
PP2C-Mediated ABA Signaling Pathway Underlies Exogenous Abscisic Acid-Induced Enhancement of Saline–Alkaline Tolerance in Potato (Solanum tuberosum L.)

 and
and
Abstract
1. Introduction
2. Results
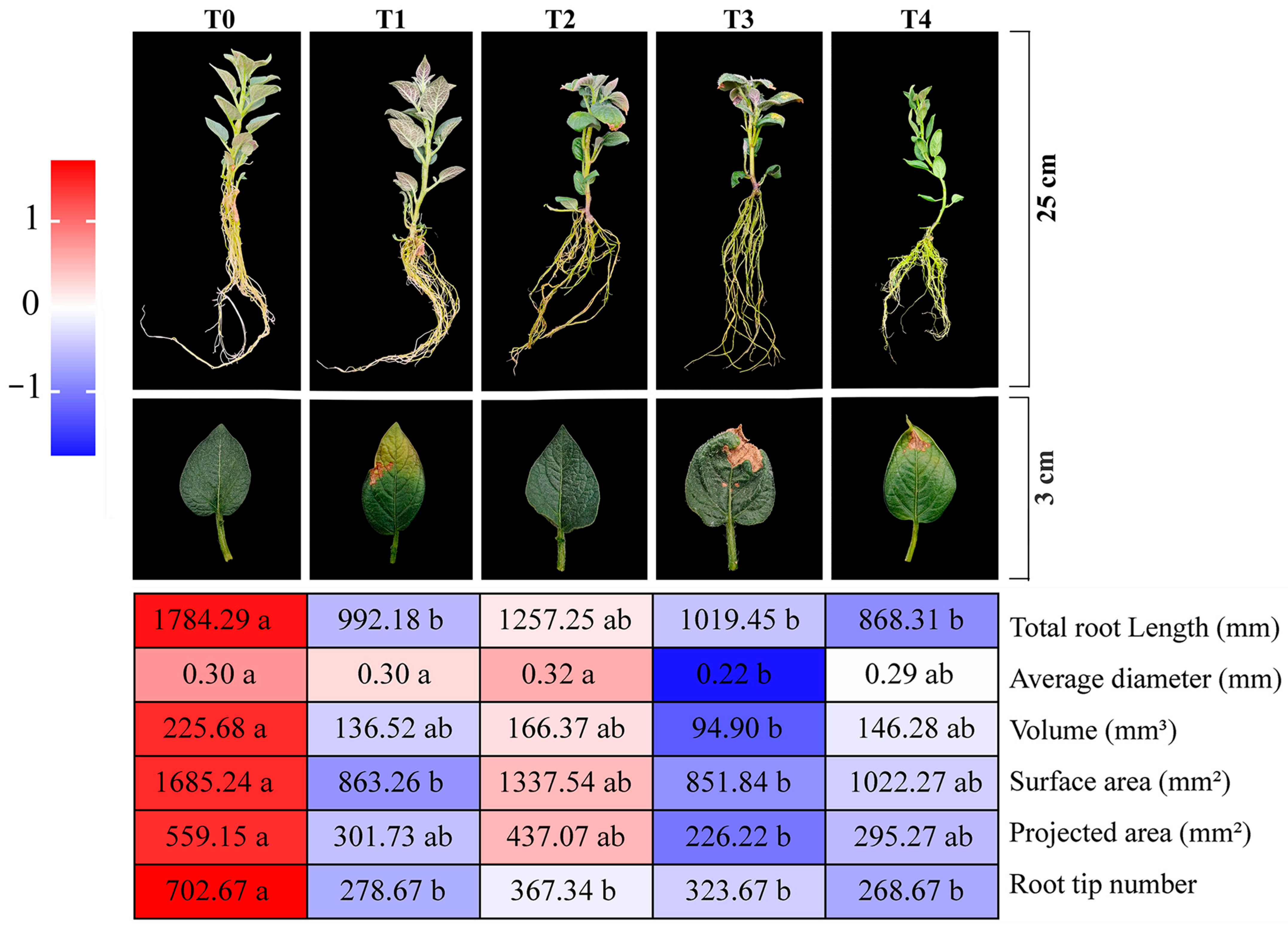
2.1. Effects of Exogenous ABA Treatment on Growth and Development of Potato Plants Under Saline–Alkali Stress
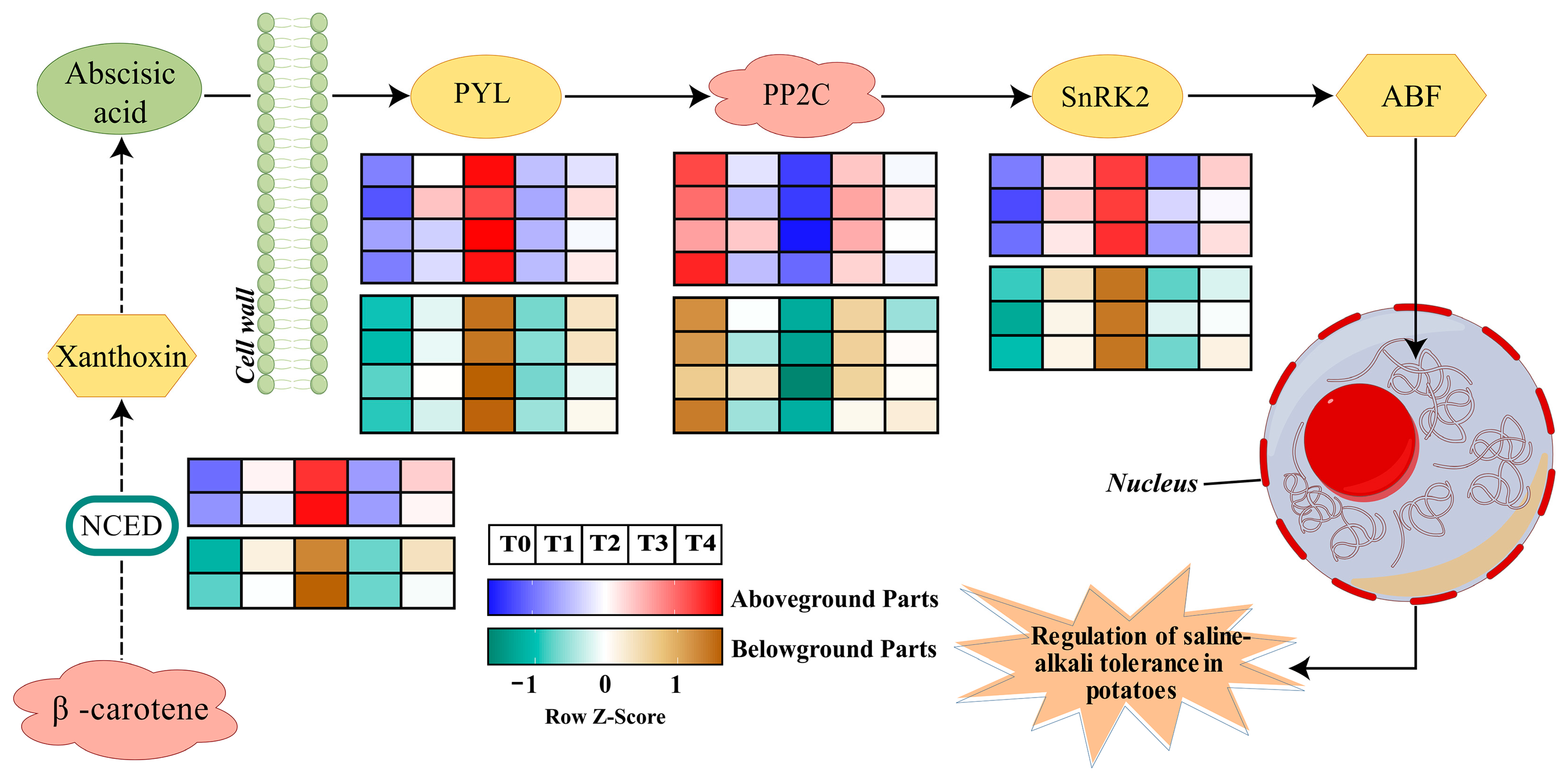
2.2. The Key Pathway of ABA in Potato Response to Saline–Alkali Stress
2.3. Identification of Transgenic Potatoes
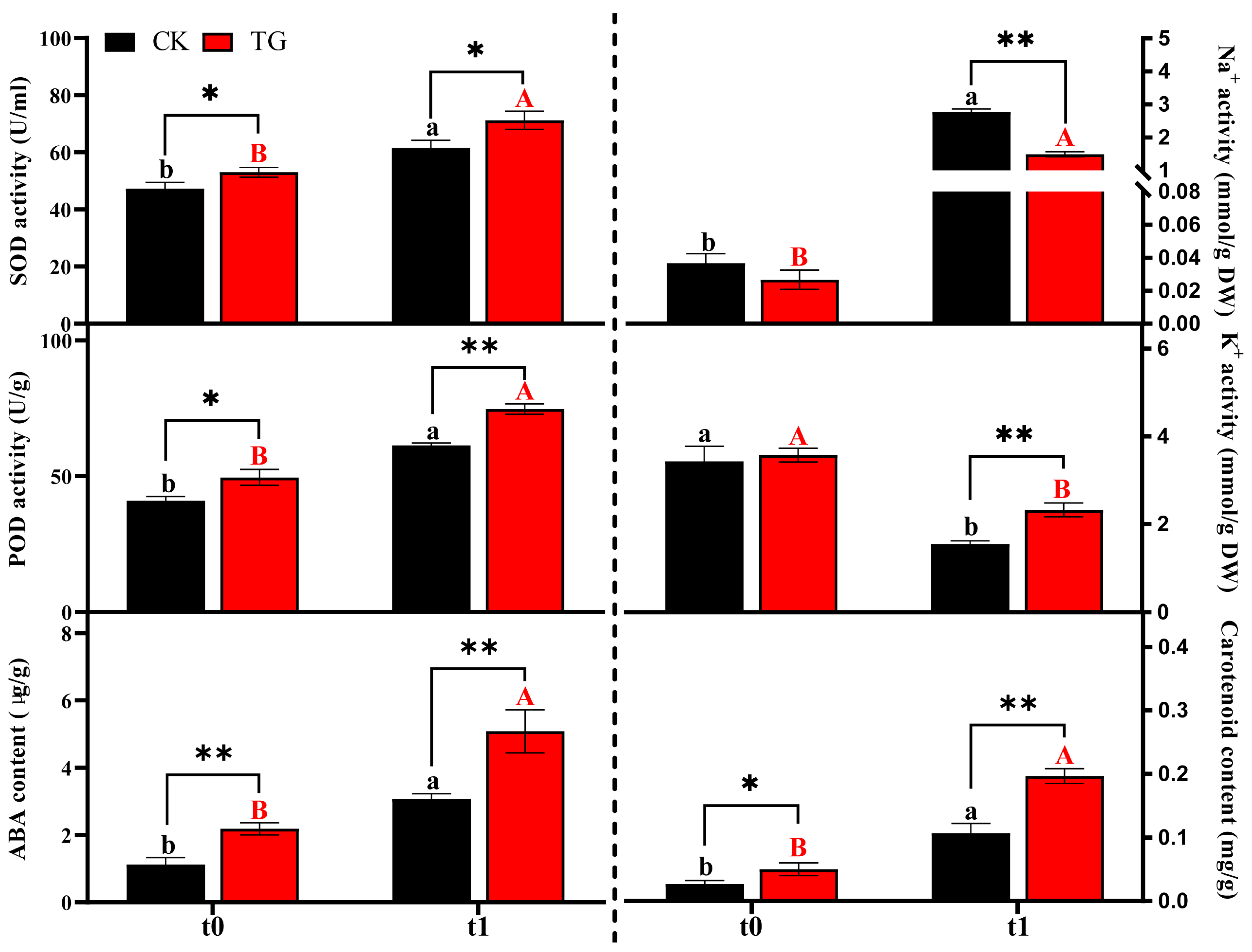
2.4. Changes in Physiological Characteristics of Transgenic Potatoes Under Saline–Alkali Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Experimental Design
4.2.1. Experimental Design of ABA for Alleviating Alkaline Salt Stress
4.2.2. Identification of Resistance of Transgenic Plants
4.3. Determination of Morphological Indicators
4.4. qRT-PCR Analysis of Key Genes
4.5. Generation of Transgenic Plants
4.6. Measurement of Physiological Index
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| PP2C | Protein phosphortase 2C |
| PT | PGSC0003DMT40 |
| SnRK2 | Sucrose non-fermenting-1-related protein kinase 2 |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| cDNA | Complementary DNA |
| HPLC | High-performance liquid chromatography |
| NCED | 9-cis epoxy carotenoid dioxygenase |
| PYL | Pyrabatin resistance 1-like proteins |
| ABF | Abscisic acid-responsive element binding factors |
| MVA | Methyl erythritol phosphate |
| XAN | Xanthoxin |
| PYR | Pyrabatin resistance |
| ABRE | Abscisic acid responsive element |
References
- Zhang, H.; Yu, F.; Xie, P.; Sun, S.; Qiao, X.; Tang, S.; Chen, C.; Yang, S.; Mei, C.; Yang, D.; et al. A Gγ protein regulates alkaline sensitivity in crops. Science 2023, 379, eade8416. [Google Scholar] [CrossRef] [PubMed]
- F.A.O. Global Status of Salt-Affected Soils. In International Soil and Water Forum 2024; F.A.O.: Bangkok, Thailand, 2024.
