
Salt Stress Responses of Different Rice Varieties at Panicle Initiation: Agronomic Traits, Photosynthesis, and Antioxidants

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Determination of Tiller Number, Plant Height, and Aboveground Dry Weight (ADW)
2.4. Canopy Interception Rate (CIR) and Leaf Area Index (LAI)
2.5. Leaf SPAD Value
2.6. Parameters of Photosynthetic Characteristics
2.7. Determination of Activities of Antioxidant Enzymes, H2O2 Content, and Malondialdehyde (MDA) Content
2.8. Data Analysis
3. Results
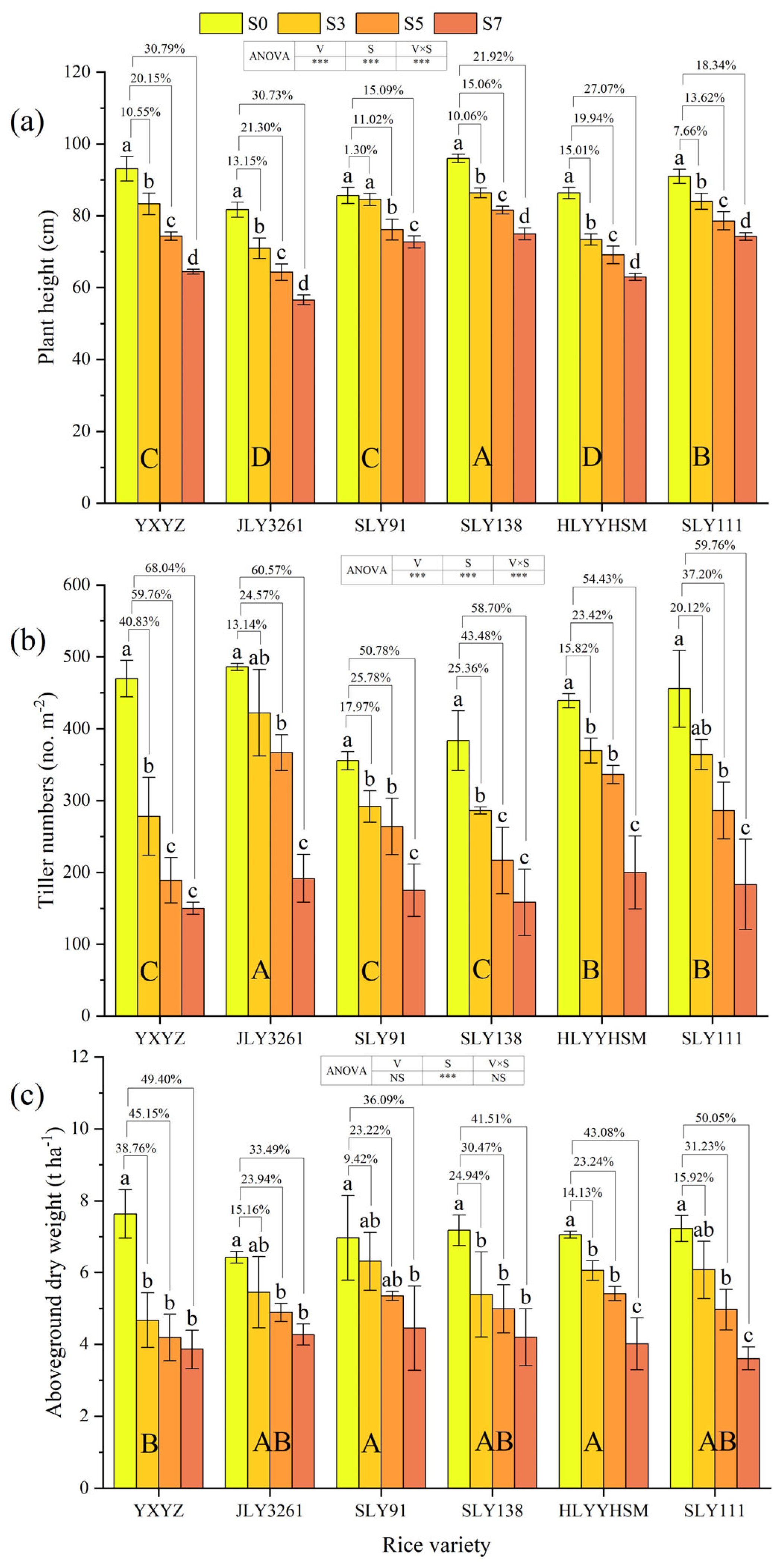
3.1. Plant Height, Tiller Numbers, and Aboveground Dry Weight
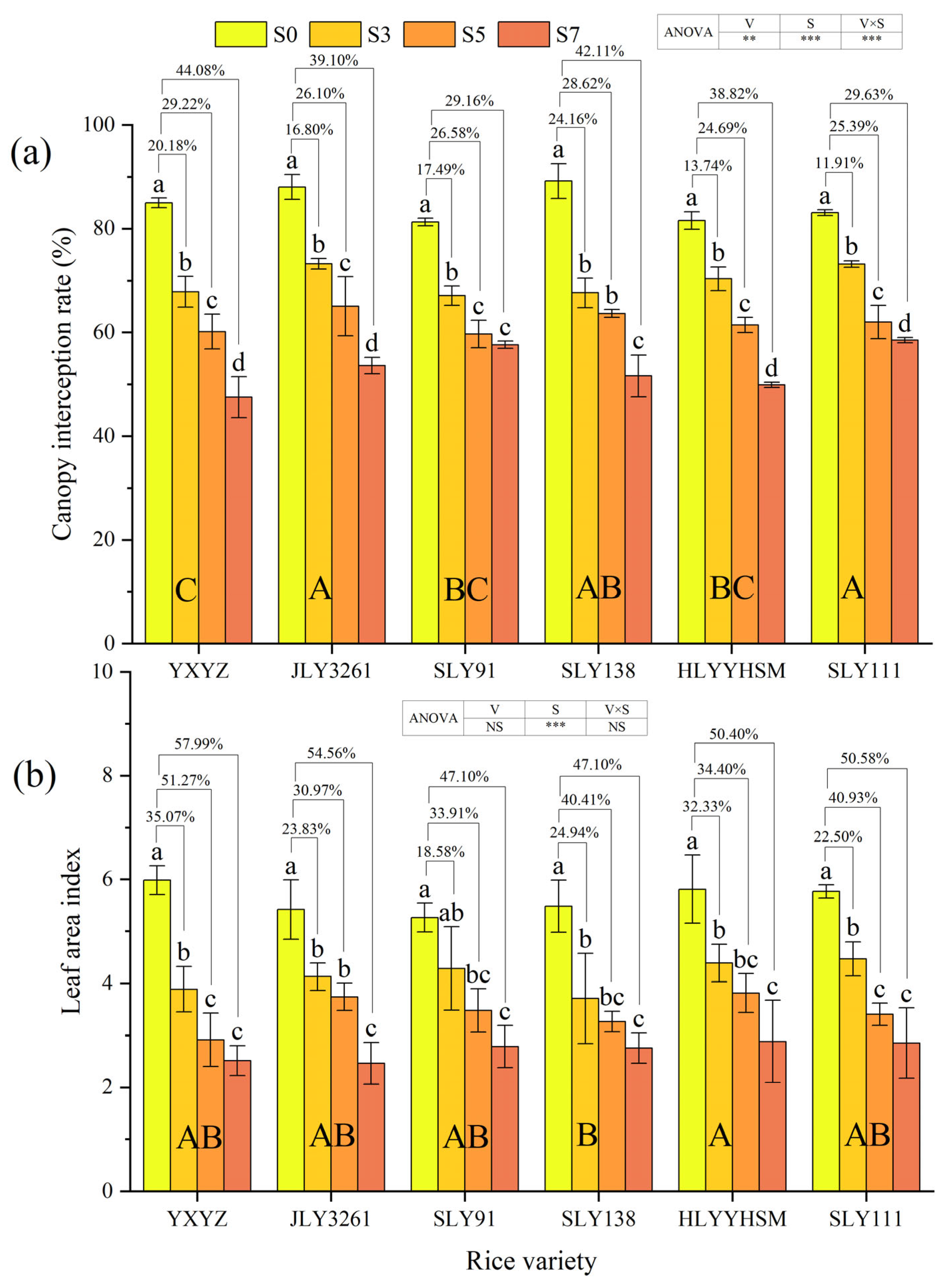
3.2. Canopy Interception Rate and Leaf Area Index
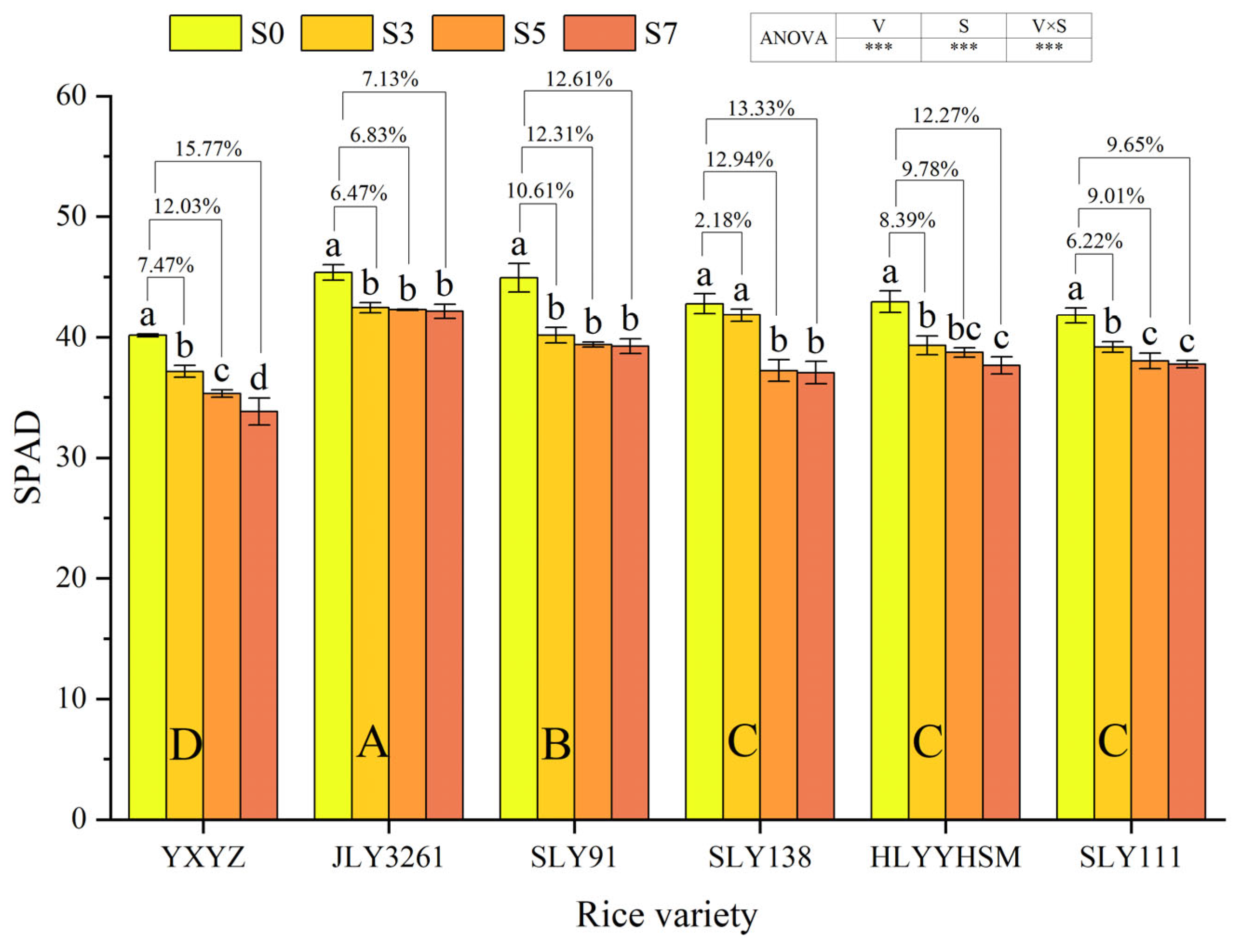
3.3. Leaf SPAD Value
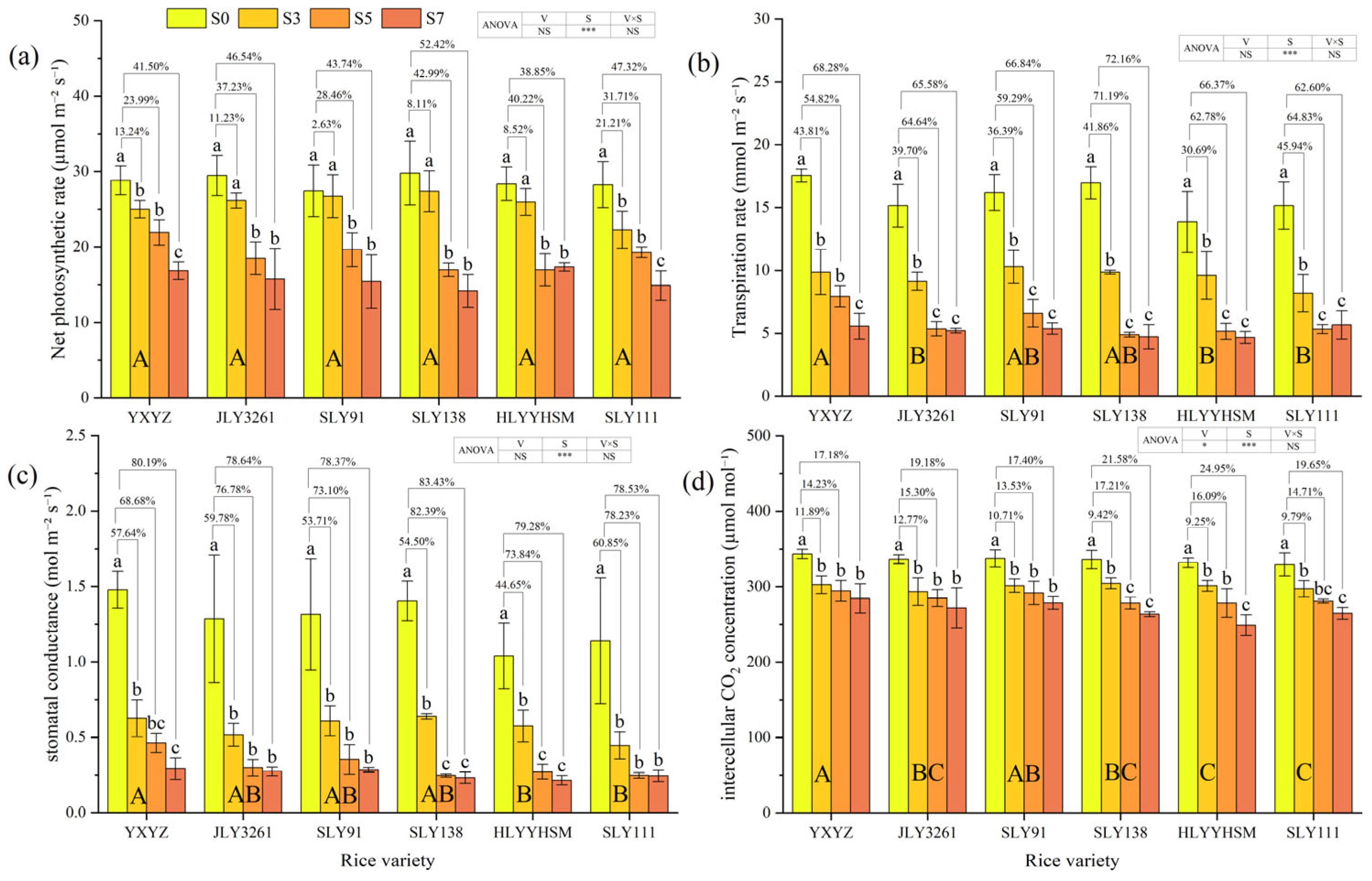
3.4. Parameters of Photosynthetic Characteristics
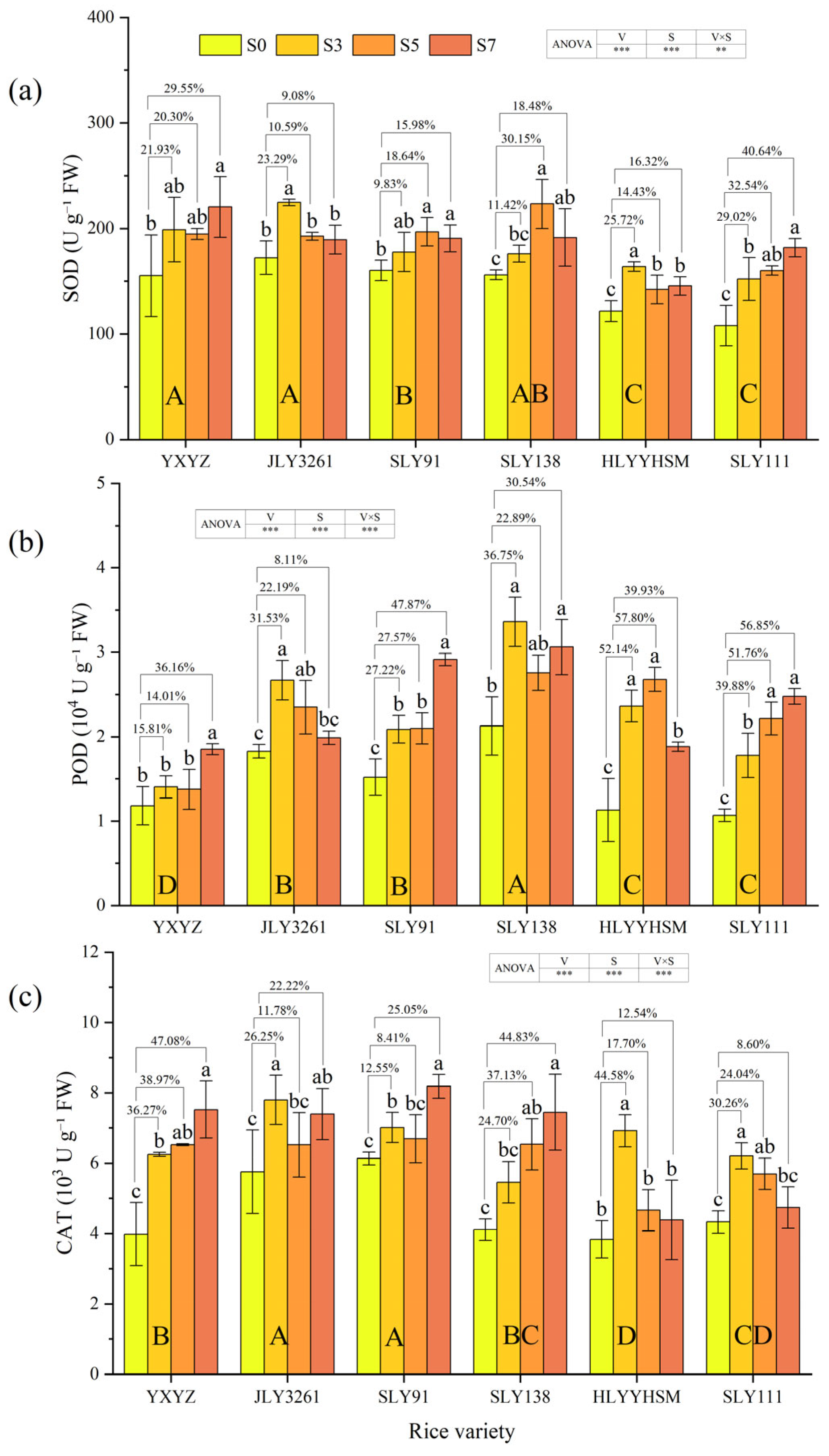
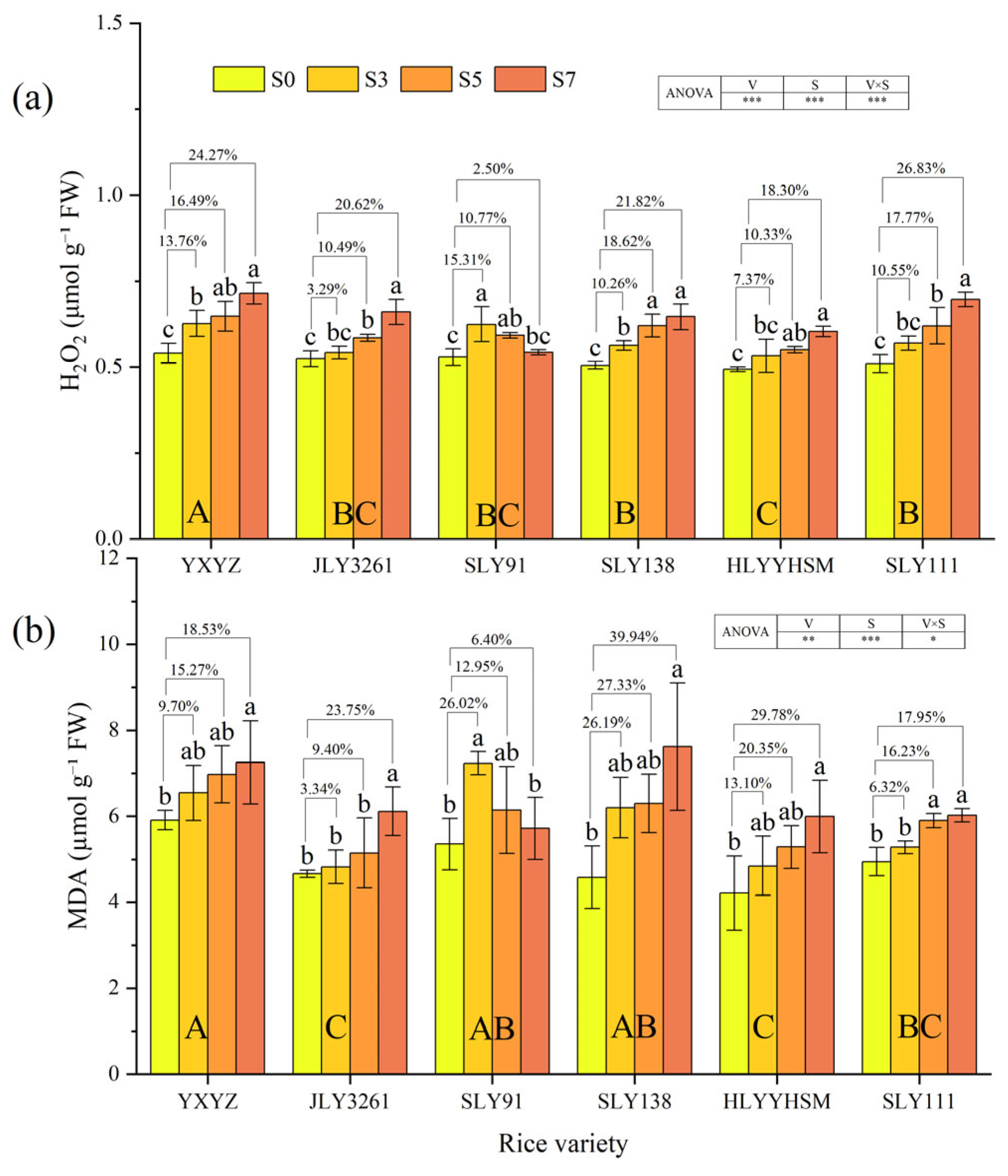
3.5. Activities of Antioxidant Enzymes, H2O2 Content, and MDA Content
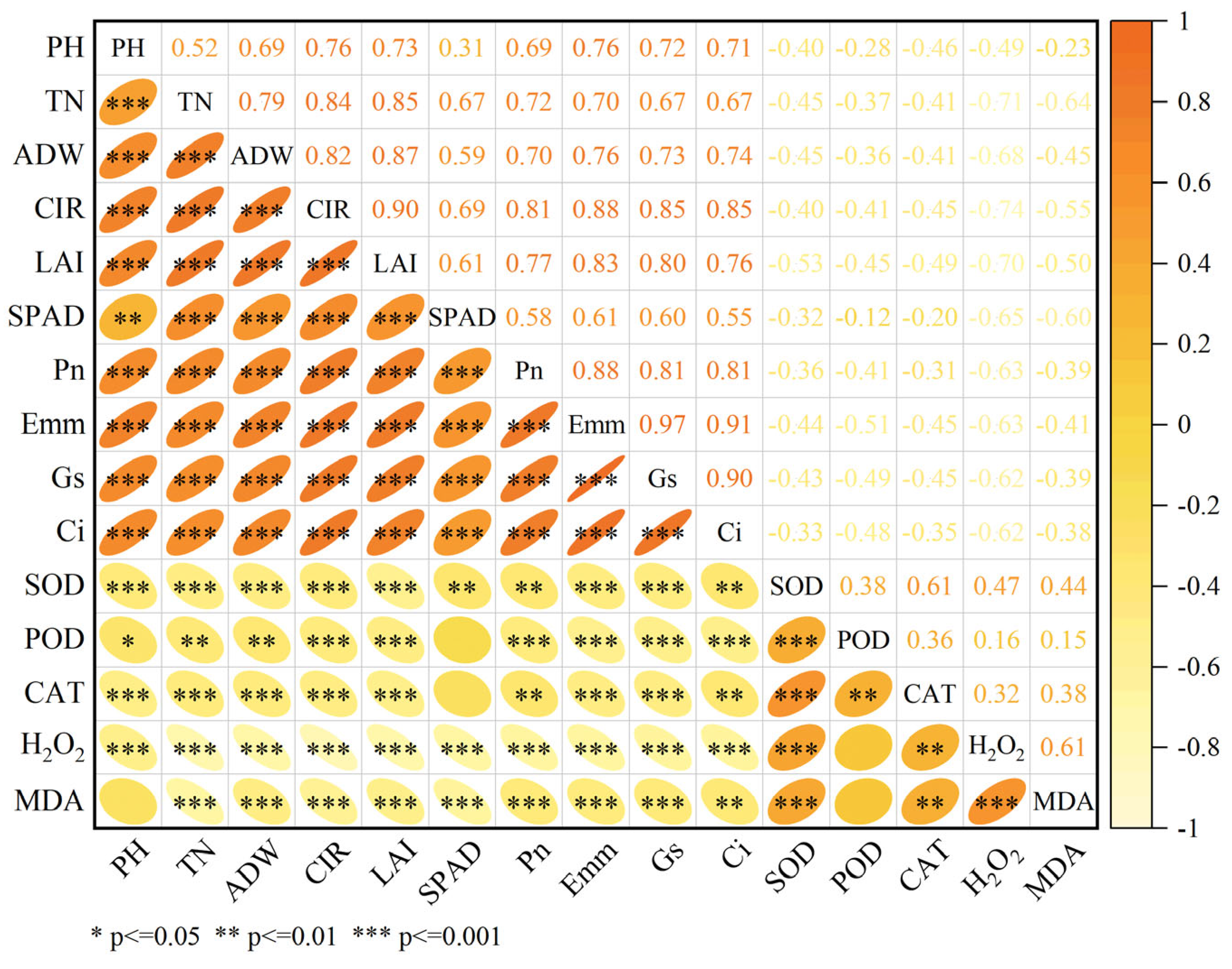
3.6. Correlation Analysis of Agronomic Traits, Photosynthetic Indices, and Antioxidant Capacity in Rice
4. Discussion
4.1. Differential Effects of Salt Stress on Agronomic Traits
4.2. The Response Mechanism of Photosynthetic Capacity to Salt Stress
4.3. Effect of Salt Stress on Antioxidant Levels in Different Salt Varieties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, L.; Emanuel, K.; Quiring, S.M. Elevated Risk of Tropical Cyclone Precipitation and Pluvial Flood in Houston under Global Warming. Environ. Res. Lett. 2021, 16, 94030. [Google Scholar] [CrossRef]
