
Conditional QTL Analysis and Fine Mapping for Thousand-Kernel Weight in Common Wheat

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Phenotypic Analysis of the Wheat RIL Population
2.2. Unconditional QTL Analysis for TKW in the Wheat RIL Population
2.3. Conditional QTL Analysis for TKW in KJ-RIL Population
2.4. Epistatic QTL Analysis for TKW in the KJ-RIL Population
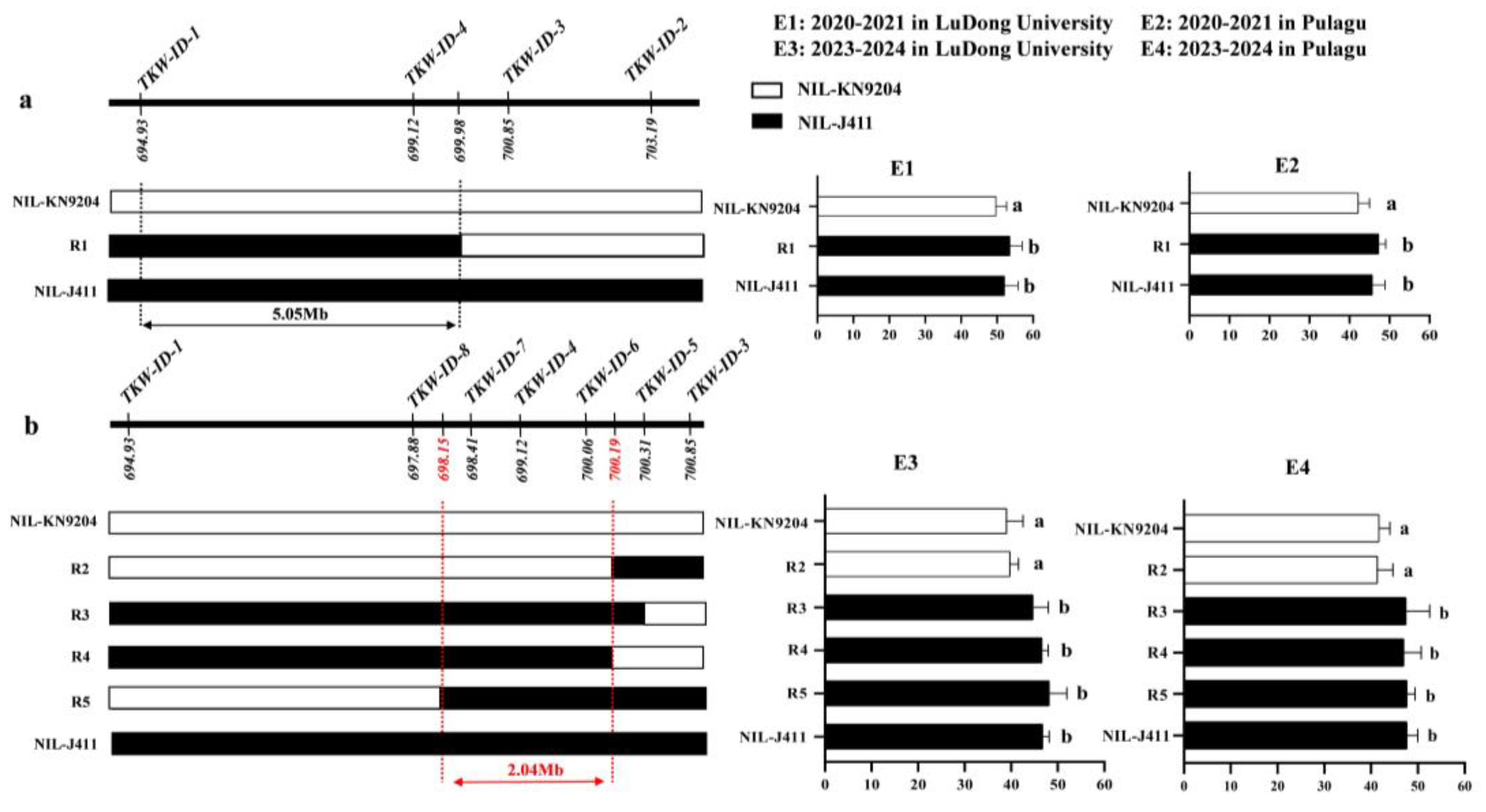
2.5. Fine Mapping of QTkw1B in the NIL Population
2.6. Genetic Effect Analysis of QTkw1B on Kernel and Yield-Related Traits
3. Discussion and Conclusions
4. Materials and Methods
4.1. Plant Materials and Field Trials
4.2. Experimental Design and Phenotypic Evaluation
4.3. Genetic Map Construction and QTL Detection
4.4. Marker Development and Genotype Identification
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| CV | Coefficient of Variation |
| KU | Kurtosis |
| KDR | Kernel Diameter Ratio |
| KL | Kernel Length |
| KW | Kernel Width |
| KYPP | Kernel Yield Per Plant |
| KNPS | Kernel Number Per Spike |
| PH | Plant Height |
| QTL | Quantitative Trait Loci |
| RIL | Recombinant Inbred Line |
| SD | Standard Deviation |
| SK | Skewness |
| SNPS | Spikelet Number Per Spike |
| SN | Spike Number |
| SL | Spike Length |
| TKW | Thousand-Kernel Weight |
References
- Acreche, M.M.; Slafer, G.A. Grain weight response to increases in number of grains in wheat in a Mediterranean area. Field Crops Res. 2005, 98, 52–59. [Google Scholar] [CrossRef]
- Walker, C.K.; Ford, R.; Muñoz-Amatriaín, M.; Panozzo, J.F. The detection of QTLs in barley associated with endosperm hardness, grain density, grain size and malting quality using rapid phenotyping tools. Theor. Appl. Genet. 2013, 126, 2533–2551. [Google Scholar] [CrossRef]
- Guan, P.; Di, N.; Mu, Q.; Shen, X.; Wang, Y.; Wang, X.; Yu, K.; Song, W.; Chen, Y.; Xin, M.; et al. Use of near-isogenic lines to precisely map and validate a major QTL for grain weight on chromosome 4AL in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2019, 132, 2367–2379. [Google Scholar] [CrossRef]
- Brinton, J.; Simmonds, J.; Minter, F.; Leverington-Waite, M.; Snape, J.; Uauy, C. Increased pericarp cell length underlies a major quantitative trait locus for grain weight in hexaploid wheat. New Phytol. 2017, 215, 1026–1038. [Google Scholar] [CrossRef]
- Zhao, J.; Becker, H.C.; Zhang, D.; Zhang, Y.; Ecke, W. Conditional QTL mapping of oil content in rapeseed with respect to protein content and traits related to plant development and grain yield. Theor. Appl. Genet. 2006, 113, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Dholakia, B.B.; Ammiraju, J.S.S.; Singh, H.; Lagu, M.D.; Der, M.S.R.; Rao, V.S.; Dhaliwal, H.S.; Ranjekar, P.K.; Gupta, V.S.; Weber, W.E. Molecular marker analysis of kernel size and shape in bread wheat. Plant Breed. 2003, 122, 392–395. [Google Scholar] [CrossRef]