- Xu, W.; Jia, L.; Shi, W.; Balu¡ka, F.e.; Kronzucker, H.J.; Liang, J.; Zhang, J. The Tomato 14-3-3 Protein TFT4 Modulates H+ Efflux, Basipetal Auxin Transport, and the PKS5-J3 Pathway in the Root Growth Response to Alkaline Stress+. Plant Physiol. 2013, 163, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Termaat, A.; Munns, R.; Termaat, A. Whole-Plant Responses to Salinity. Funct. Plant Biol. 1986, 13, 143–160. [Google Scholar] [CrossRef]
- Wang, C.; Gao, C.; Wang, L.; Zheng, L.; Yang, C.; Wang, Y. Comprehensive transcriptional profiling of NaHCO3-stressed Tamarix hispida roots reveals networks of responsive genes. Plant Mol. Biol. 2014, 84, 145–157. [Google Scholar] [CrossRef]
- Ge, Y.; Li, Y.; Zhu, Y.-M.; Bai, X.; Lv, D.-K.; Guo, D.; Ji, W.; Cai, H. Global transcriptome profiling of wild soybean (Glycine soja) roots under NaHCO3 treatment. BMC Plant Biol. 2010, 10, 153. [Google Scholar] [CrossRef]
- Kumari, R.; Sharma, V.K. Seed Culture of Rice Cultivars under Salt Stress. Kome 2015, 3, 191–202. [Google Scholar]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef]
- Hong, J.H.; Seah, S.W.; Xu, J. The root of ABA action in environmental stress response. Plant Cell Rep. 2013, 32, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Z.; Gao, J.; Wang, P.; Hu, T.; Wang, Z.; Hou, Y.-J.; Wan, Y.; Liu, W.; Xie, S.; et al. Arabidopsis Duodecuple Mutant of PYL ABA Receptors Reveals PYL Repression of ABA-Independent SnRK2 Activity. Cell Rep. 2018, 23, 3340–3351. [Google Scholar] [CrossRef]
- Dar, N.A.; Amin, I.; Wani, W.; Wani, S.A.; Shikari, A.B.; Wani, S.H.; Masoodi, K.Z. Abscisic acid: A key regulator of abiotic stress tolerance in plants. Plant Gene 2017, 11, 106–111. [Google Scholar] [CrossRef]
