
Synergistic Interaction Between Endophytic Bacillus pumilus and Indigenous Arbuscular Mycorrhizal Fungi Complex Improves Photosynthetic Activity, Growth, and Yield of Pisum sativum


 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mycorrhizal Inoculum Production
2.2. AM Fungal Identification and Diversity
2.3. Bacterial Inoculum Production
2.4. Bacterial Identification
2.5. Seed Treatment
2.6. Experimental Design
2.7. Morphological Features and Biomass Production
2.8. Photosynthetic Activity and Yield
2.9. Mycorrhizal Colonization Rate of Pea Roots
2.10. Statistical Analysis
3. Results
3.1. Composition and Diversity of the AMF Complex Associated with Zea mays in the Argania Spinosa Rhizosphere
3.2. Mycorrhizal Frequency and Intensity in Pea Seedlings Following Various Treatments
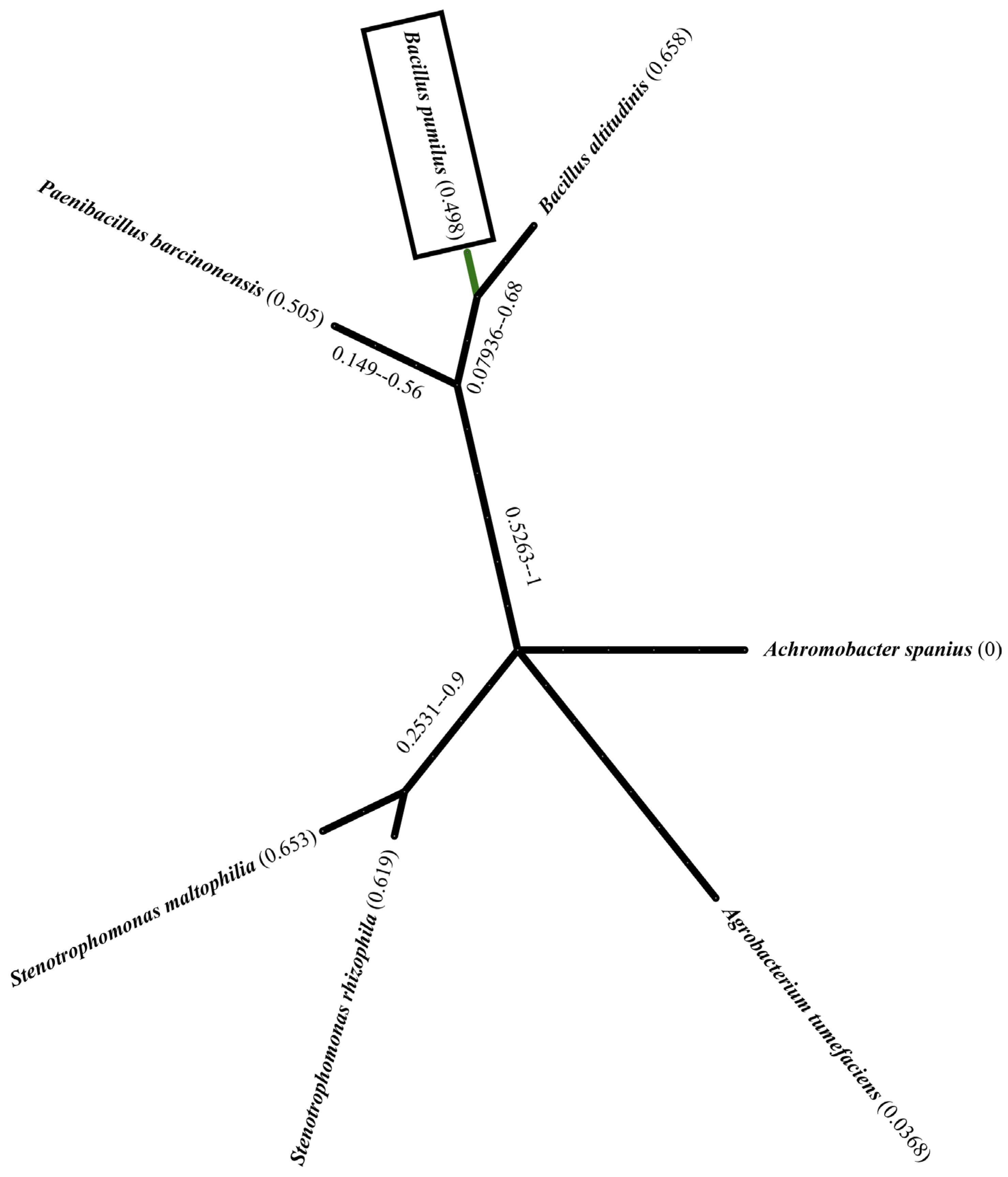
3.3. Morphological and Biochemical Characterization of the Bacillus Pumilus Strain
3.4. Demonstration of the Stimulation of the Growth Metrics of Peas
3.5. Plant Height
3.6. Collar Diameter
3.7. Leaf Production
3.8. Flower and Pod Production
3.9. Biomass Production of Peas
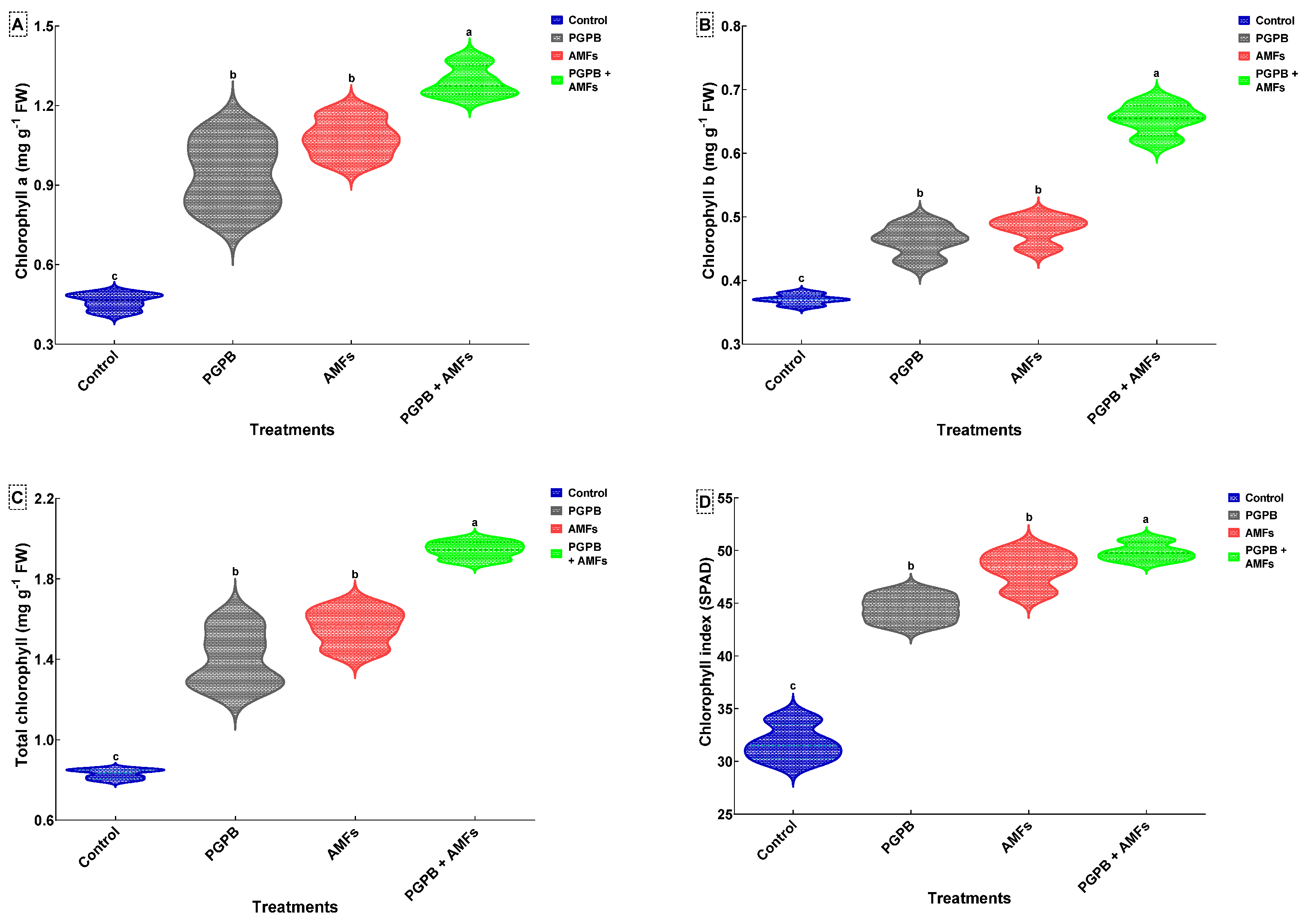
3.10. Chlorophyll Contents of Peas
3.11. Photosynthetic Yield
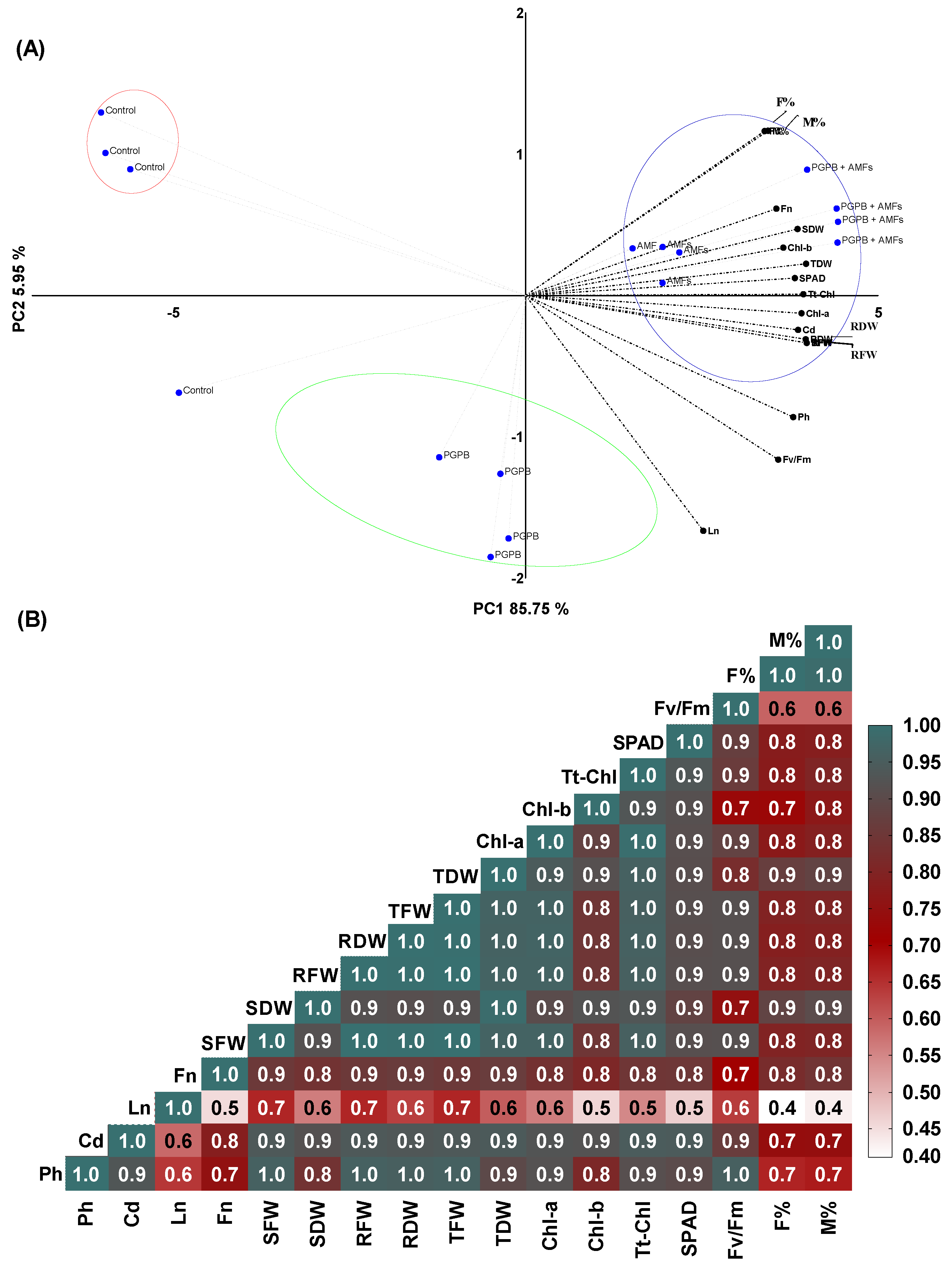
3.12. Multivariate Analysis of Trait Interactions in Pisum Sativum Following the Microbial Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. The Future of Food and Agriculture—Alternative Pathways to 2050, Summary Version; FAO: Rome, Italy, 2017. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Daoui, K. Development of Food and Fodder Legumes in Morocco: Retrospective and Prospective Study; FAO Report: Rome, Italy, 2021. [Google Scholar]
- Krimi Bencheqroun, S.; Kumari, S.G.; Moukahel, A.; Ahmed, S. Major Viruses Affecting Food Legumes in Morocco; (Internal Report); International Center for Agricultural Research in the Dry Areas (ICARDA): Beirut, Lebanon, 2023; Available online: https://hdl.handle.net/10568/138172 (accessed on 15 April 2025).
