

Identification of Salt Tolerance-Related NAC Genes in Wheat Roots Based on RNA-Seq and Association Analysis
Abstract
1. Introduction
2. Results
2.1. Isolation and Systematic Numbering of the TaNAC Gene Family
2.2. Transcriptional Response to Salt Stress of TaNACs
2.3. Association Analysis Between TaNACs and Root Salt-Tolerance Phenotypes
2.4. Candidate TaNAC Genes Responding to Salt Stress in Roots
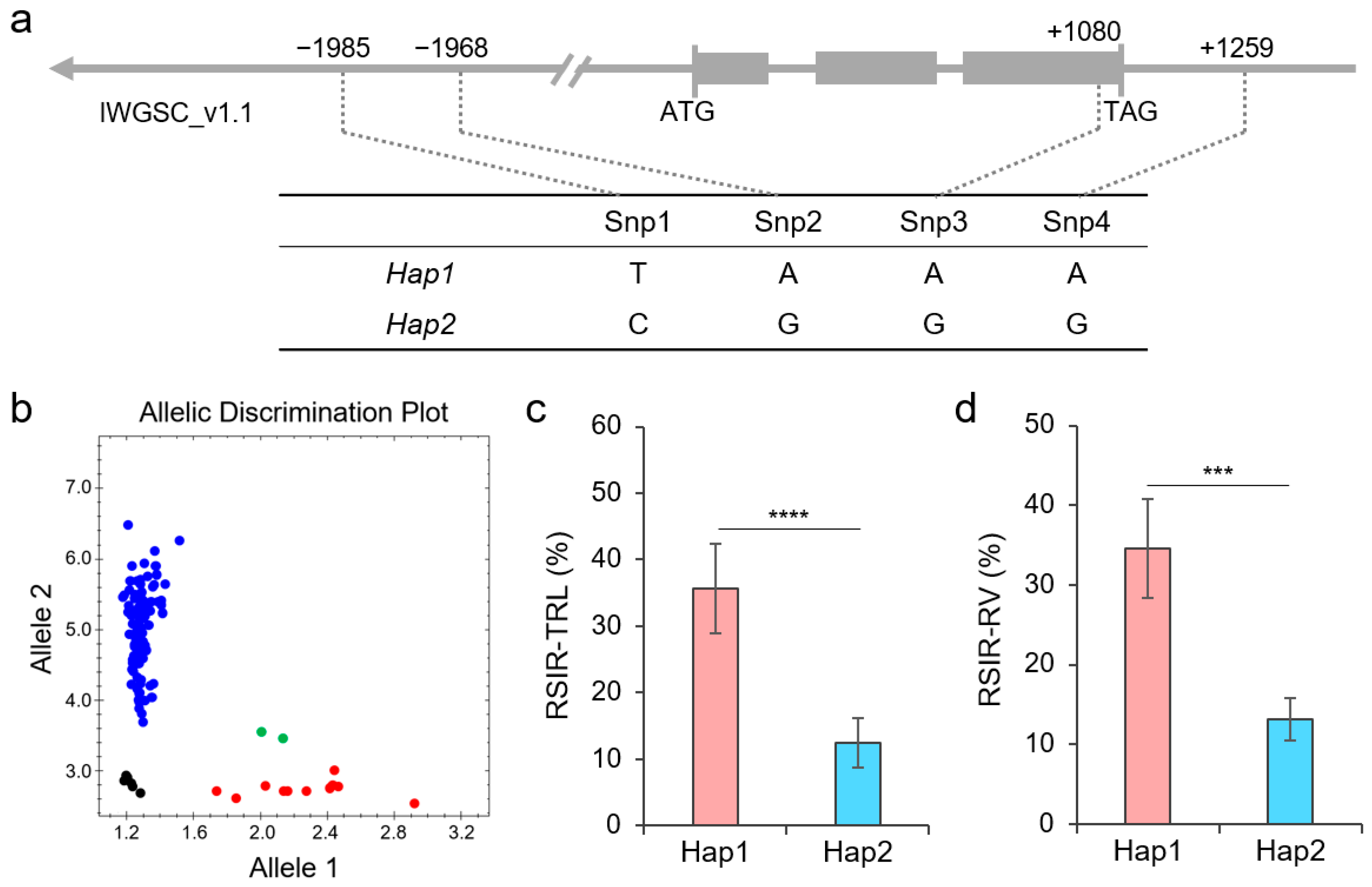
2.5. Functional Verification of TaNAC26-D3.2
2.6. Haplotypes of TaNAC26-D3.2
3. Discussion
3.1. Systematic Numbering of Wheat NAC Family
3.2. Response of TaNAC Genes to Salt Stress in Wheat Roots
3.3. TaNAC26-D3.2 Positively Regulates Salt Tolerance
4. Materials and Methods
4.1. Plant Materials
4.2. Isolation of NAC Sequences from Wheat Genome
4.3. Transcriptional Data of TaNAC Genes After Salt Treatment
4.4. RT-qPCR
4.5. Association Analysis of TaNAC Genes
4.6. Association Analysis of TaNAC16-B
4.7. Investigate for EMS-Mutant
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Roy, S.J.; Negrão, S.; Tester, M. Salt Resistant Crop Plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Zhang, Z.; Xia, Z.; Zhou, C.; Wang, G.; Meng, X.; Yin, P. Insights into Salinity Tolerance in Wheat. Genes 2024, 15, 573. [Google Scholar] [CrossRef]
- Butcher, K.; Wick, A.F.; Desutter, T.; Chatterjee, A.; Harmon, J. Soil Salinity: A Threat to Global Food Security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef]
- Han, K.; Zhao, Y.; Sun, Y.; Li, Y. NACs, Generalist in Plant Life. Plant Biotechnol. J. 2023, 21, 2433–2457. [Google Scholar] [CrossRef]
- Iqbal, A.; Bocian, J.; Hameed, A.; Orczyk, W.; Nadolska-Orczyk, A. Cis-Regulation by NACs: A Promising Frontier in Wheat Crop Improvement. Int. J. Mol. Sci. 2022, 23, 15431. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes Involved in Organ Separation in Arabidopsis: An Analysis of the Cup-Shaped Cotyledon Mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef]
- Sablowski, R.W.M.; Meyerowitz, E.M. A Homolog of NO APICAL MERISTEM Is an Immediate Target of the Floral Homeotic Genes APETALA3/PISTILLATA. Cell 1998, 92, 93–103. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Klíma, M.; Roy, A.; Tom Prášil, I. Biological Networks Underlying Abiotic Stress Tolerance in Temperate Crops-a Proteomic Perspective. Int. J. Mol. Sci. 2015, 16, 20913–20942. [Google Scholar] [CrossRef]
- Singh, S.; Grover, A.; Nasim, M. Biofuel Potential of Plants Transformed Genetically with NAC Family Genes. Front. Plant Sci. 2016, 7, 22. [Google Scholar] [CrossRef]