- Lu, C.; Chen, M.X.; Liu, R.; Zhang, L.; Hou, X.; Liu, S.; Ding, X.; Jiang, Y.; Xu, J.; Zhang, J.; et al. Abscisic Acid Regulates Auxin Distribution to Mediate Maize Lateral Root Development Under Salt Stress. Front. Plant Sci. 2019, 10, 716. [Google Scholar] [CrossRef]
- Vighi, I.L.; Crizel, R.L.; Perin, E.C.; Rombaldi, C.V.; Galli, V. Crosstalk During Fruit Ripening and Stress Response Among Abscisic Acid, Calcium-Dependent Protein Kinase and Phenylpropanoid. Crit. Rev. Plant Sci. 2019, 38, 99–116. [Google Scholar] [CrossRef]
- Fu, J.; Wu, Y.; Miao, Y.; Xu, Y.; Zhao, E.; Wang, J.; Sun, H.; Liu, Q.; Xue, Y.; Xu, Y.; et al. Improved cold tolerance in Elymus nutans by exogenous application of melatonin may involve ABA-dependent and ABA-independent pathways. Sci. Rep. 2017, 7, 39865. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Wang, Y.; Zhang, S.; Zhang, J. ABA signaling in stress-response crosstalk. Curr. Opin. Plant Biol. 2016, 33, 25–32. [Google Scholar]
- Zhang, X.; Zhu, W.; Liu, Z.; Ren, X.; Li, Y.; Li, G.; Wang, J.; Zhu, X.; Shi, Y.; Wang, C.; et al. Transcriptomic analysis of melatonin-mediated drought stress response genes in alfalfa during germination period. BMC Plant Biol. 2025, 25, 637. [Google Scholar] [CrossRef]
- Bai, Y.; Wei, Y.; Yin, H.; Hu, W.; Cheng, X.; Guo, J.; Dong, Y.; Zhang, L.; Xie, H.; Zeng, H.; et al. PP2C1 fine-tunes melatonin biosynthesis and phytomelatonin receptor PMTR1 binding to melatonin in cassava. J. Pineal Res. 2022, 73, e12804. [Google Scholar] [CrossRef]
- Carlos, B.H.; Vicent, A.; Aurelio, G. Jasmonic acid transient accumulation is needed for abscisic acid increase in citrus roots under drought stress conditions. Physiol. Plantarum. 2012, 147, 296–306. [Google Scholar]
- Rachmawati, D.; Ramadhani, A.N.; Fatikhasari, Z. The effect of silicate fertilizer on the root development of rice and its tolerance to salinity stress. Carpath. J. Earth Environ. 2021, 724, 012004. [Google Scholar] [CrossRef]