- Kharrat, M.; Ben Salah, H.; Maatougui, M.H. Study on the Food and Fodder Legume Sector in North Africa; Food & Agriculture Org.—FAO: Rome, Italy, 2023; p. 72. [Google Scholar] [CrossRef]
- Duan, H.X.; Luo, C.L.; Wang, X.; Cheng, Y.S.; Abrar, M.; Batool, A. Responses of legumes to rhizobia and arbuscular mycorrhizal fungi under abiotic stresses: A global meta-analysis. Agronomy 2024, 14, 2597. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Y.; Jia, B.; Tao, L.; Li, H.; Wang, J.; Yao, Y. Four decades of Bacillus biofertilizers: Advances and future prospects in agriculture. Microorganisms 2025, 13, 187. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Xiang, D.; Li, X.; Gao, Q.; Chen, Y.; Wang, K.; Xiang, H. Effects of combined inoculation of arbuscular mycorrhizal fungi and plant growth-promoting rhizosphere bacteria on seedling growth and rhizosphere microecology. Front. Microbiol. 2025, 15, 1475485. [Google Scholar] [CrossRef]
- Ilahi, H.; Calvo, A.; Dhane, S.; El Idrissi, M.M.; Ouahmane, L.; Alfeddy, M.N.; Mnasri, B. Impact of bacterial inoculations on Pisum sativum L. biometric parameters and associated bacterial and AM fungal communities under semi-arid field conditions in Tunisia. Appl. Soil Ecol. 2025, 205, 105757. [Google Scholar] [CrossRef]
- Masood, S.; Zhao, X.Q.; Shen, R.F. Bacillus pumilus promotes the growth and nitrogen absorption of tomato plants under nitrogen fertilization. Sci. Hortic. 2020, 272, 109581. [Google Scholar] [CrossRef]
- Gutiérrez-Mañero, F.J.; Ramos-Solano, B.; Probanza, A.N.; Mehouachi, J.; Tadeo, F.R.; Talon, M. Plant growth-promoting rhizobacteria, Bacillus pumilus, and Bacillus licheniformis produce large quantities of physiologically active gibberellins. Physiol. Plant. 2001, 111, 206–211. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Hernandez, J.P.; Bashan, Y.; Maier, R.M. Bacillus pumilus ES4: Candidate plant growth-promoting bacteria to improve plant establishment in mine tailings. Approx. Exp. Bot. 2010, 69, 343–352. [Google Scholar] [CrossRef]
- Kaushal, M.; Kumar, A.; Kaushal, R. Bacillus pumilus strain YSPMK11 as a plant growth promoter and biocontrol agent against Sclerotinia sclerotiorum. 3 Biotech 2017, 7, 90–99. [Google Scholar] [CrossRef]
- Shaffique, S.; Imran, M.; Injamum-Ul-Hoque, M.; Zainurin, N.; Peter, O.; Alomrani, S.O.; Lee, I.J. Unraveling the new member Bacillus pumilus SH-9 of Bacillaceae family and its potential role in seed biopriming to mitigate drought stress in Oryza sativa. Plant Stress 2024, 11, 100318. [Google Scholar] [CrossRef]
- Shaffique, S.; Imran, M.; Rahim, W.; Alomrani, S.O.; Yun, B.W.; Lee, I.J. A newly isolated Bacillus pumilus strain SH-9 modulates response to drought stress in soybean via endogenous phytohormones and gene expression (Daegu, South Korea). Plant Stress 2023, 10, 100279. [Google Scholar] [CrossRef]
- Bouskout, M.; Bourhia, M.; Al Feddy, M.N.; Dounas, H.; Salamatullah, A.M.; Soufan, W.; Ouahmane, L. Mycorrhizal fungi inoculation improves Capparis spinosa’s yield, nutrient uptake and photosynthetic efficiency under water deficit. Agronomy 2022, 12, 149. [Google Scholar] [CrossRef]
- Bouskout, M.; Dounas, H.; Alfeddy, M.N.; Ouahmane, L. Autochthonous Arbuscular Mycorrhizal Fungi Enhance Growth, Nutritional Homeostasis, and Antioxidant Machinery of Caper-Bush (Capparis spinosa L.) Plants Under Salt Stress. J. Soil Sci. Plant Nutr. 2025, 25, 728–749. [Google Scholar] [CrossRef]
- Bouskout, M.; ElJebri, S.; Khan, Y.; Saleh, I.A.; Okla, M.K.; Al-Amri, S.S.; Ouahmane, L. Unveiling native mycorrhizal fungi diversity: Insights into growth performance, nutrient uptake, and root system robustness in caper-bush (Capparis spinosa L.) seedlings. J. Agric. Food Res. 2025, 21, 101984. [Google Scholar] [CrossRef]
- Barwant, M.M.; Singh, B.; Sharma, S.; Gore, N.T.; Mohammad, A. Role of Mycorrhizal Association in Leguminous Plant Growth. In Recent Trends and Applications of Leguminous Microgreens as Functional Foods; Springer Nature Switzerland: Cham, Switzerland, 2025; pp. 277–295. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Njeri, K.V.; Shen, Q.; Zhang, N.; Zhang, R. Tolerance induced by corn salt by inoculation into the rhizosphere of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Fasusi, O.A.; Babalola, O.O.; Adejumo, T.O. Harnessing of plant growth-promoting rhizobacteria and arbuscular mycorrhizal fungi in agroecosystem sustainability. CABI Agric. Biosci. 2023, 4, 26. [Google Scholar] [CrossRef]
- Bortolot, M.; Buffoni, B.; Mazzarino, S.; Hoff, G.; Martino, E.; Fiorilli, V.; Salvioli Di Fossalunga, A. The Importance of Mycorrhizal Fungi and Their Associated Bacteria in Promoting Crops’ Performance: An Applicative Perspective. Horticulturae 2024, 10, 1326. [Google Scholar] [CrossRef]