- Yang, S.D.; Seo, P.J.; Yoon, H.K.; Park, C.M. The Arabidopsis NAC Transcription Factor VNI2 Integrates Abscisic Acid Signals into Leaf Senescence via the COR/RD Genes. Plant Cell 2011, 23, 2155–2168. [Google Scholar] [CrossRef]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.I.; Asensi-Fabado, M.A.; Munná-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a Reactive Oxygen Species-Responsive NAC Transcription Factor, Regulates Longevity in Arabidopsis. Plant Cell 2012, 24, 482–506. [Google Scholar] [CrossRef]
- He, L.; Shi, X.; Wang, Y.; Guo, Y.; Yang, K.; Wang, Y. Arabidopsis ANAC069 Binds to C[A/G]CG[T/G] Sequences to Negatively Regulate Salt and Osmotic Stress Tolerance. Plant Mol. Biol. 2017, 93, 369–387. [Google Scholar] [CrossRef]
- Kim, S.G.; Lee, A.K.; Yoon, H.K.; Park, C.M. A Membrane-Bound NAC Transcription Factor NTL8 Regulates Gibberellic Acid-Mediated Salt Signaling in Arabidopsis Seed Germination. Plant J. 2008, 55, 77–88. [Google Scholar] [CrossRef]
- He, X.J.; Mu, R.L.; Cao, W.H.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. AtNAC2, a Transcription Factor Downstream of Ethylene and Auxin Signaling Pathways, Is Involved in Salt Stress Response and Lateral Root Development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef]
- Balazadeh, S.; Siddiqui, H.; Allu, A.D.; Matallana-Ramirez, L.P.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Köhler, B.; Mueller-Roeber, B. A Gene Regulatory Network Controlled by the NAC Transcription Factor ANAC092/AtNAC2/ORE1 during Salt-Promoted Senescence. Plant J. 2010, 62, 250–264. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, Z.; Lai, J.; Zhang, Y.; Yang, C.; Yin, B.; Zhao, Q.; Zhang, L.; Li, Y.; Yang, C.; et al. Dual Function of Arabidopsis ATAF1 in Abiotic and Biotic Stress Responses. Cell Res. 2009, 19, 1279–1290. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, J.; Wu, Y. Arabidopsis ATAF1 Enhances the Tolerance to Salt Stress and ABA in Transgenic Rice. J. Plant Res. 2016, 129, 955–962. [Google Scholar] [CrossRef]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) Transcription Factor Enhances Drought Resistance and Salt Tolerance in Rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef]
- Saad, A.S.I.; Li, X.; Li, H.P.; Huang, T.; Gao, C.S.; Guo, M.W.; Cheng, W.; Zhao, G.Y.; Liao, Y.C. A Rice Stress-Responsive NAC Gene Enhances Tolerance of Transgenic Wheat to Drought and Salt Stresses. Plant Sci. 2013, 203–204, 33–40. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, B.; Lu, G.; Han, B. Overexpression of a NAC Transcription Factor Enhances Rice Drought and Salt Tolerance. Biochem. Biophys. Res. Commun. 2009, 379, 985–989. [Google Scholar] [CrossRef]