- Eswar, D.; Karuppusamy, R.; Chellamuthu, S. Drivers of Soil Salinity and Their Correlation with Climate Change. Curr. Opin. Environ. Sustain. 2021, 50, 310–318. [Google Scholar] [CrossRef]
- Wang, R.; Li, L.; Gentine, P.; Zhang, Y.; Chen, J.; Chen, X.; Chen, L.; Ning, L.; Yuan, L.; Lü, G. Recent Increase in the Observation-Derived Land Evapotranspiration Due to Global Warming. Environ. Res. Lett. 2022, 17, 24020. [Google Scholar] [CrossRef]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Kempen, B.; de Sousa, L. Global Mapping of Soil Salinity Change. Remote Sens. Environ. 2019, 231, 111260. [Google Scholar] [CrossRef]
- FAO. Global Status of Salt-Affected Soils—Main Report; FAO: Rome, Italy, 2024. [Google Scholar]
- Devkota, K.P.; Devkota, M.; Rezaei, M.; Oosterbaan, R. Managing Salinity for Sustainable Agricultural Production in Salt-Affected Soils of Irrigated Drylands. Agric. Syst. 2022, 198, 103390. [Google Scholar] [CrossRef]
- Ganie, S.A.; Molla, K.A.; Henry, R.J.; Bhat, K.V.; Mondal, T.K. Advances in Understanding Salt Tolerance in Rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, Z.; Zhang, Y.; Mu, Y.; Li, Y.; Nie, L. Regulation of 2-Acetyl-1-Pyrroline (2-AP) Biosynthesis and Grain Quality in Fragrant Rice under Salt Stress. F. Crop. Res. 2025, 322, 109747. [Google Scholar] [CrossRef]
- Yan, F.; Wei, H.; Li, W.; Liu, Z.; Tang, S.; Chen, L.; Ding, C.; Jiang, Y.; Ding, Y.; Li, G. Melatonin Improves K+ and Na+ Homeostasis in Rice under Salt Stress by Mediated Nitric Oxide. Ecotoxicol. Environ. Saf. 2020, 206, 111358. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and Salt Tolerance: Bringing Them Together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- MILLER, G.A.D.; SUZUKI, N.; CIFTCI-YILMAZ, S.; MITTLER, R.O.N. Reactive Oxygen Species Homeostasis and Signalling during Drought and Salinity Stresses. Plant. Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Fricke, W.; Akhiyarova, G.; Wei, W.; Alexandersson, E.; Miller, A.; Kjellbom, P.O.; Richardson, A.; Wojciechowski, T.; Schreiber, L.; Veselov, D.; et al. The Short-Term Growth Response to Salt of the Developing Barley Leaf. J. Exp. Bot. 2006, 57, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. The Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Sirajuddin; Zhao, X.Q.; Javed, M.T.; Khan, K.S.; Bano, A.; Shen, R.F.; Masood, S. Bacillus Pumilus Enhances Tolerance in Rice (Oryza sativa L.) to Combined Stresses of NaCl and High Boron Due to Limited Uptake of Na+. Environ. Exp. Bot. 2016, 124, 120–129. [Google Scholar] [CrossRef]
- Juan, M.; Rivero, R.M.; Romero, L.; Ruiz, J.M. Evaluation of Some Nutritional and Biochemical Indicators in Selecting Salt-Resistant Tomato Cultivars. Environ. Exp. Bot. 2005, 54, 193–201. [Google Scholar] [CrossRef]
- Ji, J.; Zhang, J.; Wang, X.; Song, W.; Ma, B.; Wang, R.; Li, T.; Wang, G.; Guan, C.; Gao, X. The alleviation of salt stress on rice through increasing photosynthetic capacity, maintaining redox homeostasis and regulating soil enzyme activities by Enterobacter sp. JIV1 assisted with putrescine. Microbiol. Res. 2024, 280, 127590. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Liu, J.; Meng, W.; Sun, Z.; Tan, Y.; Liu, Y.; Tan, M.; Wang, B.; Yang, J. Integrated Analysis of Transcriptome and Metabolome Reveals Molecular Mechanisms of Rice with Different Salinity Tolerances. Plants 2023, 12, 3359. [Google Scholar] [CrossRef] [PubMed]
