

Overexpression of SEPALLATA3-like Gene SnMADS37 Generates Green Petal-Tip Flowers in Solanum nigrum
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Isolation of SEPALLATA3-like Gene SnMADS37
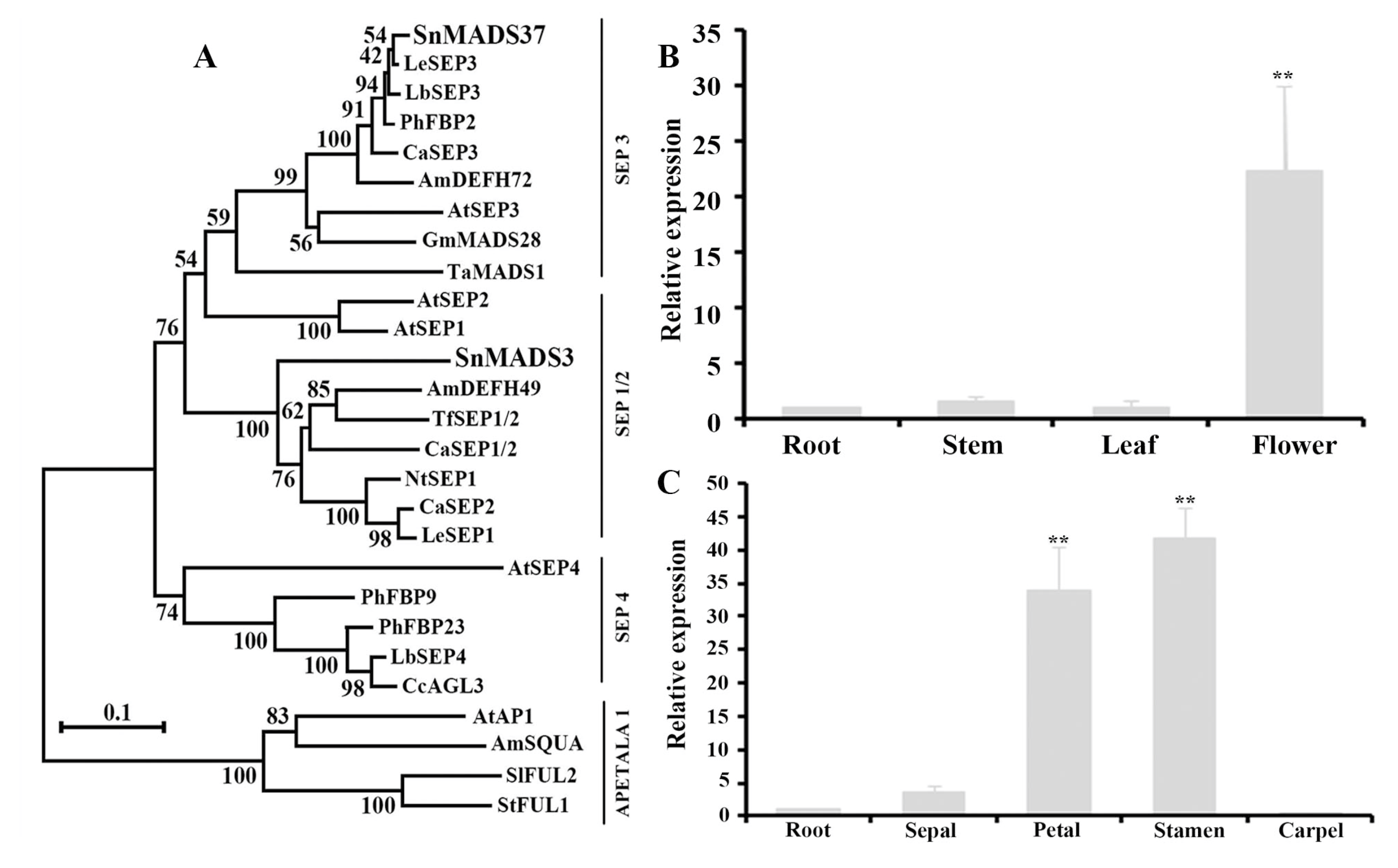
2.2. Phylogenetic Tree Analysis
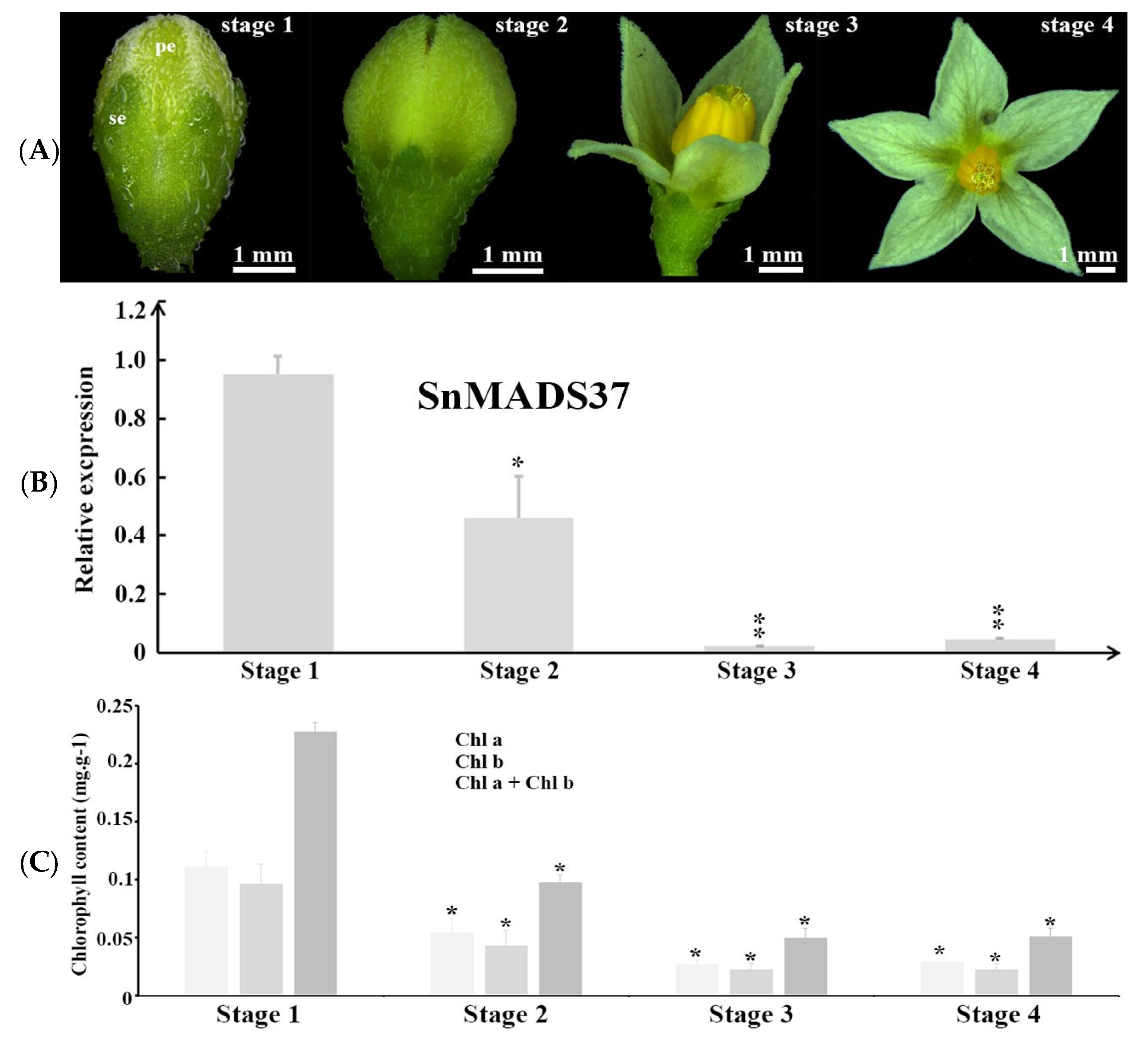
2.3. Expression Analysis of SnMADS37
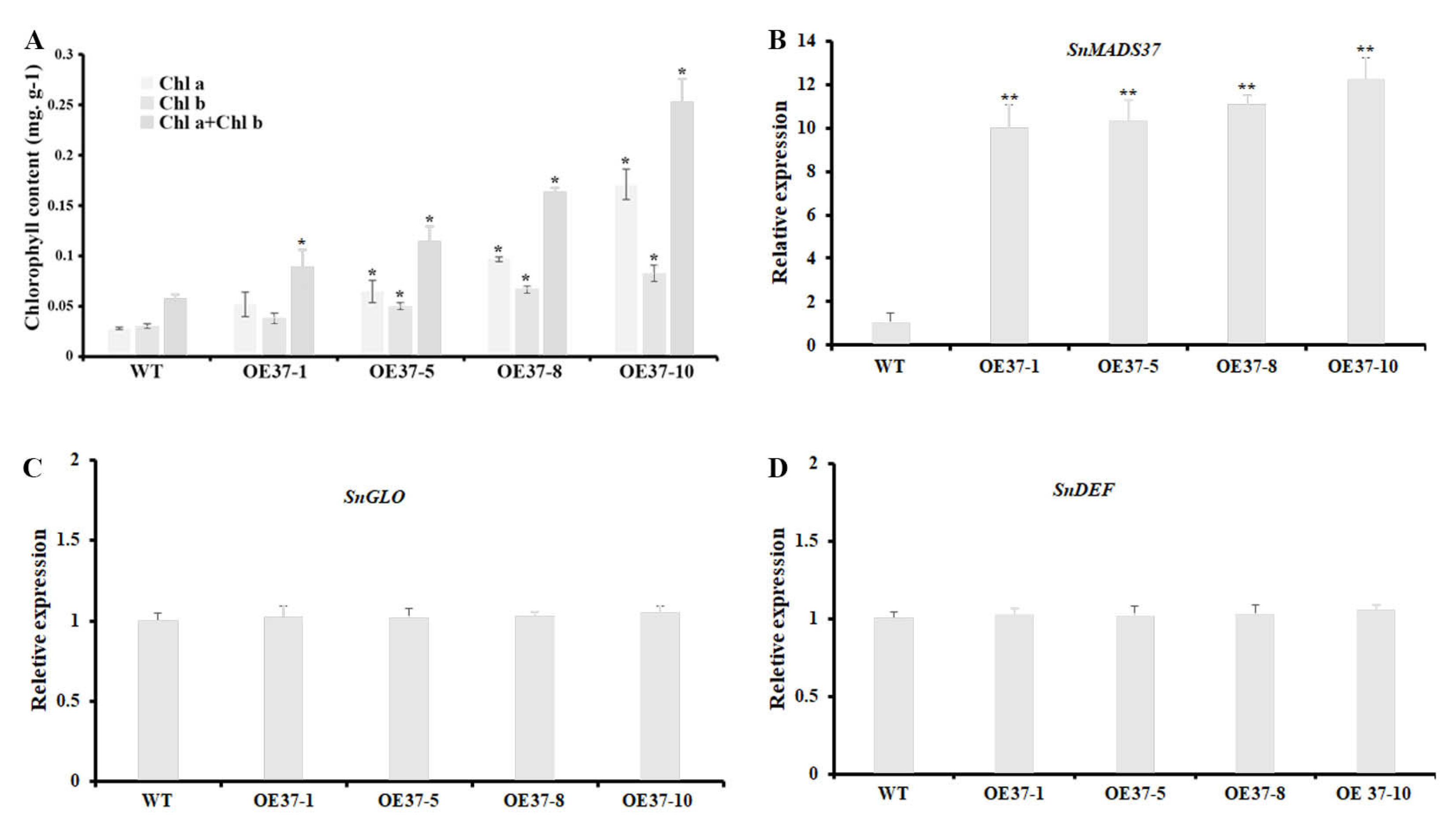
2.4. Overexpression of SnMADS37 Leads to Morphological Changes in the Flower
2.5. SEM and Sliced Section Analysis
2.6. Expression of SnMADS37 Affects Chlorophyll Content in Flower
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. SnMADS37 Gene Cloning
4.3. Phylogenetic Tree Construction
4.4. Transcriptional Analysis of SnMADS37
4.5. Construction of the Overexpression Vector pBI-35S::SnMADS37
4.6. Transformation of S. nigrum
4.7. SnMADS37 Expression Analysis in the Transgenic S. nigrum
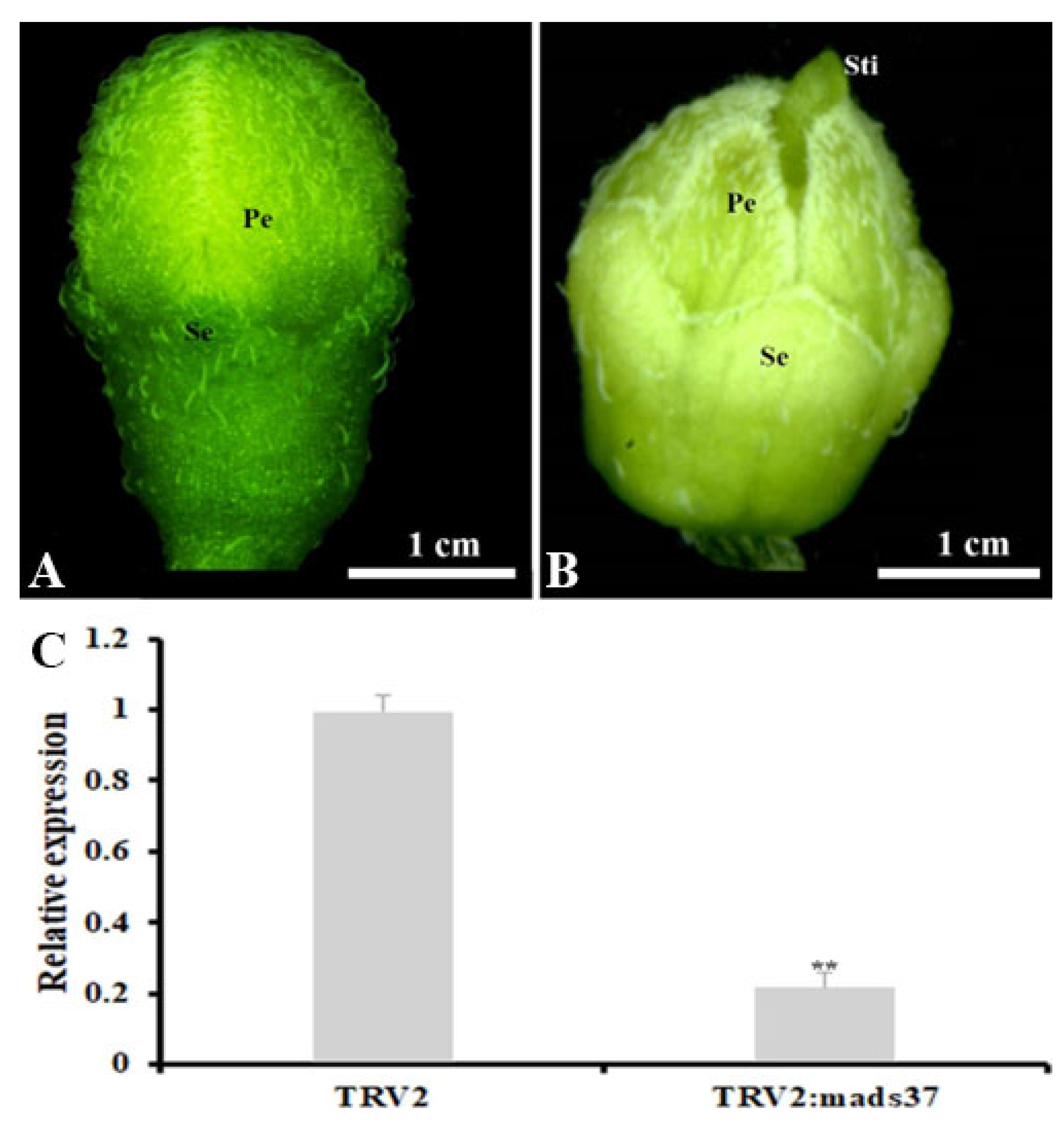
4.8. VIGS-Mediated Silencing of SnMADS37 in Solanum nigrum Flowers
4.9. Scanning Electron Microscopy (SEM) Observations
4.10. Slice Analysis of Petals
4.11. Measurement of Chlorophyll Pigment Content
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Edmonds, J.; Chweya, J.A. Black nightshades. Solanum nigrum L. and related species. In Promoting the Conservation and Use of Underutilized and Neglected Crops, 15th ed.; Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute: Rome, Italy, 1997; ISBN 92-9043-321-3. [Google Scholar]
- Lehmann, C.; Biela, C.; Töpfl, S.; Jansen, G.; Vögel, R. Solanum scabrum a potential source of a coloring plant extract. Euphytica 2007, 158, 189–199. [Google Scholar] [CrossRef]