- Takasaki, H.; Maruyama, K.; Kidokoro, S.; Ito, Y.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; Nakashima, K. The Abiotic Stress-Responsive NAC-Type Transcription Factor OsNAC5 Regulates Stress-Inducible Genes and Stress Tolerance in Rice. Mol. Genet. Genom. 2010, 284, 173–183. [Google Scholar] [CrossRef]
- Nakashima, K.; Tran, L.S.P.; Van Nguyen, D.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional Analysis of a NAC-Type Transcription Factor OsNAC6 Involved in Abiotic and Biotic Stress-Responsive Gene Expression in Rice. Plant J. 2007, 51, 617–630. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC Family Transcription Factor OsNAP Confers Abiotic Stress Response through the ABA Pathway. Plant Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Piao, W.; Lim, J.H.; Han, S.H.; Kim, Y.S.; An, G.; Paek, N.C. Rice ONAC106 Inhibits Leaf Senescence and Increases Salt Tolerance and Tiller Angle. Plant Cell Physiol. 2015, 56, 2325–2339. [Google Scholar] [CrossRef]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a Stress-Responsive NAC Transcription Factor Gene ONAC022 Improves Drought and Salt Tolerance in Rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef]
- Mao, X.; Zhang, H.; Qian, X.; Li, A.; Zhao, G.; Jing, R. TaNAC2, a NAC-Type Wheat Transcription Factor Conferring Enhanced Multiple Abiotic Stress Tolerances in Arabidopsis. J. Exp. Bot. 2012, 63, 2933–2946. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC Transcription Factor from Wheat, Enhances Salt and Drought Tolerance in Transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268. [Google Scholar] [CrossRef]
- Xu, Z.; Gongbuzhaxi; Wang, C.; Xue, F.; Zhang, H.; Ji, W. Wheat NAC Transcription Factor TaNAC29 Is Involved in Response to Salt Stress. Plant Physiol. Bioch. 2015, 96, 356–363. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y. Overexpression of TaNAC2D Displays Opposite Responses to Abiotic Stresses between Seedling and Mature Stage of Transgenic Arabidopsis. Front. Plant Sci. 2016, 7, 1754. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Jia, J.; Kong, X. The Novel Wheat Transcription Factor TaNAC47 Enhances Multiple Abiotic Stress Tolerances in Transgenic Plants. Front. Plant Sci. 2016, 6, 1174. [Google Scholar] [CrossRef]
- Mao, X.; Chen, S.; Li, A.; Zhai, C.; Jing, R. Novel NAC Transcription Factor TaNAC67 Confers Enhanced Multi-Abiotic Stress Tolerances in Arabidopsis. PLoS ONE 2014, 9, e84359. [Google Scholar] [CrossRef]
- Chen, D.; Chai, S.; McIntyre, C.L.; Xue, G.P. Overexpression of A Predominantly Root-Expressed NAC Transcription Factor in Wheat Roots Enhances Root Length, Biomass and Drought Tolerance. Plant Cell Rep. 2018, 37, 225–237. [Google Scholar] [CrossRef]
- Chi, Q.; Du, L.Y.; Ma, W.; Niu, R.Y.; Wu, B.W.; Guo, L.J.; Ma, M.; Liu, X.L.; Zhao, H.X. The miR164-TaNAC14 Module Regulates Root Development and Abiotic-Stress Tolerance in Wheat Seedlings. J. Integr. Agr. 2023, 22, 981–998. [Google Scholar] [CrossRef]
