
Niche Overlap in Forest Tree Species Precludes a Positive Diversity–Productivity Relationship

 and
and
Abstract
1. Introduction
2. Results
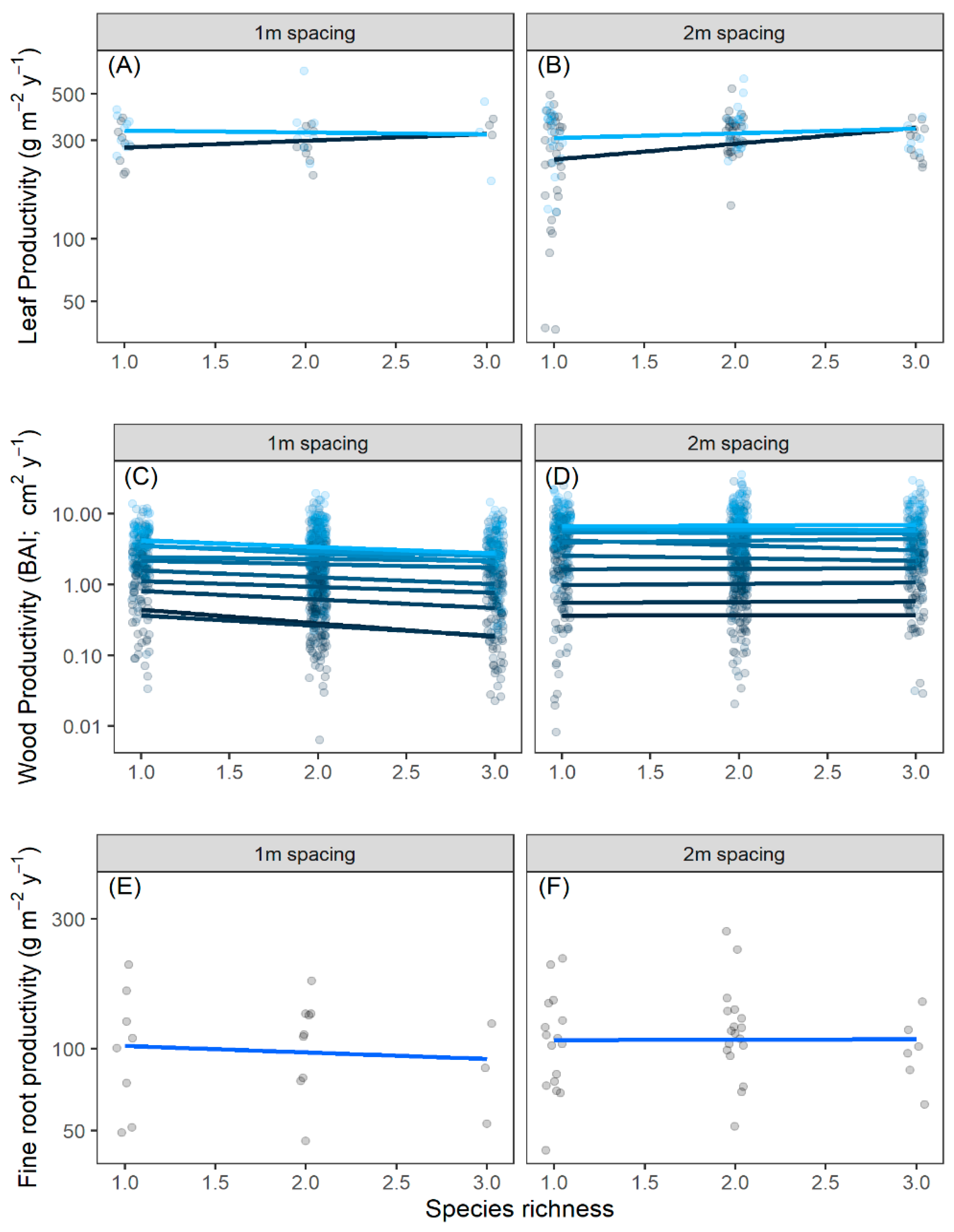
2.1. Diversity–Productivity Relationship
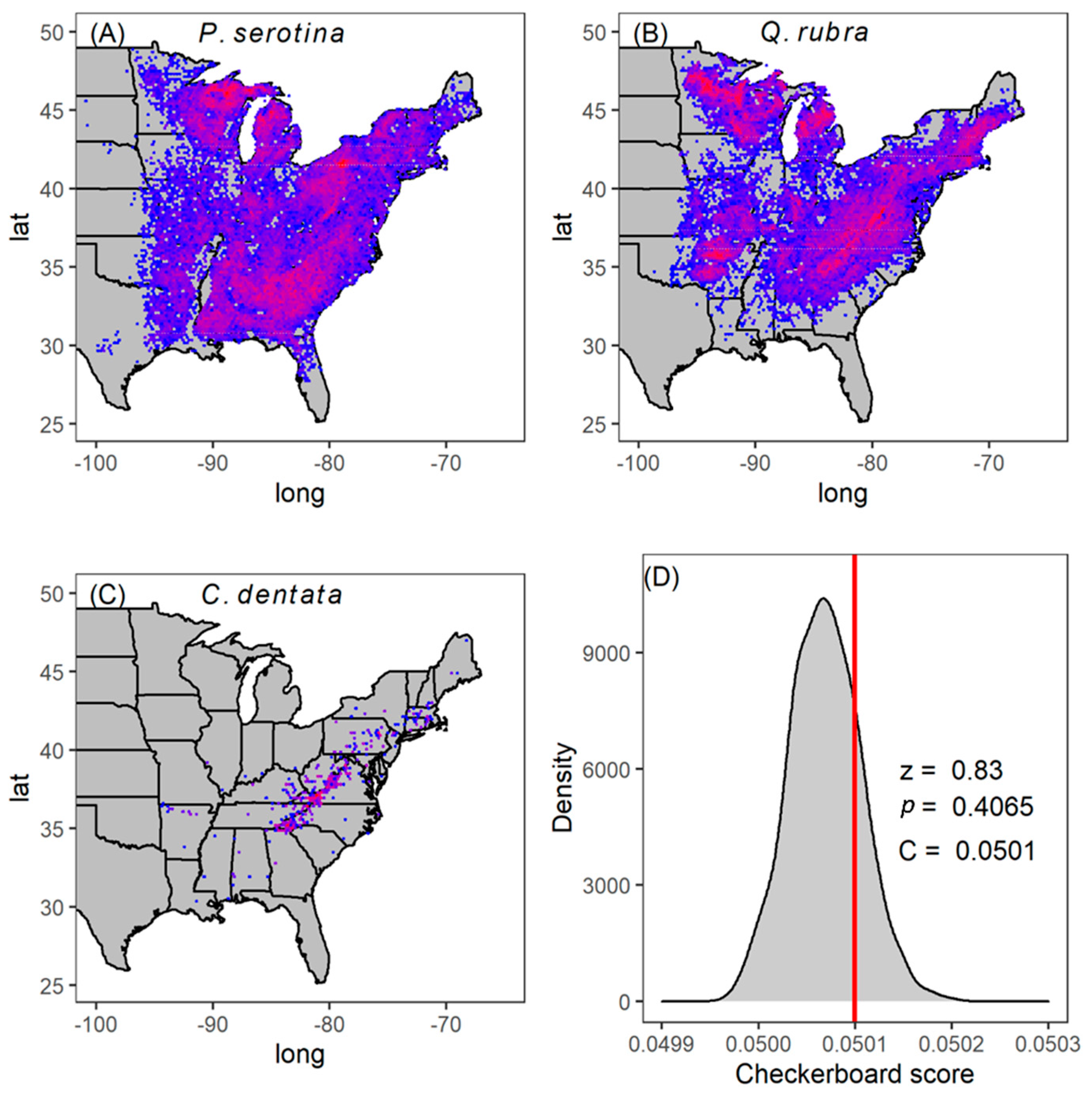
2.2. Patterns of Co-Occurrence
