
“End-to-End Chromosome Fusion” as the Main Driver of Descending Dysploidy in Vigna lasiocarpa (Mart. ex Benth.) Verdc. (Leguminosae Juss.)


 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
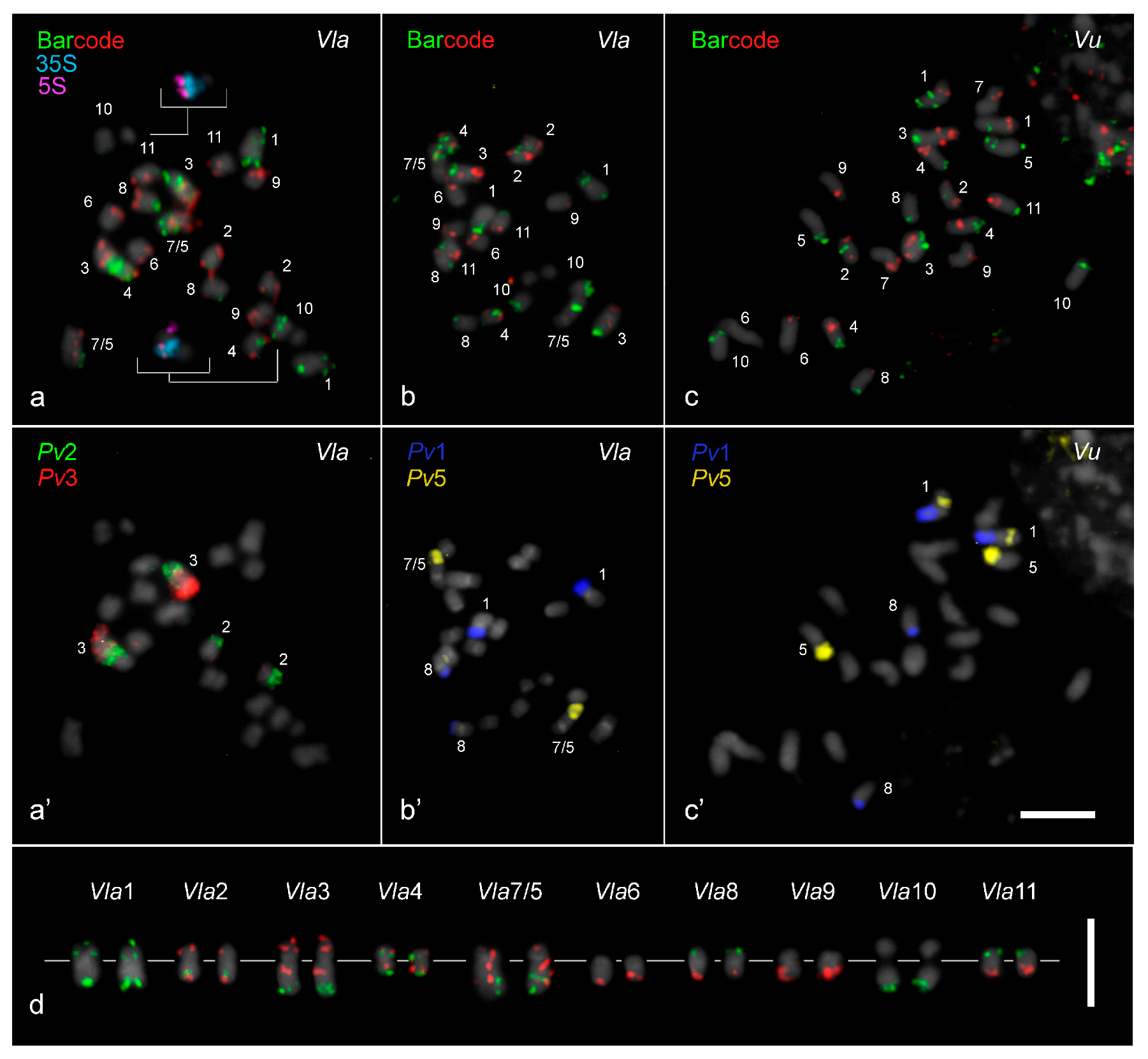
2.1. Chromosome Mapping and rDNA Sites in V. lasiocarpa
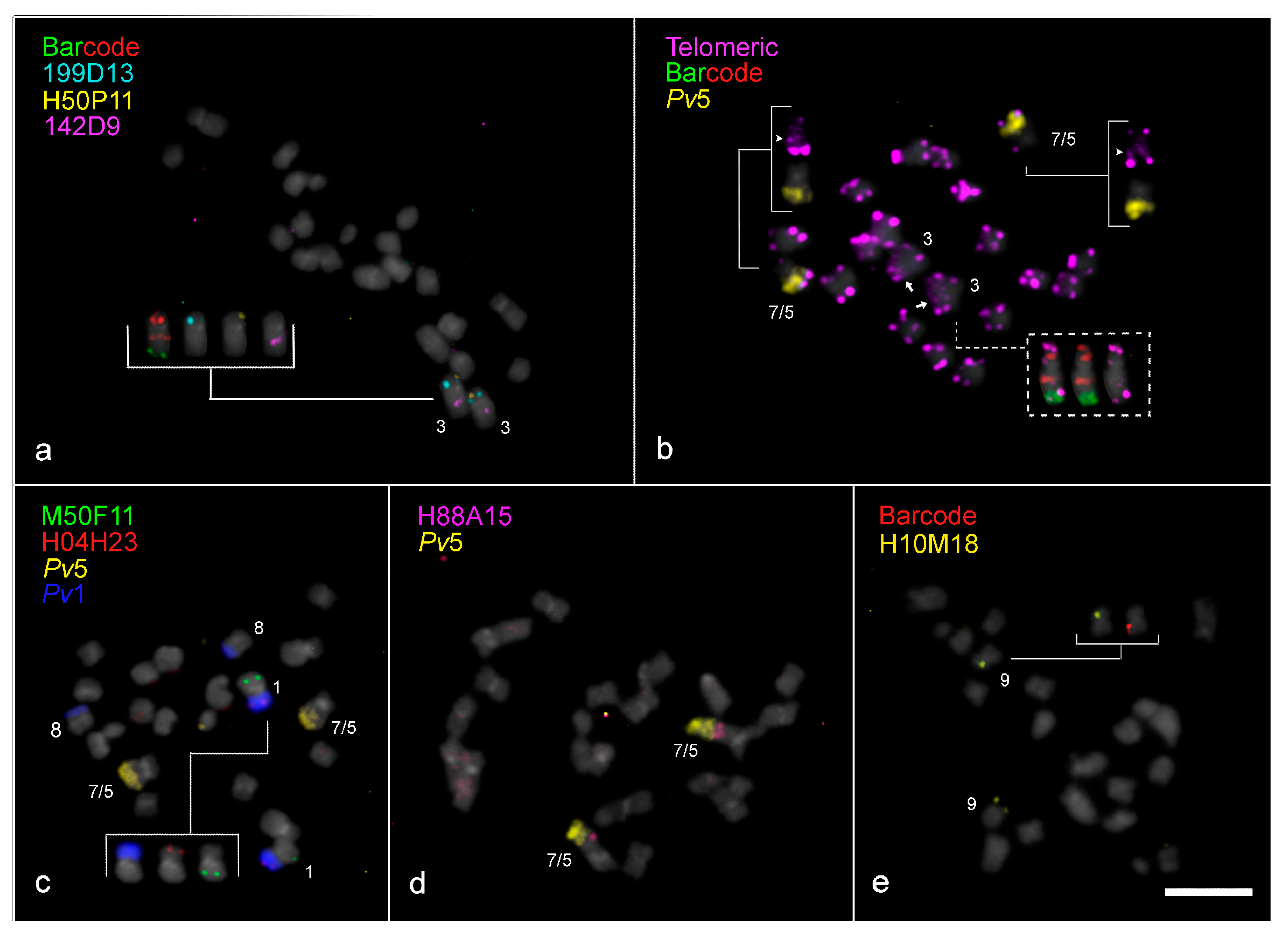
2.2. Translocation Among Vigna Chromosomes 1, 5, and 8
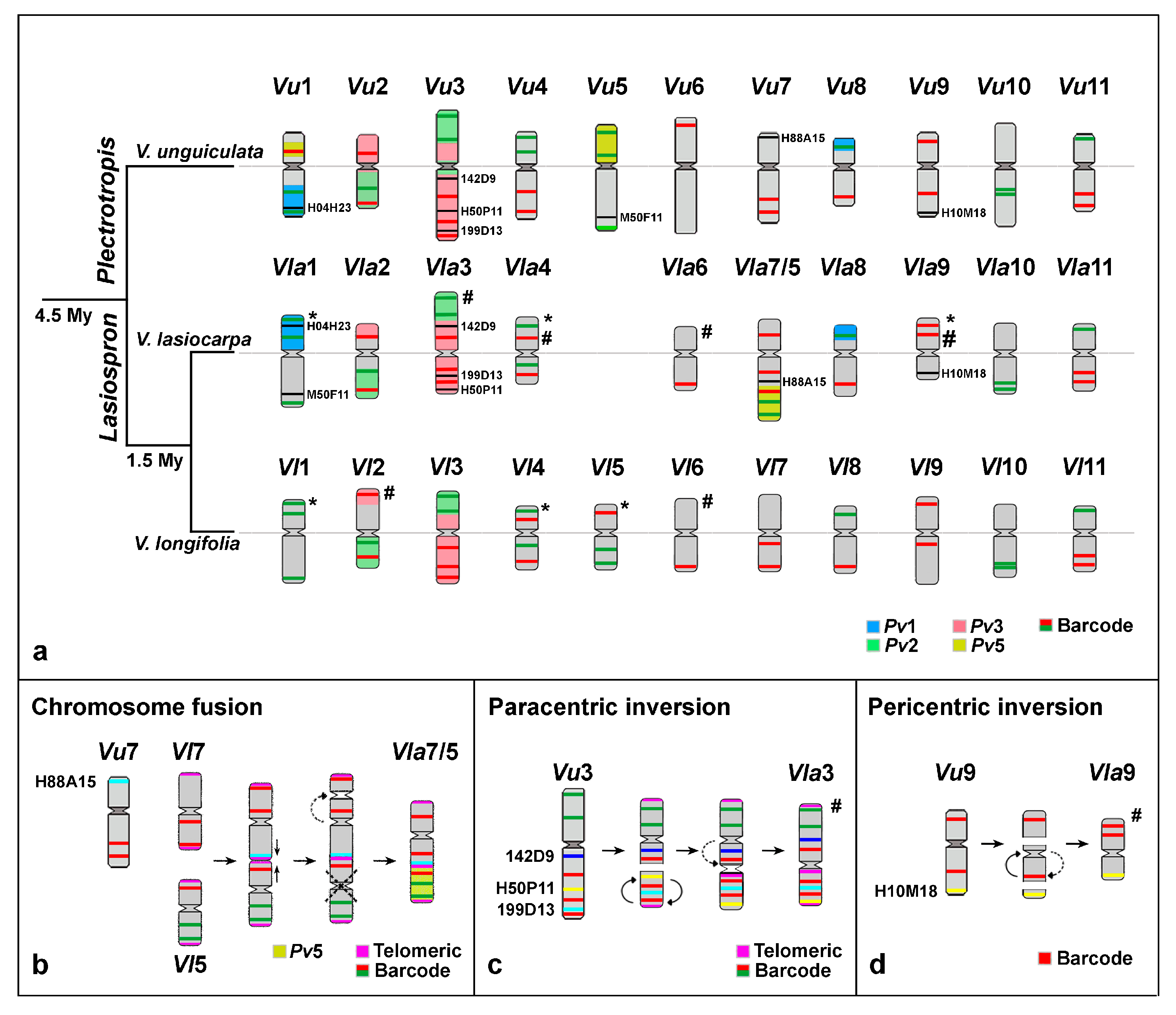
2.3. Chromosomal Rearrangements Associated with the Evolution of V. lasiocarpa
2.3.1. Paracentric Inversion on Vla3 Confirmed by the Presence of an Additional Interstitial Telomeric Signal
2.3.2. “End-to-End Vla7/5 Chromosome Fusion” as the Major Driver for Descending Dysploidy in V. lasiocarpa
2.3.3. Exclusive Pericentric Inversion on Vla9
3. Discussion
3.1. Chromosomal Rearrangements Associated with Descending Dysploidy and Evolution of V. lasiocarpa
3.2. Positional Centromere Changes on V. lasiocarpa Chromosomes
3.3. Comparative Analysis Between Two Species of the American subg. Lasiospron: V. longifolia and V. lasiocarpa
4. Materials and Methods
4.1. Plant Material and Chromosome Preparation
4.2. FISH Probe Sets
4.3. FISH and Image Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FISH | Fluorescence in situ hybridization |
| BAC | Bacterial artificial chromosome |
| EMBRAPA | Empresa Brasileira de Pesquisa Agropecuária |
| NGPS | National Plant Germplasm System |
| RT | Room temperature |
| EEF | End-to-end fusion |
| rDNA | Ribosomal DNA |
| IPA | Instituto Agronômico de Pernambuco |
| APK | Ancestral Phaseoleae karyotype |
| GB | Genomic blocks |
| EC | Evolutionary new centromeres |
| DSB | Double-strand breaks |
| ITR | Interstitial telomeric repeats |
| SSC | Saline sodium citrate |
| ChIP-seq | Chromatin immunoprecipitation sequencing |
| Pv | Phaseolus vulgaris |
| Vu | Vigna unguiculata |
| Vl | Vigna longifolia |
| Vla | Vigna lasiocarpa |
References
- Lysak, M.A. Celebrating Mendel, McClintock, and Darlington: On end-to-end chromosome fusions and nested chromosome fusions. Plant Cell 2022, 34, 2475–2491. [Google Scholar] [CrossRef] [PubMed]