- Poczai, P.; Hyvönen, J. On the origin of Solanum nigrum, can networks help? Mol. Biol. Rep. 2011, 38, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Jabamalairaj, A.; Priatama, R.A.; Heo, J.; Park, S.J. Medicinal metabolites with common biosynthetic pathways in Solanum nigrum. Plant Biotech. Rep. 2019, 13, 315–327. [Google Scholar] [CrossRef]
- Sommer, H.; Beltrán, J.P.; Huijser, P.; Pape, H.; Lönnig, W.E.; Saedler, H.; Schwarz-Sommer, Z. Defciens, a homeotic gene involved in the control of flower morphogenesis in Antirrhinum majus, the protein shows homology to transcription factors. EMBO J. 1990, 9, 605–613. [Google Scholar] [CrossRef]
- Yanofsky, M.F.; Ma, H.; Bowman, J.L.; Drews, G.N.; Feldmann, K.A.; Meyerowitz, E.M. The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors. Nature 1990, 346, 35–39. [Google Scholar] [CrossRef]
- Gan, Y.; Bernreiter, A.; Filleur, S.; Abram, B.; Forde, B.G. Overexpressing the ANR1 MADS-box gene in transgenic plants provides new insights into its role in the nitrate regulation of root development. Plant Cell Physiol. 2012, 53, 1003–1016. [Google Scholar] [CrossRef]
- Lightfoot, D.J.; Malone, K.M.; Timmis, J.N.; Orford, S.J. Evidence for alternative splicing of MADS-box transcripts in developing cotton fibre cells. Mol. Gent. Genom. 2008, 279, 75–85. [Google Scholar] [CrossRef]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31–37. [Google Scholar] [CrossRef]
- Garceau, D.C.; Batson, M.K.; Pan, I.L. Variations on theme in fruit development the PLE lineage of MADS-box genes in tomato (TAGL1) and other species. Planta 2017, 246, 313–321. [Google Scholar] [CrossRef]
- Martin-Pizarro, C.; Trivino, J.C.; Pose, D. Functional analysis of the TM6 MADS-box gene in the octoploid strawberry by CRISPR/Cas9-directed mutagenesis. J. Exp. Bot. 2019, 70, 885–895. [Google Scholar] [CrossRef]
