
Comparison of Deep Learning Models for LAI Simulation and Interpretable Hydrothermal Coupling in the Loess Plateau

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Datasets and Preprocessing
2.2.1. Meteorological Data
2.2.2. LAI Data
2.2.3. Land Cover Type Data
2.2.4. Data Preprocessing
2.3. Model Architecture
2.3.1. Artificial Neural Network·
2.3.2. Long Short-Term Memory
2.3.3. Interpretable Multivariable Long Short-Term Memory
2.4. Experimental Setup
2.5. Statistical Analysis
3. Results
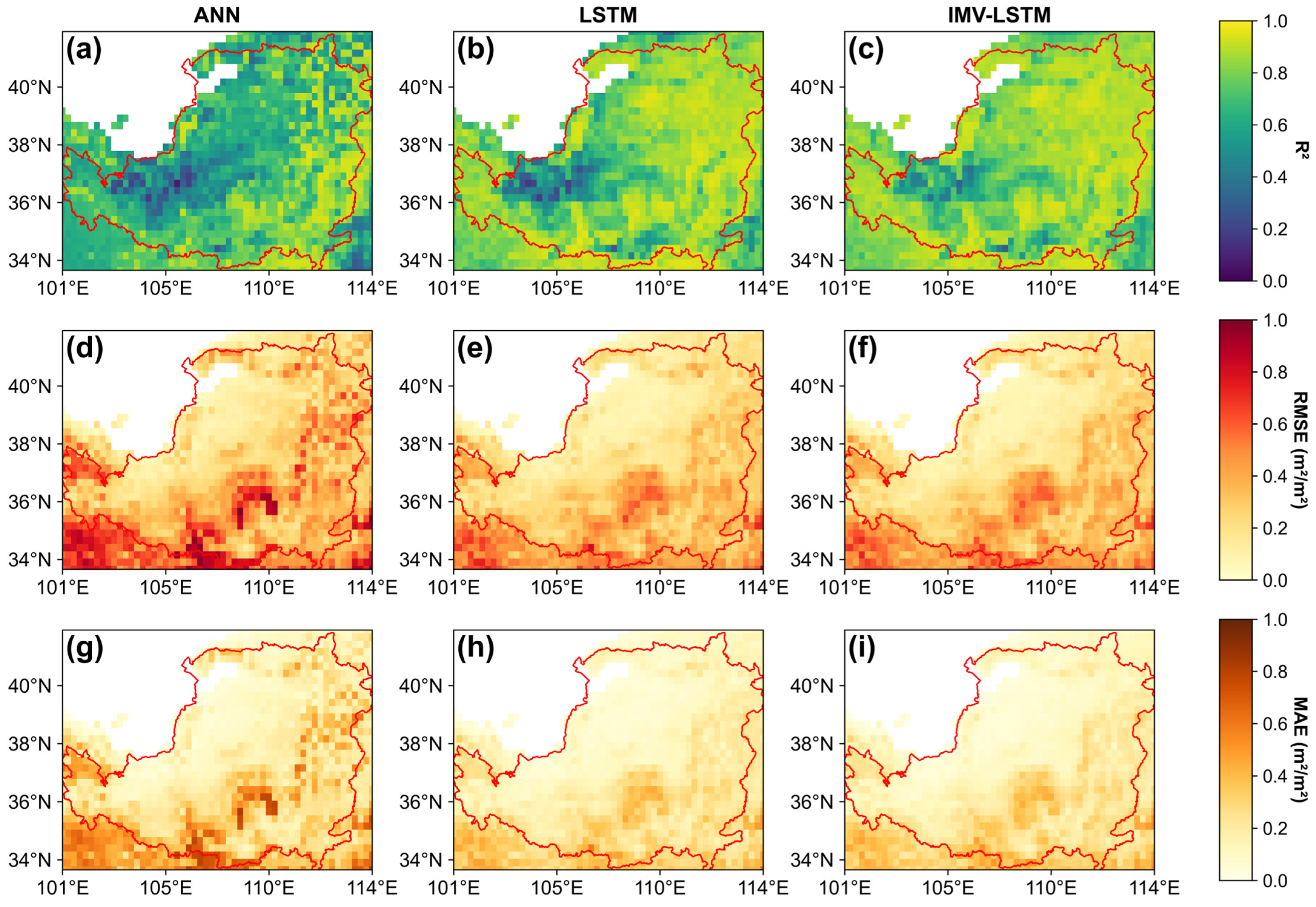
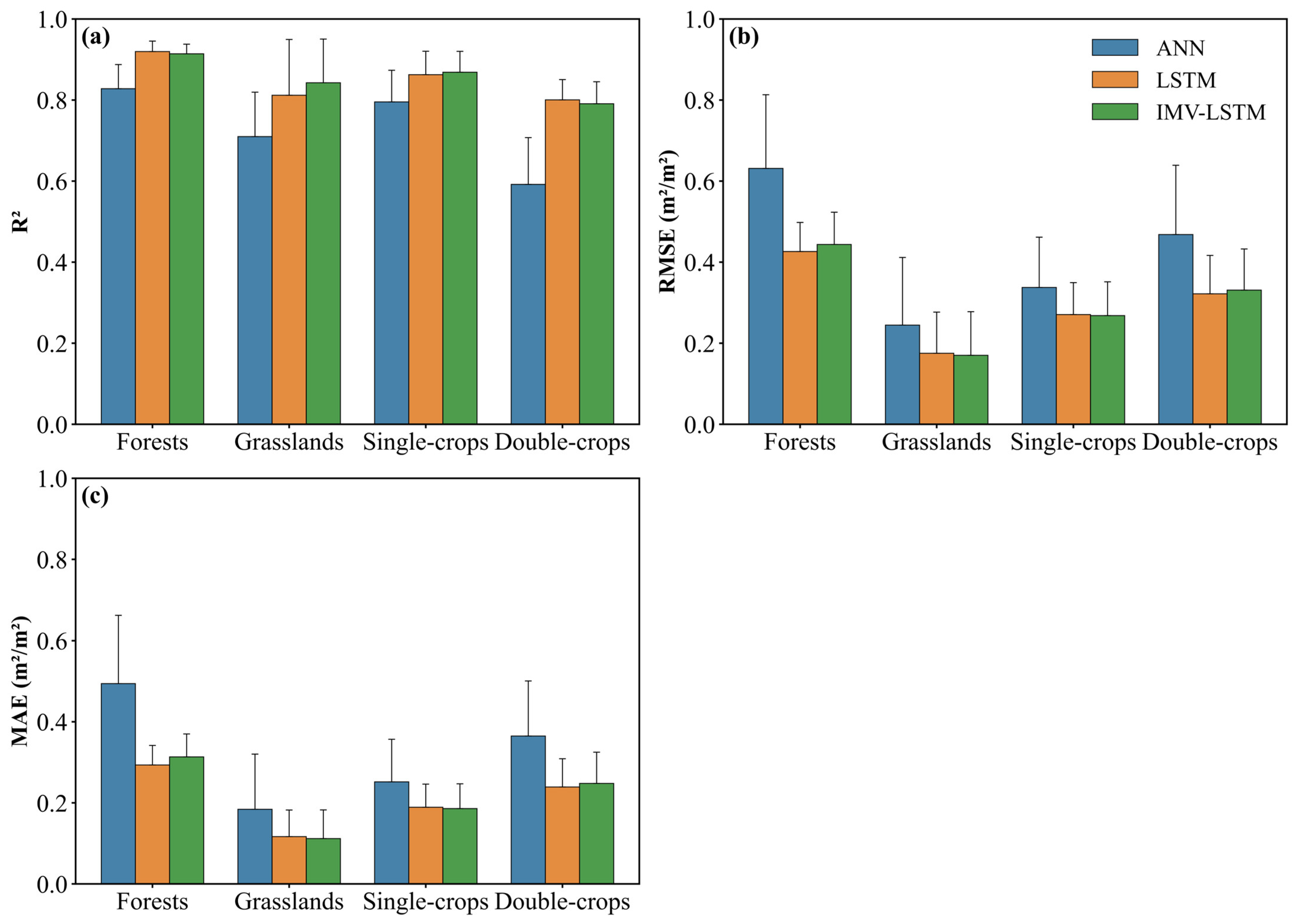
3.1. Model Evaluation
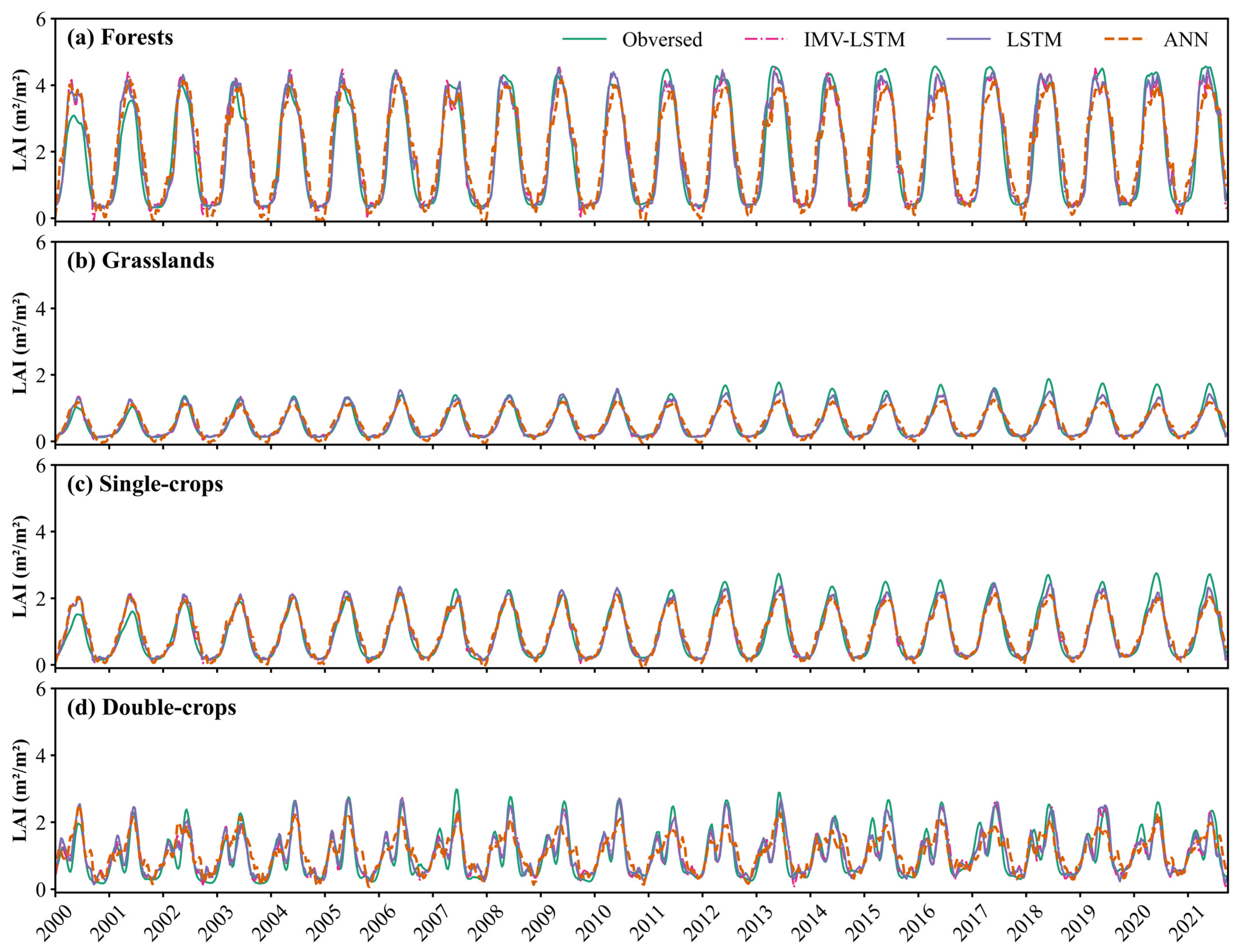
3.2. LAI Preditions for Different Vegetations
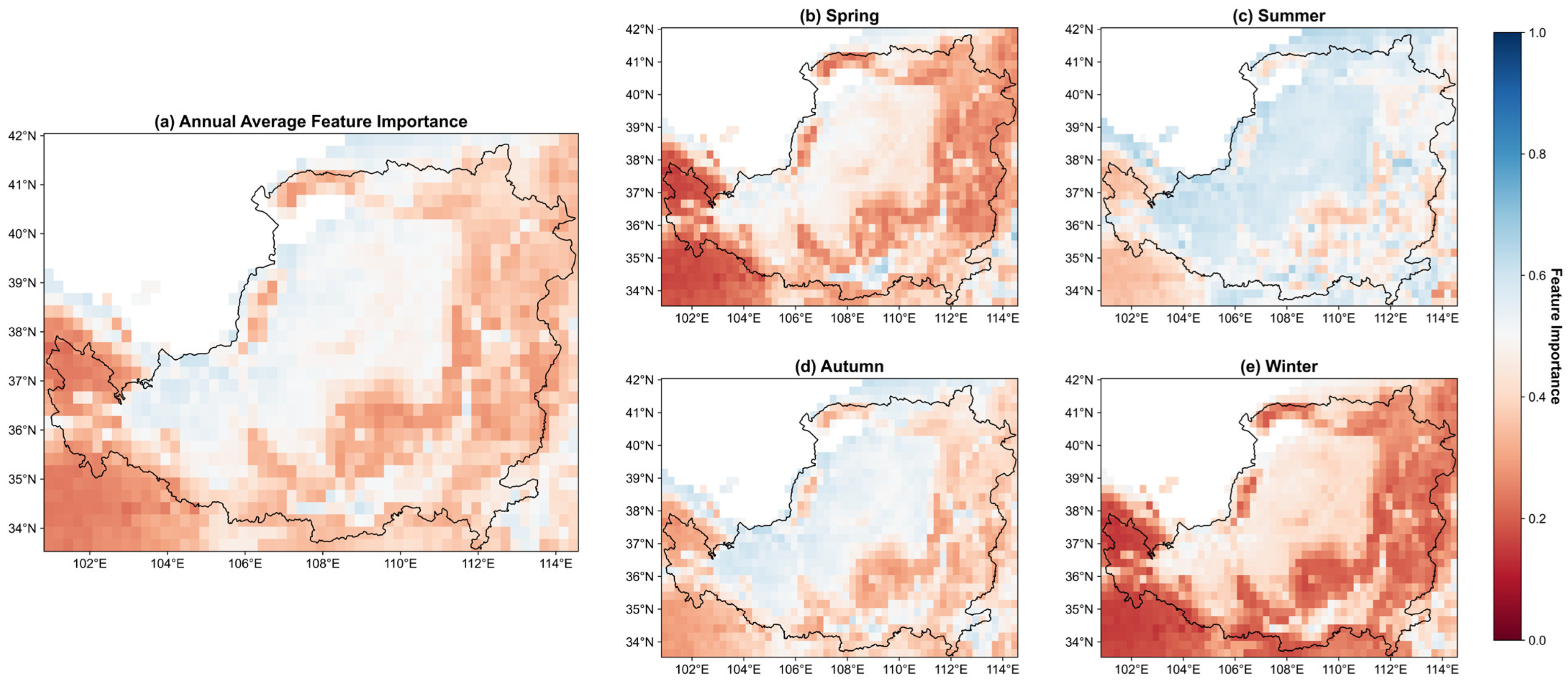
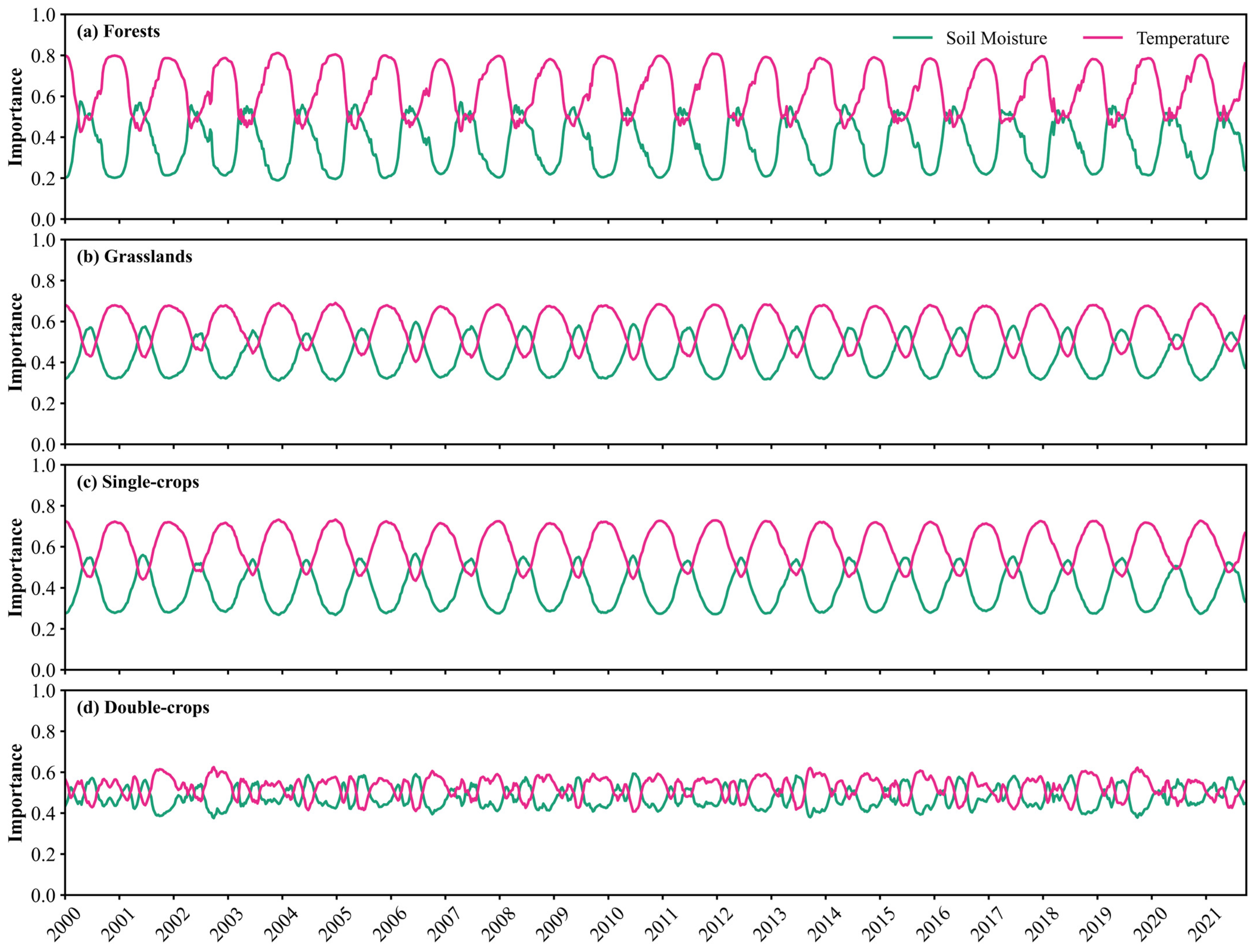
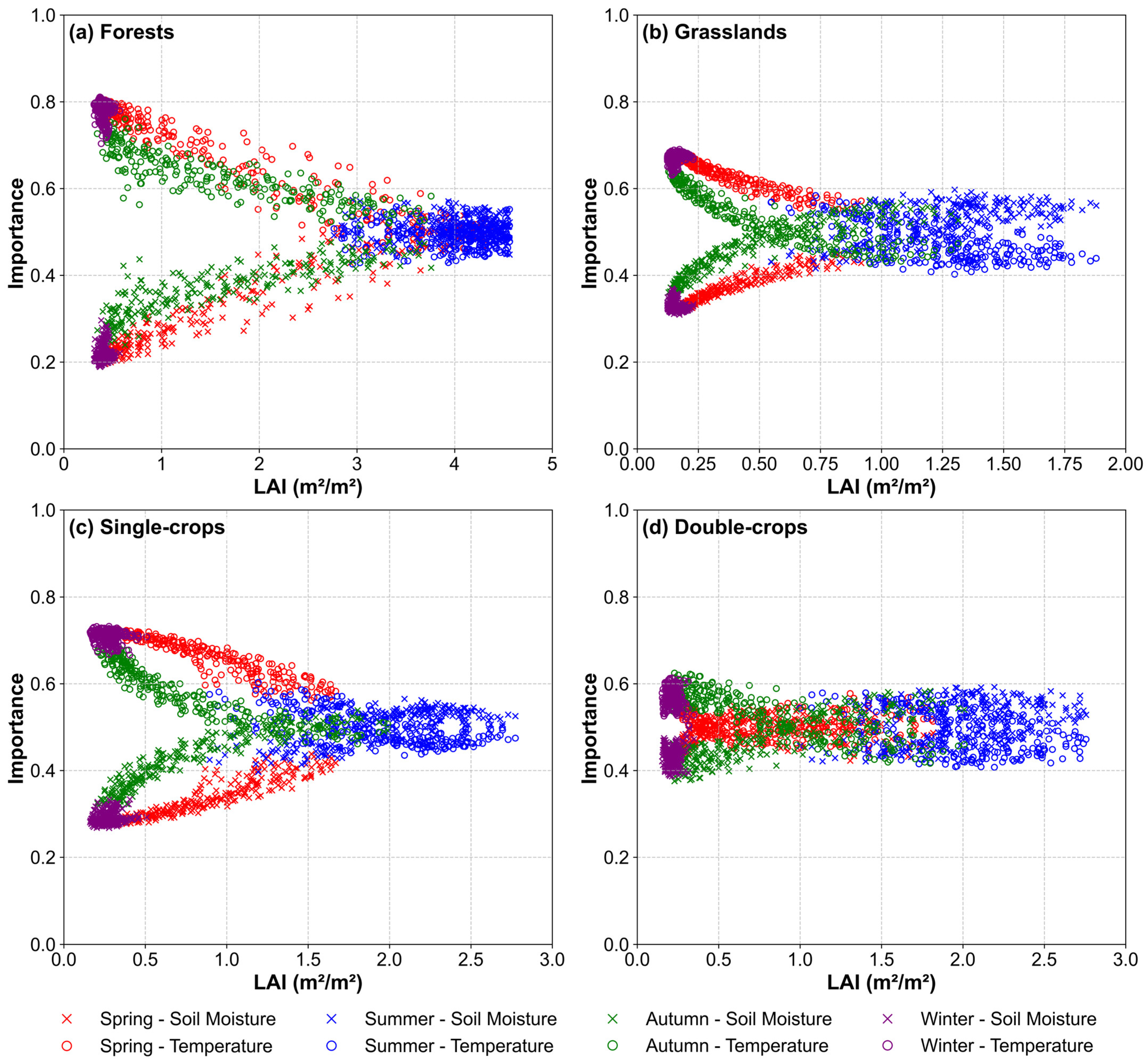
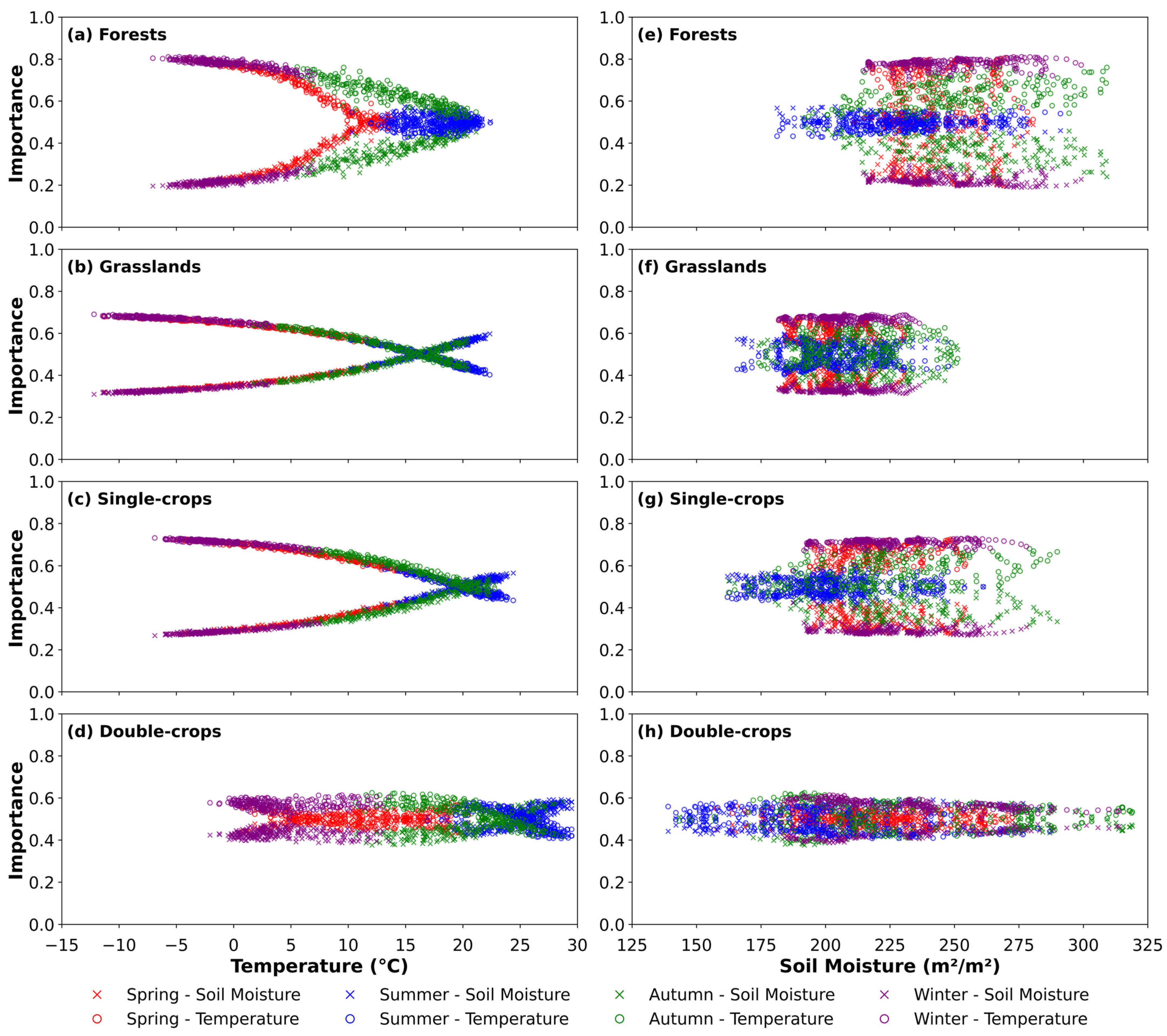
3.3. Variable Importance Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiao, Y.; Lei, H.; Yang, D.; Huang, M.; Liu, D.; Yuan, X. Impact of Vegetation Dynamics on Hydrological Processes in a Semi-Arid Basin by Using a Land Surface-Hydrology Coupled Model. J. Hydrol. 2017, 551, 116–131. [Google Scholar] [CrossRef]
- Zhang, B.; Tian, L.; Zhao, X.; Wu, P. Feedbacks between Vegetation Restoration and Local Precipitation over the Loess Plateau in China. Sci. China Earth Sci. 2021, 64, 920–931. [Google Scholar] [CrossRef]
- Chen, M.; Willgoose, G.R.; Saco, P.M. Investigating the Impact of Leaf Area Index Temporal Variability on Soil Moisture Predictions Using Remote Sensing Vegetation Data. J. Hydrol. 2015, 522, 274–284. [Google Scholar] [CrossRef]
- Zhang, G.; Ming, B.; Shen, D.; Xie, R.; Hou, P.; Xue, J.; Wang, K.; Li, S. Optimizing Grain Yield and Water Use Efficiency Based on the Relationship between Leaf Area Index and Evapotranspiration. Agriculture 2021, 11, 313. [Google Scholar] [CrossRef]
- Ek, M.B.; Mitchell, K.E.; Lin, Y.; Rogers, E.; Grunmann, P.; Koren, V.; Gayno, G.; Tarpley, J.D. Implementation of Noah Land Surface Model Advances in the National Centers for Environmental Prediction Operational Mesoscale Eta Model. J. Geophys. Res. Atmos. 2003, 108, 2002JD003296. [Google Scholar] [CrossRef]