- de Storme, N.; Mason, A. Plant speciation through chromosome instability and ploidy change: Cellular mechanisms, molecular factors and evolutionary relevance. Curr. Plant Biol. 2014, 1, 10–33. [Google Scholar] [CrossRef]
- Mandáková, T.; Lysak, M.A. Post-polyploid diploidization and diversification through dysploid changes. Curr. Opin. Plant Biol. 2018, 42, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, W.; Lin, L.; Zhu, X.; Li, J.; Zhu, X.; Chen, Z. Diversification of the Phaseoloid legumes: Effects of climate change, range expansion and habit shift. Front. Plant Sci. 2013, 4, 386. [Google Scholar] [CrossRef]
- Boukar, O.; Belko, N.; Chamarthi, S.; Togola, A.; Batieno, J.; Owusu, E.; Haruna, M.; Diallo, S.; Umar, M.L.; Olufajo, O.; et al. Cowpea (Vigna unguiculata): Genetics, genomics and breeding. Plant Breed. 2019, 138, 415–424. [Google Scholar] [CrossRef]
- Somta, P.; Laosatit, K.; Yuan, X.; Chen, X. Thirty years of mungbean genome research: Where do we stand and what have we learned? Front. Plant Sci. 2022, 13, 944721. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, X.; Shen, H.; Zhao, R.; Li, Z.; Shen, X.; Wang, F.; Chen, K.; Zhou, Y.; Li, B.; et al. Nutritional composition, efficacy, and processing of Vigna angularis (Adzuki Bean) for the human diet: An overview. Molecules 2022, 27, 6079. [Google Scholar] [CrossRef]
- Delgado-Salinas, A.; Thulin, M.; Pasquet, R.; Weeden, N.; Lavin, M. Vigna (Leguminosae) Sensu lato: The names and identities of the American segregate genera. Am. J. Bot. 2011, 98, 1694–1715. [Google Scholar] [CrossRef]
- Delgado-Salinas, A.; Lavin, M.; Snak, C.; Lewis, G.P. Systematics of Vigna subgenus Lasiospron (Leguminosae: Papilionoideae: Phaseolinae). Syst. Bot. 2022, 47, 97–124. [Google Scholar] [CrossRef]
- Horton, D.M.; Feleke, Y.; Pasquet, R.S.; Javadi, F.; Melville, K.A.; Delgado-Salinas, A.; Thulin, M.; Mithen, R.F.; Gepts, P.; Egan, A.N. Phylogenetic systematics of Vigna sensu stricto in the context of Physostigma and allies. Am. J. Bot. 2024, 111, e16381. [Google Scholar] [CrossRef]
- Forni-Martins, E.R. New chromosome number in the genus Vigna Savi (Leguminosae-Papilionoideae). Bull. Jard. Bot. Natl. Belg. /Bull. Natl. Plantentuin Belg. 1986, 56, 129. [Google Scholar] [CrossRef]