- Li, J.; Xie, L.; Tian, X.; Liu, S.; Xu, D.; Jin, H.; Song, J.; Dong, Y.; Zhao, D.; Li, G.; et al. TaNAC100 Acts as an Integrator of Seed Protein and Starch Synthesis Exerting Pleiotropic Effects of Agronomic Traits in Wheat. Plant J. 2021, 108, 829–840. [Google Scholar] [CrossRef]
- Wang, F.; Lin, R.; Feng, J.; Chen, W.; Qiu, D.; Xu, S. TaNAC1 acts as a negative regulator of stripe rust resistance in wheat, enhances susceptibility to Pseudomonas syringae, and promotes lateral root development in transgenic Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 108. [Google Scholar] [CrossRef]
- Zhao, D.; Derkx, A.P.; Liu, D.C.; Buchner, P.; Hawkesford, M.J. Overexpression of a NAC Transcription Factor Delays Leaf Senescence and Increases Grain Nitrogen Concentration in Wheat. Plant Biol. 2015, 17, 904–913. [Google Scholar] [CrossRef]
- Ma, J.; Tang, X.; Sun, B.; Wei, J.; Ma, L.; Yuan, M.; Zhang, D.; Shao, Y.; Li, C.; Chen, K.M. A NAC Transcription Factor, TaNAC5D-2, Acts as a Positive Regulator of Drought Tolerance Through Regulating Water Loss in Wheat (Triticum aestivum L.). Environ. Exp. Bot. 2022, 196, 104805. [Google Scholar] [CrossRef]
- Perochon, A.; Kahla, A.; Vranić, M.; Jia, J.; Malla, K.B.; Craze, M.; Wallington, E.; Doohan, F.M. A Wheat NAC Interacts with an Orphan Protein and Enhances Resistance to Fusarium Head Blight Disease. Plant Biotechnol. J. 2019, 17, 1892–1904. [Google Scholar] [CrossRef]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC Gene Regulating Senescence Improves Grain Protein, Zinc, and Iron Content in Wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef]
- Mao, H.; Li, S.; Wang, Z.; Cheng, X.; Li, F.; Mei, F.; Chen, N.; Kang, Z. Regulatory Changes in TaSNAC8-6A are Associated with Drought Tolerance in Wheat Seedlings. Plant Biotechnol. J. 2020, 18, 1078–1092. [Google Scholar] [CrossRef]
- Chen, D.; Richardson, T.; Chai, S.; Lynne McIntyre, C.; Rae, A.L.; Xue, G.P. Drought-Up-Regulated TaNAC69-1 is a Transcriptional Repressor of TaSHY2 and TaIAA7, and Enhances Root Length and Biomass in Wheat. Plant Cell Physiol. 2016, 57, 2076–2090. [Google Scholar] [CrossRef]
- He, X.; Qu, B.; Li, W.; Zhao, X.; Teng, W.; Ma, W.; Ren, Y.; Li, B.; Li, Z.; Tong, Y. The Nitrate-Inducible NAC Transcription Factor TaNAC2-5A Controls Nitrate Response and Increases Wheat Yield. Plant Physiol. 2015, 169, 1991–2005. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, M.; Gao, S.; Zhang, Z.; Zhao, X.; Zhao, C.; Zhang, F.; Chen, X. Molecular Characterization of Novel TaNAC Genes in Wheat and Overexpression of TaNAC2a Confers Drought Tolerance in Tobacco. Physiol Plant. 2012, 144, 210–224. [Google Scholar] [CrossRef]
