BLH3 Regulates the ABA Pathway and Lignin Synthesis Under Salt Stress in Lilium pumilum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cloning and Bioinformatic Analysis of the LpBLH3 Gene
2.2. Analysis of LpBLH3 Expression
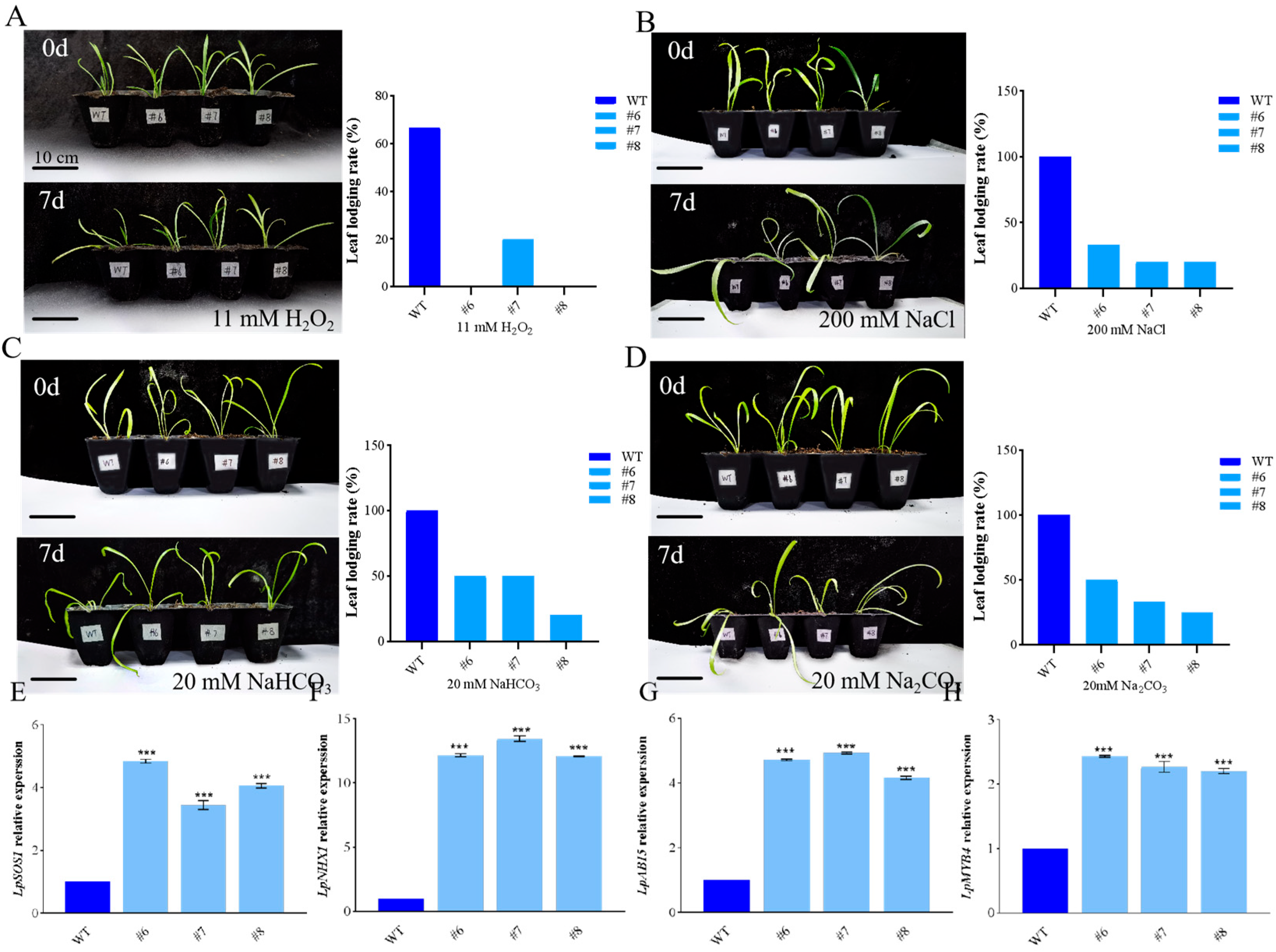
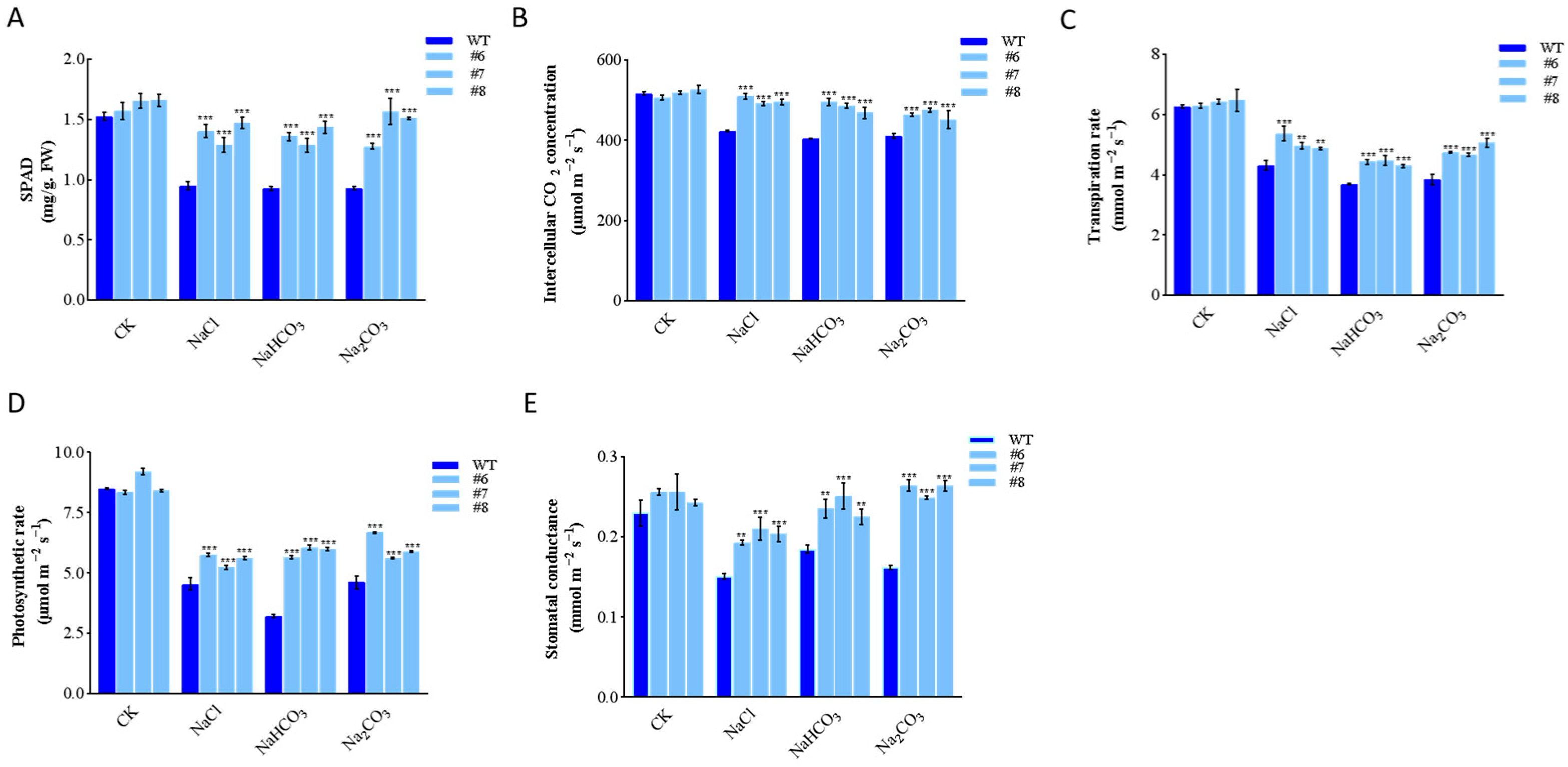
2.3. Generation of LpBLH3 Over-Expressing Lines and Measure of Physiological Indexes
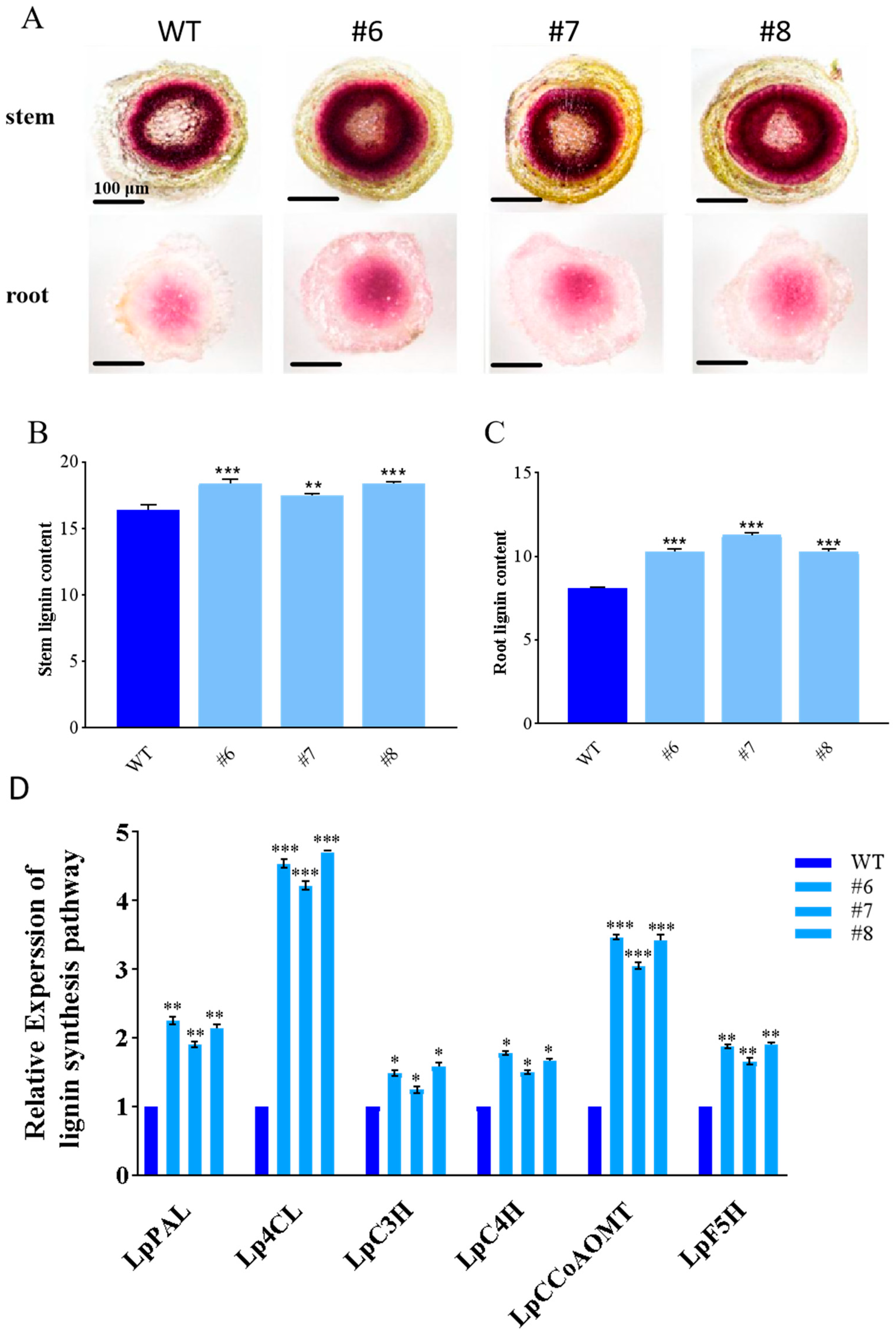
2.4. The Effect of the LpBLH3 Gene on Lignin Content
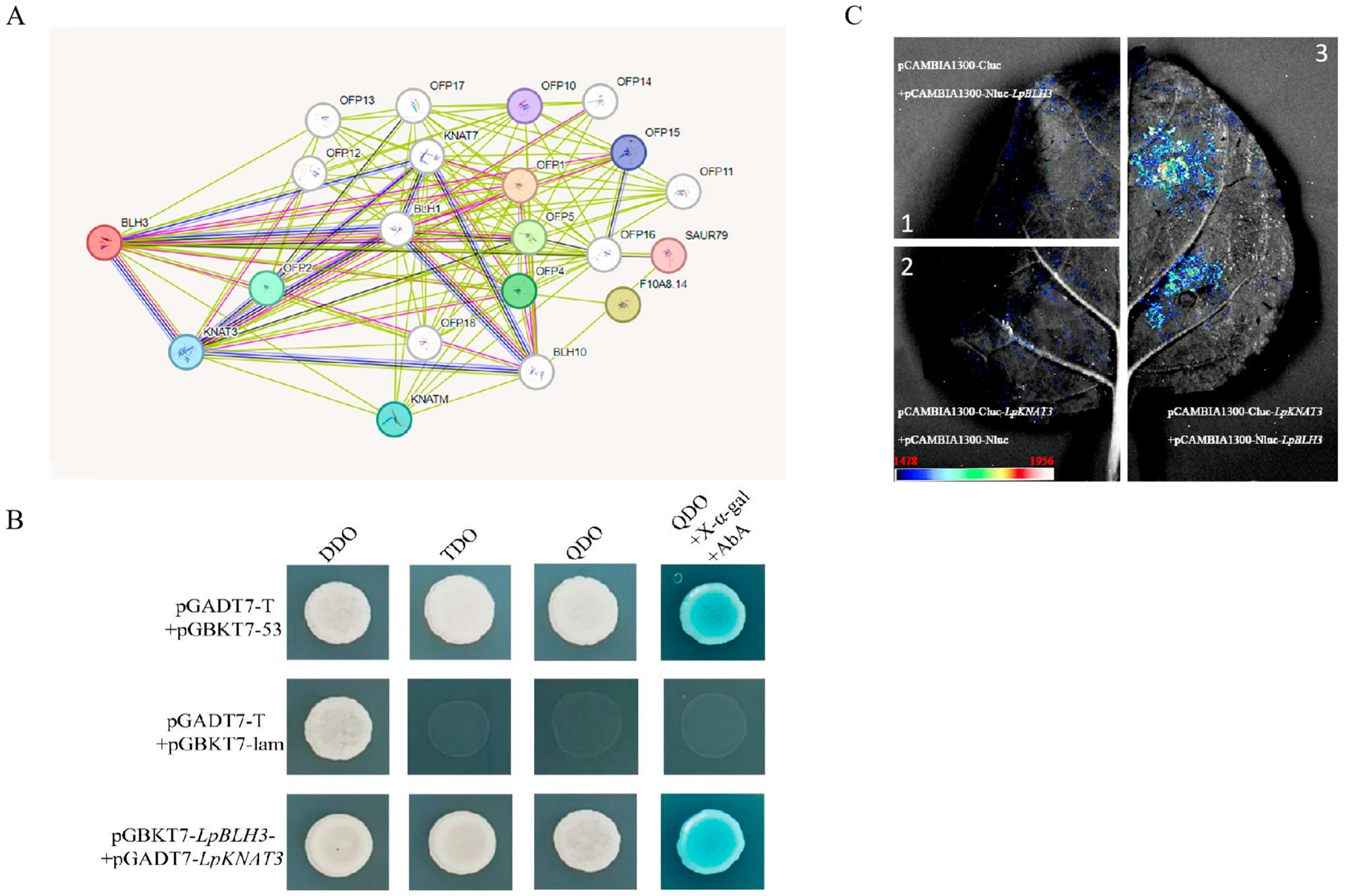
2.5. Analysis of LpBLH3-Interacting Proteins
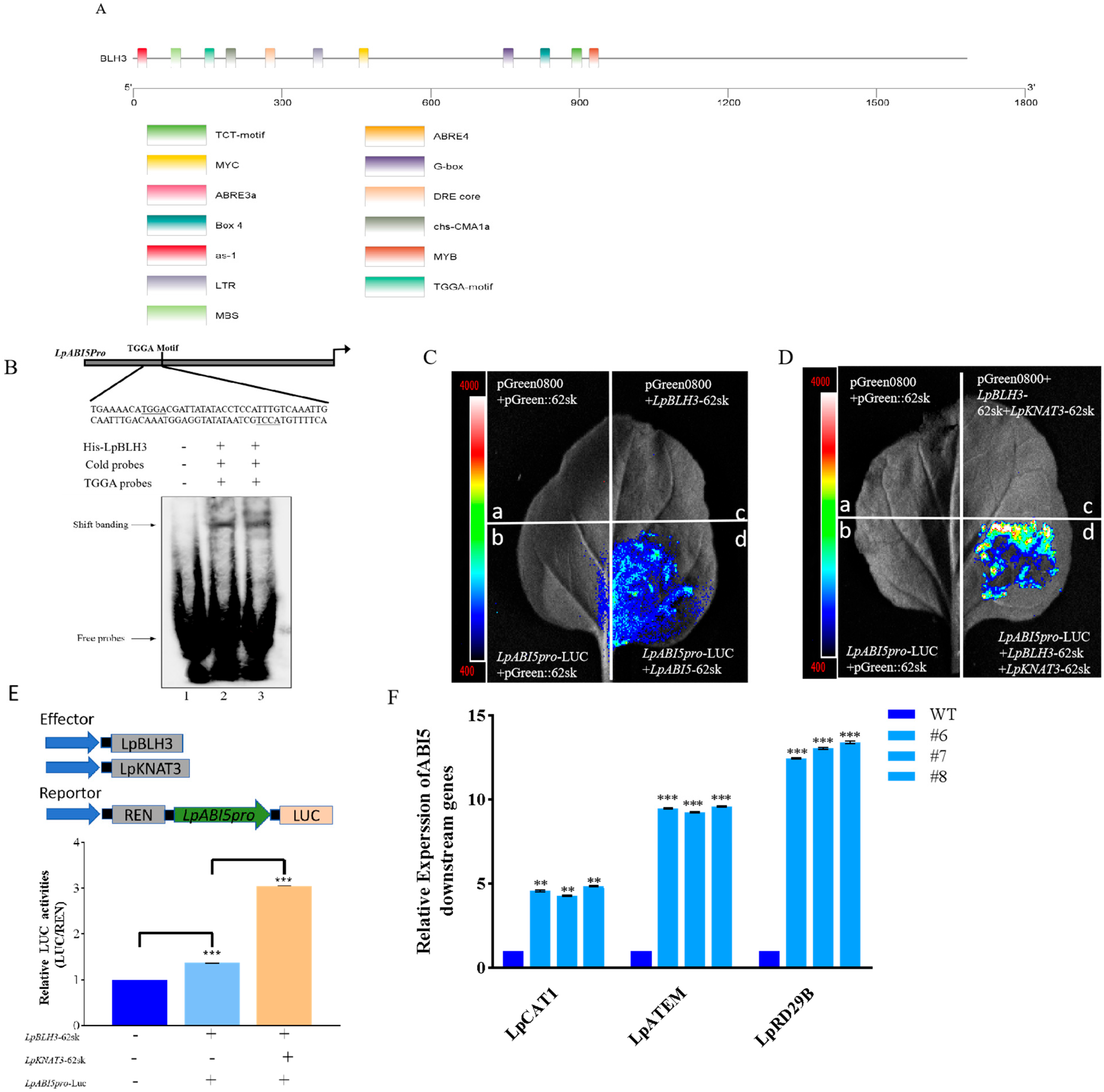
2.6. BLH3 Regulates the ABI5 Expression
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Cloning and Bio-Informatics Analysis of LpBLH3 Gene
4.3. RT-qPCR Analysis of LpBLH3 Expression
4.4. Subcellular Localization of LpBLH3
4.5. Acquisition of LpBLH3 Overexpressed Lines
4.6. Determination of Physiological Indexes of LpBLH3 OverExperssing L. pumilum
4.7. LpBLH3 Regulates the Lignin Content in L. pumilum
4.8. Screening of LpBLH3 Interacting Protein
4.9. Validation of the Interaction Between LpBLH3 and LpKNAT3
4.10. Cloning of the LpABI5 Promoter and Analysis
4.11. Dual-Luciferase Reporter Gene Assay
4.12. DNA Electrophoretic Mobility Shift Assay (EMSA)
4.13. Data Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J. How saline-alkaline land transforms into “black soil”: Placing equal emphasis on improvement and cultivation. The People’s Daily, 1 November 2023. [Google Scholar]
- Yao, D.; Wu, J.; Hu, Z.; Bai, B.; Zhuang, W.; Li, J.; Deng, Q. Physiological mechanisms and breeding strategies for saline-alkali tolerance in rice. Hybrid Rice 2019, 34, 1–7. [Google Scholar] [CrossRef]