- Jian, G.; Mo, Y.; Hu, Y.; Huang, Y.; Ren, L.; Zhang, Y.; Hu, H.; Zhou, S.; Liu, G.; Guo, J.; et al. Variety-Specific Transcriptional and Alternative Splicing Regulations Modulate Salt Tolerance in Rice from Early Stage of Stress. Rice 2022, 15, 56. [Google Scholar] [CrossRef] [PubMed]
- Taleisnik, E.; Rodríguez, A.A.; Bustos, D.; Erdei, L.; Ortega, L.; Senn, M.E. Leaf Expansion in Grasses under Salt Stress. J. Plant Physiol. 2009, 166, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Hakim, M.A.; Juraimi, A.S.; Hanafi, M.M.; Ismail, M.R.; Selamat, A.; Rafii, M.Y.; Latif, M.A. Biochemical and Anatomical Changes and Yield Reduction in Rice (Oryza sativa L.) under Varied Salinity Regimes. Biomed Res. Int. 2014, 2014, 208584. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Shabala, S.; Niu, Y.; Chen, Z.H.; Shabala, L.; Meinke, H.; Venkataraman, G.; Pareek, A.; Xu, J.; Zhou, M. Molecular Mechanisms of Salinity Tolerance in Rice. Crop J. 2021, 9, 506–520. [Google Scholar] [CrossRef]
- Li, Y.; Ai, Z.; Mu, Y.; Zhao, T.; Zhang, Y.; Li, L.; Huang, Z.; Nie, L.; Khan, M.N. Rice Yield Penalty and Quality Deterioration Is Associated with Failure of Nitrogen Uptake from Regreening to Panicle Initiation Stage under Salinity. Front. Plant Sci. 2023, 14, 1120755. [Google Scholar] [CrossRef] [PubMed]
- Rasel, M.; Tahjib-Ul-Arif, M.; Hossain, M.A.; Sayed, M.A.; Hassan, L. Discerning of Rice Landraces (Oryza Sativa L.) for Morpho-Physiological, Antioxidant Enzyme Activity, and Molecular Markers’ Responses to Induced Salt Stress at the Seedling Stage. J. Plant Growth Regul. 2020, 39, 41–59. [Google Scholar] [CrossRef]