- Ouahmane, L.; Ndoye, I.; Morino, A.; Ferradous, A.; Sfairi, Y.; Al Faddy, M.N.; Abourouh, M. Inoculation of Ceratonia siliqua L. with native arbuscular mycorrhizal fungi mixture improves seedling establishment under greenhouse conditions. Afr. J. Biotechnol. 2012, 11, 16422–16426. [Google Scholar]
- Lee, J.; Lee, S.; Young, J.P.W. Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef]
- Simon, L.; Lalonde, M.; Bruns, T. Specific amplification of 18S fungal ribosomal genes from vesicular-arbuscular endomycorrhizal fungi colonizing roots. Appl. Environ. Microbiol. 1992, 58, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Thioye, B.; Legras, M.; Castel, L.; Hirissou, F.; Chaftar, N.; Trinsoutrot-Gattin, I. Understanding arbuscular mycorrhizal colonization in walnut plantations: The contribution of cover crops and soil microbial communities. Agriculture 2022, 12, 1. [Google Scholar] [CrossRef]
- Slimani, A.; Harkousse, O.; Mazri, M.A.; Zouahri, A.; Ouahmane, L.; Koussa, T.; Al Feddy, M.N. Impact of a selected mycorrhizal complex and a rhizobacterial species on tomato plants’ growth under water stress conditions. Indian J. Agric. Res. 2022, 56, 696–704. [Google Scholar] [CrossRef]
- Willems, A.; Coopman, R.; Gillis, M. Comparison of sequence analysis of 16S-23S rDNA spacer regions, AFLP analysis and DNA-DNA hybridizations in Bradyrhizobium. Int. J. Syst. Evol. Microbiol. 2001, 51, 623–632. [Google Scholar] [CrossRef]
- Ponsonnet, C.; Nesme, X. Identification of Agrobacterium strains by PCR-RFLP analysis of pTi and chromosomal regions. Arch. Microbiol. 1994, 161, 300–309. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae, Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA Press: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Lipková, N.; Cinkocki, R.; Maková, J.; Medo, J.; Javoreková, S. Characterization of endophytic bacteria of the Bacillus genus and their influence on the growth of corn (Zea mays) in vivo. J. Microbiol. Biotechnol. Food Sci. 2021, 10, e3602. [Google Scholar] [CrossRef]
- Gupta, R.; Singal, R.; Shankar, A.; Kuhad, R.C.; Saxena, R.L. A modified plate assay for screening phosphate solubilizing microorganisms. J. Gen. Appl. Microbiol. 1994, 40, 255–260. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Xu, W.; Zeng, J.; Li, L.; Li, S.; Gao, Z. Bacillus pumilus LZP02 promotes rice root growth by improving carbohydrate metabolism and phenylpropanoid biosynthesis. Mol. Plant Microbe Interact. 2020, 33, 1222–1231. [Google Scholar] [CrossRef] [PubMed]
- Pudova, D.S.; Toymentseva, A.A.; Gogoleva, N.E.; Shagimardanova, E.I.; Mardanova, A.M.; Sharipova, M.R. Comparative analysis of the genome of two strains of Bacillus pumilus producing a high level of extracellular hydrolases. Genes 2022, 13, 409. [Google Scholar] [CrossRef]
- Yakovleva, G.; Kurdy, W.; Gorbunova, A.; Khilyas, I.; Lochnit, G.; Ilinskaya, O. The Bacillus pumilus proteome changes in response to 2,4,6-trinitrotoluene-induced stress. Biodegradation 2022, 33, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lai, Q.; Dong, C.; Sun, F.; Wang, L.; Li, G.; Shao, Z. Phylogenetic diversity of the Bacillus pumilus group and marine ecotype revealed by multilocus sequence analysis. PLoS ONE 2013, 8, e80097. [Google Scholar] [CrossRef]
- Cuong, P.V.; Hoa, N.P. Optimization of culture conditions for the production of IAA by Bacillus sp. isolated from a cassava field in Vietnam. Vietnam J. Sci. Technol. 2021, 59, 312–323. [Google Scholar] [CrossRef]
- Bessai, S.A.; Bensidhoum, L.; Nabti, E.H. Optimization of IAA production by soil bacteria isolated from northern Algeria. Biocatal. Agric. Biotechnol. 2022, 41, 102319. [Google Scholar] [CrossRef]
- Madigan, M.; Martinko, J. Biology of Microorganisms, 11th ed.; Pearson Education: Chennai, India, 2007; p. 655. [Google Scholar]
- Figueiredo, V.B.; Burity, H.A.; Martinez, C.R.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paeni Bacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]
- Devi, K.K.; Seth, N.; Kothamasi, S.; Kothamasi, D. Hydrogen cyanide-producing rhizobacteria kill the subterranean termite Odontotermes obesus (Rambur) by cyanide poisoning under in vitro conditions. Curr. Microbiol. 2007, 54, 74–78. [Google Scholar] [CrossRef]
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol. Agron. Soc. Environ. 2011, 15, 327–337. [Google Scholar]
- Shahzad, A.; Qin, M.; Elahie, M.; Naeem, M.; Bashir, T.; Yasmin, H.; Younas, M.; Areeb, A.; Irfan, M.; Billah, M.; et al. Bacillus pumilus induced tolerance of maize (Zea mays L.) to cadmium (Cd) stress. Sci. Rep. 2021, 11, 17196. [Google Scholar] [CrossRef]