- Peng, J.; Ronin, Y.; Fahima, T.; Roder, M.S.; Li, Y.; Nevo, E.; Korol, A. Domestication quantitative trait loci in Triticum dicoccoides, the progenitor of wheat. Proc. Natl. Acad. Sci. USA 2003, 100, 2489–2494. [Google Scholar] [CrossRef] [PubMed]
- Ramya, P.; Chaubal, A.; Kulkarni, K.; Gupta, L.; Kadoo, N.; Dhaliwal, H.S.; Chhuneja, P.; Lagu, M.; Gupta, V. QTL mapping of 1000-kernel weight, kernel length, and kernel width in bread wheat (Triticum aestivum L.). J. Appl. Genet. 2010, 51, 421–429. [Google Scholar] [CrossRef]
- Sun, X.; Wu, K.; Zhao, Y.; Kong, F.; Han, G.; Jiang, H.; Huang, X.; Li, R.; Wang, H.; Li, S. QTL analysis of kernel shape and weight using recombinant inbred lines in wheat. Euphytica 2009, 165, 615–624. [Google Scholar] [CrossRef]
- Wen, Y.X.; Zhu, J. Multivariable conditional analysis for complex trait and its components. Acta. Genet. Sin. 2005, 32, 289–296. [Google Scholar]
- Cui, F.; Li, J.; Ding, A.M.; Zhao, C.H.; Wang, L.; Wang, X.Q.; Li, S.S.; Bao, Y.G.; Li, X.F.; Feng, D.S.; et al. Conditional QTL mapping for plant height with respect to the length of the spike and internode in two mapping populations of wheat. Theor. Appl. Genet. 2011, 122, 1517–1536. [Google Scholar] [CrossRef]
- Fan, X.L.; Liu, X.F.; Guo, S.D.; Feng, B.; Zhou, Q.; Deng, G.B.; Long, H.; Xu, Z.B.; Wang, T. Traditional and Conditional QTL Analysis of Kernel Size- and Shape-Related Traits in Wheat (Triticum aestivum L.). Agriculture 2022, 12, 1718. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Deng, Z.Y.; Wang, Y.R.; Li, J.F.; Tian, J.C. Unconditional and conditional QTL analysis of kernel weight related traits in wheat (Triticum aestivum L.) in multiple genetic backgrounds. Genetica 2014, 142, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Dong, Y.B.; Cui, D.Q.; Wang, Y.Z.; Liu, Y.Y.; Wei, M.G.; Li, X.H. The genetic relationship between popping expansion volume and two yield components in popcorn using unconditional and conditional QTL analysis. Euphytica 2008, 162, 345–351. [Google Scholar] [CrossRef]
- Guo, X.; Wu, C.N.; Wang, D.H.; Wang, G.Y.; Jin, K.T.; Zhao, Y.J.; Tian, J.C.; Deng, Z.Y. Conditional QTL mapping for seed germination and seedling traits under salt stress and candidate gene prediction in wheat. Sci. Rep. 2022, 12, 21010. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cui, F.; Wang, J.P.; Jun, L.; Ding, A.M.; Zhao, C.H.; Li, X.F.; Feng, D.S.; Gao, J.R.; Wang, H.G. Conditional QTL mapping of protein content in wheat with respect to grain yield and its components. J. Genet. 2012, 91, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Wang, J.; Liu, Q.; Zhang, M.; Zou, K.; Fu, X. Genetic Relationships Among Panicle Characteristics of Rice (Oryza sativa L.) Using Unconditional and Conditional QTL Analyses. J. Plant Biol. 2009, 52, 259–267. [Google Scholar] [CrossRef]
- Li, Q.F.; Zhang, Y.; Liu, T.T.; Wang, F.F.; Liu, K.; Chen, J.S.; Tian, J.C. Genetic analysis of kernel weight and kernel size in wheat (Triticum aestivum L.) using unconditional and conditional QTL mapping. Mol. Breed. 2015, 35, 10. [Google Scholar] [CrossRef]