- Venoral, G.; Blangifortil, S.; Cremonini, R. Karyotype analysis of twelve species belonging to genus Vigna. Cytologia 1999, 64, 117–127. [Google Scholar] [CrossRef]
- Shamurailatpam, A.; Madhavan, L.; Yadav, S.R.; Bhat, K.V.; Rama Rao, S. Heterochromatin distribution and comparative karyo-morphological studies in Vigna umbellata Thunberg, 1969 and V. aconitifolia Jacquin, 1969 (Fabaceae) accessions. Comp. Cytogenet. 2015, 9, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Galasso, I.; Saponetti, L.S.; Pignone, D. Cytotaxonomic studies in Vigna. III. Chromosomal distribution and reacting properties of the heterochromatin in five wild species of the section Vigna. Caryologia 1996, 49, 311–319. [Google Scholar] [CrossRef]
- Jiang, J. Fluorescence in Situ Hybridization in plants: Recent developments and future applications. Chromosome Res. 2019, 27, 153–165. [Google Scholar] [CrossRef]
- Braz, G.T.; He, L.; Zhao, H.; Zhang, T.; Semrau, K.; Rouillard, J.-M.; Torres, G.A.; Jiang, J. Comparative oligo-FISH mapping: An efficient and powerful methodology to reveal karyotypic and chromosomal evolution. Genetics 2018, 208, 513–523. [Google Scholar] [CrossRef]
- He, L.; Braz, G.T.; Torres, G.A.; Jiang, J. Chromosome painting in meiosis reveals pairing of specific chromosomes in polyploid Solanum Species. Chromosoma 2018, 127, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef]
- Zhao, Q.; Meng, Y.; Wang, P.; Qin, X.; Cheng, C.; Zhou, J.; Yu, X.; Li, J.; Lou, Q.; Jahn, M.; et al. Reconstruction of ancestral karyotype illuminates chromosome evolution in the genus Cucumis. Plant J. 2021, 107, 1243–1259. [Google Scholar] [CrossRef]
- do Vale Martins, L.; Yu, F.; Zhao, H.; Dennison, T.; Lauter, N.; Wang, H.; Deng, Z.; Thompson, A.; Semrau, K.; Rouillard, J.-M.; et al. Meiotic crossovers characterized by haplotype-specific chromosome painting in maize. Nat. Commun. 2019, 10, 4604. [Google Scholar] [CrossRef]
- Wang, K.; Cheng, H.; Han, J.; Esh, A.; Liu, J.; Zhang, Y.; Wang, B. A comprehensive molecular cytogenetic analysis of the genome architecture in modern sugarcane cultivars. Chromosome Res. 2022, 30, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Doležalová, A.; Sládeková, L.; Šimoníková, D.; Holušová, K.; Karafiátová, M.; Varshney, R.K.; Doležel, J.; Hřibová, E. Karyotype differentiation in cultivated chickpea revealed by oligopainting fluorescence in situ hybridization. Front. Plant Sci. 2022, 12, 791303. [Google Scholar] [CrossRef]