- Theißen, G.; Saedler, H. Floral quartets. Nature 2001, 409, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Goto, K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 2001, 409, 525–529. [Google Scholar] [CrossRef]
- Krizek, B.A.; Fletcher, J.C. Molecular mechanisms of flower development, an armchair guide. Nat. Rev. Genet. 2005, 6, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Theißen, G.; Melzer, R.; Rümpler, F. MADS-domain transcription factors and the floral quartet model of flower development: Linking plant development and evolution. Development 2016, 143, 3259–3271. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, M.; Zethof, J.; Souer, E.; Koes, R.; Tornielli, G.B.; Pezzotti, M.; Ferrario, S.; Angenent, G.C.; Gerats, T. Toward the analysis of the petunia MADS box gene family by reverse and forward transposon insertion mutagenesis approaches: B, C, and D floral organ identity functions require SEPALLATA-like MADS box genes in petunia. Plant Cell 2003, 15, 2680–2693. [Google Scholar] [CrossRef]
- Ampomah-Dwamena, C.; Morris, B.A.; Sutherland, P.; Veit, B.; Yao, J.L. Down-regulation of TM29, a tomato SEPALLATA homolog, causes parthenocarpic fruit development and floral reversion. Plant Physiol. 2002, 130, 605–617. [Google Scholar] [CrossRef]
- Matsubara, K.; Shimamura, K.; Kodama, H.; Kokubun, H.; Watanabe, H.; Basualdo, I.L.; Ando, T. Green corolla segments in a wild Petunia species caused by a mutation in FBP2, a SEPALLATA-like MADS box gene. Planta 2008, 228, 401–409. [Google Scholar] [CrossRef]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef]
- Ditta, G.; Pinyopich, A.; Robles, P.; Pelaz, S.; Yanofsky, M.F. The SEP4 gene of Arabidopsis thaliana functions in floral organ and meristem identity. Curr. Biol. 2004, 14, 1935–1940. [Google Scholar] [CrossRef]
