
Precise Identification and Analysis of Maize Germplasm Resistance to Ear Rot Caused by Six Fusarium Species

 ,
, 
Abstract
1. Introduction
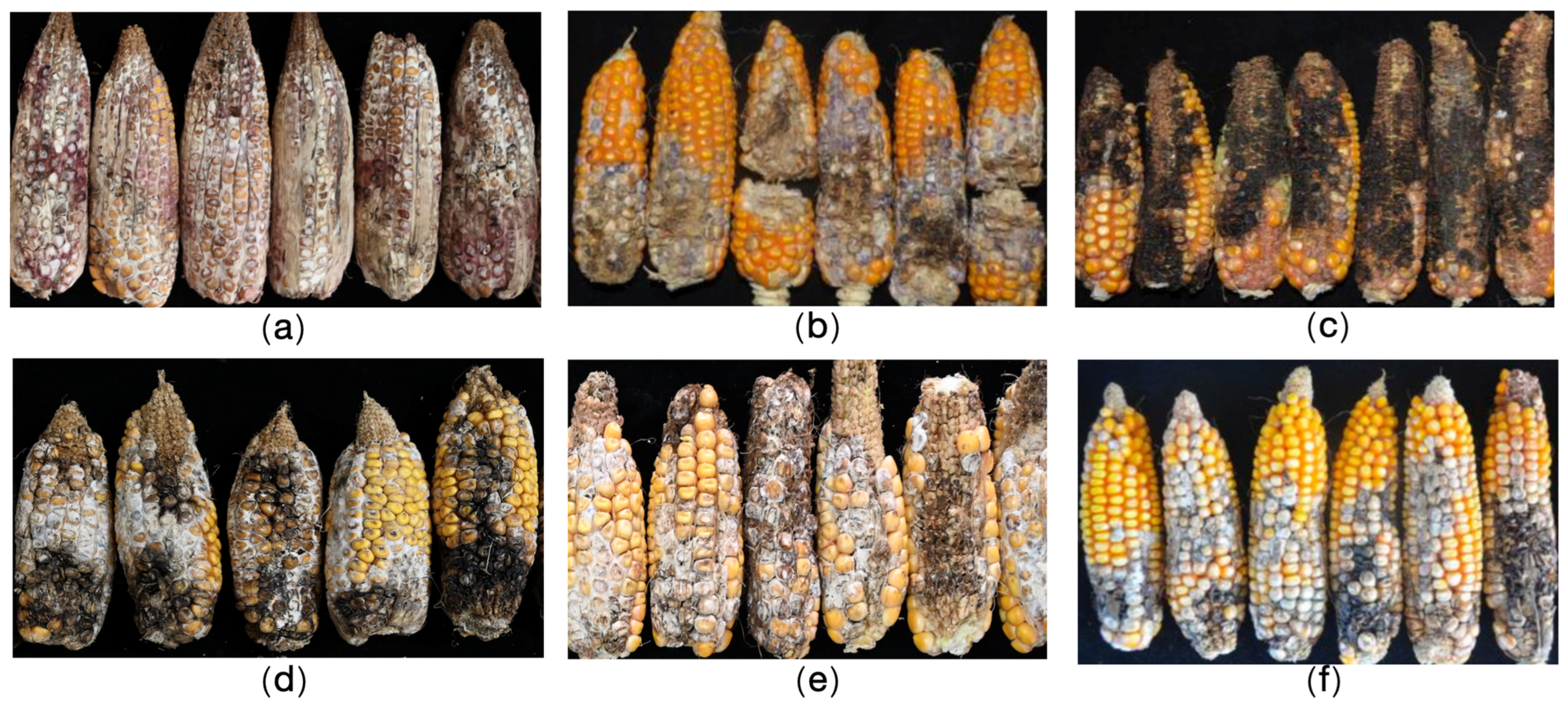
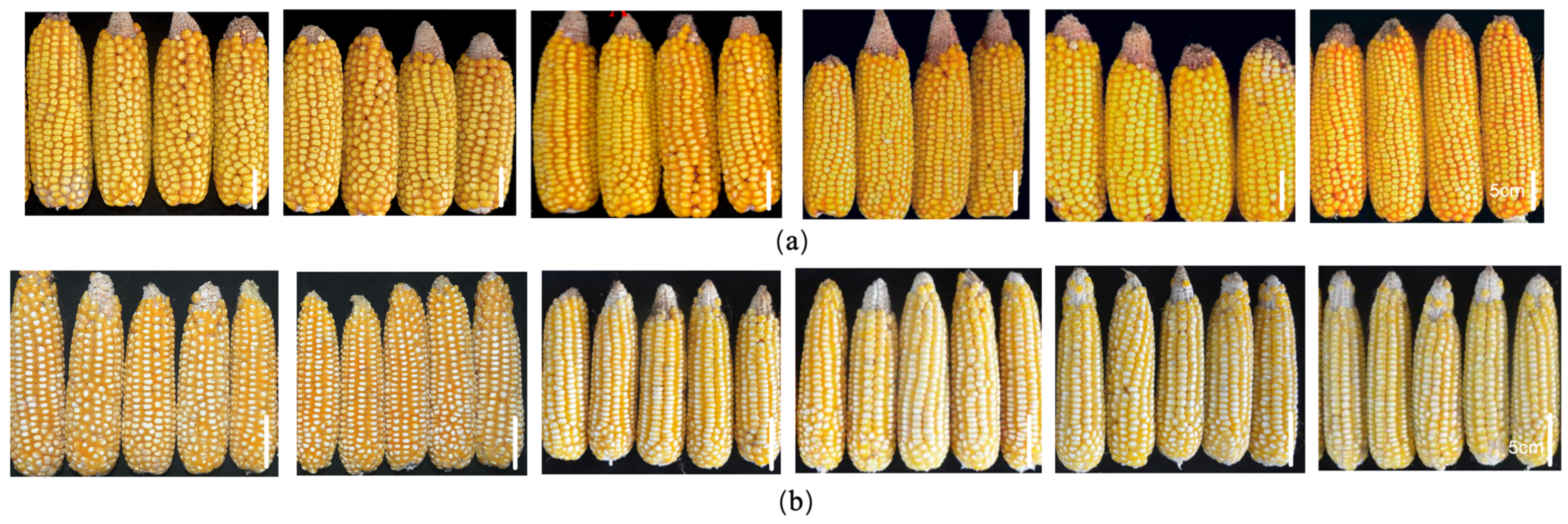
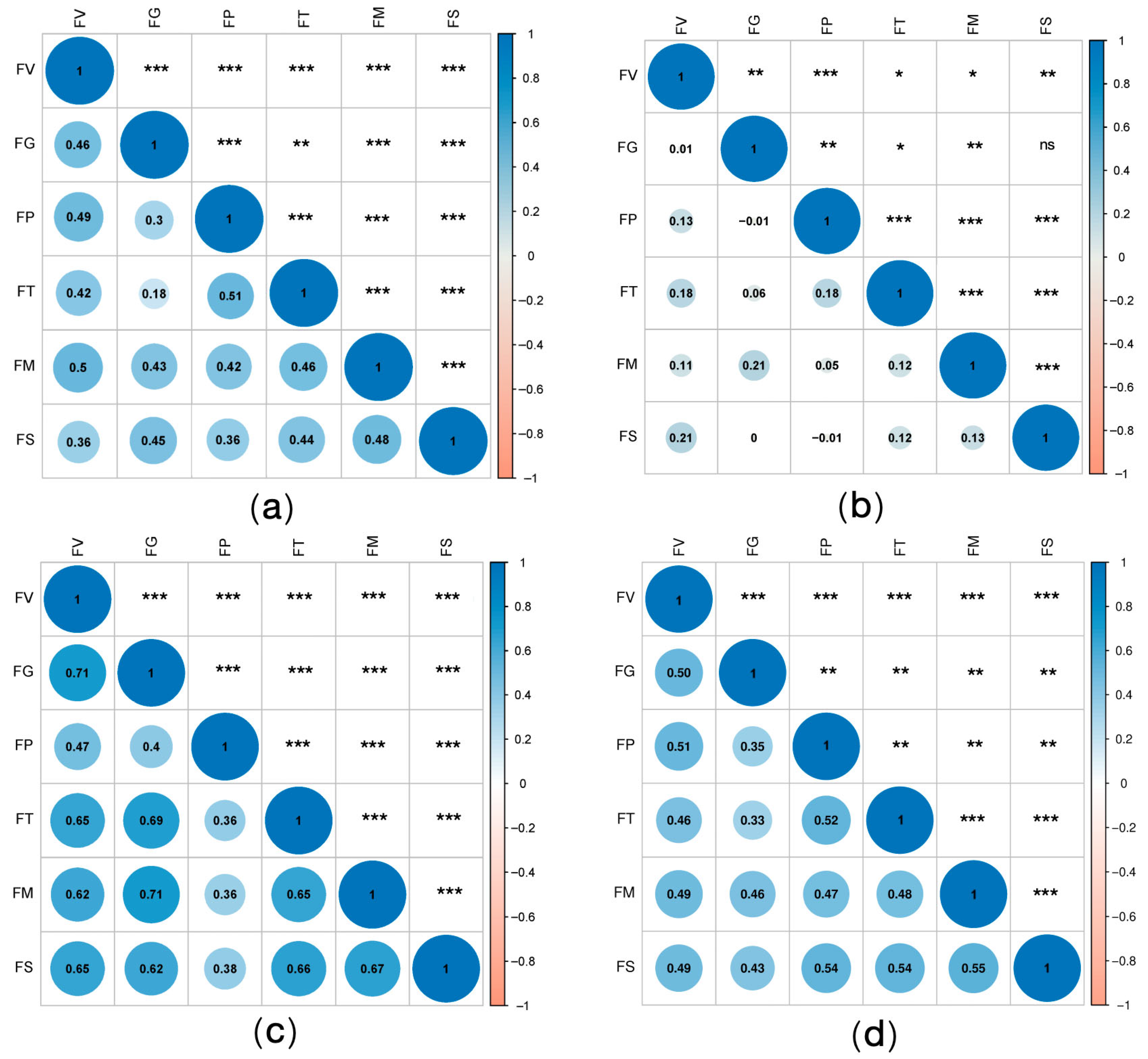
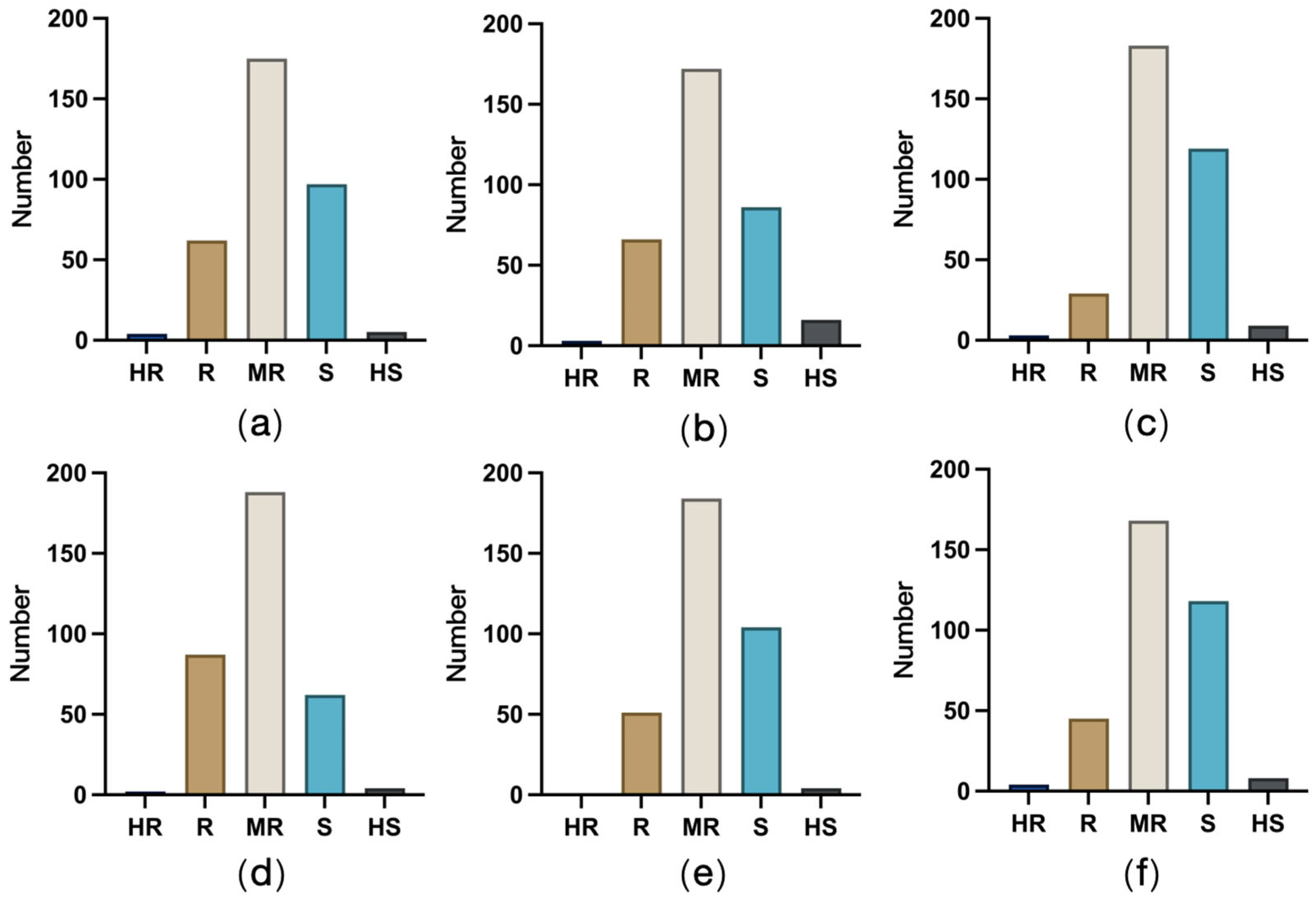
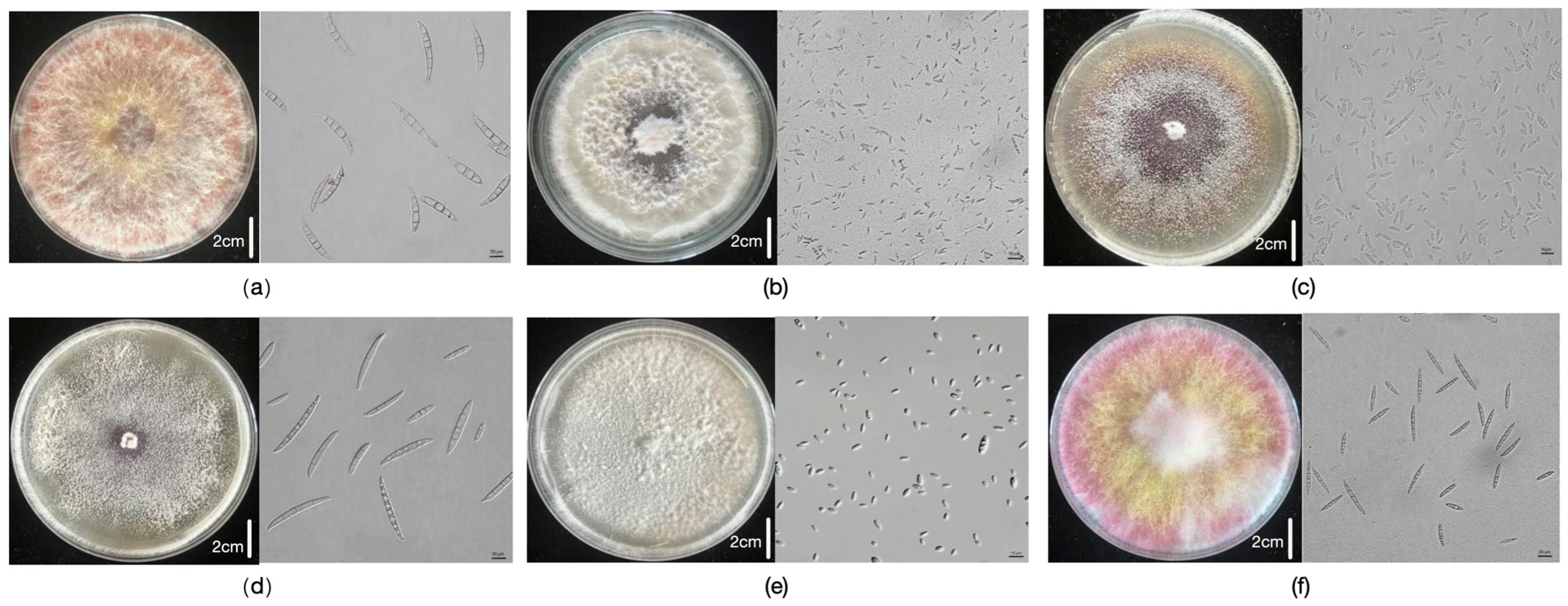
2. Results
3. Discussion
4. Materials and Methods
4.1. Test Materials
4.2. Pathogen and Inoculum Preparation
4.3. Disease Identification Plot
4.4. Inoculum Preparation
4.5. Inoculation Method
4.6. Disease Resistance Survey and Evaluation
4.7. Statistical Analysis of Genotypic Variance and G × E Interaction
4.8. Heterotic Group Classification
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Heterotic Group | Code | Heterotic Group | Code | Heterotic Group |
|---|---|---|---|---|---|
| K21HZD0021 | NSS | K21HZD1517 | Pioneer Female | K21HZD5457 | Pioneer Male B |
| K21HZD0044 | NSS | K21HZD1548 | Pioneer Female | K21HZD5461 | Pioneer Male B |
| K21HZD0071 | NSS | K21HZD1550 | Pioneer Female | K21HZD5970 | Pioneer Male B |
| K21HZD0187 | NSS | K21HZD1558 | Pioneer Female | K21HZD5973 | Pioneer Male B |
| K21HZD0271 | NSS | K21HZD1978 | Pioneer Female | K21HZD5976 | Pioneer Male B |
| K21HZD0275 | NSS | K21HZD1983 | Pioneer Female | K21HZD6054 | Pioneer Male B |
| K21HZD0375 | NSS | K21HZD2118 | Pioneer Female | K21HZD6057 | Pioneer Male B |
| K21HZD0394 | NSS | K21HZD2123 | Pioneer Female | K21HZD6063 | Pioneer Male B |
| K21HZD0395 | NSS | K21HZD2124 | Pioneer Female | K21HZD0012 | SS |
| K21HZD0401 | NSS | K21HZD2130 | Pioneer Female | K21HZD0105 | SS |
| K21HZD0402 | NSS | K21HZD2131 | Pioneer Female | K21HZD0452 | SS |
| K21HZD0482 | NSS | K21HZD2228 | Pioneer Female | K21HZD0611 | SS |
| K21HZD0566 | NSS | K21HZD2239 | Pioneer Female | K21HZD0700 | SS |
| K21HZD0751 | NSS | K21HZD2253 | Pioneer Female | K21HZD0736 | SS |
| K21HZD0759 | NSS | K21HZD2258 | Pioneer Female | K21HZD0762 | SS |
| K21HZD0766 | NSS | K21HZD2278 | Pioneer Female | K21HZD1623 | SS |
| K21HZD0777 | NSS | K21HZD2357 | Pioneer Female | K21HZD1693 | SS |
| K21HZD0792 | NSS | K21HZD2365 | Pioneer Female | K21HZD1786 | SS |
| K21HZD0847 | NSS | K21HZD2415 | Pioneer Female | K21HZD1971 | SS |
| K21HZD0850 | NSS | K21HZD2588 | Pioneer Female | K21HZD1985 | SS |
| K21HZD1131 | NSS | K21HZD2598 | Pioneer Female | K21HZD2127 | SS |
| K21HZD1133 | NSS | K21HZD4687 | Pioneer Female | K21HZD2146 | SS |
| K21HZD1145 | NSS | K21HZD4698 | Pioneer Female | K21HZD2199 | SS |
| K21HZD1172 | NSS | K21HZD4705 | Pioneer Female | K21HZD2388 | SS |
| K21HZD1214 | NSS | K21HZD4731 | Pioneer Female | K21HZD2405 | SS |
| K21HZD1463 | NSS | K21HZD4757 | Pioneer Female | K21HZD2442 | SS |
| K21HZD1465 | NSS | K21HZD5217 | Pioneer Female | K21HZD2612 | SS |
| K21HZD1467 | NSS | K21HZD5267 | Pioneer Female | K21HZD3969 | SS |