- Yan, X.; Yu, R.; Wang, J.; Jiao, Y. Ancestral Genome Reconstruction and the Evolution of Chromosomal Rearrangements in Triticeae. J. Genet. Genom. 2025, 52, 761–773. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Fuertes-Aguilar, J.; Matilla, A.J. Transcriptional Control of Seed Life: New Insights into the Role of the NAC Family. Int. J. Mol. Sci. 2024, 25, 5369. [Google Scholar] [CrossRef]
- Baloglu, M.C.; Oz, M.T.; Oktem, H.A.; Yucel, M. Expression Analysis of TaNAC69-1 and TtNAMB-2, Wheat NAC Family Transcription Factor Genes Under Abiotic Stress Conditions in Durum Wheat (Triticum turgidum). Plant Mol. Biol. Rep. 2012, 30, 1246–1252. [Google Scholar] [CrossRef]
- Wang, M.; Wang, M.; Zhao, M.; Wang, M.; Liu, S.; Tian, Y.; Moon, B.; Liang, C.; Li, C.; Shi, W.; et al. TaSRO1 Plays a Dual Role in Suppressing TaSIP1 to Fine Tune Mitochondrial Retrograde Signaling and Enhance Salinity Stress Tolerance. New Phytol. 2022, 236, 495–511. [Google Scholar] [CrossRef]
- Wang, M.; Yuan, J.; Qin, L.; Shi, W.; Xia, G.; Liu, S. TaCYP81D5, One Member in a Wheat Cytochrome P450 Gene Cluster, Confers Salinity Tolerance via Reactive Oxygen Species Scavenging. Plant Biotechnol. J. 2020, 18, 791–804. [Google Scholar] [CrossRef]
- Jung, J.K.H.; McCouch, S. Getting to the Roots of It: Genetic and Hormonal Control of Root Architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef]
- Zheng, L.; Hu, Y.; Yang, T.; Wang, Z.; Wang, D.; Jia, L.; Xie, Y.; Luo, L.; Qi, W.; Lv, Y.; et al. A Root Cap-Localized NAC Transcription Factor Controls Root Halotropic Response to Salt Stress in Arabidopsis. Nat. Commun. 2024, 15, 2061. [Google Scholar] [CrossRef]
- Wang, B.; Wei, J.; Song, N.; Wang, N.; Zhao, J.; Kang, Z. A Novel Wheat NAC Transcription Factor, TaNAC30, Negatively Regulates Resistance of Wheat to Stripe Rust. J. Integr. Plant Biol. 2018, 60, 432–443. [Google Scholar] [CrossRef]
- Qiao, L.; Li, Y.; Wang, L.; Gu, C.; Luo, S.; Li, X.; Yan, J.; Lu, C.; Chang, Z.; Gao, W.; et al. Identification of Salt-Stress-Responding Genes by Weighted Gene Correlation Network Analysis and Association Analysis in Wheat Leaves. Plants 2024, 13, 2642. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, L.; Liu, Y.; Shen, X.; Guo, Y.; Ma, X.; Zhang, X.; Li, X.; Cheng, T.; Wen, H.; et al. RNA-Seq-Based WGCNA and Association Analysis Reveal the Key Regulatory Module and Genes Responding to Salt Stress in Wheat Roots. Plants 2024, 13, 274. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, A.; Chen, J.; Wu, L.; Li, T.; Qiao, L. Identification of Ethylene Response Factors in Wheat Reveals That TaERF16-B Contributes to Salt Tolerance. Plants 2025, 14, 621. [Google Scholar] [CrossRef]