- Li, R.; Guo, X.; Qi, Y.; Wang, Y.; Wang, J.; Zhang, P.; Cheng, S.; He, W.; Zhao, T.; Li, Y.; et al. Soil Amendments and Slow-Release Urea Improved Growth, Physiological Characteristics, and Yield of Salt-Tolerant Rice Under Salt Stress Conditions. Plants 2025, 14, 543. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wang, J.; Li, R.; Zhang, P.; Guo, X.; Qi, Y.; Li, Y.; Cheng, S.; Ji, J.; He, A.; et al. Effects of Nitrogen Fertilizer Types and Planting Density on the Yield and Nitrogen Use Efficiency of Salt-Tolerant Rice Under Salt Stress Conditions. Plants 2025, 14, 501. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt Tolerance in Rice: Physiological Responses and Molecular Mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Zhang, J.-L.; Flowers, T.J.; Wang, S.-M. Mechanisms of Sodium Uptake by Roots of Higher Plants. Plant Soil 2010, 326, 45–60. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Li, Y.; Huang, R. Advances and Challenges in the Breeding of Salt-Tolerant Rice. Int. J. Mol. Sci. 2020, 21, 8385. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, M.; Tian, J.; Xiao, F.; Xu, S.; Zuo, W.; Zhang, W. Improved Photosynthetic Capacity during the Mid- and Late Reproductive Stages Contributed to Increased Cotton Yield across Four Breeding Eras in Xinjiang, China. F. Crop. Res. 2019, 240, 177–184. [Google Scholar] [CrossRef]