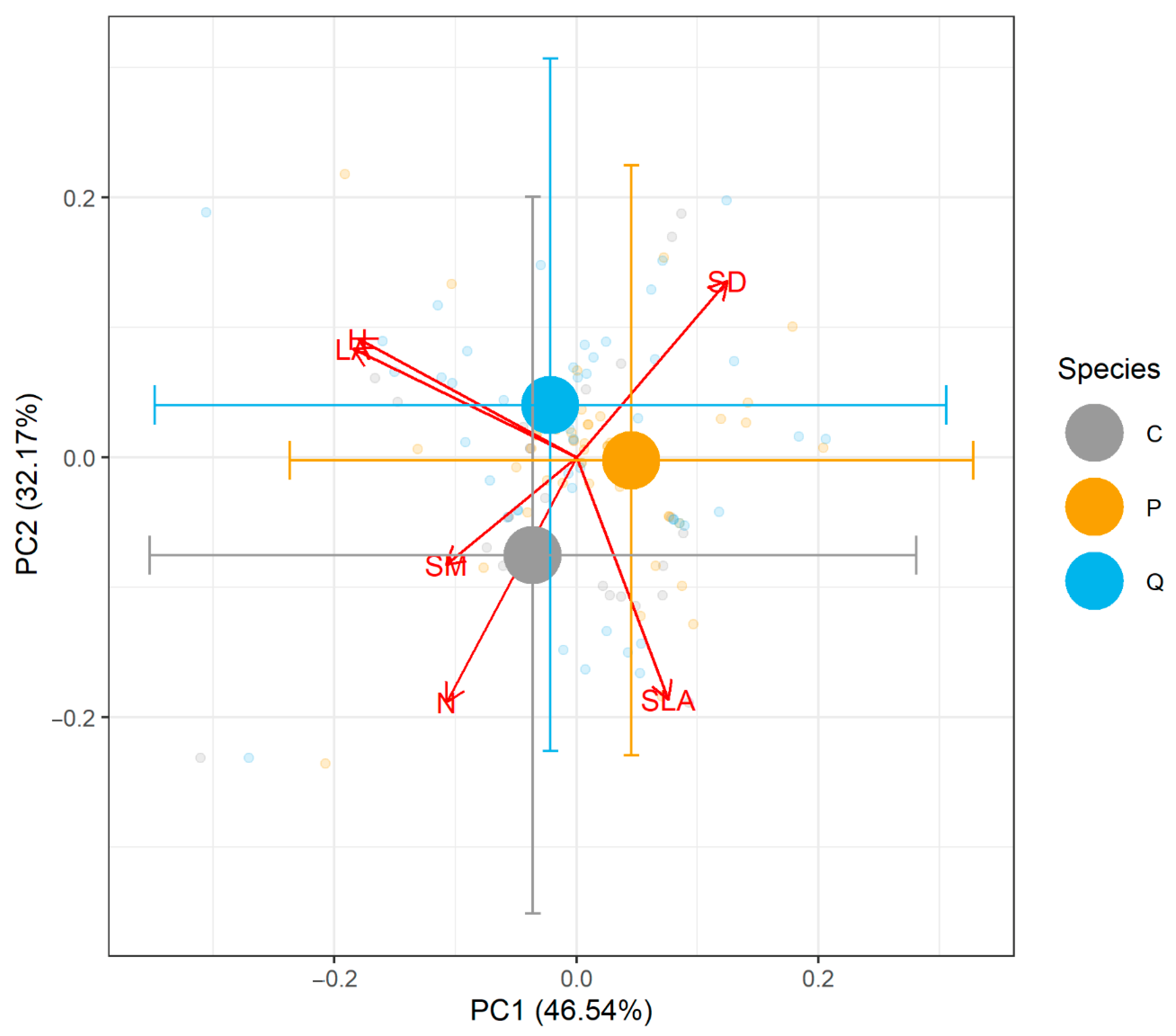
2.3. Functional Traits and Site Nutrient Availability
3. Discussion
Conclusions
4. Materials and Methods
4.1. Study Species and Experimental Plot Design
4.2. Productivity Estimates
4.3. USDA Forest Inventory and Analysis Data
4.4. Checkerboard Score Analysis
4.5. Functional Traits and Soil Nutrient and Pollutant Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Details of Planting Methods
Appendix A.2. Detailed Data Collection Methods
Appendix A.3. Measurement of Functional Traits in Our Plots
Appendix A.4. Detailed Description of Study Species
Appendix A.4.1. Quercus rubra (L.)