- Nascimento, T.; Pedrosa-Harand, A. High rates of structural rearrangements have shaped the chromosome evolution in dysploid Phaseolus beans. Theor. Appl. Genet. 2023, 136, 215. [Google Scholar] [CrossRef]
- Oliveira, A.R.D.S.; Martins, L.D.V.; Bustamante, F.D.O.; Muñoz-Amatriaín, M.; Close, T.; Da Costa, A.F.; Benko-Iseppon, A.M.; Pedrosa-Harand, A.; Brasileiro-Vidal, A.C. Breaks of macrosynteny and collinearity among moth bean (Vigna aconitifolia), cowpea (V. unguiculata), and common bean (Phaseolus vulgaris). Chromosome Res. 2020, 28, 293–306. [Google Scholar] [CrossRef] [PubMed]
- do Vale Martins, L.; de Oliveira Bustamante, F.; da Silva Oliveira, A.R.; da Costa, A.F.; de Lima Feitoza, L.; Liang, Q.; Zhao, H.; Benko-Iseppon, A.M.; Muñoz-Amatriaín, M.; Pedrosa-Harand, A.; et al. BAC- and oligo-FISH mapping reveals chromosome evolution among Vigna angularis, V. unguiculata, and Phaseolus vulgaris. Chromosoma 2021, 130, 133–147. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Bustamante, F.; do Nascimento, T.H.; Montenegro, C.; Dias, S.; do Vale Martins, L.; Braz, G.T.; Benko-Iseppon, A.M.; Jiang, J.; Pedrosa-Harand, A.; Brasileiro-Vidal, A.C. Oligo-FISH barcode in beans: A new chromosome identification system. Theor. Appl. Genet. 2021, 134, 3675–3686. [Google Scholar] [CrossRef]
- Dias, S.; de Oliveira Bustamante, F.; do Vale Martins, L.; Da Costa, V.A.; Montenegro, C.; Oliveira, A.R.D.S.; de Lima, G.S.; Braz, G.T.; Jiang, J.; Da Costa, A.F.; et al. Translocations and inversions: Major chromosomal rearrangements during Vigna (Leguminosae) evolution. Theor. Appl. Genet. 2024, 137, 29. [Google Scholar] [CrossRef]
- Hu, H.; Scheben, A.; Wang, J.; Li, F.; Li, C.; Edwards, D.; Zhao, J. Unravelling inversions: Technological advances, challenges, and potential impact on crop breeding. Plant Biotechnol. J. 2024, 22, 544–554. [Google Scholar] [CrossRef]