- Kamran, M.; Ahmad, S.; Ahmad, I.; Hussain, I.; Meng, X.; Zhang, X.; Javed, T.; Ullah, M.; Ding, R.; Xu, P.; et al. Paclobutrazol Application Favors Yield Improvement of Maize Under Semiarid Regions by Delaying Leaf Senescence and Regulating Photosynthetic Capacity and Antioxidant System During Grain-Filling Stage. Agronomy 2020, 10, 187. [Google Scholar] [CrossRef]
- Qin, H.; Huang, R. The Phytohormonal Regulation of Na+/K+ and Reactive Oxygen Species Homeostasis in Rice Salt Response. Mol. Breed. 2020, 40, 47. [Google Scholar] [CrossRef]
- Seo, D.H.; Seomun, S.; Choi, Y.D.; Jang, G. Root Development and Stress Tolerance in Rice: The Key to Improving Stress Tolerance without Yield Penalties. Int. J. Mol. Sci. 2020, 21, 1807. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Huang, J.; Peng, S.; Xiong, D. Diffusional Conductance to CO2 Is the Key Limitation to Photosynthesis in Salt-Stressed Leaves of Rice (Oryza sativa L.). Physiol. Plant. 2018, 163, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, Y.; Liu, X.; Korpelainen, H.; Li, C. Ammonium and Nitrate Affect Sexually Different Responses to Salt Stress in Populus Cathayana. Physiol. Plant. 2022, 174, e13626. [Google Scholar] [CrossRef] [PubMed]
- Ganganelli, I.; Galatro, A.; Gergoff Grozeff, G.E.; Bartoli, C.G.; Senn, M.E. Reactive Oxygen Species (ROS): Chemistry and Role in Plant Physiology. Oxyg. Nitrogen Sulfur Species Post-Harvest. Physiol. Hortic. Crop. 2024, 2024, 43–73. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Gong, H.-J.; Yin, J.-L. Role of Silicon in Mediating Salt Tolerance in Plants: A Review. Plants 2019, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Bilal, M.; Hassani, D.; Iqbal, H.M.N.; Wang, H.; Huang, D. Mitigation of Salt Stress in White Clover (Trifolium Repens) by Azospirillum Brasilense and Its Inoculation Effect. Bot. Stud. 2017, 58, 5. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive Oxygen Species, Abiotic Stress and Stress Combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Ali, A.; Safdar, L.B.; Zafar, M.M.; Rui, Y.; Shakeel, A.; Shaukat, A.; Ashraf, M.; Gong, W.; Yuan, Y. Salt Stress Induces Physiochemical Alterations in Rice Grain Composition and Quality. J. Food Sci. 2020, 85, 14–20. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Brief Name | Rice Type | Rice Parentage | Incubation Unit | Trait |
|---|---|---|---|---|---|
| Yuxiangyouzhan | YXYZ | Indica conventional rice | TY36/IR100 (♀) × IR100 (♂) | Rice Research Institute of Guangdong Academy of Agricultural Sciences | The control variety of the national salt-tolerant rice district test consortium, with a salt resistance lower than JLY3261 |
| Jingliangyou3261 | JLY3261 | Indica two-line hybrid rice | Jing196S (♀) × R3261 (♂) | Hunan Hybrid Rice Research Center; National Salt-Tolerant Rice Technology Innovation Center | High-quality disease-resistant and salt-tolerant hybrid rice |
| Shuangliangyou138 | SLY138 | Indica two-line hybrid rice | Shuang1S (♀) × Qianghui38 (♂) | Xike Agricultural Group Co., Ltd.; Hunan Hybrid Rice Research Center; Hunan Academy of Agricultural Sciences | High yield, but salt tolerance has not been reported |
| Shuangliangyou111 | SLY111 | Indica two-line hybrid rice | Shuang1S (♀) × Shuanghui11 (♂) | Xike Agricultural Group Co., Ltd.; Hunan Hybrid Rice Research Center; Hunan Academy of Agricultural Sciences | Not tolerant to rice blast, salt tolerance not reported |