- Hu, C.; Zheng, Y.; Tong, C.; Zhang, D. Effects of exogenous melatonin on plant growth, root hormones and photosynthetic characteristics of trifoliate orange subjected to salt stress. Plant Growth Regul. 2022, 97, 551–558. [Google Scholar] [CrossRef]
- Parveen, A.; Ahmar, S.; Kamran, M.; Malik, Z.; Ali, A.; Riaz, M.; Abbasi, G.H.; Khan, M.; Sohail, A.B.; Rizwan, M.; et al. Abscisic acid signaling reduced transpiration flow, regulated Na+ ion homeostasis and antioxidant enzyme activities to induce salinity tolerance in wheat (Triticum aestivum L.) seedlings. Environ. Technol. Innov. 2021, 24, 101808. [Google Scholar] [CrossRef]
- Bartley, G.E.; Scolnik, P.A.; Giuliano, G. Molecular Biology of Carotenoid Biosynthesis in Plants. Annu. Rev. Plant Biol. 1994, 45, 287–301. [Google Scholar] [CrossRef]
- Milborrow, B.V. The pathway of biosynthesis of abscisic acid in vascular plants: A review of the present state of knowledge of ABA biosynthesis. J. Exp. Bot. 2001, 52, 1145–1164. [Google Scholar] [CrossRef] [PubMed]
- Swapnil, P.; Meena, M.; Singh, S.K.; Dhuldhaj, U.P.; Harish; Marwal, A. Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Curr. Plant Biol. 2021, 26, 100203. [Google Scholar] [CrossRef]
- Johnson, M.P.; Havaux, M.; Triantaphylides, C.; Ksas, B.; Pascal, A.A.; Robert, B.; Davison, P.A.; Ruban, A.V.; Horton, P. Elevated zeaxanthin bound to oligomeric LHCII enhances the resistance of arabidopsis to photooxidative stress by a lipid-protective, antioxidant mechanism. J. Biol. Chem. 2007, 282, 22605–22618. [Google Scholar] [CrossRef]
- Kang, C.; Zhai, H.; Xue, L.; Zhao, N.; He, S.; Liu, Q. A lycopene β-cyclase gene, IbLCYB2, enhances carotenoid contents and abiotic stress tolerance in transgenic sweetpotato. Plant Sci. 2018, 272, 243–254. [Google Scholar] [CrossRef]
- Juan, C.; Moreno, J.M.; Alagoz, Y.; Al-Babili, S. Plant apocarotenoids: From retrograde signaling to interspecific communication. Plant J. 2020, 105, 351–375. [Google Scholar]