- Qiao, L.; Zhang, X.; Li, X.; Yang, Z.; Li, R.; Jia, J.; Yan, L.; Chang, Z. Genetic Incorporation of Genes for the Optimal Plant Architecture in Common Wheat. Mol. Breed. 2022, 42, 66. [Google Scholar] [CrossRef]
- Qiao, L.; Li, T.; Liu, S.; Zhang, X.; Fan, M.; Zhang, X.; Li, X.; Yang, Z.; Jia, J.; Qiao, L.; et al. Ali-A1 and TPL1 Proteins Interactively Modulate Awn Development in Wheat. Crop J. 2025, 13, 468–479. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme 1. | Chromosome | Position (IWGSC v1.0) | TaNAC Genes | MAF | RSIR | q Value |
|---|---|---|---|---|---|---|
| 1430[A/T] | 1B | 478581430 | TaNAC3-B | 0.26 | RD | 5.11 × 10−5 |
| 0779[A/G] | 2D | 639820779 | TaNAC26-D3.2 | 0.11 | TRL | 4.42 × 10−5 |
| RV | 1.67 × 10−6 | |||||
| 2830[A/G] | 3B | 199702830 | TaNAC31-B1 | 0.42 | RSA | 5.16 × 10−5 |
| RD | 5.73 × 10−5 | |||||
| 3581[G/A] | 3B | 214283581 | TaNAC32-B | 0.11 | RV | 1.71 × 10−6 |
| RSA | 1.64 × 10−5 | |||||
| RTN | 1.91 × 10−5 | |||||
| RD | 1.45 × 10−5 | |||||
| 4718[G/T] | 3B | 244384718 | TaNAC33-B | 0.16 | RBN | 9.14 × 10−5 |
| 5088[T/C] | 3B | 647245088 | TaNAC40-B | 0.11 | RTN | 2.09 × 10−5 |
| Gene Name in This Study | Reported Names in References |
|---|---|
| TaNAC9-A6 | TaNAC29 [28] |
| TaNAC68-D2 | TaNAC29 [29] |
| TaNAC15 | TaRNAC1 [33], TaNAC14 [34], TaNAC100 [35] |
| TaNAC106 | TaNAC1 [36], TaNAC-S [37] |
| TaNAC66-D | TaNAC5D-2 [38], TaNACL-D1 [39] |
| TaNAC85-A | NAM-A1 [40], TaSNAC8-6A [41] |
| TaNAC68 | TaNAC69 [42], TaNAC29 [29] |
| TaNAC80 | TaNAC2-5A [43], TaNAC2 [27], TaNAC2a [44], TaNAC2D [30] |
| TaNAC83 | TaNAC47 [31], TaNAC67 [32] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Wei, A.; Wang, W.; Zhang, X.; Zhao, Z.; Qiao, L. Identification of Salt Tolerance-Related NAC Genes in Wheat Roots Based on RNA-Seq and Association Analysis. Plants 2025, 14, 2318. https://doi.org/10.3390/plants14152318
Zhang L, Wei A, Wang W, Zhang X, Zhao Z, Qiao L. Identification of Salt Tolerance-Related NAC Genes in Wheat Roots Based on RNA-Seq and Association Analysis. Plants. 2025; 14(15):2318. https://doi.org/10.3390/plants14152318
Chicago/Turabian StyleZhang, Lei, Aili Wei, Weiwei Wang, Xueqi Zhang, Zhiyong Zhao, and Linyi Qiao. 2025. "Identification of Salt Tolerance-Related NAC Genes in Wheat Roots Based on RNA-Seq and Association Analysis" Plants 14, no. 15: 2318. https://doi.org/10.3390/plants14152318
APA StyleZhang, L., Wei, A., Wang, W., Zhang, X., Zhao, Z., & Qiao, L. (2025). Identification of Salt Tolerance-Related NAC Genes in Wheat Roots Based on RNA-Seq and Association Analysis. Plants, 14(15), 2318. https://doi.org/10.3390/plants14152318