- Ma, W.; Appels, R.; Bekes, F.; Larroque, O.; Morell, M.K.; Gale, K.R. Genetic characterisation of dough rheological properties in a wheat doubled haploid population: Additive genetic effects and epistatic interactions. Theor. Appl. Genet. 2005, 111, 410–422. [Google Scholar] [CrossRef]
- Sun, N.; Liu, W.; Shi, D.; Zhao, C.; Ou, J.; Song, Y.; Yang, Z.; Sun, H.; Wu, Y.; Qin, R.; et al. Mapping QTLs with additive and epistatic effects for awn length and their effects on kernel-related traits in common wheat. Front. Plant Sci. 2024, 15, 1417588. [Google Scholar] [CrossRef]
- Parveen, M.; Jitendra, K.; Shiveta, S.; Kumar, M.P.; Singh, B.H.; Kumar, G.P.; Shailendra, S. GWAS for main effects and epistatic interactions for grain morphology traits in wheat. Physiol. Mol. Biol. Plants. 2022, 28, 651–668. [Google Scholar]
- Huang, X.Q.; Kempf, H.; Ganal, M.W.; Röder, M.S. Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Borner, A.; Schumann, E.; Furste, A.; Coster, H.; Leithold, B.; Roder, S.; Weber, E. Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat ( Triticum aestivum L.). Theor. Appl. Genet. 2002, 105, 921–936. [Google Scholar] [CrossRef] [PubMed]
- Groos, C.; Robert, N.; Bervas, E.; Charmet, G. Genetic analysis of grain protein-content, grain yield and thousand-kernel weight in bread wheat. Theor. Appl. Genet. 2003, 106, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Marza, F.; Bai, G.H.; Carver, B.F.; Zhou, W.C. Quantitative trait loci for yield and related traits in the wheat population Ning7840 x Clark. Theor. Appl. Genet. 2006, 112, 688–698. [Google Scholar] [CrossRef]
- McIntyre, C.L.; Mathews, K.L.; Rattey, A.; Chapman, S.C.; Drenth, J.; Ghaderi, M.; Reynolds, M.; Shorter, R. Molecular detection of genomic regions associated with grain yield and yield-related components in an elite bread wheat cross evaluated under irrigated and rainfed conditions. Theor. Appl. Genet. 2010, 120, 527–541. [Google Scholar] [CrossRef]
- Meng, D.; Batool, A.; Xuan, Y.; Pan, R.; Zhang, N.; Zhang, W.; Zhi, L.; Ren, X.; Li, W.; Li, J.; et al. Fine mapping and validation of a stable QTL for thousand-kernel weight in wheat(Triticum aestivum L.). Crop J. 2023, 11, 1491–1500. [Google Scholar] [CrossRef]
- Wang, R.X.; Hai, L.; Zhang, X.Y.; You, G.X.; Yan, C.S.; Xiao, S.H. QTL mapping for grain filling rate and yield-related traits in RILs of the Chinese winter wheat population Heshangmai x Yu8679. Theor. Appl. Genet. 2009, 118, 313–325. [Google Scholar] [CrossRef]
- Gegas, V.C.; Nazari, A.; Griffiths, S.; Simmonds, J.; Fish, L.; Orford, S.; Sayers, L.; Doonan, J.H.; Snape, J.W. A Genetic Framework for Grain Size and Shape Variation in Wheat. Plant. Cell 2010, 22, 1046–1056. [Google Scholar] [CrossRef]
- Chang, C.; Zhang, H.P.; Zhang, X.Y.; Yan, C.S.; Xiao, S.H. Study on relationship between allelic variation in PEBP-like gene and grain size and weight in common wheat. Mol. Plant Breed. 2009, 7, 23–37. [Google Scholar]
