. Acta Hortic. 2000, 528, 511–516. [Google Scholar] [CrossRef]
- Leng, H.; Guo, Y.; Su, K.; Shi, G.; Zhao, Y.; Song, J.; Liu, Z.; Li, K.; Guo, X. Genetic analysis of Downy mildew resistance for grape hybrid offspring. J. Shenyang Agric. Univ. 2017, 48, 713–718. [Google Scholar]
- Ren, Z.; Guo, Y.; Zhang, J.; Liu, Z.; Gao, H.; Li, K.; Guo, X. Gray mold resistance identification and genetic analysis for offspring of grapevine. Sino-Overseas Grapevine Wine 2018, 4, 24–28. [Google Scholar] [CrossRef]
- Song, R.; Zheng, Y.; Lu, W.; Shen, Y.; Fan, S.; Yang, Y.; Li, X.; Li, X. Genetic analysis of resistance to Plasmopara viticola disease in progenies (F1-F4) derived from interspecific hybridization with Vitis amurensis. J. Fruit Sci. 2008, 25, 33–39. [Google Scholar] [CrossRef]
- Zhang, J. Detection of QTLs for Resistance to Downy Mildew in Grape (Vitis vinifera L. × Vitis labrusca L.) and Screening and Analysis of Candidate Genes. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2020. [Google Scholar]
- Ai, J.; Shen, Y.J.; Li, X.H.; Zang, P.; Ge, Y.X.; Li, C.Y. Relationship between leaf surface stomata and downy mildew of Vitis amurensis Rupr. Spec. Wild Econ. Anim. Plant Res. 1995, 2, 14–16. [Google Scholar] [CrossRef]
- Atallah, S.S.; Gómez, M.I.; Fuchs, M.F.; Martinson, T.E. Economic Impact of Grapevine Leafroll Disease on Vitis vinifera cv. Cabernet franc in Finger Lakes Vineyards of New York. Am. J. Enol. Vitic. 2012, 63, 73–79. [Google Scholar] [CrossRef]
- Gao, Z.; Khot, L.R.; Naidu, R.A.; Zhang, Q. Early detection of grapevine leafroll disease in a red-berried wine grape cultivar using hyperspectral imaging. Comput. Electron. Agric. 2020, 179, 105807. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Ren, Y.; Ji, L.; Li, G. Comparisons and correlation analysis of different identifying methods for grape cultivars resistance to Plasmopara viticola. Xinjiang Agric. Sci. 2012, 7, 1244–1249. [Google Scholar] [CrossRef]
- Bayo-Canha, A.; Fernández-Fernández, J.I.; Martínez-Cutillas, A.; Ruiz-García, L. Phenotypic segregation and relationships of agronomic traits in Monastrell × Syrah wine grape progeny. Euphytica 2012, 186, 393–407. [Google Scholar] [CrossRef]
- Wang, Z.L.; Yao, F.; Hui, M.; Wu, D.; Wang, Y.; Han, X.; Cao, X.; Li, Y.H.; Li, H.; Wang, H. Fertility analysis of intraspecific hybrids in Vitis vinifera and screening of superior hybrid combinations. Front. Plant Sci. 2022, 13, 940540. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.S.; Li, D.M.; Huang, W.D.; Liu, C.H.; Wang, L.J.; Wang, X.Q.; Wen, P.F. Wine Grape Varieties; China Agricultural University Press: Beijing, China, 2015. [Google Scholar]
- Li, H. The utilization of Desaymard classification method in identification of the grape resistance to grape downy mildew. J. Sichuan Agric. Univ. 1991, 9, 303–307. [Google Scholar]