- Isayenkov, S.; Maathuis, F. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, J.; Wang, C.; Han, K.; Hu, L.; Niu, T.; Yang, Y.; Chang, Y.; Xie, J. Exogenous Proline Enhances Systemic Defense against Salt Stress in Celery by Regulating Photosystem, Phenolic Compounds, and Antioxidant System. Plants 2023, 12, 928. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Xia, X.; Li, H.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Tan, M.; Sun, S.; Wang, J. Bioinformatics and Stress-Responsive Expression Analysis of DREB Transcription Factors in Lilium pumilum. J. Northwest For. Univ. 2023, 38, 95–101+198. [Google Scholar]
- Tao, Y.; Chen, M.; Shu, Y.; Zhu, Y.; Wang, S.; Huang, L.; Yu, X.; Wang, Z.; Qian, P.; Gu, W.; et al. Identification and functional characterization of a novel BEL1-like homeobox transcription factor GmBLH4 in soybean. Plant Cell 2018, 134, 331–344. [Google Scholar] [CrossRef]
- Hamant, O.; Pautot, V. Plant development: A TALE story. Comptes Rendus Biol. 2010, 333, 371–381. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lin, H.; Joo, S.; Goodenough, U. Early sexual origins of homeoprotein heterodimerization and evolution of the plant KNOX/BELL family. Cell 2008, 133, 829–840. [Google Scholar] [CrossRef]
- Niu, X.; Fu, D. The Roles of BLH Transcription Factors in Plant Development and Environmental Response. Int. J. Mol. Sci. 2022, 23, 3731. [Google Scholar] [CrossRef]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A Review on Plant Responses to Salt Stress and Their Mechanisms of Salt Resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Blein, T.; Hasson, A.; Laufs, P. Leaf development: What it needs to be complex. Curr. Opin. Plant Biol. 2010, 13, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kushalappa, K.; Godt, D.; Pidkowich, M.S.; Pastorelli, S.; Hepworth, S.R.; Haughn, G.W. The Arabidopsis BEL1-LIKE HOMEODOMAIN proteins SAW1 and SAW2 act redundantly to regulate