- Ma, X.Q.; Tang, J.H.; Teng, W.T.; Yan, J.B.; Meng, Y.J.; Li, J.S. Epistatic interaction is an important genetic basis of grain yield and its components in maize. Mol. Breed. 2007, 20, 41–51. [Google Scholar] [CrossRef]
- Zhang, K.; Tian, J.; Zhao, L.; Wang, S. Mapping QTL with epistatic effects and QTL × environment interactions for plant height using a doubled haploid population in cultivated wheat. J. Genet. Genomics 2008, 35, 119–127. [Google Scholar] [CrossRef]
- Cao, G.Q.; Zhu, J.; He, C.X.; Gao, Y.M.; Wu, P. QTL analysis for epistatic effects and QTL x environment interaction effects on final height of rice (Oryza sativa L.). Yi Chuan Xue Bao 2001, 28, 135–143. [Google Scholar] [PubMed]
- Song, J.; Xu, D.; Dong, Y.; Li, F.; Bian, Y.; Li, L.; Luo, X.; Fei, S.; Li, L.; Zhao, C.; et al. Fine mapping and characterization of a major QTL for grain weight on wheat chromosome arm 5DL. Theor. Appl. Genet. 2022, 135, 3237–3246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yang, L.; Liu, D.; Zeng, J.; Cao, S.; Xia, X.; Yan, J.; Song, X.; He, Z.; Zhang, Y. Fine mapping and validation of a major QTL for grain weight on chromosome 5B in bread wheat. Theor. Appl. Genet. 2021, 134, 1–11. [Google Scholar] [CrossRef]
- Mahdi Nezhad, N.; Jalal Kamali, M.R.; McIntyre, C.L.; Fakheri, B.A.; Omidi, M.; Masoudi, B. Mapping QTL with main and epistatic effect on Seri M82 × Babax"wheat population under salt stress. Euphytica 2019, 215, 130. [Google Scholar] [CrossRef]
- Cao, J.; Shang, Y.; Xu, D.; Xu, K.; Cheng, X.; Pan, X.; Liu, X.; Liu, M.; Gao, C.; Yan, S.; et al. Identification and Validation of New Stable QTL for Grain Weight and Size by Multiple Mapping Models in Common Wheat. Front. Genetics 2020, 11, 584859. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Xu, Y.; Ma, F.; Zhang, J.; Cao, Y.; Li, L.; An, D. Identification and validation of quantitative trait loci for kernel traits in common wheat (Triticum aestivum L.). BMC Plant Biol. 2020, 20, 529. [Google Scholar] [CrossRef]
- Qin, R.; Cao, M.; Dong, J.; Chen, L.; Guo, H.; Guo, Q.; Cai, Y.; Han, L.; Huang, Z.; Xu, N.; et al. Fine mapping of a major QTL, qKl-1BL controlling kernel length in common wheat. Theor. Appl. Genet. 2024, 137, 67. [Google Scholar] [CrossRef]
- Cui, F.; Fan, X.; Chen, M.; Zhang, N.; Zhao, C.; Zhang, W.; Han, J.; Ji, J.; Zhao, X.; Yang, L.; et al. QTL detection for wheat kernel size and quality and the responses of these traits to low nitrogen stress. Theor. Appl. Genet. 2016, 129, 469–484. [Google Scholar] [CrossRef]
- Cui, F.; Fan, X.; Zhao, C.; Zhang, W.; Chen, M.; Ji, J.; Li, J. A novel genetic map of wheat: Utility for mapping QTL for yield under different nitrogen treatments. BMC Genet. 2014, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Zhang, N.; Fan, X.; Zhang, W.; Zhao, C.; Yang, L.; Pan, R.; Chen, M.; Han, J.; Zhao, X.; et al. Utilization of a Wheat 660K SNP array-derived high-density genetic map for high-resolution mapping of a major QTL for kernel number. Sci. Rep. 2017, 7, 3788. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Cui, F.; Han, X.; He, Y.; Zhao, L.; Zhang, N.; Zhang, H.; Zhu, H.; Liu, Z.; Ma, B.; et al. Comparative genomic and transcriptomic analyses uncover the molecular basis of high nitrogen-use efficiency in the wheat cultivar Kenong 9204. Mol. Plant. 2022, 15, 1440–1456. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Maaty, A.N.; Oraby, A.H. Extraction of high-quality genomic DNA from different plant orders applying a modified CTAB-based method. Bull. Natl. Res. Cent. 2019, 43, 25. [Google Scholar] [CrossRef]
- Huang, L.; Deng, X.; Li, R.; Xia, Y.; Bai, G.; Siddique, K.H.; Guo, P. A Fast Silver Staining Protocol Enabling Simple and Efficient Detection of SSR Markers using a Non-denaturing Polyacrylamide Gel. J. Vis. Exp. JoVE 2018, 134, 57192. [Google Scholar]
- Fan, X.; Cui, F.; Zhao, C.; Zhang, W.; Yang, L.; Zhao, X.; Han, J.; Su, Q.; Ji, J.; Zhao, Z.; et al. QTLs for flag leaf size and their influence on yield-related traits in wheat (Triticum aestivum L.). Mol. Breed. 2015, 35, 24. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Env | Parents | RIL Population | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| KN9204 | J411 | Min | Max | Mean | SD | SK | KU | CV (%) | ||
| TKW (g) | E1 | 43.41 ± 0.05 | 45.18 ± 1.38 | 36.38 | 52.94 | 45.92 | 3.43 | −0.35 | −0.07 | 7.46 |
| E2 | 45.31 ± 1.56 | 40.94 ± 0.02 | 34.50 | 54.55 | 45.87 | 3.46 | −0.25 | 0.39 | 7.55 | |
| E3 | 40.17 ± 1.90 | 42.46 ± 4.60 | 33.03 | 51.84 | 41.89 | 4.10 | 0.12 | −0.46 | 9.79 | |
| E4 | 38.71 ± 1.20 | 41.83 ± 1.72 ** | 30.14 | 53.56 | 41.09 | 4.78 | 0.33 | −0.26 | 11.63 | |
| E5 | 39.02 ± 4.23 | 39.89 ± 1.36 | 30.52 | 50.44 | 40.99 | 4.30 | 0.08 | −0.29 | 10.49 | |
| E6 | 34.85 ± 2.98 | 34.65 ± 3.50 | 27.09 | 49.82 | 37.34 | 5.13 | 0.20 | −0.61 | 13.75 | |
| E7 | 47.02 ± 0.74 | 47.5 ± 1.41 | 35.16 | 55.95 | 46.19 | 3.94 | −0.32 | −0.20 | 8.53 | |
| E8 | 46.33 ± 2.25 | 46.83 ± 2.64 | 30.04 | 60.91 | 46.98 | 4.65 | −0.49 | 2.03 | 9.90 | |
| E9 | - | - | 36.36 | 57.86 | 46.39 | 3.92 | 0.05 | −0.17 | 8.45 | |
| E10 | - | - | 35.43 | 59.28 | 47.99 | 5.10 | −0.37 | −0.14 | 10.62 | |
| Unconditional QTL | Environment | Chromosome | Position (Mb) | Marker Interval | LOD | PVE(%) | Add |
|---|---|---|---|---|---|---|---|
| QTkw1A.1 | E7/BLUE-LN/BLUE-HN | 1A | 35.13–51.13 | AX-109492724-AX-110471342 | 4.94/3.55/2.88 | 8.52/4.02/3.34 (5.29) | 1.34/0.64/0.70 |
| QTkw1A.2 | E10 | 1A | 90.13 | AX-111003384-AX-111659850 | 4.05 | 3.13 | 1.04 |
| QTkw1A.4 | E4/E6 | 1A | 526.13 | AX-109400055-AX-110567456 | 3.48/4.46 | 4.76/5.07 (4.91) | 1.16/1.33 |
| QTkw1B | BLUE-LN/E1/E7/E10/BLUE-HN | 1B | 693.16–700.16 | AX-109307953-AX-109435037 | 2.72/5.51/2.79/7.51/3.96 | 2.73/9.50/4.80/5.74/4.96 (5.55) | −0.52/−1.06/−1.00/−1.41/−0.85 |
| QTkw1D.3 | E5 | 1D | 477.13 | AX-95097609-AX-110376491 | 3.04 | 3.43 | −0.92 |
| QTkw2A.1 | E2/BLUE-HN/E5/E10 | 2A | 21.93–55.93 | AX-95163150-AX-95633238 | 2.52/3.98/4.04/3.51 | 4.64/ 5.28/4.08/2.52 (4.13) | −0.97/−0.88/−1.00/−0.93 |