| K21HZD1979 | NSS | K21HZD5274 | Pioneer Female | K21HZD4182 | SS |
| K21HZD2550 | NSS | K21HZD5279 | Pioneer Female | K21HZD4303 | SS |
| K21HZD4741 | NSS | K21HZD5290 | Pioneer Female | K21HZD4790 | SS |
| K21HZD5318 | NSS | K21HZD5291 | Pioneer Female | K21HZD5082 | SS |
| K21HZD5376 | NSS | K21HZD5375 | Pioneer Female | K21HZD5296 | SS |
| K21HZD5988 | NSS | K21HZD5946 | Pioneer Female | K21HZD5454 | SS |
| K21HZD0010 | PB | K21HZD5949 | Pioneer Female | K21HZD5650 | SS |
| K21HZD0018 | PB | K21HZD5952 | Pioneer Female | K21HZD0051 | TSPT |
| K21HZD0334 | PB | K21HZD5955 | Pioneer Female | K21HZD0103 | TSPT |
| K21HZD0471 | PB | K21HZD5958 | Pioneer Female | K21HZD0761 | TSPT |
| K21HZD0569 | PB | K21HZD5991 | Pioneer Female | K21HZD0844 | TSPT |
| K21HZD0715 | PB | K21HZD6006 | Pioneer Female | K21HZD0871 | TSPT |
| K21HZD0897 | PB | K21HZD6018 | Pioneer Female | K21HZD1179 | TSPT |
| K21HZD1854 | PB | K21HZD6021 | Pioneer Female | K21HZD1380 | TSPT |
| K21HZD1948 | PB | K21HZD6027 | Pioneer Female | K21HZD1410 | TSPT |
| K21HZD1967 | PB | K21HZD0074 | Pioneer Male A | K21HZD1429 | TSPT |
| K21HZD1993 | PB | K21HZD0095 | Pioneer Male A | K21HZD1485 | TSPT |
| K21HZD2220 | PB | K21HZD0224 | Pioneer Male A | K21HZD1499 | TSPT |
| K21HZD2417 | PB | K21HZD0236 | Pioneer Male A | K21HZD1634 | TSPT |
| K21HZD2473 | PB | K21HZD0279 | Pioneer Male A | K21HZD2154 | TSPT |
| K21HZD2475 | PB | K21HZD0341 | Pioneer Male A | K21HZD2177 | TSPT |
| K21HZD2595 | PB | K21HZD0397 | Pioneer Male A | K21HZD2243 | TSPT |
| K21HZD2596 | PB | K21HZD0398 | Pioneer Male A | K21HZD2423 | TSPT |
| K21HZD2779 | PB | K21HZD0444 | Pioneer Male A | K21HZD2425 | TSPT |
| K21HZD2817 | PB | K21HZD0511 | Pioneer Male A | K21HZD2429 | TSPT |
| K21HZD2839 | PB | K21HZD0562 | Pioneer Male A | K21HZD2439 | TSPT |
| K21HZD2844 | PB | K21HZD0691 | Pioneer Male A | K21HZD2509 | TSPT |
| K21HZD2879 | PB | K21HZD0731 | Pioneer Male A | K21HZD2555 | TSPT |
| K21HZD3076 | PB | K21HZD1027 | Pioneer Male A | K21HZD2556 | TSPT |
| K21HZD3156 | PB | K21HZD1132 | Pioneer Male A | K21HZD3945 | TSPT |
| K21HZD3435 | PB | K21HZD1155 | Pioneer Male A | K21HZD5342 | TSPT |
| K21HZD4242 | PB | K21HZD1344 | Pioneer Male A | K21HZD5629 | TSPT |
| K21HZD4690 | PB | K21HZD1970 | Pioneer Male A | K21HZD5961 | TSPT |
| K21HZD5166 | PB | K21HZD2373 | Pioneer Male A | K21HZD5964 | TSPT |
| K21HZD5459 | PB | K21HZD2374 | Pioneer Male A | K21HZD5967 | TSPT |
| K21HZD5982 | PB | K21HZD2376 | Pioneer Male A | K21HZD5979 | TSPT |
| K21HZD5985 | PB | K21HZD2377 | Pioneer Male A | K21HZD6042 | TSPT |
| K21HZD0057 | PH4CV | K21HZD4780 | Pioneer Male A | K21HZD6048 | TSPT |
| K21HZD0171 | PH4CV | K21HZD0025 | Pioneer Male B | 2021S0915 | |
| K21HZD0196 | PH4CV | K21HZD0068 | Pioneer Male B | 2021S0919 | |
| K21HZD0197 | PH4CV | K21HZD0072 | Pioneer Male B | 2021S0925 | |