- Kotilainen, M.; Elomaa, P.; Uimari, A.; Albert, V.A.; Yu, D.; Teeri, T.H. GRCD1, an AGL2-like MADS box gene, participates in the C function during stamen development in Gerbera hybrida. Plant Cell 2000, 12, 1893–1902. [Google Scholar] [CrossRef]
- Uimari, A.; Kotilainen, M.; Elomaa, P.; Yu, D.; Albert, V.A.; Teeri, T.H. Integration of reproductive meristem fates by a SEPALLATA-like MADS-box gene. Pro. Natl. Acad. Sci. USA 2004, 101, 15817–15822. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.; Juntheikki, I.; Mouhu, K.; Broholm, S.K.; Rijpkema, A.S.; Kins, L.; Lan, T.; Albert, V.A.; Teeri, T.H.; et al. Dissecting functions of SEPALLATA-like MADS box genes in patterning of the pseudanthial inflorescence of Gerbera Hybrida. New Phytol. 2017, 216, 939–954. [Google Scholar] [CrossRef]
- Castillejo, C.; Romera-Branchat, M.; Pelaz, S. A new role of the Arabidopsis SEPALLATA3 gene revealed by its constitutive expression. Plant J. 2005, 43, 586–596. [Google Scholar] [CrossRef]
- Adal, A.M.; Binson, E.; Remedios, L.; Mahmoud, S.S. Expression of lavender AGAMOUS-like and SEPALLATA3-like genes promote early flowering and alter leaf morphology in Arabidopsis thaliana. Planta 2021, 254, 54. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Cheng, Z.J.; Zhang, X.S. Overexpression of TaMADS1, a SEPALLATA-like gene in wheat, causes early flowering and the abnormal development of floral organs in Arabidopsis. Planta 2006, 223, 698–707. [Google Scholar] [CrossRef]
- Lemmetyinen, J.; Hassinen, M.; Elo, A.; Porali, I.; Sopanen, T. Functional characterization of SEPALLATA3 and AGAMOUS orthologues in silver birch. Physiol. Plant. 2004, 121, 149–162. [Google Scholar] [CrossRef]