- Liu, Y.; Du, H.; Li, P.; Shen, Y.; Peng, H.; Liu, S.; Zhou, G.-A.; Zhang, H.; Liu, Z.; Shi, M.; et al. Pan-genome of wild and cultivated soybeans. Cell 2020, 182, 162–176.e13. [Google Scholar] [CrossRef]
- Vasconcelos, E.V.; de Andrade Fonsêca, A.F.; Pedrosa-Harand, A.; de Andrade Bortoleti, K.C.; Benko-Iseppon, A.M.; Da Costa, A.F.; Brasileiro-Vidal, A.C. Intra- and Interchromosomal rearrangements between cowpea [Vigna unguiculata (L.) Walp.] and common bean (Phaseolus vulgaris L.) revealed by BAC-FISH. Chromosome Res. 2015, 23, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Fonsêca, A.; Pedrosa-Harand, A. karyotype stability in the genus Phaseolus evidenced by the comparative mapping of the wild species Phaseolus microcarpus. Genome 2013, 56, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.; Brandes, A.; Schubert, I. Telomere sequence localization and karyotype evolution in higher plants. Plant Syst. Evol. 1995, 196, 227–241. [Google Scholar] [CrossRef]
- Sousa, A.; Renner, S.S. Interstitial telomere-like repeats in the monocot family Araceae: Chromosome evolution in the Araceae. Bot. J. Linn. Soc. 2015, 177, 15–26. [Google Scholar] [CrossRef]
- Schubert, I.; Lysak, M.A. Interpretation of karyotype evolution should consider chromosome structural constraints. Trends Genet. 2011, 27, 207–216. [Google Scholar] [CrossRef]
- Wang, X.; Jin, D.; Wang, Z.; Guo, H.; Zhang, L.; Wang, L.; Li, J.; Paterson, A.H. Telomere-centric genome repatterning determines recurring chromosome number reductions during the evolution of eukaryotes. New Phytol. 2015, 205, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Mandáková, T.; Pouch, M.; Harmanová, K.; Zhan, S.H.; Mayrose, I.; Lysak, M.A. Multispeed genome diploidization and diversification after an ancient allopolyploidization. Mol. Ecol. 2017, 26, 6445–6462. [Google Scholar] [CrossRef]
- Mercado-Ruaro, P.; Delgado-Salinas, A. Karyological studies in several mexican species of Phaseolus L. and Vigna Savi (Phaseolinae, Fabaceae). Adv. Legume Syst. 1996, 8, 83–87. [Google Scholar]