| K21HZD0210 | PH4CV | K21HZD0214 | Pioneer Male B | 2021S0926 | |
| K21HZD0390 | PH4CV | K21HZD0512 | Pioneer Male B | 2021S0928 | |
| K21HZD0415 | PH4CV | K21HZD0526 | Pioneer Male B | 2021S0929 | |
| K21HZD0749 | PH4CV | K21HZD0730 | Pioneer Male B | 2021S0931 | |
| K21HZD0904 | PH4CV | K21HZD0836 | Pioneer Male B | 2021S0933 | |
| K21HZD1193 | PH4CV | K21HZD0842 | Pioneer Male B | 2021S0935 | |
| K21HZD1205 | PH4CV | K21HZD0852 | Pioneer Male B | 2021S0940 | |
| K21HZD1208 | PH4CV | K21HZD0857 | Pioneer Male B | 2021S0941 | |
| K21HZD1210 | PH4CV | K21HZD0866 | Pioneer Male B | 2021S0943 | |
| K21HZD1237 | PH4CV | K21HZD0868 | Pioneer Male B | 2021S0945 | |
| K21HZD1245 | PH4CV | K21HZD0885 | Pioneer Male B | 2021S0960 | |
| K21HZD1514 | PH4CV | K21HZD0898 | Pioneer Male B | 2021S0961 | |
| K21HZD1526 | PH4CV | K21HZD0899 | Pioneer Male B | 22NHRB001 | |
| K21HZD1617 | PH4CV | K21HZD0903 | Pioneer Male B | 22NHRB016 | |
| K21HZD1636 | PH4CV | K21HZD1149 | Pioneer Male B | 22NHRB039 | |
| K21HZD1637 | PH4CV | K21HZD1182 | Pioneer Male B | 22NHRB048 | |
| K21HZD1640 | PH4CV | K21HZD1183 | Pioneer Male B | 22NHRB066 | |
| K21HZD1944 | PH4CV | K21HZD1200 | Pioneer Male B | 22NHRB081 | |
| K21HZD1987 | PH4CV | K21HZD1217 | Pioneer Male B | 22NHRB105 | |
| K21HZD2300 | PH4CV | K21HZD1520 | Pioneer Male B | 22NHRB108 | |
| K21HZD2567 | PH4CV | K21HZD1573 | Pioneer Male B | 22NHRB155 | |
| K21HZD2578 | PH4CV | K21HZD1581 | Pioneer Male B | 22NHRB214 | |
| K21HZD2581 | PH4CV | K21HZD1590 | Pioneer Male B | 22NHRB315 | |
| K21HZD2602 | PH4CV | K21HZD1598 | Pioneer Male B | 22NHRB322 | |
| K21HZD2610 | PH4CV | K21HZD1615 | Pioneer Male B | CNH3323 | |
| K21HZD4732 | PH4CV | K21HZD1962 | Pioneer Male B | CY103 | |
| K21HZD0170 | Pioneer Female | K21HZD1965 | Pioneer Male B | CY105 | |
| K21HZD0226 | Pioneer Female | K21HZD2234 | Pioneer Male B | CY109 | |
| K21HZD0296 | Pioneer Female | K21HZD2244 | Pioneer Male B | K21HXY045 | |
| K21HZD0400 | Pioneer Female | K21HZD2260 | Pioneer Male B | K21HXY085 | |
| K21HZD0742 | Pioneer Female | K21HZD2274 | Pioneer Male B | K21HXY145 | |
| K21HZD0786 | Pioneer Female | K21HZD2350 | Pioneer Male B | K21HXY221 | |
| K21HZD0804 | Pioneer Female | K21HZD2371 | Pioneer Male B | K21HXY259 | |
| K21HZD0849 | Pioneer Female | K21HZD2398 | Pioneer Male B | K21HZD0276 | |
| K21HZD0861 | Pioneer Female | K21HZD2463 | Pioneer Male B | K21HZD0887 | |
| K21HZD0915 | Pioneer Female | K21HZD2482 | Pioneer Male B | K21HZD1353 | |
| K21HZD1261 | Pioneer Female | K21HZD2534 | Pioneer Male B | K21HZD1470 | |
| K21HZD1295 | Pioneer Female | K21HZD2553 | Pioneer Male B | K21HZD1521 | |
| K21HZD1302 | Pioneer Female | K21HZD2616 | Pioneer Male B | K21HZD1528 | |
| K21HZD1307 | Pioneer Female | K21HZD2618 | Pioneer Male B | K21HZD2219 | |