KNOX expression spatially in leaf margins. Plant Cell 2007, 19, 2719–2735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, L.; Zhang, X.; Zhang, S.; Xie, D.; Liang, C.; Huang, W.; Fan, L.; Fang, Y.; Chang, Y. OFP1 Interaction with ATH1 Regulates Stem Growth, Flowering Time and Flower Basal Boundary Formation in Arabidopsis. Genes 2018, 9, 399. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.; Ju, H.; Chen, J.; Wang, S.; Wang, H.; Zhao, Y.; Chang, Y. Ovate family protein1 interaction with BLH3 regulates transition timing from vegetative to reproductive phase in Arabidopsis. Biochem. Biophys. Res. Commun. 2016, 470, 492–497. [Google Scholar] [CrossRef]
- Zhang, J. Functional Studies of GhBLH5-AO5 and GhHDT4D in Cotton Drought Stress Response. Ph.D. Thesis, Huazhong Normal University, Wuhan, China, 2021. [Google Scholar] [CrossRef]
- Chen, S.; Jia, Y.; Yang, Y.; Liu, H.; Chen, H.; Liu, J.; Yin, H.; Zhuo, R.; Han, X. Genome-wide analysis of the TsBLH gene family reveals TsBLH4 involved the regulation of abiotic stresses by interacting with KNOX6 in Toona sinensis. Plant Stress 2025, 15, 100721. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, X.; Cheng, Z.; Yao, W.; Li, R.; Jiang, T.; Zhou, B. Comprehensive analysis of the three-amino-acid-loop-extension gene family and its tissue-differential expression in response to saline stress in poplar. Plant Physiol. Biochem. 2019, 136, 1–12. [Google Scholar] [CrossRef]
- Liu, Y. Functional analysis of homeodomain transcription factors in secondary cell wall formation in Arabidopsis thaliana. Plant Cell 2015, 26, 480–485. [Google Scholar]
- Chun, H.J.; Baek, D.; Cho, H.M.; Lee, S.H.; Jin, B.J.; Yun, D.-J.; Hong, Y.-S.; Kim, M.C. Lignin biosynthesis genes play critical roles in the adaptation of Arabidopsis plants to high-salt stress. Plant Signal. Behav. 2019, 14, 1625697. [Google Scholar] [CrossRef]