- Kim, Y.; Hwang, I.; Jung, H.J.; Park, J.I.; Kang, J.G.; Nou, I.S. Genome-Wide Classification and Abiotic Stress-Responsive Expression Profiling of Carotenoid Oxygenase Genes in Brassica rapa and Brassica oleracea. J. Plant Growth Regul. 2016, 35, 202–214. [Google Scholar] [CrossRef]
- Assmann, S.M.; Jegla, T. Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid, and CO2. Curr. Opin. Plant Biol. 2016, 33, 157–167. [Google Scholar] [CrossRef]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F.; et al. Abscisic acid inhibits Type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C Phosphatase Activity Function as Abscisic Acid Sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Yu, Y.; Zhu, L.; Liu, H.; Chai, Q.; Guo, W. A cotton NAC transcription factor GhirNAC2 plays positive roles in drought tolerance via regulating ABA biosynthesis. Plant Sci. 2020, 296, 110498. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Peng, D.; Liu, M.; Wei, A.; Li, X. TaFDL2-1A interacts with TabZIP8-7A protein to cope with drought stress via the abscisic acid signaling pathway. Plant Sci. 2021, 311, 111022. [Google Scholar] [CrossRef]
- Chen, J.; Gong, Y.; Gao, Y.; Zhou, Y.; Chen, M.; Xu, Z.; Guo, C.; Ma, Y. TaNAC48 positively regulates drought tolerance and ABA responses in wheat (Triticum aestivum L.). Crop J. 2021, 9, 785–793. [Google Scholar] [CrossRef]
- Yan, J.; Li, J.; Zhang, H.; Liu, Y.; Zhang, A. ZmWRKY104 positively regulates salt tolerance by modulating ZmSOD4 expression in maize. Crop J. 2022, 10, 555–564. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Q.; Gao, Z.; Wang, Y.; Li, J. Analysis of Phytohormone Signal Transduction in Sophora alopecuroides under Salt Stress. Int. J. Mol. Sci. 2021, 22, 7313. [Google Scholar] [CrossRef]