Appendix A.4.2. Prunus serotina (Ehrh.)
Appendix A.4.3. Castanea dentata ((Marsh.) Borkh.)
References
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Adler, P.B.; HilleRisLambers, J.; Levine, J.M. A niche for neutrality. Ecol. Lett. 2007, 10, 95–104. [Google Scholar] [CrossRef]
- Hart, S.P.; Freckleton, R.P.; Levine, J.M. How to quantify competitive ability. J. Ecol. 2018, 106, 1902–1909. [Google Scholar] [CrossRef]
- Wright, S.J. Plant diversity in tropical forests: A review of mechanisms of species coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Avolio, M.L.; Carroll, I.T.; Collins, S.L.; Houseman, G.R.; Hallett, L.M.; Isbell, F.; Koerner, S.E.; Komatsu, K.J.; Smith, M.D.; Wilcox, K.R. A comprehensive approach to analyzing community dynamics using rank abundance curves. Ecosphere 2019, 10, e02881. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Cazelles, K.; Gravel, D. Co-occurrence is not evidence of ecological interactions. Ecol. Lett. 2020, 23, 1050–1063. [Google Scholar] [CrossRef]
- Ulrich, W.; Gotelli, N.J. Pattern detection in null model analysis. Oikos 2013, 122, 2–18. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Godoy, O.; Levine, J.M. Plant functional traits and the multidimensional nature of species coexistence. Proc. Natl. Acad. Sci. USA 2015, 112, 797–802. [Google Scholar] [CrossRef]
- Liang, J.J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 12. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Homage to Santa-Rosalia or Why Are There So Many Kinds of Animals. Am. Nat. 1959, 93, 145–159. [Google Scholar] [CrossRef]
- HilleRisLambers, J.; Adler, P.B.; Harpole, W.S.; Levine, J.M.; Mayfield, M.M. Rethingking community assembly through the lens of coexistence theory. Annu. Rev. Ecol. Syst. 2012, 43, 227–248. [Google Scholar] [CrossRef]
- Grime, J.P. Control of species density in herbaceous vegetation. J. Environ. Manag. 1973, 1, 151–167. [Google Scholar]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef]
- Grace, J.B.; Anderson, T.M.; Seabloom, E.W.; Borer, E.T.; Adler, P.B.; Harpole, W.S.; Hautier, Y.; Hillebrand, H.; Lind, E.M.; Pärtel, M.; et al. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 2016, 529, 390–393. [Google Scholar] [CrossRef]
- Lotka, A.J. The growth of mixed populations: Two species competing for a common food supply. J. Wash. Acad. Sci. 1932, 22, 461–469. [Google Scholar]
- MacArthur, R.H.; Levins, R. Limiting Similarity Convergence and Divergence of Coexisting Species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Tilman, D. Resources—A graphical-mechanistic approach to competition and predation. Am. Nat. 1980, 116, 362–393. [Google Scholar] [CrossRef]
- van Ruijven, J.; Berendse, F. Diversity–productivity relationships: Initial effects, long-term patterns, and underlying mechanisms. Proc. Natl. Acad. Sci. USA 2005, 102, 695–700. [Google Scholar] [CrossRef]
- Diamond, J. Assembly of species communities. In Ecology and Evolution of Communities; Cody, M., Diamond, J., Eds.; Belknap Press: Cambridge, MA, USA, 1975; pp. 342–444. [Google Scholar]
- Stone, L.; Roberts, A. The checkerboard score and species distributions. Oecologia 1990, 85, 74–79. [Google Scholar] [CrossRef]
- Cheng, Y.X.; Zhang, C.Y.; Zhao, X.H.; von Gadow, K. Biomass-dominant species shape the productivity-diversity relationship in two temperate forests. Ann. For. Sci. 2018, 75, 9. [Google Scholar] [CrossRef]
- Head, M.L.; Holman, L.; Lanfear, R.; Kahn, A.T.; Jennions, M.D. The Extent and Consequences of P-Hacking in Science. PLoS Biol. 2015, 13, e1002106. [Google Scholar] [CrossRef]