- Mercado-Ruaro, P.; Delgado-Salinas, A. Karyotypic studies on species of Phaseolus (Fabaceae: Phaseolinae). Am. J. Bot. 1998, 85, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.; Vasconcelos, E.; dos Santos, K.G.B.; Vaio, M.; Brasileiro-Vidal, A.C.; Pedrosa-Harand, A. Diversity of repetitive sequences within compact genomes of Phaseolus L. Beans and Allied Genera Cajanus L. and Vigna Savi. Chromosome Res. 2020, 28, 139–153. [Google Scholar] [CrossRef]
- Ferraz, M.E.; Fonsêca, A.; Pedrosa-Harand, A. Multiple and independent rearrangements revealed by comparative Cytogenetic mapping in the dysploid Leptostachyus group (Phaseolus L., Leguminosae). Chromosome Res. 2020, 28, 395–405. [Google Scholar] [CrossRef]
- Allshire, R.C.; Karpen, G.H. Epigenetic regulation of centromeric chromatin: Old dogs, new tricks? Nat. Rev. Genet. 2008, 9, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef]
- Keçeli, B.N.; Jin, C.; van Damme, D.; Geelen, D. Conservation of centromeric histone 3 interaction partners in plants. J. Exp. Bot. 2020, 71, 5237–5246. [Google Scholar] [CrossRef]
- Schubert, I. What is behind “Centromere repositioning”? Chromosoma 2018, 127, 229–234. [Google Scholar] [CrossRef]
- Mandáková, T.; Hlousková, P.; Koch, M.A.; Lysak, M.A. Genome evolution in Arabideae was marked by frequent centromere repositioning. Plant Cell 2020, 32, 650–665. [Google Scholar] [CrossRef] [PubMed]
- Steckenborn, S.; Marques, A. Centromere diversity and its evolutionary impacts on plant karyotypes and Plant Reproduction. New Phytol. 2025, 245, 1879–1886. [Google Scholar] [CrossRef]
- Montenegro, C.; do Vale Martins, L.; Bustamante, F.D.O.; Brasileiro-Vidal, A.C.; Pedrosa-Harand, A. Comparative cytogenomics reveals genome reshuffling and centromere repositioning in the legume tribe Phaseoleae. Chromosome Res. 2022, 30, 477–492. [Google Scholar] [CrossRef] [PubMed]
- Leo, L.; Marchetti, M.; Giunta, S.; Fanti, L. Epigenetics as an evolutionary tool for centromere flexibility. Genes 2020, 11, 809. [Google Scholar] [CrossRef]
- Marechal, R. Donnees Cytologiques Sur Les Especes de La Sous-Tribu Des Papilionaceae—Phaseoleae—Phaseolinae Premiere Serie. Bull. Jard. Bot. Natl. Belg. 1969, 39, 125. [Google Scholar] [CrossRef]
- Senff, M.; Hickenbick, M.M.; Paim, N.R. Cytogenetic studies in species of the genus Vigna Savi (Leguminosae-Papilionoideae). Rev. Bras. Genét. 1992, 15, 2. [Google Scholar]