- Slugina, M.A.; Dyachenko, E.A.; Kochieva, E.Z.; Shchennikova, A.V. Structural and functional diversification of SEPALLATA genes TM5 and RIN in tomato species (Section Lycopersicon). Dokl. Biochem.Biophys. 2020, 492, 152–158. [Google Scholar] [CrossRef]
- Ruelens, P.; Zhang, Z.; van Mourik, H.; Maere, S.; Kaufmann, K.; Geuten, K. The origin of floral organ identity quartets. Plant Cell 2017, 29, 229–242. [Google Scholar] [CrossRef]
- Huang, F.; Xu, G.; Chi, Y.; Liu, H.; Xue, Q.; Zhao, T.; Gai, J.; Yu, D. A soybean MADS-box protein modulates floral organ numbers, petal identity and sterility. BMC Plant Biol. 2014, 14, 89. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, S.; Yi, S.; Han, H.; Liu, L.; Zhang, J.; Bao, M.; Liu, G. Functional conservation and divergence of five SEPALLATA-like genes from a basal eudicot tree, Platanus acerifolia. Planta 2017, 245, 439–457. [Google Scholar]
- Angenent, G.C.; Franken, J.; Busscher, M.; Weiss, D.; Van Tunen, A.J. Co-suppression of the Petunia homeotic gene Fbp2 affects the identity of the generative meristem. Plant J. 1994, 5, 33–44. [Google Scholar] [CrossRef]
- Ferrario, S.; Immink, R.G.H.; Shchennikova, A.; Busscher-Lange, J.; Angenent, G.C. The MADS-box gene FBP2 is required for SEPALLATA function in Petunia. Plant Cell 2003, 15, 914–925. [Google Scholar] [CrossRef]
- Pnueli, L.; Hareven, D.; Broday, L.; Hurwitz, C.; Lifschitz, E. The TM5 MADS box gene mediates organ differentiation in the three inner whorls of tomato flowers. Plant Cell 1994, 6, 175–186. [Google Scholar] [CrossRef]
- Zhao, M.L.; Zhou, Z.F.; Chen, M.S.; Xu, C.J.; Xu, Z.F. An ortholog of the MADS-box gene SEPALLATA3 regulates stamen development in the woody plant Jatropha curcas. Planta 2022, 255, 111. [Google Scholar] [CrossRef]