- Chen, H.; Wang, J.P.; Liu, H.; Li, H.; Lin, Y.-C.J.; Shi, R.; Yang, C.; Gao, J.; Zhou, C.; Li, Q.; et al. Hierarchical transcription factor and chromatin binding network for wood formation in Populus trichocarpa. Plant Cell 2019, 31, 602–626. [Google Scholar] [CrossRef]
- Yan, C.; Hu, Z.; Nie, Z.; Li, J.; Yao, X.; Yin, H. CcBLH6, a BELl-like homeodomain-containing transcription factor, regulates the fruit lignification pattern. Planta 2021, 253, 90. [Google Scholar] [CrossRef] [PubMed]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APXB genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Yin, Q.; Chen, J.; Zhao, X.; Yue, F.; He, J.; Yang, L.; Liu, L.; Zeng, Q.; Lu, F.; et al. The class II KNOX transcription factors KNAT3 and KNAT7 synergistically regulate monolignol biosynthesis in Arabidopsis. J. Exp. Bot. 2020, 71, 5469–5483. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yamaguchi, M.; Grienenberger, E.; Martone, P.T.; Samuels, A.L.; Mansfield, S.D. The Class II KNOX genes KNAT3 and KNAT7 work cooperatively to influence deposition of secondary cell walls that provide mechanical support to arabidopsis stems. Plant J. 2020, 101, 293–309. [Google Scholar] [CrossRef]
- Hoth, S.; Morgante, M.; Sanchez, J.-P.; Hanafey, M.K.; Tingey, S.V.; Chua, N.-H. Genome-we gene expression profiling in Arabidopsis thaliana reveals new targets of abscisic acid and largely impaired gene regulation in the abi1-1 mutant. J. Cell Sci. 2002, 115, 4891–4900. [Google Scholar] [CrossRef]
- Zhang, D.; Ding, X.; Wang, Z.; Li, W.; Li, L.; Liu, L.; Zhou, H.; Yu, J.; Zheng, C.; Wu, H.; et al. A C2H2 Zinc Finger Protein, OsZOS2-19, Modulates ABA Sensitivity and Cold Response in Rice. Plant Cell Physiol. 2025, 66, 753–765. [Google Scholar] [CrossRef]