| Shuangliangyou91 | SLY91 | Indica two-line hybrid rice | Shuang1S (♀) × Z91 (♂) | Hunan Hybrid Rice Research Center | / |
| Hualiangyouyuexiangsimiao | HLYYXSM | Indica two-line hybrid rice | Hua9S (♀) × Yuehesimiao (♂) | National Salt-Tolerant Rice Technology Innovation Center in Sanya; Hunan Hybrid Rice Research Center | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Xue, Y.; Guan, Z.; Wang, Z.; Hou, D.; Zhao, T.; Lu, X.; Qi, Y.; Hao, Y.; Liu, J.; et al. Salt Stress Responses of Different Rice Varieties at Panicle Initiation: Agronomic Traits, Photosynthesis, and Antioxidants. Plants 2025, 14, 2278. https://doi.org/10.3390/plants14152278
Li Y, Xue Y, Guan Z, Wang Z, Hou D, Zhao T, Lu X, Qi Y, Hao Y, Liu J, et al. Salt Stress Responses of Different Rice Varieties at Panicle Initiation: Agronomic Traits, Photosynthesis, and Antioxidants. Plants. 2025; 14(15):2278. https://doi.org/10.3390/plants14152278
Chicago/Turabian StyleLi, Yusheng, Yuxiang Xue, Zhuangzhuang Guan, Zhenhang Wang, Daijie Hou, Tingcheng Zhao, Xutong Lu, Yucheng Qi, Yanbo Hao, Jinqi Liu, and et al. 2025. "Salt Stress Responses of Different Rice Varieties at Panicle Initiation: Agronomic Traits, Photosynthesis, and Antioxidants" Plants 14, no. 15: 2278. https://doi.org/10.3390/plants14152278
APA StyleLi, Y., Xue, Y., Guan, Z., Wang, Z., Hou, D., Zhao, T., Lu, X., Qi, Y., Hao, Y., Liu, J., Li, L., Sultan, H., Guo, X., Ai, Z., & He, A. (2025). Salt Stress Responses of Different Rice Varieties at Panicle Initiation: Agronomic Traits, Photosynthesis, and Antioxidants. Plants, 14(15), 2278. https://doi.org/10.3390/plants14152278