- Wang, Y.; Liu, Y.; He, P.; Lamikanra, O.; Lu, J. Resistance of Chinese Vitis species to Elsinoë ampelina (de Bary) Shear. HortScience 1998, 33, 123–126. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Hybrid Combinations (♀ × ♂) | Disease Index of Parents | Bias in Progeny | CV (%) | H2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| P1 (♀) | P2 (♂) | HH (%) | HM (%) | LL (%) | |||||
| 2021A | C1 (CS × ECL) | 51.85 ± 8.42 | 7.39 ± 2.75 | 29.62 | 70.59 | 88.24 | 0 | 49.60 | 67.23 |
| C7 (ECL × CS) | 7.39 ± 2.75 | 51.85 ± 8.42 | 13.85 | 70.77 | 7.81 | 62.92 | 63.60 | ||
| C3 (MSL × ECL) | 48.14 ± 5.62 | 7.39 ± 2.75 | 27.77 | 71.93 | 91.23 | 5.26 | 58.06 | 76.65 | |
| C9 (ECL × MSL) | 7.39 ± 2.75 | 48.14 ± 5.62 | 25.93 | 59.26 | 7.41 | 51.42 | 65.21 | ||
| C6 (DKF × ECL) | 21.40 ± 4.89 | 7.39 ± 2.75 | 14.40 | 28.57 | 96.43 | 0 | 35.86 | 62.14 | |
| C10 (ECL × DKF) | 7.39 ± 2.75 | 21.40 ± 4.89 | 33.33 | 76.19 | 7.14 | 46.62 | 59.52 | ||
| 2022A | C1 (CS × ECL) | 47.22 ± 9.22 | 4.26 ± 1.45 | 25.74 | 47.06 | 88.24 | 5.88 | 63.76 | 76.73 |
| C7 (ECL × CS) | 4.26 ± 1.45 | 47.22 ± 9.22 | 12.5 | 29.69 | 6.25 | 62.86 | 62.09 | ||
| C3 (MSL × ECL) | 33.33 ± 5.62 | 4.26 ± 1.45 | 18.80 | 43.86 | 91.23 | 3.51 | 51.92 | 65.88 | |
| C9 (ECL × MSL) | 4.26 ± 1.45 | 33.33 ± 5.62 | 14.81 | 40.74 | 14.81 | 77.45 | 78.92 | ||
| C6 (DKF × ECL) | 9.09 ± 3.28 | 4.26 ± 1.45 | 6.68 | 28.57 | 64.29 | 10.71 | 59.34 | 59.47 | |
| C10 (ECL × DKF) | 4.26 ± 1.45 | 9.09 ± 3.28 | 11.43 | 37.14 | 14.29 | 57.86 | 57.39 | ||
| 2022B | C1 (CS × ECL) | 47.22 ± 9.22 | 4.26 ± 1.45 | 25.74 | 25.01 | 81.68 | 8.4 | 61.90 | 58.89 |
| C7 (ECL × CS) | 4.26 ± 1.45 | 47.22 ± 9.22 | 11.76 | 62.75 | 39.22 | 84.26 | 55.23 | ||
| C3 (MSL × ECL) | 33.33 ± 5.62 | 4.26 ± 1.45 | 18.80 | 12.74 | 66.41 | 27.41 | 88.34 | 68.82 | |
| C9 (ECL × MSL) | 4.26 ± 1.45 | 33.33 ± 5.62 | 15.38 | 61.54 | 30.76 | 78.44 | 79.99 | ||
| C6 (DKF × ECL) | 9.09 ± 3.28 | 4.26 ± 1.45 | 6.68 | 22.39 | 52.99 | 8.21 | 64.62 | 57.46 | |
| C10 (ECL × DKF) | 4.26 ± 1.45 | 9.09 ± 3.28 | 15.56 | 55.56 | 24.44 | 77.92 | 61.23 | ||
| Year | Hybrid Combinations (♀ × ♂) | Disease Index of Parents | Disease Index of Progeny | CV (%) | H2 | |||
|---|---|---|---|---|---|---|---|---|
| P1 (♀) | P2 (♂) | Range | Average | |||||
| 2021A | C1 (CS × ECL) | 49.29 ± 6.87 | 9.40 ± 4.85 | 29.35 | 2.22–70.37 | 34.32 ± 14.32 | 57.07 | 66.83 |
| C7 (ECL × CS) | 9.40 ± 4.85 | 49.29 ± 6.87 | 0–55.55 | 11.08 ± 11.05 | 99.70 | 60.90 | ||
| C3 (MSL × ECL) | 48.21 ± 9.56 | 9.40 ± 4.85 | 28.81 | 0–88.89 | 42.89 ± 25.10 | 58.52 | 70.34 | |
| C9 (ECL × MSL) | 9.40 ± 4.85 | 48.21 ± 9.56 | 0–15.56 | 6.77 ± 5.10 | 75.30 | 56.31 | ||
| C6 (DKF × ECL) | 29.40 ± 4.36 | 9.40 ± 5.36 | 19.40 | 0–55.56 | 21.92 ± 9.95 | 45.39 | 62.60 | |