- Xiu, Y.W.; Tian, L. A casein kinase II phosphorylation site in AtYY1 affects its activity, stability, and function in the ABA response. Front. Plant Sci. 2017, 8, 323. [Google Scholar]
- Xue, T.; Wang, D.; Zhang, S.; Ehlting, J.; Ni, F.; Jakab, S.; Zheng, C.; Zhong, Y. Genome-wide and expression analysis of protein phosphatase 2C in rice and Arabidopsis. BMC Genom. 2008, 9, 550. [Google Scholar] [CrossRef]
- Chu, M.; Chen, P.; Meng, S.; Xu, P.; Lan, W. The Arabidopsis phosphatase PP2C49 negatively regulates salt tolerance through inhibition of AtHKT1;1. J. Integr. Plant Biol. 2021, 63, 528–542. [Google Scholar] [CrossRef]
- Hiroki, S.; Satoshi, K.; Tomoko, T.; Chie, I.; Nobuhiko, M.; Etsuko, H.; Ken’Ichi, O.; Norihiro, M.; Chikara, O. Overexpression of a novel Arabidopsis PP2C isoform, AtPP2CF1, enhances plant biomass production by increasing inflorescence stem growth. J. Exp. Bot. 2014, 65, 5385–5400. [Google Scholar]
- Zhang, P.; Yuan, Z.; Wei, L.; Qiu, X.; Wang, G.; Liu, Z.; Fu, J.; Cao, L.; Wang, T. Overexpression of ZmPP2C55 positively enhances tolerance to drought stress in transgenic maize plants. Plant Sci. 2022, 314, 111127. [Google Scholar] [CrossRef]
- Sang, K.S.; Myeong, M.L.; Clark, S.E. POL and PLL1 phosphatases are CLAVATA1 signaling intermediates required for Arabidopsis shoot and floral stem cells. Development 2006, 133, 4691–4698. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.; Bouvier, D.M.; Morris, P.C.; Guerrier, D.; Chefdor, F.; Giraudat, J. Arabidopsis ABA response gene ABI1: Features of a calcium-modulated protein phosphatase. Science 1994, 264, 1448–1452. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Leube, M.P.; Grill, E. A Protein Phosphatase 2C Involved in ABA Signal Transduction in Arabidopsis thaliana. Science 1994, 264, 1452–1455. [Google Scholar] [CrossRef]