| K21HZD1309 | Pioneer Female | K21HZD3093 | Pioneer Male B | K21HZD4411 | |
| K21HZD1323 | Pioneer Female | K21HZD4339 | Pioneer Male B | K21HZD4653 | |
| K21HZD1370 | Pioneer Female | K21HZD4737 | Pioneer Male B | K21HZD4739 | |
| K21HZD1418 | Pioneer Female | K21HZD5309 | Pioneer Male B | K21HZD5266 | |
| K21HZD1511 | Pioneer Female | K21HZD5366 | Pioneer Male B | K21HZD5308 | |
| K21HZD5467 |
References
- Duan, C.X.; Wang, X.M.; Song, F.J.; Sun, S.L.; Zhou, D.N.; Zhu, Z.D. Advances in research on maize resistance to ear rot. Sci. Agric. Sin. 2015, 48, 2152–2164. [Google Scholar]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Song, L.Q.; Wei, L.M.; Wang, Z.Y.; He, K.L. Effect of infestation by the Asian corn borer together with Fusarium verticillioides on corn yield loss. Acta Phytophylacica Sin. 2009, 36, 6. [Google Scholar]
- Wang, B.B. Identification of Pathogenic Fusarium Species Causing Maize Ear Rot and Study on Host Resistance. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2020. [Google Scholar]
- Wang, X.M.; Shi, J.; Jin, Q.M.; Li, X.; Sun, S.X. The Field Visual of Maize Diseases and Pests; China Agriculture Science and Technology Press: Beijing, China, 2010; pp. 55–56. [Google Scholar]
- Bi, X.L. Research progress on resistance to fusarium infection in maize. South China Agric. 2023, 17, 237–240. [Google Scholar]
- Song, W.B.; Dong, H.F.; Chen, W.; Dai, X.D.; Liu, C.Y.; Li, J.J.; Wu, J.Y. Progress on maize ear rot. J. Henan Agric. Univ. 2005, 4, 3. [Google Scholar]
- Tan, L.P.; Liu, Y.; Xing, F.G. Removal of fumonisins from maize and maize products. Cereal Feed Ind. 2008, 8, 17–19. [Google Scholar]
- Duan, C.X.; Qin, Z.H.; Yang, Z.H.; Li, W.X.; Sun, S.L.; Zhu, Z.D. Identification of pathogenic Fusarium spp. causing maize ear rot and potential mycotoxin production in China. Toxins 2016, 8, 186. [Google Scholar] [CrossRef]
- Wang, X.C.; He, C.H.; Liu, H.M.; Zhang, H.B. Mycotoxin contamination, hazards, and detection technologies. Anim. Husb. Vet. Med. 2009, 41, 8. [Google Scholar]
- Li, H.; Xiang, K.; Zhang, Z.M.; Yuan, G.S.; Pan, G.T. Research progress on ear rot resistant mechanism and resistant breeding in maize. J. Maize Sci. 2019, 27, 167–174. [Google Scholar]
- Qin, Z.H.; Ren, X.; Jiang, K.; Wu, X.F.; Yang, Z.H.; Wang, X.M. Identification of Fusarium species and F. graminearum species complex causing maize ear rot in China. Acta Phytophylacica Sin. 2014, 41, 589–596. [Google Scholar]
- Ren, X. Diversity Analyses of Fusarium spp. the Main Causal Agents of Maize Ear Rot in China. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2011. [Google Scholar]
- Zhou, D.N.; Wang, X.M.; Li, D.D.; Yang, Y. Isolation and identification of Fusarium species causing maize ear rot in Chongqing and its vicinity. J. Plant Prot. 2016, 43, 782–788. [Google Scholar]