| C10 (ECL × DKF) | 9.40 ± 5.36 | 29.40 ± 4.36 | 0–22.16 | 7.89 ± 5.81 | 73.64 | 61.91 | ||
| 2022A | C1 (CS × ECL) | 29.12 ± 7.26 | 4.12 ± 1.36 | 16.62 | 0–70.37 | 35.31 ± 20.97 | 59.39 | 77.39 |
| C7 (ECL × CS) | 4.12 ± 1.36 | 29.12 ± 7.26 | 0–55.55 | 6.14 ± 9.66 | 157.45 | 66.25 | ||
| C3 (MSL × ECL) | 26.54 ± 4.13 | 4.12 ± 1.36 | 15.33 | 0–55.55 | 22.71 ± 15.48 | 68.18 | 79.72 | |
| C9 (ECL × MSL) | 4.12 ± 1.36 | 26.54 ± 4.13 | 0–22.22 | 4.52 ± 4.85 | 107.45 | 64.57 | ||
| C6 (DKF × ECL) | 7.22 ± 1.78 | 4.12 ± 1.36 | 5.67 | 0–34.44 | 6.70 ± 7.50 | 107.31 | 76.26 | |
| C10 (ECL × DKF) | 4.12 ± 1.36 | 7.22 ± 1.78 | 0–11.08 | 3.22 ± 2.68 | 83.06 | 76.50 | ||
| 2022B | C1 (CS × ECL) | 29.12 ± 7.26 | 4.12 ± 1.36 | 16.62 | 0–86.67 | 15.28 ± 12.98 | 85.00 | 61.71 |
| C7 (ECL × CS) | 4.12 ± 1.36 | 29.12 ± 7.26 | 0–13.89 | 3.24 ± 3.54 | 109.17 | 53.00 | ||
| C3 (MSL × ECL) | 26.54 ± 4.13 | 4.12 ± 1.36 | 15.33 | 0–63.89 | 9.02 ± 12.97 | 143.91 | 75.63 | |
| C9 (ECL × MSL) | 4.12 ± 1.36 | 26.54 ± 4.13 | 0–19.44 | 4.76 ± 5.95 | 124.87 | 62.60 | ||
| C6 (DKF × ECL) | 7.22 ± 1.78 | 4.12 ± 1.36 | 5.67 | 0–53.09 | 8.55 ± 10.19 | 119.13 | 82.27 | |
| C10 (ECL × DKF) | 4.12 ± 1.36 | 7.22 ± 1.78 | 0–30.56 | 5.77 ± 7.55 | 130.89 | 76.24 | ||
| Population | Orthogonal Combinations | Correspondence Number | Inverse Combinations | Correspondence Number |
|---|---|---|---|---|
| A | C1 (CS × ECL) | 2001001–2001017 | C7 (ECL × CS) | 2007001–2007064 |
| C3 (MSL × ECL) | 2003001–2003057 | C9 (ECL × MSL) | 2009001–2009027 | |
| C6 (DKF × ECL) | 2006001–2006029 | C10 (ECL × DKF) | 2010001–2010035 | |
| B | C1 (CS × ECL) | 2101001–2101726 | C7 (ECL × CS) | 2107001–2107051 |
| C3 (MSL × ECL) | 2103001–2103459 | C9 (ECL × MSL) | 2109001–2109013 | |
| C6 (DKF × ECL) | 2106001–2106134 | C10 (ECL × DKF) | 2110001–2101045 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Li, Y.; Wang, Z.; Li, Z.; Li, N.; Li, H.; Duan, X. Identification and Genetic Analysis of Downy Mildew Resistance in Intraspecific Hybrids of Vitis vinifera L. Plants 2025, 14, 2415. https://doi.org/10.3390/plants14152415
Han X, Li Y, Wang Z, Li Z, Li N, Li H, Duan X. Identification and Genetic Analysis of Downy Mildew Resistance in Intraspecific Hybrids of Vitis vinifera L. Plants. 2025; 14(15):2415. https://doi.org/10.3390/plants14152415
Chicago/Turabian StyleHan, Xing, Yihan Li, Zhilei Wang, Zebin Li, Nanyang Li, Hua Li, and Xinyao Duan. 2025. "Identification and Genetic Analysis of Downy Mildew Resistance in Intraspecific Hybrids of Vitis vinifera L." Plants 14, no. 15: 2415. https://doi.org/10.3390/plants14152415
APA StyleHan, X., Li, Y., Wang, Z., Li, Z., Li, N., Li, H., & Duan, X. (2025). Identification and Genetic Analysis of Downy Mildew Resistance in Intraspecific Hybrids of Vitis vinifera L. Plants, 14(15), 2415. https://doi.org/10.3390/plants14152415