- Bruns, S.B.; Ioannidis, J.P.A. p-Curve and p-Hacking in Observational Research. PLoS ONE 2016, 11, e0149144. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, D. The reproducibility of research and the misinterpretation of p-values. R. Soc. Open Sci. 2017, 4, 171085. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.M.; Zellers, K.E.; Lof, M.; Jacobs, D.F. Inter- and intra-specific competitiveness of plantation-grown American chestnut (Castanea dentata). For. Ecol. Manag. 2013, 291, 289–299. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Emily, W.B.R. Pre-Blight Distribution of Castanea dentata (Marsh.) Borkh. Bull. Torrey Bot. Club 1987, 114, 183–190. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Nelson, C.D.; Scrivani, J.A.; Jacobs, D.F. Consequences of Shifts in Abundance and Distribution of American Chestnut for Restoration of a Foundation Forest Tree. Forests 2016, 7, 4. [Google Scholar] [CrossRef]
- Hooper, D.U. The role of complementarity and competition in ecosystem reponses to variation in plant diversity. Ecology 1998, 79, 704–719. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Adler, P.B. Demography of perennial grassland plants: Survival, life expectancy and life span. J. Ecol. 2008, 96, 1023–1032. [Google Scholar] [CrossRef]
- Grainger, T.N.; Letten, A.D.; Gilbert, B.; Fukami, T. Applying modern coexistence theory to priority effects. Proc. Natl. Acad. Sci. USA 2019, 116, 6205–6210. [Google Scholar] [CrossRef] [PubMed]
- Narwani, A.; Alexandrou, M.A.; Oakley, T.H.; Carroll, I.T.; Cardinale, B.J. Experimental evidence that evolutionary relatedness does not affect the ecological mechanisms of coexistence in freshwater green algae. Ecol. Lett. 2013, 16, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Popper, K. Conjectures and Refutations: The Growth of Scientific Knowledge; Routledge and Kegan Paul: New York, NY, USA, 1963. [Google Scholar]
- Bates, D.M. Linear Mixed Model Implementation in lme4. 2007. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=e01c8f1cad9da9b8a7a38524d637933e7a2a921c (accessed on 6 July 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 26. [Google Scholar] [CrossRef]
- Gray, A.N.; Brandeis, T.J.; Shaw, J.D.; McWilliams, W.H.; Miles, P.D. Forest Inventory and Analysis Database of the United States of America (FIA). Biodivers. Ecol. 2012, 4, 225–231. [Google Scholar] [CrossRef]
- Ulrich, W.; Gotelli, N.J. Null model analysis of species associations using abundance data. Ecology 2010, 91, 3384–3397. [Google Scholar] [CrossRef]
- Gotelli, N.J. Null model analysis of species co-occurrence patterns. Ecology 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Hardy, O.J. Testing the spatial phylogenetic structure of local communities: Statistical performances of different null models and test statistics on a locally neutral community. J. Ecol. 2008, 96, 914–926. [Google Scholar] [CrossRef]
- Oksanen, J.F.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package v.2.7.1. 2025. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 6 July 2022).
- Miklós, I.; Podani, J. Randomization of presence–absence matrices: Comments and new algorithms. Ecology 2004, 85, 86–92. [Google Scholar] [CrossRef]
- Ulrich, W.; Gotelli, N.J. Disentangling community patterns of nestedness and species co-occurrence. Oikos 2007, 116, 2053–2061. [Google Scholar] [CrossRef]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Change Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef]
- Díaz, S.; Hodgson, J.G.; Thompson, K.; Cabido, M.; Cornelissen, J.H.C.; Jalili, A.; Montserrat-Martí, G.; Grime, J.P.; Zarrinkamar, F.; Asri, Y.; et al. The plant traits that drive ecosystems: Evidence from three continents. J. Veg. Sci. 2004, 15, 295–304. [Google Scholar] [CrossRef]
- Josse, J.; Husson, F. missMDA: A Package for Handling Missing Values in Multivariate Data Analysis. J. Stat. Softw. 2016, 70, 1–31. [Google Scholar] [CrossRef]
- Josse, J.; Husson, F. Handling missing values in exploratory multivariate data analysis methods. J. La Société Française Stat. 2012, 153, 79–99. [Google Scholar]
- Fox, J.; Weisberg, S. Package 'car'. 2010. Available online: http://cran.r-project.org/web/packages/car/car.pdf (accessed on 6 July 2022).