- Kim, D.; Cho, Y.; Ryu, H.; Kim, Y.; Kim, T.; Hwang, I. BLH 1 and KNAT 3 modulate ABA responses during germination and early seedling development in Arabidopsis. Plant J. 2013, 75, 755–766. [Google Scholar] [CrossRef]
- Liu, C.; Li, Z.; Dou, L.; Yuan, Y.; Zou, C.; Shang, H.; Cui, L.; Xiao, G. A genome-wide identification of the BLH gene family reveals BLH1 involved in cotton fiber development. J. Cotton Res. 2020, 3, 26. [Google Scholar] [CrossRef]
- Jia, T.; Wang, H.; Cui, S.; Li, Z.; Shen, Y.; Li, H.; Xiao, G. Cotton BLH1 and KNOX6 antagonistically modulate fiber elongation via regulation of linolenic acid biosynthesis. Plant Commun. 2024, 5, 100887. [Google Scholar] [CrossRef]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef]
- Yan, F.; Deng, W.; Wang, X.; Yang, C.; Li, Z. Maize (Zea mays L.) homologue of ABA-insensitive (ABI) 5 gene plays a negative regulatory role in abiotic stresses response. Plant Growth Regul. 2012, 68, 383–393. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-Insensitive 5) in plant development, abiotic stress responses and phytohormone crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-C.; Tsai, M.-C.; Wu, S.-S.; Chang, I.-F. Regulation of ABI5 expression by ABF3 during salt stress responses in Arabidopsis thaliana. Bot. Stud. 2019, 60, 16. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.; Ma, Y.; Wu, Z.; Yu, Y.-T.; Liang, S.; Lu, K.; Wang, X.-F. Arabidopsis ABI5 plays a role in regulating ROS homeostasis by activating CATALASE 1 transcription in seed germination. Plant Mol. Biol. 2017, 94, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Bensmihen, S.; To, A.; Lambert, G.; Kroj, T.; Giraudat, J.; Parcy, F. Analysis of an activated ABI5 allele using a new selection method for transgenic Arabidopsis seeds. FEBS 2004, 561, 127–131. [Google Scholar] [CrossRef]