- Rodell, M.; Houser, P.R.; Jambor, U.; Gottschalck, J.; Mitchell, K.; Meng, C.-J.; Arsenault, K.; Cosgrove, B.; Radakovich, J.; Bosilovich, M.; et al. The Global Land Data Assimilation System. Bull. Am. Meteorol. Soc. 2004, 85, 381–394. [Google Scholar] [CrossRef]
- Kolby Smith, W.; Reed, S.C.; Cleveland, C.C.; Ballantyne, A.P.; Anderegg, W.R.L.; Wieder, W.R.; Liu, Y.Y.; Running, S.W. Large Divergence of Satellite and Earth System Model Estimates of Global Terrestrial CO2 Fertilization. Nat. Clim. Change 2016, 6, 306–310. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant Phenology and Global Climate Change: Current Progresses and Challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, L.; Tian, X.; Zhang, J.; Yuan, W.; Zheng, Y.; Zhao, X.; Kato, T. Assimilation of Remotely-Sensed Leaf Area Index into a Dynamic Vegetation Model for Gross Primary Productivity Estimation. Remote Sens. 2017, 9, 188. [Google Scholar] [CrossRef]
- Luo, Y.; Ogle, K.; Tucker, C.; Fei, S.; Gao, C.; LaDeau, S.; Clark, J.S.; Schimel, D.S. Ecological Forecasting and Data Assimilation in a Data-Rich Era. Ecol. Appl. 2011, 21, 1429–1442. [Google Scholar] [CrossRef]
- Fisher, R.A.; Koven, C.D.; Anderegg, W.R.L.; Christoffersen, B.O.; Dietze, M.C.; Farrior, C.E.; Holm, J.A.; Hurtt, G.C.; Knox, R.G.; Lawrence, P.J.; et al. Vegetation Demographics in Earth System Models: A Review of Progress and Priorities. Glob. Change Biol. 2018, 24, 35–54. [Google Scholar] [CrossRef]
- Müller, C.; Elliott, J.; Chryssanthacopoulos, J.; Arneth, A.; Balkovic, J.; Ciais, P.; Deryng, D.; Folberth, C.; Glotter, M.; Hoek, S.; et al. Global Gridded Crop Model Evaluation: Benchmarking, Skills, Deficiencies and Implications. Geosci. Model. Dev. 2017, 10, 1403–1422. [Google Scholar] [CrossRef]
- Thiery, W.; Davin, E.L.; Lawrence, D.M.; Hirsch, A.L.; Hauser, M.; Seneviratne, S.I. Present-day Irrigation Mitigates Heat Extremes. J. Geophys. Res. Atmos. 2017, 122, 1403–1422. [Google Scholar] [CrossRef]