- Armada, E.; Azcón, R.; López-Castillo, O.M.; Calvo-Polanco, M.; Ruiz-Lozano, J.M. Autochthonous arbuscular mycorrhizal fungi and Bacillus thuringiensis from a degraded Mediterranean area can be used to improve physiological traits and performance of a plant of agronomic interest under drought conditions. Plant Physiol. Biochem. 2015, 90, 64–74. [Google Scholar] [CrossRef]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef]
- Sánchez-Navarro, V.; Zornoza, R.; Faz, Á.; Egea-Gilabert, C.; Ros, M.; Pascual, J.A.; Fernández, J.A. Inoculation with different nitrogen-fixing bacteria and arbuscular mycorrhiza affects grain protein content and nodule bacterial communities of a fava bean crop. Agronomy 2020, 10, 768. [Google Scholar] [CrossRef]
- Liu, X.Q.; Xie, M.M.; Hashem, A.; Abd-Allah, E.F.; Wu, Q.S. Arbuscular mycorrhizal fungi and rhizobia synergistically promote root colonization, plant growth, and nitrogen acquisition. Plant Growth Regul. 2023, 100, 691–701. [Google Scholar] [CrossRef]
- Abkenar, M.B.; Mozafari, H.; Karimzadeh, K.; Rajabzadeh, F.; Azimi, R. Arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) as an alternative to mineral fertilizers to improve the growth, essential oil profile, and phenolic content of Satureja macrantha L. J. Crop Health 2024, 76, 347–356. [Google Scholar] [CrossRef]
- Ain, Q.U.; Hussain, H.A.; Zhang, Q.; Maqbool, F.; Ahmad, M.; Mateen, A.; Imran, A. Coordinated influence of Funneliformis mosseae and different plant growth-promoting bacteria on growth, root functional traits, and nutrient acquisition by maize. Mycorrhiza 2024, 34, 477–488. [Google Scholar] [CrossRef]
- Gebreslassie, S.; Jida, M.; Puente, M.L.; Covacevich, F.; Belay, Z. Inoculation of native arbuscular mycorrhizae and Bacillus subtilis can improve growth in vegetable crops. Int. J. Microbiol. 2024, 2024, 9226715. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Jakubowska, Z.; Dybek, B. Potential of Bacillus pumilus to directly promote plant growth. Front. Microbiol. 2022, 13, 1069053. [Google Scholar] [CrossRef]
- Chakraborty, U.; Chakraborty, B.N.; Roychowdhury, P. Plant growth-promoting activity of Bacillus pumilus in tea (Camellia sinensis) and its biocontrol potential against Poria hypobrunnea. Indian Phytopathol. 2013, 66, 387–396. Available online: https://epubs.icar.org.in/index.php/IPPJ/article/view/36017 (accessed on 24 June 2025).
- Hafeez, F.Y.; Yasmin, S.; Ariani, D.; Renseigné, N.; Zafar, Y.; Malik, K.A. Bacteria promoting plant growth as a biofertilizer. Agron. Sustain. Dev. 2006, 26, 143–150. [Google Scholar] [CrossRef]
- Ahmad, F.; Iqbal, S.; Anwar, S.; Afzal, M.; Islam, E.; Mustafa, T.; Khan, Q.M. Improving soil chlorpyrifos remediation using ryegrass (Lollium multiflorum) and the chlorpyrifos-degrading bacteria Bacillus pumilus C2A1. J. Hazard. Mater. 2012, 237–238, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Induction of rhizosphere microbiome by application of biofertilizers to suppress Fusarium wilt disease of banana. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Costa-Santos, M.; Mariz-Ponte, N.; Dias, M.C.; Moura, L.; Marques, G.; Santos, C. Effect of Bacillus spp. and Brevibacillus sp. on the photosynthesis and redox status of Solanum lycopersicum. Horticulturae 2021, 7, 24. [Google Scholar] [CrossRef]
- Awasthi, A.; Bharti, N.; Nair, P.; Singh, R.; Shukla, A.K.; Gupta, M.M.; Darokar, M.P.; Kalra, A. Synergistic effect of Glomus mosseae and nitrogen-fixing Bacillus subtilis strain Daz26 on artemisinin content in Artemisia annua L. Appl. Soil Ecol. 2011, 49, 125–130. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improves seed yield and quality of soybean plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef]
- Khalid, M.; Hassani, D.; Bilal, M.; Asad, F.; Huang, D. Influence of bio-fertilizer containing beneficial fungi and rhizospheric bacteria on health-promoting compounds and antioxidant activity of Spinacia oleracea L. Bot. Stud. 2017, 58, 1–9. [Google Scholar] [CrossRef]
- Moreira, H.; Pereira, S.I.; Vega, A.; Castro, P.M.; Marques, A.P. Synergistic effects of arbuscular mycorrhizal fungi and plant growth-promoting bacteria benefit maize growth under increasing soil salinity. J. Environ. Manag. 2020, 257, 109982. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A. Synergistic effects of the inoculation with plant growth-promoting rhizobacteria and an arbuscular mycorrhizal fungus on the performance of wheat. Turk. J. Agric. For. 2007, 31, 355–362. Available online: https://journals.tubitak.gov.tr/agriculture/vol31/iss6/1 (accessed on 24 June 2025).