- Lozano, J.J.; Infantes, L.; García-Maquilón, I.; Ruiz, P.R.; Merilo, E.; Benavente, J.L.; Velázquez-Campoy, A.; Coego, A.; Bono, M.; Forment, J. Structure-guided engineering of a receptor-agonist pair for inducible activation of the ABA adaptive response to drought. Sci. Adv. 2023, 9, 9948. [Google Scholar]
- Xiang, Y.; Sun, X.; Gao, S.; Qin, F.; Dai, M. Deletion of an endoplasmic reticulum stress response element in a ZmPP2C-A gene facilitates drought tolerance of maize seedlings. Mol. Plant 2017, 10, 456–469. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; Wu, J.F.; Sun, X.P.; Dai, M.Q. The maize clade a PP2C phosphatases play critical roles in multiple abiotic stress responses. Int. J. Mol. Sci. 2019, 20, 3573. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Shi, W.S.; Liu, Y.; Gao, X.M.; Hu, B.; Sun, H.R.; Li, X.Y.; Yang, Y.; Li, X.F.; Liu, Z.B.; et al. MdPP2C24/37, Protein Phosphatase Type 2Cs from apple, interact with MdPYL2/12 to negatively regulate ABA signaling in transgenic arabidopsis. Int. J. Mol. Sci. 2022, 23, 14375. [Google Scholar] [CrossRef]
- Kang, Y.C.; Yang, X.Y.; Liu, Y.H.; Shi, M.F.; Zhang, W.N.; Fan, Y.L.; Yao, Y.H.; Zhang, J.L.; Qin, S.H. Integration of mRNA and miRNA analysis reveals the molecular mechanism of potato (Solanum tuberosum L.) response to alkali stress. Int. J. Biol. Macromol. 2021, 182, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | Time | Cycles | |
|---|---|---|---|
| Step 1 | 95 °C | 30 s | 1 |
| Step 2 | 95 °C | 5 s | 50 |
| 60 °C | 30 s |
| Gene ID | Forward Primer | Reverse Primer |
|---|---|---|
| PGSC0003DMT400067426 | ATGGAGGAAAAGTATGAGCTTTTGA | TCAGACATAAACAGCAAAGTCA |
| PGSC0003DMT400060760 | ATGGAAAGATATGAAATTCAGAAAGAC | TCATACCATTGAATGACAGTAACTCAC |
| PGSC0003DMT400060264 | ATGCAGAATTACGAAGTTGTGAAGG | CGAGGTGCTGGACTTCCATC |
| PGSC0003DMT400046381 | ATGACAGTTGCTGATTGCCA | CTACGTTTTCTTCTTGAATTTCCTCTG |
| PGSC0003DMT400075332 | ATGGAGGAGATGTATATGGTTGCA | TTAGCTAGGAGAAAACATACCGA |