- Bi, K.; Xie, L.; Zhang, H.; Chen, X.; Gu, X.; Tian, Q. Accurate Medium-Range Global Weather Forecasting with 3D Neural Networks. Nature 2023, 619, 533–538. [Google Scholar] [CrossRef]
- Zhang, Y.; Long, M.; Chen, K.; Xing, L.; Jin, R.; Jordan, M.I.; Wang, J. Skilful Nowcasting of Extreme Precipitation with NowcastNet. Nature 2023, 619, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Ham, Y.-G.; Kim, J.-H.; Min, S.-K.; Kim, D.; Li, T.; Timmermann, A.; Stuecker, M.F. Anthropogenic Fingerprints in Daily Precipitation Revealed by Deep Learning. Nature 2023, 622, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Zhang, Z.; Gui, H.; Zhu, P.; Sun, Y.; Zhou, X.; Xiao, K.; Xin, Q. Predicting Time Series of Vegetation Leaf Area Index across North America Based on Climate Variables for Land Surface Modeling Using Attention-Enhanced LSTM. Int. J. Digit. Earth 2024, 17, 2372317. [Google Scholar] [CrossRef]
- Garofalo, S.P.; Modugno, A.F.; De Carolis, G.; Sanitate, N.; Negash Tesemma, M.; Scarascia-Mugnozza, G.; Tekle Tegegne, Y.; Campi, P. Explainable Artificial Intelligence to Predict the Water Status of Cotton (Gossypium hirsutum L., 1763) from Sentinel-2 Images in the Mediterranean Area. Plants 2024, 13, 3325. [Google Scholar] [CrossRef]
- Laroche-Pinel, E.; Duthoit, S.; Albughdadi, M.; Costard, A.D.; Rousseau, J.; Chéret, V.; Clenet, H. Towards Vine Water Status Monitoring on a Large Scale Using Sentinel-2 Images. Remote Sens. 2021, 13, 1837. [Google Scholar] [CrossRef]
- Shin, T.; Ko, J.; Jeong, S.; Kang, J.; Lee, K.; Shim, S. Assimilation of Deep Learning and Machine Learning Schemes into a Remote Sensing-Incorporated Crop Model to Simulate Barley and Wheat Productivities. Remote Sens. 2022, 14, 5443. [Google Scholar] [CrossRef]
- Ahlström, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The Dominant Role of Semi-Arid Ecosystems in the Trend and Variability of the Land CO2 Sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef]