- Wilkes, T.I.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G. Species-specific interactions of Bacillus innocula and arbuscular mycorrhizal fungi symbiosis with winter wheat. Microorganisms 2020, 8, 1795. [Google Scholar] [CrossRef]
- El-Azeem, A.; Abd El-Malik, S.; Bucking, H. Effectiveness of combined inoculation of rhizobacteria and arbuscular mycorrhizal fungi on wheat performance. Egypt. J. Soil Sci. 2023, 63, 35–45. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; El-Enany, A.W.E.; Nafady, N.A.; Khalaf, D.M.; Morsy, F.M. Synergistic interaction of Rhizobium leguminosarum bv. viciae and arbuscular mycorrhizal fungi as plant growth-promoting biofertilizers for faba bean (Vicia faba L.) in alkaline soil. Microbiol. Res. 2014, 169, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef]
- Da Costa Neto, V.P.; de Melo, A.R.P.; Alencar, C.E.S.; de Lima, V.B.C.; Zilli, J.E.; Rodrigues, A.C.; Bonifacio, A. Bacterial consortia among Bradyrhizobium species, Azospirillum baldaniorum, and Bacillus pumilus promote plant growth and efficient symbiotic nitrogen fixation in mung bean. Symbiosis 2024, 93, 255–267. [Google Scholar] [CrossRef]
- Alam, M.; Khaliq, A.; Sattar, A.; Shukla, R.S.; Anwar, M.; Dharni, S. Synergistic effect of arbuscular mycorrhizal fungi and Bacillus subtilis on the biomass and essential oil yield of rose-scented geranium (Pelargonium graveolens). Arch. Agron. Soil Sci. 2011, 57, 889–898. [Google Scholar] [CrossRef]
- Ngosong, C.; Tatah, B.N.; Olougou, M.N.E.; Suh, C.; Nkongho, R.N.; Ngone, M.A.; Achiri, D.T.; Tchakounté, G.V.T.; Ruppel, S. Inoculating plant growth-promoting bacteria and arbuscular mycorrhizal fungi modulates rhizosphere acid phosphatase and nodulation activities and enhances the productivity of soybean (Glycine max). Front. Plant Sci. 2022, 13, 934339. [Google Scholar] [CrossRef]
- Savastano, N.; Bais, H. Synergism or antagonism: Do arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria work together to benefit plants? Int. J. Plant Biol. 2024, 15, 944–958. [Google Scholar] [CrossRef]
- Nader, A.A.; Hauka, F.I.; Afify, A.H.; El-Sawah, A.M. Drought-tolerant bacteria and arbuscular mycorrhizal fungi mitigate the detrimental effects of drought stress induced by withholding irrigation at critical growth stages of soybean (Glycine max L.). Microorganisms 2024, 12, 1123. [Google Scholar] [CrossRef]
- Shaffique, S.; Shah, A.A.; Odongkara, P.; Elansary, H.O.; Khan, A.L.; Adhikari, A.; Kang, S.M.; Lee, I.J. Deciphering the ABA and GA biosynthesis approach of Bacillus pumilus, a mechanistic approach, explaining the role of the metabolic region as an aid in improving stress tolerance. Sci. Rep. 2024, 14, 28923. [Google Scholar] [CrossRef]
- Yadav, R.; Ror, P.; Rathore, P.; Kumar, S.; Ramakrishna, W. Bacillus subtilis CP4, isolated from native soil in combination with arbuscular mycorrhizal fungi promotes biofortification, yield, and metabolite production in wheat under field conditions. J. Appl. Microbiol. 2021, 131, 339–359. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | PGPB | AMFs | PGPB + AMFs | |
|---|---|---|---|---|
| F (%) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 100 ± 0.0 a | 100 ± 0.0 a |
| M (%) | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 36.25 ± 2.50 a | 41.25 ± 4.79 a |
| Macroscopic Characteristics | ||||||
| Odor | Form | Color | Margin | Elevation | Surface | Size (mm) |