- Senff, M.; Schifino-Wittmann, M.T.S.-W.M.; Paim, N.R. Cytogenetic studies of populations of Arachis, Desmodium and Vigna Species (Leguminosae, Papilionoideae) from Rio Grande Do Sul. Braz. J. Genet. 1995, 18, 4. [Google Scholar]
- Dias, S.; Souza, R.C.; Vasconcelos, E.V.; Vasconcelos, S.; da Silva Oliveira, A.R.; do Vale Martins, L.; de Oliveira Bustamante, F.; da Costa, V.A.; Souza, G.; da Costa, A.F.; et al. Cytomolecular diversity among Vigna Savi (Leguminosae) subgenera. Protoplasma 2024, 261, 859–875. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.R.; Saraiva, L.S. An Air Drying technique for maize chromosomes without enzymatic maceration. Biotech. Histochem. 1993, 68, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, C.; Ibiapino, A.; Nascimento, T.; Costa, A.F.D.; Brasileiro-Vidal, A.C.; Pedrosa-Harand, A. Cytogenomic and phylogenomic evidence for new Infrageneric relationships in Macroptilium (Benth.) beans. Ann. Bot. 2024; submitted. [Google Scholar] [CrossRef]
- Lonardi, S.; Muñoz-Amatriaín, M.; Liang, Q.; Shu, S.; Wanamaker, S.I.; Lo, S.; Tanskanen, J.; Schulman, A.H.; Zhu, T.; Luo, M.; et al. The genome of cowpea (Vigna unguiculata [L.] Walp.). Plant J. 2019, 98, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Braz, G.T.; Yu, F.; do Vale Martins, L.; Jiang, J. Fluorescent in situ hybridization using oligonucleotide-based probes. In In Situ Hybridization Protocols, 5th ed.; Nielsen, B.S., Jones, J., Eds.; Springer: New York, NY, USA, 2020; pp. 71–83. [Google Scholar]
- Pedrosa-Harand, A.; Kami, J.; Gepts, P.; Geffroy, V.; Schweizer, D. Cytogenetic mapping of common bean chromosomes reveals a less compartmentalized small-genome plant species. Chromosome Res. 2009, 17, 405–417. [Google Scholar] [CrossRef]
- Pedrosa, A.; Sandal, N.; Stougaard, J.; Schweizer, D.; Bachmair, A. Chromosomal map of the model legume Lotus japonicus. Genetics 2002, 161, 1661–1672. [Google Scholar] [CrossRef]
- Wanzenböck, E.; Schöfer, C.; Schweizer, D.; Bachmair, A. Ribosomal transcription units integrated via T-DNA transformation associate with the nucleolus and do not require upstream repeat sequences for activity in Arabidopsis thaliana. Plant J. 1997, 11, 1007–1016. [Google Scholar] [CrossRef]