- Sun, H.; Guo, N.; Shi, J. Characterization of the maize ear rot pathogens and genetic diversity analysis of dominant species in Hainan. Acta Phytophylacica Sin. 2017, 47, 577–583. [Google Scholar]
- Wang, B.B.; Bi, S.G.; Xiao, M.G. Isolation and identification of pathogenic Fusarium spp. causing maize ear rot and analysis of their toxin-producing genotype in Heilongjiang Province. Acta Prataculturae Sin. 2020, 29, 163–174. [Google Scholar]
- Chai, H.Y.; Jia, J.; Bai, X.; Meng, L.M.; Zhang, W.; Jin, R.; Wu, H.B.; Su, Q.F. Identification of pathogenic Fusarium spp. causing maize ear rot and susceptibility of some strains to fungicides in Jilin Province. Acta Agron. Sin. 2023, 56, 64–78. [Google Scholar]
- Chai, H.Y.; Pan, Y.Y.; Bai, X.; Meng, L.M.; Zhang, W.; Wu, H.B.; Wang, Y.S.; Gao, Y.B.; Jia, B.; Su, Q.F. Analysis of the relationship between aphid-damage and Fusarium composition, mycotoxin pollution on maize ears. Acta Agron. Sin. 2024, 57, 3171–3181. [Google Scholar]
- Wang, C.M.; Guo, C.; Luo, H.S.; Zhou, T.W.; Hong, L. Population structure of maize ear rot pathogens and genetic diversity of dominant pathogens in Gansu Province. Chin. J. Appl. Environ. Biol. 2025. [Google Scholar]
- Wang, B.B.; Guo, C.; Sun, S.L.; Xia, Y.S.; Zhu, Z.D.; Duan, C.X. The genetic diversity, pathogenicity, and toxigenic chemotypes of Fusarium graminearum species complex causing maize ear rot. Sci. Agric. Sin. 2020, 53, 4777–4790. [Google Scholar]
- Sun, H.; Zhang, H.J.; Ma, H.X.; Shi, J.; Guo, N.; Chen, D.; Li, P. Composition and distribution of pathogens causing ear rot in spring maize regionand identification of Fusarium graminearum species complex. Acta Phytopathol. Sin. 2018, 48, 8–15. [Google Scholar]
- Andriolli, C.F.; Casa, R.T.; Kuhnem, P.R.; Bogo, A.; Zancan, R.L.; Reis, E.M. Timing of fungicide application for the control of Gibberella ear rot of maize. Trop. Plant Pathol. 2016, 41, 264–269. [Google Scholar] [CrossRef]
- Duan, C.X.; Zhu, Z.D.; Wu, X.F.; Yang, Z.H.; Wang, X.M. Screening and evaluation of maize germplasm for resistance to five diseases and Asian corn borer. J. Plant Genet. Resour. 2012, 13, 169–174. [Google Scholar]
- Duan, C.X.; Wang, X.M.; Wu, X.F.; Yang, Z.H.; Song, F.J.; Zhao, L.P. Analysis of maize accessions resistance to Pythium stalk rot and Fusarium ear rot. J. Plant Genet. Resour. 2015, 16, 947–954. [Google Scholar]
- Guo, C.; Guo, M.K.; Wei, H.Y.; Guo, J.G. Identification of resistance of maize germplasm resources to ear rot. Acta Agric. Jiangxi 2015, 27, 50–52. [Google Scholar]
- Han, Y.C.; Wei, Y.J.; Zhang, Z.; Wang, T.Y. Large-scale identification of maize germplasm resources for resistance to ear rot caused by two Fusarium species. J. Plant Genet. Resour. 2024, 25, 1613–1626. [Google Scholar]
- Zhang, Y.; Zhang, Y.; Wang, Z.Y.; Wen, J.; Han, S.P.; Guo, J.; Xing, Y.X. Evaluation of resistance to Fusarium ear rot in 44 maize inbred lines. J. Plant Genet. Resour. 2019, 20, 276–283. [Google Scholar]