- Li, W.; Migliavacca, M.; Forkel, M.; Walther, S.; Reichstein, M.; Orth, R. Revisiting Global Vegetation Controls Using Multi-layer Soil Moisture. Geophys. Res. Lett. 2021, 48, e2021GL092856. [Google Scholar] [CrossRef]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Peñuelas, J.; Seneviratne, S.I. Quantifying Soil Moisture Impacts on Light Use Efficiency across Biomes. New Phytol. 2018, 218, 1430–1449. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential Responses to Changes in Growth Temperature between Trees from Different Functional Groups and Biomes: A Review and Synthesis of Data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef]
- Wang, Q.; Fan, X.; Qin, Z.; Wang, M. Change Trends of Temperature and Precipitation in the Loess Plateau Region of China, 1961–2010. Glob. Planet. Change 2012, 92–93, 138–147. [Google Scholar] [CrossRef]
- Liu, T. Loess and the Environment; China Ocean Press: Beijing, China, 1985. [Google Scholar]
- Guo, R.; Li, F.; He, W.; Yang, S.; Sun, G. Spatial and Temporal Variability of Annual Precipitation during 1958–2007 in Loess Plateau, China. In Computer and Computing Technologies in Agriculture IV; Li, D., Liu, Y., Chen, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 551–560. [Google Scholar]
- Li, Z.; Zheng, F.; Liu, W.; Flanagan, D.C. Spatial Distribution and Temporal Trends of Extreme Temperature and Precipitation Events on the Loess Plateau of China during 1961–2007. Quat. Int. 2010, 226, 92–100. [Google Scholar] [CrossRef]
- Zhao, G.; Mu, X.; Wen, Z.; Wang, F.; Gao, P. Soil Erosion, Conservation, and Eco-environment Changes in the Loess Plateau of China. Land. Degrad. Dev. 2013, 24, 499–510. [Google Scholar] [CrossRef]
- Fu, B. Soil Erosion and Its Control in the Loess Plateau of China. Soil. Use Manag. 1989, 5, 76–82. [Google Scholar] [CrossRef]
- Huang, L.; Shao, M. Advances and Perspectives on Soil Water Research in China’s Loess Plateau. Earth Sci. Rev. 2019, 199, 102962. [Google Scholar] [CrossRef]
- Tao, Z.; Li, H.; Neil, E.; Si, B. Groundwater Recharge in Hillslopes on the Chinese Loess Plateau. J. Hydrol. Reg. Stud. 2021, 36, 100840. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, M.; Jin, Z.; Zhang, S.; Sun, P.; Gu, T.; Liu, X.; Lin, H.; An, Z.; Peng, J.; et al. The Genesis, Development, and Evolution of Original Vertical Joints in Loess. Earth Sci. Rev. 2021, 214, 103526. [Google Scholar] [CrossRef]