- Nakashima, K.; Fujita, Y.; Katsura, K.; Maruyama, K.; Narusaka, Y.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulation of ABI3-and ABA-responsive genes including RD29B and RD29A in seeds, germinating embryos, and seedlings of Arabidopsis. Plant Mol. Biol. 2006, 60, 51–68. [Google Scholar] [CrossRef]
- He, H. Transcriptome analysis of Lilium pumilum and Preliminary Exploration of the Function of LpPEX5 and LpPEX7 Genes. Master’s Thesis, Northeast Foreastry University, Harbin, China, 2020. [Google Scholar]
- Tadjouri, H.; Amiri, O.; Medjedded, H.; Nemmiche, S.; Benati, F.Z. Ecophysiological responses of Glycine max L. under single and combined cadmium and salinity stresses. Ecotoxicology 2023, 32, 802–810. [Google Scholar] [CrossRef]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J.; Wang, Y.; Yang, Q. Melatonin Improves Drought Resistance in Maize Seedlings by Enhancing the Antioxidant System and Regulating Abscisic Acid Metabolism to Maintain Stomatal Opening Under PEG-Induced Drought. J. Plant Biol. 2022, 64, 299–312. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Huang, X.; Xing, J.; Yao, J.; Yin, T.; Jiang, J.; Wang, P.; Xu, B. STAYGREEN-mediated chlorophyll a catabolism is critical for photosystem stability during heat-induced leaf senescence in perennial ryegrass. Plant Cell Environ. 2022, 45, 1412–1427. [Google Scholar] [CrossRef]
- Chen, H.; Wu, W.; Du, K.; Ling, A.; Kang, X. The interplay of growth-regulating factor 5 and BZR1 in coregulating chlorophyll degradation in poplar. Plant Cell Environ. 2018, 47, 3766–3779. [Google Scholar] [CrossRef]
- Hu, P.; Zhang, K.; Yang, C. BpNAC012 positively regulates abiotic stress responses and secondary wall biosynthesis. Plant Physiol. 2019, 179, 700–717. [Google Scholar] [CrossRef]