- Whitney, H.M.; Bennett, K.M.; Dorling, M.; Sandbach, L.; Prince, D.; Chittka, L.; Glover, B.J. Why do so many petals have conical epidermal cells? Annal. Bot. 2011, 108, 609–616. [Google Scholar] [CrossRef]
- Wang, Z.; Shen, Y.; Yang, X.; Pan, Q.; Ma, G.; Bao, M.; Zheng, B.; Duanmu, D.; Lin, R.; Larkin, R.M.; et al. Overexpression of particular MADS-box transcription factors in heat-stressed plants induces chloroplast biogenesis in petals. Plant Cell Environ. 2019, 42, 1545–1560. [Google Scholar] [CrossRef]
- Qu, G.Z.; Zheng, T.; Liu, G.; Wang, W.; Zang, L.; Liu, H.; Yang, C. Overexpression of a MADS-Box Gene from Birch (Betula platyphylla) Promotes Flowering and Enhances Chloroplast Development in Transgenic Tobacco. PLoS ONE 2013, 8, e63398. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 1962, 15, 81–84. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetic analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.L.; Liu, C.; Piao, C.; Liu, C.L. A stable Agrobacterium rhizogenes-mediated transformation of cotton (Gossypium hirsutum L.) and plant regeneration from transformed hairy root via embryogenesis. Front. Plant Sci. 2020, 11, 604255. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.; Gao, Z.; Yuan, S.; Cui, M.L. The R2R3-MYB gene CgMYB4 is involved in the regulation of cell differentiation and fiber development in the stamens of Chelone glabra L. Protoplasma 2022, 259, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Zhang, C.; Jiang, X.; Kang, M.; Yin, X.; Lü, P.; Zhang, X.; Zheng, Y.; Gao, J. RhNAC2 and RhEXPA4 are involved in the regulation of dehydration tolerance during the expansion of rose petals. Plant Physiol. 2012, 160, 2064–2082. [Google Scholar] [CrossRef]