- Friedl, M.; Sulla-Menashe, D. MODIS/Terra+Aqua Land Cover Type Yearly L3 Global 500m SIN Grid V061 [Data set]; NASA Land Processes Distributed Active Archive Center: Sioux Falls, SD, USA, 2022. [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-Driven Increases in Global Terrestrial Net Primary Production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef]
- Seddon, A.W.R.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of Global Terrestrial Ecosystems to Climate Variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A Global Analysis of Root Distributions for Terrestrial Biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Yuan, H.; Dong, W.; Zhang, S.; Liu, S.; Wei, N.; Lu, X.; Wei, Z.; Hu, Y.; Dai, Y. Reprocessed MODIS Version 6.1 Leaf Area Index Dataset and Its Evaluation for Land Surface and Climate Modeling. Remote Sens. 2023, 15, 1780. [Google Scholar] [CrossRef]
- Myneni, R.; Knyazikhin, Y.; Park, T. MODIS/Terra Leaf Area Index/FPAR 8-Day L4 Global 500m SIN Grid V061; NASA Land Processes Distributed Active Archive Center: Sioux Falls, SD, USA, 2021. [CrossRef]
- Yuan, H.; Dai, Y.; Xiao, Z.; Ji, D.; Shangguan, W. Reprocessing the MODIS Leaf Area Index Products for Land Surface and Climate Modelling. Remote Sens. Environ. 2011, 115, 1171–1187. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-lag Effects of Global Vegetation Responses to Climate Change. Glob. Change Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Z.; Peng, S. Global Analysis of Time-Lag and -Accumulation Effects of Climate on Vegetation Growth. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102179. [Google Scholar] [CrossRef]
- Gardner, M.W.; Dorling, S.R. Artificial Neural Networks (the Multilayer Perceptron)—A Review of Applications in the Atmospheric Sciences. Atmos. Environ. 1998, 32, 2627–2636. [Google Scholar] [CrossRef]
- Hochreiter, S.; Schmidhuber, J. Long Short-Term Memory. Neural Comput. 1997, 9, 1735–1780. [Google Scholar] [CrossRef]