| QTkw2A.4 | E3/E10/BLUE-LN | 2A | 641.93–662.93 | AX-111193777-AX-111217859 | 5.99/3.76/6.42 | 9.26/2.66/6.95 (6.29) | −1.41/−0.96/−0.84 |
| QTkw2A.5 | E10/BLUE-LN | 2A | 754.93–755.93 | AX-111717528-AX-111218685 | 7.30/2.79 | 5.56/2.66 (4.11) | 1.39/0.52 |
| QTkw2D.3 | E3/E2/E1 | 2D | 548.75–565.75 | AX-109945455-AX-108737061 | 8.18/3.86/4.81 | 12.63/5.44/8.32 (8.80) | 1.65/1.05/0.99 |
| QTkw2D.4 | E5/E9/BLUE-LN/E4/E6/BLUE-HN | 2D | 618.75 | AX-110548845-AX-110399362 | 6.64/4.74/10.26/9.46/6.12/7.20 | 6.40/9.06/11.48/13.47/6.91/8.91 (9.37) | 1.26/1.33/1.08/1.96/1.56/1.14 |
| QTkw3A.5 | E2 | 3A | 726.76 | AX-110026070-AX-10 9307603 | 2.83 | 4.46 | −0.95 |
| QTkw3D.2 | E5/E10/BLUE-LN | 3D | 522.08–555.08 | AX-94785859-AX-108969398 | 3.80/4.44/4.06 | 3.52/3.22/4.25 (3.66) | 0.93/1.05/0.66 |
| QTkw4A.1 | E4 | 4A | 2.91 | AX-109011322-AX-109270124 | 2.64 | 3.27 | −0.97 |
| QTkw4A.4 | E2/BLUE-HN | 4A | 567.91–568.91 | AX-111591351-AX-94881516 | 6.86/3.10 | 11.23/3.68 (7.46) | 1.50/0.73 |
| QTkw4A.6 | E1/E4/BLUE-LN/E3/E6/E5 | 4A | 618.91–644.91 | AX-111693688-AX-111148488 | 3.08/5.93/7.89/4.27/3.48/9.79 | 5.22/8.20/8.75/6.32/3.76/9.76 (7.00) | 0.79/1.53/0.94/1.17/1.15/1.55 |
| QTkw4A.8 | E1/E10 | 4A | 682.91–684.91 | AX-89599752-AX-110614619 | 4.01/4.48 | 6.81/3.25 (5.03) | 0.90/1.06 |
| QTkw4B.1 | E4/BLUE-HN/E10 | 4B | 23.70–30.70 | AX-111148234-AX-109284839 | 3.42/14.79/24.23 | 4.51/20.43/22.43 (15.79) | −1.14/−1.73/−2.79 |
| QTkw4B.2 | E5 | 4B | 66.70 | AX-109909104-AX-110066172 | 14.37 | 15.24 | −1.94 |
| QTkw4B.3 | E6/E2 | 4B | 173.70–178.70 | AX-111780236-AX-110022862 | 16.95/3.68 | 21.82/5.16 (13.49) | −2.78/−1.02 |
| QTkw4B.4 | BLUE-LN | 4B | 423.70 | AX-110574141-AX-111683846 | 5.23 | 5.43 | −0.74 |
| QTkw4B.6 | E4 | 4B | 591.70 | AX-108911622-AX-108731280 | 3.90 | 5.12 | −1.21 |
| QTkw4D.1 | E5 | 4D | 16.53 | AX-109343336-AX-111616151 | 2.84 | 3.04 | 0.86 |
| QTkw4D.2 | BLUE-HN/E6 | 4D | 52.53–67.53 | AX-108874010-AX-111072884 | 2.89/3.99 | 3.34/4.34 (3.84) | 0.70/1.23 |
| QTkw4D.3 | E10 | 4D | 236.53 | AX-110710664-AX-110025501 | 6.13 | 4.54 | 1.25 |
| QTkw5A.4 | E5/E6 | 5A | 551.67–554.67 | AX-111479128-AX-110692951 | 2.80/3.59 | 2.57/3.92 (3.25) | −0.80/−1.17 |
| QTkw5A.5 | E10 | 5A | 602.67 | AX-109524552-AX-109031208 | 13.20 | 10.59 | −1.91 |
| QTkw5A.7 | E10 | 5A | 669.67 | AX-111020300-AX-110038009 | 3.35 | 2.39 | 0.92 |
| QTkw5D.1 | E10/E6 | 5D | 0.61–2.62 | AX-94938800-AX-111475778 | 2.77/2.71 | 1.94/3.02 (2.48) | 0.83/1.04 |
| QTkw5D.3 | BLUE-LN | 5D | 431.62 | AX-110391491-AX-109732378 | 2.88 | 2.90 | −0.54 |
| QTkw5D.5 | E7 | 5D | 513.62 | AX-108810919-AX-110023145 | 3.70 | 6.24 | 1.14 |
| QTkw6A.2 | E5 | 6A | 368.01 | AX-109086661-AX-94716600 | 6.52 | 6.24 | −1.27 |
| QTkw6A.3 | E9/BLUE-LN | 6A | 413.01 | AX-111050302-AX-108817049 | 3.33/5.82 | 6.21/6.09 (6.15) | −1.13/−0.81 |
| QTkw6B.1 | BLUE-HN/BLUE-LN | 6B | 271.76–279.76 | AX-109901945-AX-109871569 | 4.41/7.43 | 5.22/7.96 (6.59) | −0.87/−0.89 |