- Yang, Q.; Yuan, C.; Cong, T.; Wang, J.; Zhang, Q. Genome-wide identification of three-amino-acid-loop-extension gene family and their expression profile under hormone and abiotic stress treatments during stem development of Prunus mume. Front. Plant Sci. 2022, 13, 1006360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H. The Molecular Mechanisms of Plant Responses to salt stress. Front. Plant Sci. 2020, 13, 2022. [Google Scholar] [CrossRef]
- Nookaraju, A.; Pandey, S.K.; Ahlawat, Y.K.; Joshi, C.P. Understanding the modus operandi of Class II KNOX transcription factors in secondary cell wall biosynthesis. Plants 2022, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- Collin, A.; Daszkowska-Golec, A.; Szarejko, I. Updates on the Role of ABSCISIC ACID INSENSITIVE 5 (ABI5) and ABSCISIC ACID-RESPONSIVE ELEMENT BINDING FACTORs (ABFs) in ABA Signaling in Different Developmental Stages in Plants. Cells 2021, 10, 1996. [Google Scholar] [CrossRef]
- Zhang, J.; Gai, M.; Xue, B.; Jia, N.; Wang, C.; Wang, J.; Sun, H. The use of miRNAs as reference genes for miRNA expression normalization during Lilium somatic embryogenesis by real-time reverse transcription PCR analysis. Plant Cell 2017, 129, 105–118. [Google Scholar] [CrossRef]
- Liang, S.; Liu, Y.Y.; Lin, N.F. Saline-alkaline Grassland Improvement through the Bio-engineering Technology. Adv. Mater. Res. 2013, 807, 1318–1321. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Z.; Ji, S.; Zhu, G.; Dong, Y.; Li, J.; Jing, Y.; Jin, S. Ferric reduction oxidase in Lilium pumilum affects plant saline-alkaline tolerance by regulating ROS homeostasis. Plant Physiol. Biochem. 2024, 207, 108305. [Google Scholar] [CrossRef]
- Senthilkumar, M.; Amaresan, N.; Sankaranarayanan, A. Estimation of Malondialdehyde (MDA) by Thiobarbituric Acid (TBA) Assay. In Plant-Microbe Interactions; Springer Protocols Handbooks; Springer Nature: New York, NY, USA, 2021; pp. 103–105. [Google Scholar] [CrossRef]