- Richards, E.J.; Ausubel, F.M. Isolation of a higher eukaryotic telomere from Arabidopsis thaliana. Cell 1988, 53, 127–136. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S.; Schwarzacher, T.; Anamthawat-Jónsson, K.; Leitch, A.R.; Shi, M.; Leitch, I.J. In Situ Hybridization with automated chromosome denaturation. Tecnhique 1991, 3, 109–115. [Google Scholar]
- Kirov, I.; Khrustaleva, L.; van Laere, K.; Soloviev, A.; Meeus, S.; Romanov, D.; Fesenko, I. DRAWID: User-friendly java software for chromosome measurements and idiogram drawing. Comp. Cytogenet. 2017, 11, 747–757. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serafim, L.; Silva, J.H.; Dias, S.; Oliveira, A.R.d.S.; Nunes, M.C.; da Costa, A.F.; Benko-Iseppon, A.M.; Jiang, J.; do Vale Martins, L.; Brasileiro-Vidal, A.C. “End-to-End Chromosome Fusion” as the Main Driver of Descending Dysploidy in Vigna lasiocarpa (Mart. ex Benth.) Verdc. (Leguminosae Juss.). Plants 2025, 14, 1872. https://doi.org/10.3390/plants14121872
Serafim L, Silva JH, Dias S, Oliveira ARdS, Nunes MC, da Costa AF, Benko-Iseppon AM, Jiang J, do Vale Martins L, Brasileiro-Vidal AC. “End-to-End Chromosome Fusion” as the Main Driver of Descending Dysploidy in Vigna lasiocarpa (Mart. ex Benth.) Verdc. (Leguminosae Juss.). Plants. 2025; 14(12):1872. https://doi.org/10.3390/plants14121872
Chicago/Turabian StyleSerafim, Lazaro, Jarbson Henrique Silva, Sibelle Dias, Ana Rafaela da Silva Oliveira, Maria Clara Nunes, Antônio Félix da Costa, Ana Maria Benko-Iseppon, Jiming Jiang, Lívia do Vale Martins, and Ana Christina Brasileiro-Vidal. 2025. "“End-to-End Chromosome Fusion” as the Main Driver of Descending Dysploidy in Vigna lasiocarpa (Mart. ex Benth.) Verdc. (Leguminosae Juss.)" Plants 14, no. 12: 1872. https://doi.org/10.3390/plants14121872
APA StyleSerafim, L., Silva, J. H., Dias, S., Oliveira, A. R. d. S., Nunes, M. C., da Costa, A. F., Benko-Iseppon, A. M., Jiang, J., do Vale Martins, L., & Brasileiro-Vidal, A. C. (2025). “End-to-End Chromosome Fusion” as the Main Driver of Descending Dysploidy in Vigna lasiocarpa (Mart. ex Benth.) Verdc. (Leguminosae Juss.). Plants, 14(12), 1872. https://doi.org/10.3390/plants14121872