- Guo, T.; Lin, T.; Antulov-Fantulin, N. Exploring Interpretable LSTM Neural Networks over Multi-Variable Data. arXiv 2019, arXiv:1905.12034. [Google Scholar] [CrossRef]
- Kingma, D.P.; Ba, J. Adam: A Method for Stochastic Optimization. arXiv 2014. [Google Scholar] [CrossRef]
- Shu, Z.; Zhang, B.; Tian, L.; Zhao, X. Improving Dynamic Vegetation Modeling in noah-MP by Parameter Optimization and Data Assimilation over China’s Loess Plateau. J. Geophys. Res. Atmos. 2022, 127, e2022JD036703. [Google Scholar] [CrossRef]
- Shu, Z.; Zhang, B.; Yu, L.; Zhao, X. Reconciling Plant Water Stress Response Using Vegetation and Soil Moisture Data Assimilation for Vegetation-Soil-Hydrology Interaction Estimation over the Chinese Loess Plateau. Agric. For. Meteorol. 2025, 369, 110581. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of Vegetation to Drought Time-Scales across Global Land Biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Tan, J.; Chen, A.; Fu, Y.H.; Ciais, P.; Liu, Q.; Janssens, I.A.; Vicca, S.; Zeng, Z.; Jeong, S.-J.; et al. Leaf Onset in the Northern Hemisphere Triggered by Daytime Temperature. Nat. Commun. 2015, 6, 6911. [Google Scholar] [CrossRef]
- Peng, J.; Dong, W.; Yuan, W.; Zhang, Y. Responses of Grassland and Forest to Temperature and Precipitation Changes in Northeast China. Adv. Atmos. Sci. 2012, 29, 1063–1077. [Google Scholar] [CrossRef]
- Li, Q.; Dong, B.; Qiao, Y.; Liu, M.; Zhang, J. Root Growth, Available Soil Water, and Water-Use Efficiency of Winter Wheat under Different Irrigation Regimes Applied at Different Growth Stages in North China. Agric. Water Manag. 2010, 97, 1676–1682. [Google Scholar] [CrossRef]
- Yang, F.; Huang, M.; Li, C.; Wu, X.; Guo, T.; Zhu, M. Changes in Soil Moisture and Organic Carbon under Deep-Rooted Trees of Different Stand Ages on the Chinese Loess Plateau. Agric. Ecosyst. Environ. 2022, 328, 107855. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, N.; Zhang, C.; Zhang, X.; He, X.; Jiang, W.; Li, J.; Zhan, Z.; Peng, D.; Lv, D.; et al. Deep Soil Moisture Has Limited Impact on Mitigating Drought Stress Effects on Plant Transpiration in a Subtropical Secondary Forest. Plant Soil. 2025. [Google Scholar] [CrossRef]