- Zhao, Z.Q.; Zhao, Y.Q.; Lin, C.P.; Zhao, Y.Z.; Yu, Y.X.; Meng, Q.L.; Zeng, G.Y.; Xue, J.Q.; Yang, Q. Precise evaluation of 48 maize inbred lines to major diseases. Sci. Agric. Sin. 2021, 54, 2510–2522. [Google Scholar]
- Duan, C.X.; Cui, L.N.; Xia, Y.S.; Dong, H.Y.; Yang, Z.H.; Hu, Q.Y.; Sun, S.L.; Li, X.; Zhu, Z.D. Precise characterization and analysis of maize germplasm resources for resistance to Fusarium ear rot and Gibberella ear rot. Acta Agron. Sin. 2022, 48, 2155–2167. [Google Scholar] [CrossRef]
- Xia, Y.S.; Guo, C.; Wen, S.H.; Sun, S.l.; Zhu, Z.D.; Duan, C.X. Identification of maize germplasm resistant to Fusarium ear rot and Gibberella ear rot and genetic diversity analysis of resistant lines. J. Plant Genet. Resour. 2022, 23, 61–71. [Google Scholar]
- Hou, M.; Wang, W.; Zhao, W.; Chen, Y. Research progress on identification of maize resistance to ear rot and mapping of resistance loci. Heilongjiang Agric. Sci. 2024, 7, 97–107. [Google Scholar]
- Zhang, T.; Sun, X.D.; Lv, G.Z. Fusarium species and its isolation frequency from rot ears of maize in northeast China. J. Fungal Res. 2011, 36, 9–14. [Google Scholar]
- Ding, M.J.; Yang, Y.; Sun, H.; Ma, H.X.; Liu, S.S.; Shi, J. Isolation and identification of maize ear rot pathogens and phylogenetic analysis of dominant species in Shandong province. Acta Agric. Boreali-Sin. 2019, 34, 216–223. [Google Scholar]
- Hu, Y.X.; Liu, Y.B.; Wang, H.; Jin, L.P.; Lin, F.; Zhang, X.C.; Zheng, H.J. Research progress on ear rot resistant genetics and breeding in maize. J. Maize Sci. 2021, 29, 171–178. [Google Scholar]
- Gong, L.; Deng, J.H.; Jiao, Q.; Liu, X.; Qiao, Z.H. Determination of indoor toxicity and field control effect of fungicides against maize ear rot. Plant Prot. 2022, 48, 374–381. [Google Scholar]
- Guo, Z.F.; Zou, C.; Liu, X.G.; Wang, S.; Li, W.X.; Jeffers, D. Complex genetic system involved in Fusarium ear rot resistance in maize as revealed by GWAS, bulked sample analysis, and genomic prediction. Plant Dis. 2020, 104, 1725–1735. [Google Scholar] [CrossRef] [PubMed]
- Gaikpa, D.S.; Kessel, B.; Presterl, T.; Ouzunova, M.; Galiano-Carneiro, A.L.; Mayer, M. Exploiting genetic diversity in two European maize landraces for improving Gibberella ear rot resistance using genomic tools. Theor. Appl. Genet. 2021, 134, 793–805. [Google Scholar] [CrossRef]
- Yin, Z.C.; Wang, X.F.; Long, Y.; Dong, Z.Y.; Wan, X.Y. Advances on genetic research and mechanism analysis on maize resistance to ear rot. China Biotechnol. 2021, 41, 103–115. [Google Scholar]
- Tembo, E.; Minnaar-Ontong, A.; Menkir, A.; Marais, G.; Magorokosho, C.; Labuschagne, M.T. Inheritance of resistance to Fusarium verticillioides ear rot in maize inbred lines of southern, West and central Africa origin. Crop Sci. 2022, 62, 1813–1833. [Google Scholar] [CrossRef]