| PGSC0003DMT400077988 | ATGGCAGAGGTCTGTTTTGGA | TTAACGTACATGAGAGCAGCCA |
| PGSC0003DMT400046381 | ATGACAGTTGCTGATTGCCA | CTACGTTTTCTTCTTGAATTTCCTCTG |
| PGSC0003DMT400005378 | ATGGAGCAATCCGATAACTCA | TTAGGAGATCTCACCGTTACCAC |
| PGSC0003DMT400023511 | ATGAACGCTAATGGATTCTGCG | TTAGACCTGATCAATGGGTTCTGT |
| PGSC0003DMT400028658 | ATGCCTCCCAGTTCTTCAGA | TCATCTGCTTGAATTCCGTGC |
| PGSC0003DMT400045156 | ATGGATAGTAAACCGGAAACGTCA | TCACCTGTGACTTACATCACT |
| PGSC0003DMT400039054 | ATGACTTCCACAATTGCAAA | TTATACTTGATTTTGCAAGTCCT |
| PGSC0003DMT400004980 | ATGCCGAAAGTAATAGGGATAGCA | TTATAGTTTCATAAGATCATTTTCCGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Liu, Z.; Chen, J.; Zhang, W.; Yang, W.; Shen, C.; Kang, Y.; Qin, S. PP2C-Mediated ABA Signaling Pathway Underlies Exogenous Abscisic Acid-Induced Enhancement of Saline–Alkaline Tolerance in Potato (Solanum tuberosum L.). Plants 2025, 14, 1921. https://doi.org/10.3390/plants14131921
Yang X, Liu Z, Chen J, Zhang W, Yang W, Shen C, Kang Y, Qin S. PP2C-Mediated ABA Signaling Pathway Underlies Exogenous Abscisic Acid-Induced Enhancement of Saline–Alkaline Tolerance in Potato (Solanum tuberosum L.). Plants. 2025; 14(13):1921. https://doi.org/10.3390/plants14131921
Chicago/Turabian StyleYang, Xinhui, Zisheng Liu, Jun Chen, Wangjun Zhang, Wenhui Yang, Congang Shen, Yichen Kang, and Shuhao Qin. 2025. "PP2C-Mediated ABA Signaling Pathway Underlies Exogenous Abscisic Acid-Induced Enhancement of Saline–Alkaline Tolerance in Potato (Solanum tuberosum L.)" Plants 14, no. 13: 1921. https://doi.org/10.3390/plants14131921
APA StyleYang, X., Liu, Z., Chen, J., Zhang, W., Yang, W., Shen, C., Kang, Y., & Qin, S. (2025). PP2C-Mediated ABA Signaling Pathway Underlies Exogenous Abscisic Acid-Induced Enhancement of Saline–Alkaline Tolerance in Potato (Solanum tuberosum L.). Plants, 14(13), 1921. https://doi.org/10.3390/plants14131921