- Hou, Q.; Pei, T.; Yu, X.; Chen, Y.; Ji, Z.; Xie, B. The Seasonal Response of Vegetation Water Use Efficiency to Temperature and Precipitation in the Loess Plateau, China. Glob. Ecol. Conserv. 2022, 33, e01984. [Google Scholar] [CrossRef]
- Bhattacharya, A. Effect of Low-Temperature Stress on Germination, Growth, and Phenology of Plants: A Review. In Physiological Processes in Plants Under Low Temperature Stress; Bhattacharya, A., Ed.; Springer: Singapore, 2022; pp. 1–106. ISBN 978-981-16-9037-2. [Google Scholar]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of Water Stress on Growth, Biomass Partitioning, and Water-Use Efficiency in Summer Maize (Zea mays L.) throughout the Growth Cycle. Acta Physiol. Plant. 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Huang, M.; Piao, S.; Ciais, P.; Peñuelas, J.; Wang, X.; Keenan, T.F.; Peng, S.; Berry, J.A.; Wang, K.; Mao, J.; et al. Air Temperature Optima of Vegetation Productivity across Global Biomes. Nat. Ecol. Evol. 2019, 3, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Solbrig, O.T. Plant Traits and Adaptive Strategies: Their Role in Ecosystem Function. In Biodiversity and Ecosystem Function; Schulze, E.-D., Mooney, H.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 97–116. ISBN 978-3-642-58001-7. [Google Scholar]
- Wang, Y.; Woodcock, C.E.; Buermann, W.; Stenberg, P.; Voipio, P.; Smolander, H.; Häme, T.; Tian, Y.; Hu, J.; Knyazikhin, Y.; et al. Evaluation of the MODIS LAI Algorithm at a Coniferous Forest Site in Finland. Remote Sens. Environ. 2004, 91, 114–127. [Google Scholar] [CrossRef]
- Fensholt, R.; Sandholt, I.; Rasmussen, M.S. Evaluation of MODIS LAI, fAPAR and the Relation between fAPAR and NDVI in a Semi-Arid Environment Using in Situ Measurements. Remote Sens. Environ. 2004, 91, 490–507. [Google Scholar] [CrossRef]
- Li, S.; Fang, H.; Zhang, Y.; Wang, Y. Comprehensive Evaluation of Global CI, FVC, and LAI Products and Their Relationships Using High-Resolution Reference Data. Sci. Remote Sens. 2022, 6, 100066. [Google Scholar] [CrossRef]