- Lanubile, A.; Maschietto, V.; Borrelli, V.M.; Stagnati, L.; Logrieco, A.F.; Marocco, A. Molecular basis of resistance to Fusarium ear rot in maize. Front. Plant Sci. 2017, 8, 1774. [Google Scholar] [CrossRef]
- Loeffler, M.; Kessel, B.; Ouzunova, M.; Miedaner, T. Population parameters for resistance to Fusarium graminearum and Fusarium verticillioides ear rot among large sets of early, mid-late and late maturing European maize (Zea mays L.) inbred lines. Theor. Appl. Genet. 2010, 120, 1053–1062. [Google Scholar] [CrossRef]
- Presello, D.A.; Iglesias, J.; Botta, G.; Reid, L.M.; Lori, G.A.; Eyherabide, G.H. Stability of maize resistance to the ear rots caused by Fusarium graminearum and F. verticillioides in Argentinean and Canadian environments. Euphytica 2006, 147, 403–407. [Google Scholar] [CrossRef]
- Qu, Q.; Li, L.N.; Liu, J.; Wang, Z.X.; Cao, Z.Y.; Dong, J.G. Resistance evaluation of some commonly used maize germplasm resources to Fusarium diseases in China. Sci. Agric. Sin. 2019, 52, 2962–2971. [Google Scholar]
- Wang, L.X.; Xu, X.D.; Liu, Z.H.; Dong, H.Y.; Jiang, J.; Zhang, M.H. Inoculation technique and screening maize germplasm resistance to Fusarium ear rot. J. Plant Genet. Resour. 2007, 8, 145–148. [Google Scholar]
- Owen, H.; Dylan, H.; Colin, B.; Jeremy, B. The Ear Unwrapper: A maize ear image acquisition pipeline for disease severity phenotyping. AgriEngineering 2023, 5, 1216–1225. [Google Scholar] [CrossRef]
- Wen, J.; Shen, Y.Q.; Wang, Z.Y.; Li, S.J.; Mo, L.Y.; Lei, Y.H.; Zhang, Y.; Han, S.P. QTL mapping of resistance to Fusarium ear rot in maize based on image analysis. Sci. Agric. Sin. 2021, 54, 2724–2736. [Google Scholar]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
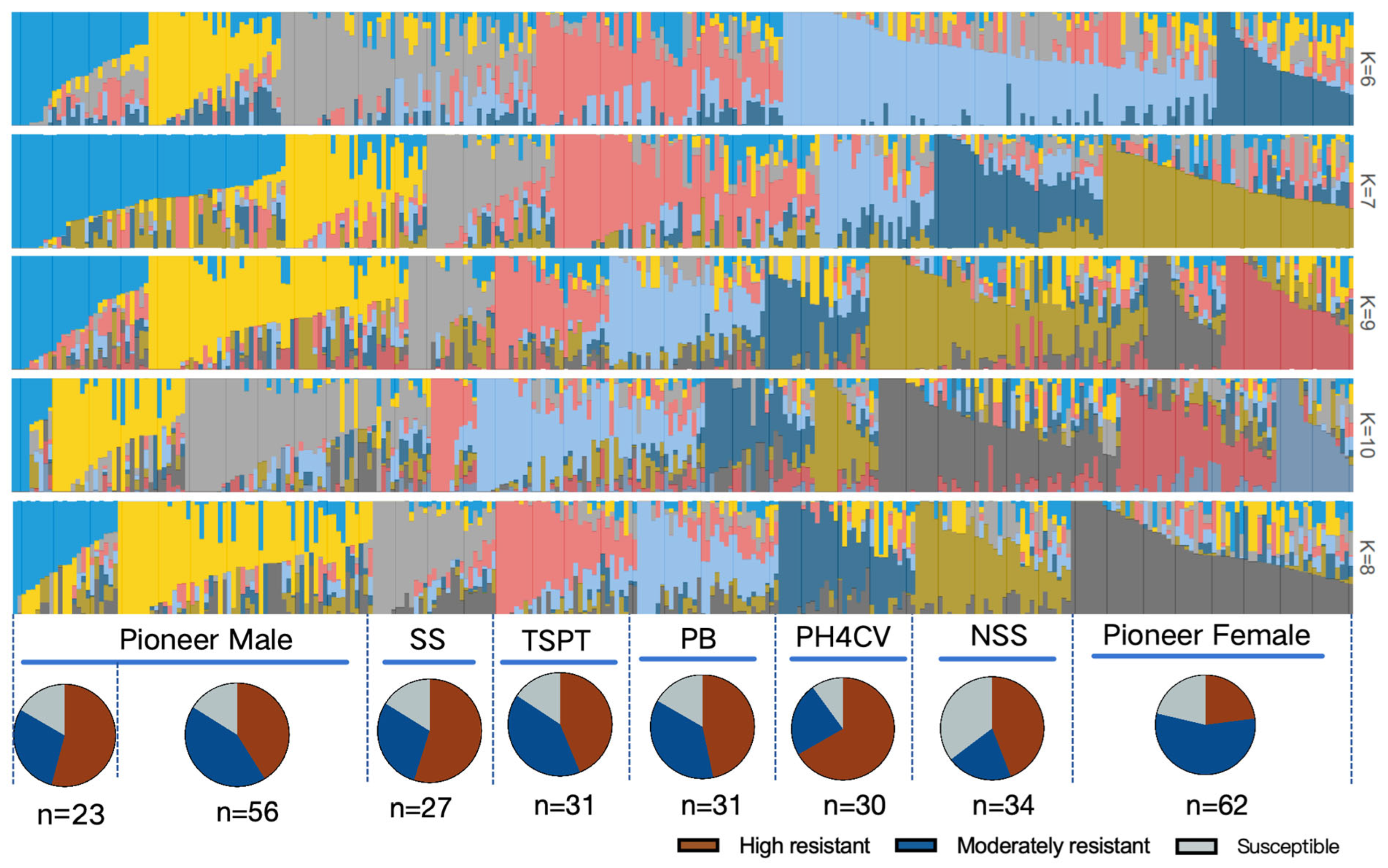
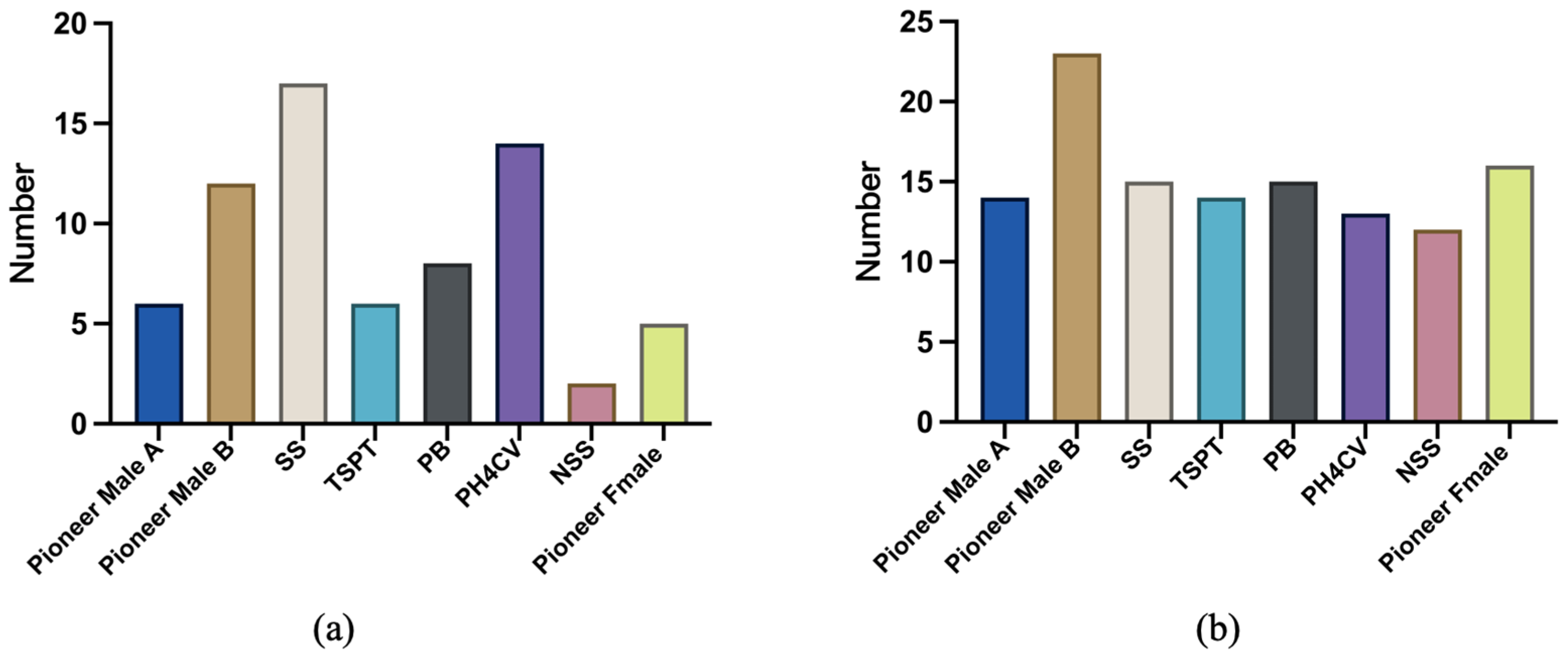
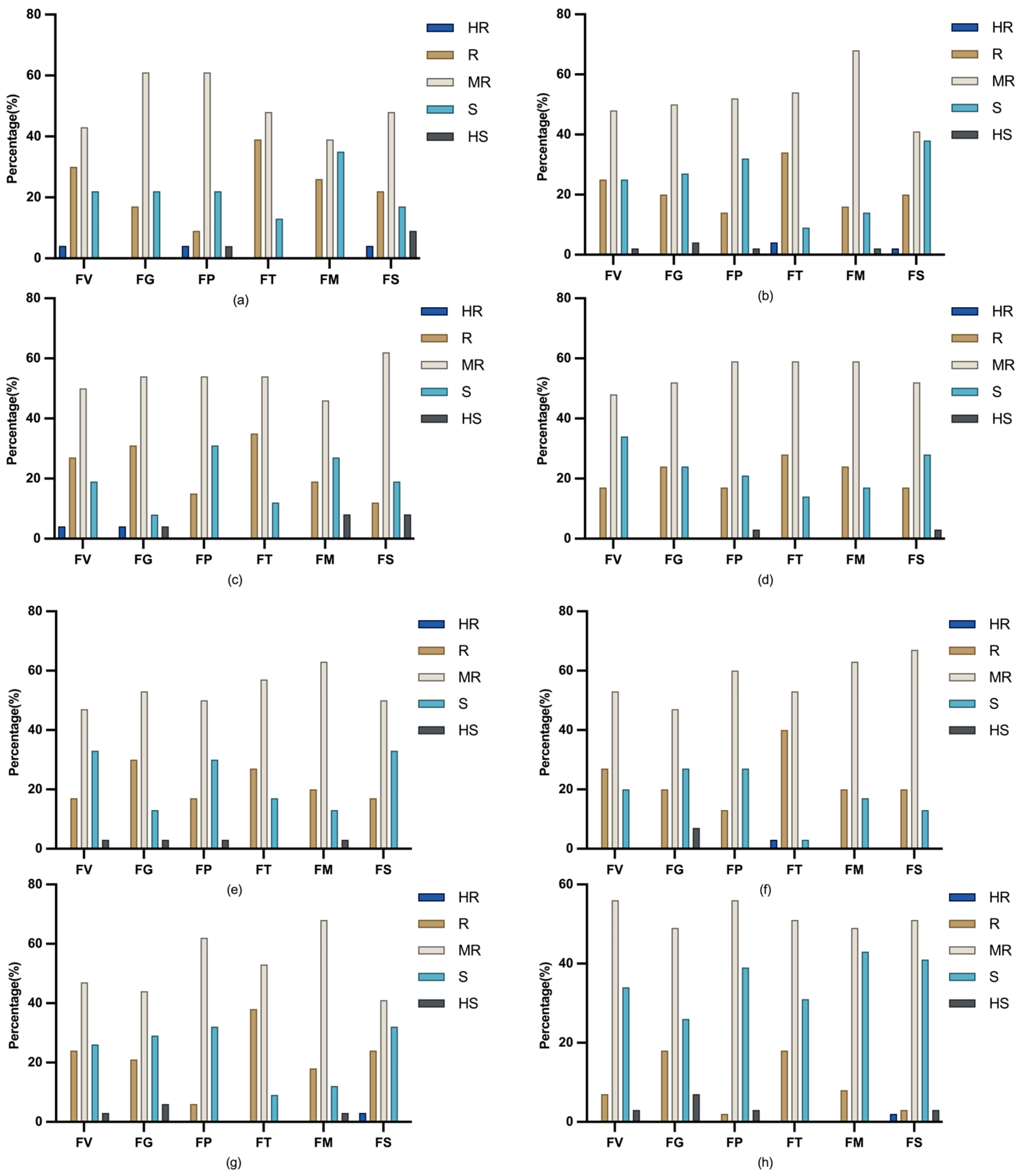
| Fusarium sp. | Geographic Distribution | Ecological Status |
|---|---|---|
| F. verticillioides | All primary maize agroecological zones | Dominant species |
| F. graminearum | All primary maize agroecological zones | Dominant species |
| F. proliferatum | Northeast, northwest, southwest | Regionally dominant species |
| F. meridionale | Southwest, southeast | Regionally dominant species |
| F. subglutinans | Northeast, northwest | Regionally dominant species |
| F. temperatum | Northeast | Regionally dominant species |
| F. oxysporum | Geographically undifferentiated distribution | Endemic species |
| F. culmorum | Geographically undifferentiated distribution | Endemic species |
| F. solani | Geographically undifferentiated distribution | Endemic species |
| F. semitectum | Geographically undifferentiated distribution | Endemic species |
| F. fujikuroi | Geographically undifferentiated distribution | Endemic species |