- Hartl, M.; Merker, H.; Schmidt, D.D.; Baldwin, I.T. Optimized virus-induced gene silencing in Solanum nigrum reveals the defensive function of leucine aminopeptidase against herbivores and the shortcomings of empty vector controls. New Phytol. 2008, 179, 356–365. [Google Scholar] [CrossRef]
- Krizek, B.A. Overexpression of AINTEGUMENTA in Arabidopsis plants results in increased growth of floral organs. Dev. Genet. 1999, 25, 224–236. [Google Scholar] [CrossRef]
- Zhou, J.; Han, P.; Pan, Y.; Wu, M.; Zhao, Y.; Jia, Y.; Jiang, B.; Zhang, L.; Xu, Q.; Liu, S.; et al. Effects of cadmium stress on photosynthetic physiology and chlorophyll fluorescence in Solanum nigrum and Solanum americanum. J. Agro-Environ. Sci. 2021, 40, 26–34. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, S.; Piao, C.-L.; Zhang, X.; Cui, M.-L. Overexpression of SEPALLATA3-like Gene SnMADS37 Generates Green Petal-Tip Flowers in Solanum nigrum. Plants 2025, 14, 1891. https://doi.org/10.3390/plants14131891
Yuan S, Piao C-L, Zhang X, Cui M-L. Overexpression of SEPALLATA3-like Gene SnMADS37 Generates Green Petal-Tip Flowers in Solanum nigrum. Plants. 2025; 14(13):1891. https://doi.org/10.3390/plants14131891
Chicago/Turabian StyleYuan, Siming, Chun-Lan Piao, Xinyu Zhang, and Min-Long Cui. 2025. "Overexpression of SEPALLATA3-like Gene SnMADS37 Generates Green Petal-Tip Flowers in Solanum nigrum" Plants 14, no. 13: 1891. https://doi.org/10.3390/plants14131891
APA StyleYuan, S., Piao, C.-L., Zhang, X., & Cui, M.-L. (2025). Overexpression of SEPALLATA3-like Gene SnMADS37 Generates Green Petal-Tip Flowers in Solanum nigrum. Plants, 14(13), 1891. https://doi.org/10.3390/plants14131891