| Odorless | Round | Cream | Whole | Raised | Mucoid | 6.5 |
| Microscopic Characteristics | ||||||
| Gram reaction | Form | Motility | Sporulation | |||
| Positive (+) | Bacillus | Mobile | Positive (+) | |||
| Biochemical Characteristic | Bacillus pumilus |
|---|---|
| Breathing type | Optional Aerobics |
| Csein | - |
| NaCl | - |
| Hydrolysis of starch | + |
| Growth at 50 °C | - |
| Growth at 60 °C | - |
| Nitrate reduction | + |
| Mannose utilization | + |
| Arabinose utilization | - |
| Fructose utilization | + |
| Maltose utilization | + |
| Citrate utilization | - |
| Arginine degradation | + |
| Gelatinase activity | + |
| Catalase activity | + |
| Strains | AIA Production | PO4 Solubilization | NH3 Production | Hydrogen Cyanide Production | Pectinase Activity | Cellulase Activity | N Fixation Activity |
|---|---|---|---|---|---|---|---|
| B. pumilus | + | - | + | + | - | + | + |
| Parameters | DF | F | ANOVA |
|---|---|---|---|
| Plant height (cm) | 3 | 53.93 | <0.0001 |
| Collar diameter (mm) | 3 | 43.47 | <0.0001 |
| Leaves number | 3 | 6.487 | 0.0074 |
| Flower and pod numbers | 3 | 17.26 | 0.0001 |
| Shoot fresh weight | 3 | 661.2 | <0.0001 |
| Soot dry weight | 3 | 107.9 | <0.0001 |
| Root fresh weight | 3 | 144.1 | <0.0001 |
| Root dry weight | 3 | 133.2 | <0.0001 |
| Total fresh weight | 3 | 445.5 | <0.0001 |
| Total dry weight | 3 | 99.46 | <0.0001 |
| Chlorophyll a | 3 | 68.89 | <0.0001 |
| Chlorophyll b | 3 | 124.6 | <0.0001 |
| Total chlorophyll (Chl a + b) | 3 | 100.9 | <0.0001 |
| Chlorophyll index (SPAD) | 3 | 131.1 | <0.0001 |
| Photosynthetic efficiency (Fv/Fm) | 3 | 67.34 | <0.0001 |
| F (%) | 3 | 273.8 | <0.0001 |
| M (%) | 3 | 276.9 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhallaa Youne, M.; Akhallaa Youne, O.; Bouskout, M.; Khan, Y.; Khassali, H.; Shah, S.; Sujat, A.; Alahoui, H.; Alfeddy, M.N.; Mnasri, B.; et al. Synergistic Interaction Between Endophytic Bacillus pumilus and Indigenous Arbuscular Mycorrhizal Fungi Complex Improves Photosynthetic Activity, Growth, and Yield of Pisum sativum. Plants 2025, 14, 1991. https://doi.org/10.3390/plants14131991
Akhallaa Youne M, Akhallaa Youne O, Bouskout M, Khan Y, Khassali H, Shah S, Sujat A, Alahoui H, Alfeddy MN, Mnasri B, et al. Synergistic Interaction Between Endophytic Bacillus pumilus and Indigenous Arbuscular Mycorrhizal Fungi Complex Improves Photosynthetic Activity, Growth, and Yield of Pisum sativum. Plants. 2025; 14(13):1991. https://doi.org/10.3390/plants14131991
Chicago/Turabian StyleAkhallaa Youne, Mounia, Oumnia Akhallaa Youne, Mohammed Bouskout, Yaseen Khan, Hamza Khassali, Sulaiman Shah, Ahmed Sujat, Hassan Alahoui, Mohamed Najib Alfeddy, Bacem Mnasri, and et al. 2025. "Synergistic Interaction Between Endophytic Bacillus pumilus and Indigenous Arbuscular Mycorrhizal Fungi Complex Improves Photosynthetic Activity, Growth, and Yield of Pisum sativum" Plants 14, no. 13: 1991. https://doi.org/10.3390/plants14131991
APA StyleAkhallaa Youne, M., Akhallaa Youne, O., Bouskout, M., Khan, Y., Khassali, H., Shah, S., Sujat, A., Alahoui, H., Alfeddy, M. N., Mnasri, B., & Ouahmane, L. (2025). Synergistic Interaction Between Endophytic Bacillus pumilus and Indigenous Arbuscular Mycorrhizal Fungi Complex Improves Photosynthetic Activity, Growth, and Yield of Pisum sativum. Plants, 14(13), 1991. https://doi.org/10.3390/plants14131991