| F. verticillioides Ear Rot | F. graminearum Ear Rot | F. proliferatum Ear Rot | F. temperatum Ear Rot | F. meridionale Ear Rot | F. subglutinans Ear Rot | |
|---|---|---|---|---|---|---|
| 2022 | 5.76 | 5.28 | 5.86 | 5.47 | 5.79 | 6.07 |
| 2023 | 3.56 | 3.61 | 4.53 | 3.03 | 3.69 | 4.24 |
| 2024 | 6.55 | 6.59 | 6.26 | 6.10 | 6.89 | 6.36 |
| Fusarium Ear Rot | Genotypic Variance | F-Value | Broad Sense Heritability Value | p-Value |
|---|---|---|---|---|
| F. graminearum ear rot | 1.85 ± 0.21 | 24.37 | 0.86 | <0.001, *** |
| F. verticillioides ear rot | 1.62 ± 0.18 | 19.83 | 0.83 | <0.001, *** |
| F. proliferatum ear rot | 1.28 ± 0.15 | 15.02 | 0.54 | <0.001, *** |
| F. temperatum ear rot | 0.97 ± 0.11 | 10.45 | 0.32 | 0.003, ** |
| F. meridionale ear rot | 0.89 ± 0.10 | 9.12 | 0.31 | 0.008, ** |
| F. subglutinans ear rot | 0.75 ± 0.09 | 7.85 | 0.29 | 0.021, * |
| Fusarium Ear Rot | G × E Interaction Variance | Percentage of Total Variance |
|---|---|---|
| F. graminearum ear rot | 0.38 ± 0.05 | 17.2% |
| F. verticillioides ear rot | 0.42 ± 0.06 | 20.6% |
| F. proliferatum ear rot | 0.61 ± 0.08 | 32.3% |
| F. temperatum ear rot | 0.75 ± 0.09 | 43.6% |
| F. meridionale ear rot | 0.68 ± 0.08 | 43.3% |
| F. subglutinans ear rot | 0.59 ± 0.07 | 44.0% |
| Scale | Description | Ear Rot Score | Resistance |
|---|---|---|---|
| 1 | 0~1% of the diseased ear surface | ≤1.5 | Highly resistant (HR) |
| 3 | 2~10% of the diseased ear surface | 1.6~3.5 | Resistant (R) |
| 5 | 11~25% of the diseased ear surface | 3.6~5.5 | Moderately resistant (MR) |
| 7 | 26~50% of the diseased ear surface | 5.6~7.5 | Susceptible (S) |
| 9 | 51~100% of the diseased ear surface | 7.6~9.0 | Highly susceptible (HS) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Zhu, L.; Li, Y.; Guo, Y.; Zhang, Y.; Huang, C.; Wu, W.; Sun, S.; Cheng, Z.; Duan, C. Precise Identification and Analysis of Maize Germplasm Resistance to Ear Rot Caused by Six Fusarium Species. Plants 2025, 14, 2280. https://doi.org/10.3390/plants14152280
Li S, Zhu L, Li Y, Guo Y, Zhang Y, Huang C, Wu W, Sun S, Cheng Z, Duan C. Precise Identification and Analysis of Maize Germplasm Resistance to Ear Rot Caused by Six Fusarium Species. Plants. 2025; 14(15):2280. https://doi.org/10.3390/plants14152280
Chicago/Turabian StyleLi, Shuai, Lihong Zhu, Yongxiang Li, Yaxuan Guo, Yuhang Zhang, Chaosong Huang, Wenqi Wu, Suli Sun, Zixiang Cheng, and Canxing Duan. 2025. "Precise Identification and Analysis of Maize Germplasm Resistance to Ear Rot Caused by Six Fusarium Species" Plants 14, no. 15: 2280. https://doi.org/10.3390/plants14152280
APA StyleLi, S., Zhu, L., Li, Y., Guo, Y., Zhang, Y., Huang, C., Wu, W., Sun, S., Cheng, Z., & Duan, C. (2025). Precise Identification and Analysis of Maize Germplasm Resistance to Ear Rot Caused by Six Fusarium Species. Plants, 14(15), 2280. https://doi.org/10.3390/plants14152280