- Serbin, S.P. Spectroscopic Determination of Leaf Nutritional, Morpholgical, and Metabolic Traits. Ph.D. Dissertation, University of Wisconsin, Madison, WI, USA, 2012. [Google Scholar]
- Burns, R.M.; Honkala, B.H.; Service, F. Technical Coordinators. Silvics of Forest Trees of the United States. In Agriculture Handbook 654 (Supersedes Agriculture Handbook 271); U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1965. Available online: https://www.srs.fs.usda.gov/pubs/misc/ag_654/table_of_contents.htm (accessed on 6 July 2022).
- USDA.gov. Quercus rubra L. Available online: https://www.srs.fs.usda.gov/pubs/misc/ag_654/volume_2/quercus/rubra.htm (accessed on 6 July 2022).
- Gilman, E.F.; Watson, D.G.; Klein, R.W.; Koeser, A.K.; Hilbert, D.R.; Mclean, D.C. Prunus Serotina: Black Cherry 1. 2016. Available online: https://edis.ifas.ufl.edu/publication/ST516 (accessed on 6 July 2022).
- Nicolescu, V.N.; Vor, T.; Mason, W.L.; Bastien, J.C.; Brus, R.; Henin, J.M.; Kupka, I.; Lavnyy, V.; LaPorta, N.; Mohren, F.; et al. Ecology and management of northern red oak (Quercus rubra L. syn. Q. borealis F. Michx.) in Europe: A review. Forestry 2020, 93, 481–494. [Google Scholar]
- Arbor Day Foundation. Northern Red Oak. Available online: https://www.Arborday.Org/Trees/Treeguide/Treedetail.Cfm?ItemID=877 (accessed on 6 July 2022).
- Cecich, R.A. The Reproductive Biology of Quercus, with an Emphasis on Q. rubra. Int. Oak Soc. 1997, 7, 10–20. [Google Scholar]
- Peterson, S.J.; Neson, G.; Anderson, K. Plant Guide Northern Red Oak, Quercus rubra L. Plant Symbol = QURU. Plant Guide. 2019. Available online: http://cygnus.tamu.edu/Texlab/oakwilt.html (accessed on 6 July 2022).
- Terwei, A. Prunus serotina (Black Cherry); CABI Compendium; CABI: Wallingford, UK, 2022. [Google Scholar]
- Burns, R.M.; Honkala, B.H. Technical coordinators. In Silvics of North America: Vol. 2 Hardwoods; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Agriculture Handbook 654. Available online: https://research.fs.usda.gov/treesearch/1548 (accessed on 6 July 2022).
- Hough, A.F. Silvical Characteristics of Black Cherry (Prunus serotina); Station Paper NE-139. Upper Darby, PA; U.S. Department of Agriculture, Forest Service, Northeastern Research Station: Washington, DC, USA, 1960; pp. 1–26. Available online: https://research.fs.usda.gov/treesearch/13728 (accessed on 6 July 2022).
- Robakowski, P.; Bielinis, E.; Stachowiak, J.; Mejza, I.; Bułaj, B. Seasonal Changes Affect Root Prunasin Concentration in Prunus serotina and Override Species Interactions between P. serotina and Quercus petraea. J. Chem. Ecol. 2016, 42, 202–214. [Google Scholar] [CrossRef]
- Kinloch, B.B. Tree breeding practices. Breeding for Disease and Insect Resistance. In Encyclopedia of Forest Sciences; Academic Press: Cambridge, MA, USA, 2004; pp. 1472–1479. [Google Scholar]
- Jacobs, D.F.; Dalgleish, H.J.; Nelson, D. Synthesis of American Chestnut (Castanea dentata) Biological, Ecological, and Genetic Attributes with Application to Forest Restoration. For. Ecol. Manag. 2016, 7, 121041. [Google Scholar] [CrossRef]
- Clark, S.L.; Marcolin, E.; Patrício, M.S.; Loewe-Muñoz, V. A silvicultural synthesis of sweet (Castanea sativa) and American (C. dentata) chestnuts. Forest Ecol. Manag. 2023, 539, 121041. [Google Scholar] [CrossRef]
- Wang, G.G.; Knapp, B.O.; Clark, S.L.; Mudder, B.T. The Silvics of Castanea dentata (Marsh.) Borkh.)), American chestnut, Fagaceae (Beech Family); General Technical Report; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2013; 18p. [CrossRef]
- Saucier, J.R. American Chestnut (Castanea dentata (Marsh. Borkh.)); FS-230; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 1973.