| QTkw6B.2 | E4/E3/E6 | 6B | 453.76–457.76 | AX-109968521-AX-108962284 | 6.50/6.68/4.88 | 8.90/10.12/5.38 (8.13) | −1.59/−1.48/−1.37 |
| QTkw6B.3 | E1/E9 | 6B | 514.76–515.76 | AX-110149880-AX-110014652 | 4.19/3.80 | 7.03/7.11 (7.07) | −0.91/−1.17 |
| QTkw6B.4 | E5 | 6B | 648.76 | AX-111797130-AX-109373555 | 5.21 | 4.90 | −1.10 |
| QTkw7A.2 | E8 | 7A | 677.95 | AX-109949333-AX-111829116 | 3.76 | 9.23 | 1.43 |
| QTL | Marker Interval | Position (Mb) | Unconditional QTL | Conditional QTL | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TKW|KL | TKW|KW | TKW|KDR | ||||||||
| Environment | PVE (%) | Environment | PVE (%) | Environment | PVE (%) | Environment | PVE (%) | |||
| QTkw1A.1 | AX-109310335-AX-111672231 | 22.13–45.50 | E7/BLUE-LN/BLUE-HN | 8.52/4.02/3.34 (5.29) | E10/E7/E6/BLUE-LN | 3.61/8.56/3.95/2.96 a (4.77) | ||||
| QTkw1A.3 | AX-110717317-AX-109083423 | 184.13–185.13 | E10/BLUE-HN | 4.16/13.91 (9.03) d | ||||||
| QTkw1A.4 | AX-109400055-AX-110567456 | 524.66–526.52 | E4/E6 | 4.75/5.07 (4.91) c | ||||||
| QTkw1B | AX-109307953-AX-111625435 | 693.16–706.16 | BLUE-LN/E1/E7/E10/BLUE-HN | 2.73/9.50/4.80/5.74/4.96 (5.55) | E6/E9/BLUE-LN/E4/E10/BLUE-HN | 4.53/8.41/15.43/10.27/12.14/8.63 b (9.90) | E2/E6/E1/BLUE-LN/E9/E7/E4/E8/E10/BLUE-HN | 5.23/5.61/12.31/8.42/5.95/6.53/8.02/5.61/6.65/4.97 b (6.93) | ||
| QTkw1D.3 | AX-89693771-AX-110671322 | 453.13–487.13 | E5 | 3.43 | E10/E6/E3/BLUE-LN/E1 | 3.49/3.68/5.54/2.94/4.60 a (4.05) | ||||
| QTkw2A.1 | AX-95163150-AX-95633238 | 21.93–55.93 | E2/BLUE-HN/E5/E10 | 2.52/3.98/4.04/3.51 c (3.51) | ||||||
| QTkw2A.2 | AX-111605603-AX-111079268 | 65.93–66.93 | BLUE-HN/E10 | 4.53/5.36 (4.95) d | ||||||
| QTkw2A.3 | AX-86179693-AX-94718925 | 616.55–619.68 | E3/BLUE-LN | 18.31/6.63 (12.47) d | ||||||
| QTkw2A.4 | AX-111193777-AX-111217859 | 641.93–662.93 | E3/E10/BLUE-LN | 9.26/2.66/6.95 (6.29) | E3/E1/E9/E10/BLUE-HN/BLUE-LN | 13.31/6.31/5.66/8.36/4.57/11.13 b (8.22) | ||||
| QTkw2A.5 | AX-111717528-AX-111218685 | 752.58–756.56 | E10/BLUE-LN | 5.56/2.66(4.11) | BLUE-LN/E10 | 3.20/3.64 a (3.42) | ||||
| QTkw2B | AX-108831932-AX-110141316 | 54.55–55.57 | E6/BLUE-HN | 4.82/4.13(4.48) d | ||||||
| QTkw2D.3 | AX-111722527-AX-111497459 | 529.75–550.75 | E3/E2/E1 | 12.63/5.44/8.32 (8.80) | E1/E9/E10/E5 | 8.18/9.95/3.25/3.56 b (6.24) | E1/E10/BLUE-LN/E9 | 7.58/2.50/7.31/6.69 b (6.02) | ||
| QTkw2D.4 | AX-110548845-AX-110399362 | 618.5–621.17 | E5/E9/BLUE-LN/E4/E6/BLUE-HN | 6.40/9.06/11.48/13.47/6.91/8.91 (9.37) | E3/E7/BLUE-LN/E4/E6/E8/BLUE-HN | 10.24/6.04/12.39/12.11/9.53/9.38/12.3 a (10.30) | E5/E4/E6/BLUE-LN | 6.37/8.51/4.33/7.70 b (6.73) | ||
| QTkw3D.2 | AX-94785859-AX-108969398 | 522.08–555.08 | E5/E10/BLUE-LN | 3.52/3.22/4.25 (3.66) | E5/BLUE-LN/E6 | 9.05/5.28/5.26 b (6.53) | E5/E3/BLUE-LN/E6 | 5.19/3.21/4.90/5.20 a (4.63) | ||
| QTkw4A.4 | AX-111591351-AX-94881516 | 563.91–589.91 | E2/BLUE-HN | 11.23/3.68 (7.46) | E2/E5/E5 | 11.58/7.91/5.88 a (8.46) | ||||