- Xu, L.; Zhu, L.; Tu, L.; Liu, L.; Yuan, D.; Jin, L.; Long, L.; Zhang, X. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by Rna-Seq-dependent transcriptional analysis and histochemistry. J. Exp. Bot. 2011, 62, 5607–5621. [Google Scholar] [CrossRef]
- Xie, X.-M.; Zhang, X.-Q.; Dong, Z.-X.; Guo, H.-R. Dynamic changes of lignin contents of MT-1 elephant grass and its closely related cultivars. Biomass Bioenergy 2011, 35, 1732–1738. [Google Scholar] [CrossRef]
- Ge, Q. Cloning of the Choline Monooxygenase Gene of Salix Salina and Its Expression in Tobacco. Master’s Thesis, Dalian University of Technology, Dalian, China, 2005. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, W.; Zhang, L.; Liu, X.; Cui, H.; Shi, M.; Sun, H.; Yang, W.; Wang, X.; Yang, F.; Jin, S. BLH3 Regulates the ABA Pathway and Lignin Synthesis Under Salt Stress in Lilium pumilum. Plants 2025, 14, 1860. https://doi.org/10.3390/plants14121860
Wan W, Zhang L, Liu X, Cui H, Shi M, Sun H, Yang W, Wang X, Yang F, Jin S. BLH3 Regulates the ABA Pathway and Lignin Synthesis Under Salt Stress in Lilium pumilum. Plants. 2025; 14(12):1860. https://doi.org/10.3390/plants14121860
Chicago/Turabian StyleWan, Wenhao, Lingshu Zhang, Xingyu Liu, Huitao Cui, Miaoxin Shi, Hao Sun, Wei Yang, Xinran Wang, Fengshan Yang, and Shumei Jin. 2025. "BLH3 Regulates the ABA Pathway and Lignin Synthesis Under Salt Stress in Lilium pumilum" Plants 14, no. 12: 1860. https://doi.org/10.3390/plants14121860
APA StyleWan, W., Zhang, L., Liu, X., Cui, H., Shi, M., Sun, H., Yang, W., Wang, X., Yang, F., & Jin, S. (2025). BLH3 Regulates the ABA Pathway and Lignin Synthesis Under Salt Stress in Lilium pumilum. Plants, 14(12), 1860. https://doi.org/10.3390/plants14121860