- Yan, K.; Park, T.; Yan, G.; Liu, Z.; Yang, B.; Chen, C.; Nemani, R.R.; Knyazikhin, Y.; Myneni, R.B. Evaluation of MODIS LAI/FPAR Product Collection 6. Part 2: Validation and Intercomparison. Remote Sens. 2016, 8, 460. [Google Scholar] [CrossRef]
- Fang, H.; Wang, Y.; Zhang, Y.; Li, S. Long-Term Variation of Global GEOV2 and MODIS Leaf Area Index (LAI) and Their Uncertainties: An Insight into the Product Stabilities. J. Remote Sens. 2021, 2021, 9842830. [Google Scholar] [CrossRef]
- Zhang, X.; Yan, K.; Liu, J.; Yang, K.; Pu, J.; Yan, G.; Heiskanen, J.; Zhu, P.; Knyazikhin, Y.; Myneni, R.B. An Insight into the Internal Consistency of MODIS Global Leaf Area Index Products. IEEE Trans. Geosci. Remote Sens. 2024, 62, 1–16. [Google Scholar] [CrossRef]
- Liu, Y.W.; Liu, Y.B.; Wang, W. Inter-Comparison of Satellite-Retrieved and Global Land Data Assimilation System-Simulated Soil Moisture Datasets for Global Drought Analysis. Remote Sens. Environ. 2019, 220, 1–18. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Si, Y.; Zhao, W.; Zhou, Z.; Jin, J.; Yan, W.; Shao, X.; Xu, Z.; Gan, J. Comparison of Deep Learning Models for LAI Simulation and Interpretable Hydrothermal Coupling in the Loess Plateau. Plants 2025, 14, 2391. https://doi.org/10.3390/plants14152391
Yu J, Si Y, Zhao W, Zhou Z, Jin J, Yan W, Shao X, Xu Z, Gan J. Comparison of Deep Learning Models for LAI Simulation and Interpretable Hydrothermal Coupling in the Loess Plateau. Plants. 2025; 14(15):2391. https://doi.org/10.3390/plants14152391
Chicago/Turabian StyleYu, Junpo, Yajun Si, Wen Zhao, Zeyu Zhou, Jiming Jin, Wenjun Yan, Xiangyu Shao, Zhixiang Xu, and Junwei Gan. 2025. "Comparison of Deep Learning Models for LAI Simulation and Interpretable Hydrothermal Coupling in the Loess Plateau" Plants 14, no. 15: 2391. https://doi.org/10.3390/plants14152391
APA StyleYu, J., Si, Y., Zhao, W., Zhou, Z., Jin, J., Yan, W., Shao, X., Xu, Z., & Gan, J. (2025). Comparison of Deep Learning Models for LAI Simulation and Interpretable Hydrothermal Coupling in the Loess Plateau. Plants, 14(15), 2391. https://doi.org/10.3390/plants14152391