- Buland, M.; Crocker, E. American Chestnut, Castanea dentata; University of Kentucky: Lexington, KY, USA, 2020; Available online: https://forestry.ca.uky.edu/files/forfs20-03american_chestnut.pdf (accessed on 6 July 2022).

{kind=link}
{kind=link}
{kind=link}
| Tissue | Treatment | F | df num | df den | p |
|---|---|---|---|---|---|
| Leaf | Diversity | 1.50 | 1 | 120.8 | 0.2231 |
| Density | 0.23 | 1 | 120.8 | 0.6325 | |
| Year | 2.75 | 1 | 83.2 | 0.1011 | |
| Diversity × Density | 0.17 | 1 | 120.8 | 0.6829 | |
| Diversity × Year | 0.53 | 1 | 83.2 | 0.4664 | |
| Density × Year | 0.05 | 1 | 83.2 | 0.8282 | |
| Diversity × Density × Year | 0.02 | 1 | 83.2 | 0.8870 | |
| Wood | Diversity | 2.59 | 1 | 197.3 | 0.1094 |
| Density | 0.45 | 1 | 197.7 | 0.5012 | |
| Year | 35.40 | 10 | 1733.0 | <0.0001 | |
| Diversity × Density | 2.41 | 1 | 197.3 | 0.1220 | |
| Diversity × Year | 0.41 | 10 | 1732.9 | 0.9434 | |
| Density × Year | 0.79 | 10 | 1733.0 | 0.6348 | |
| Diversity × Density × Year | 0.71 | 10 | 1733.0 | 0.7136 | |
| Root | Diversity | 0.17 | 1 | 56 | 0.6787 |
| Density | <0.01 | 1 | 56 | 0.9745 | |
| Diversity × Density | 0.13 | 1 | 56 | 0.7193 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blackstone, K.M.S.; McNickle, G.G.; Ritzi, M.V.; Nelson, T.M.; Hardiman, B.S.; Montague, M.S.; Jacobs, D.F.; Couture, J.J. Niche Overlap in Forest Tree Species Precludes a Positive Diversity–Productivity Relationship. Plants 2025, 14, 2271. https://doi.org/10.3390/plants14152271
Blackstone KMS, McNickle GG, Ritzi MV, Nelson TM, Hardiman BS, Montague MS, Jacobs DF, Couture JJ. Niche Overlap in Forest Tree Species Precludes a Positive Diversity–Productivity Relationship. Plants. 2025; 14(15):2271. https://doi.org/10.3390/plants14152271
Chicago/Turabian StyleBlackstone, Kliffi M. S., Gordon G. McNickle, Morgan V. Ritzi, Taylor M. Nelson, Brady S. Hardiman, Madeline S. Montague, Douglass F. Jacobs, and John J. Couture. 2025. "Niche Overlap in Forest Tree Species Precludes a Positive Diversity–Productivity Relationship" Plants 14, no. 15: 2271. https://doi.org/10.3390/plants14152271
APA StyleBlackstone, K. M. S., McNickle, G. G., Ritzi, M. V., Nelson, T. M., Hardiman, B. S., Montague, M. S., Jacobs, D. F., & Couture, J. J. (2025). Niche Overlap in Forest Tree Species Precludes a Positive Diversity–Productivity Relationship. Plants, 14(15), 2271. https://doi.org/10.3390/plants14152271