| QTkw4A.6 | AX-111201251-AX-111711476 | 624.58–657.91 | E1/E4/BLUE-LN/E3/E6/E5 | 5.22/8.20/8.75/6.32/3.76/9.76 (7.00) | E3/E5/E4/BLUE-LN/E1 | 6.75/5.09/6.92/6.28/5.22 a (6.05) | E4/BLUE-LN/E3/E6/BLUE-HN | 6.13/7.02/6.58/4.21/7.06 a (6.2) | ||
| QTkw4A.8 | AX-89599752-AX-110614619 | 682.79–686.14 | E1/E10 | 6.81/3.25 (5.03) c | ||||||
| QTkw4B.1 | AX-109357924-AX-108914451 | 15.7–35.70 | E4/BLUE-HN/E10 | 4.51/20.43/22.43 (15.79) | E4/E6/BLUE-HN/E10 | 6.29/14.00/23.20/33.55 b (19.26) | E4/BLUE-HN/E10 | 5.43/4.69/11.19 b (7.1) | E4/BLUE-HN/E10/E6 | 5.11/14.69/16.98/8.04 b (11.21) |
| QTkw4B.2 | AX-109909104-AX-109315850 | 65.70–76.70 | E5 | 15.24 | E2/E5 | 6.93/17.68 b (12.31) | ||||
| QTkw4B.3 | AX-111593917-AX-110022862 | 144.70–178.70 | E6/E2 | 21.82/5.16 (13.49) | E6/E4 | 8.86/7.90 b (8.38) | ||||
| QTkw4B.4 | AX-110998404-AX-109473770 | 402.70–438.70 | BLUE-LN | 5.43 | BLUE-LN/E1 | 9.25/10.62 b (9.94) | E6/E1 | 20.21/10.62 b (15.42) | E6/BLUE-LN | 8.55/5.83 b (7.19) |
| QTkw4D.2 | AX-108874010-AX-111072884 | 52.53–67.53 | BLUE-HN/E6 | 3.35/4.34 (3.85) | E8/E6 | 4.84/6.58 b (5.71) | ||||
| QTkw5A.4 | AX-111479128-AX-111562392 | 551.56–557.79 | E5/E6 | 2.57/3.92 (3.25) | E7/E4 | 4.71/5.27 b (4.99) | ||||
| QTkw5A.5 | AX-109524552-AX-109031208 | 601.85–602.95 | E10 | 10.59 | E10/BLUE-HN | 3.95/3.90 b(3.93) | ||||
| QTkw5B.5 | AX-110926792-AX-110417642 | 594.56–595.62 | E3/E4/E10 | 4.36/3.78/2.46 (3.53) d | ||||||
| QTkw5D.3 | AX-110391491-AX-89331886 | 429.78–438.42 | BLUE-LN | 2.90 | BLUE-HN/E3 | 4.08/3.87 b (3.98) | ||||
| QTkw6A.1 | AX-109585639-AX-109979359 | 40.60–49.46 | E9/E1 | 4.96/6.45 (5.70) d | ||||||
| QTkw6A.3 | AX-111050302-AX-108817049 | 412.90–417.96 | E9/BLUE-LN | 6.21/6.09 (6.15) | E9/BLUE-LN | 5.79/4.25 b (5.02) | ||||
| QTkw6A.4 | AX-108849013-AX-109467855 | 561.01–574.01 | BLUE-LN/E10 | 4.02/4.13 (4.08) d | ||||||
| QTkw6B.1 | AX-109901945-AX-109871569 | 271.76–279.76 | BLUE-HN/BLUE-LN | 5.22/7.96 (6.59) c | ||||||
| QTkw6B.2 | AX-109968521-AX-108962284 | 453.76–457.76 | E4/E3/E6 | 8.90/10.12/5.38 (8.13) | E4/E9/BLUE-LN/E3/E10 | 7.17/6.73/8.15/9.74/2.78 b (6.91) | E9/BLUE-LN/E3 | 8.17/5.90/5.97 b (6.68) | ||
| QTkw6B.3 | AX-110149880-AX-110014652 | 514.73–515.88 | E1/E9 | 7.03/7.11 (7.07) c | ||||||
| QTkw7A.1 | AX-108833379-AX-110397800 | 122.95–130.95 | E2/BLUE-HN | 7.64/4.23 (5.93) d | ||||||
| QTkw7A.2 | AX-110487560-AX-109420524 | 664.95–688.79 | E8 | 9.2 | E4/E8/BLUE-HN | 5.30/9.61/3.76 b (6.22) | ||||
| QTL a | ENV b | Position1 (Mb) | Flanking Markers | QTL | Position2 (Mb) | Flanking Markers | LOD | PVE% (AA) c | AA d |
|---|---|---|---|---|---|---|---|---|---|
| QTkw3A.3 | E2 | 230.76 | AX-109930806-AX-94495161 | QTkw4A.4 | 561.91 | AX-111591351-AX-94881516 | 3.24 | 9.29 | 1.08 |
| QTkw4A.4 | BLUE-HN | 541.91 | AX-111591351-AX-94881516 | QTkw5A.1 | 400.67 | AX-110393916-AX-108821970 | 3.27 | 9.97 | 0.90 |
| QKl3A.6 | E5 | 410.763 | AX-109416489-AX-111049355 | QKl5D.2 | 470.6189 | AX-111175690-AX-109382782 | 3.28 | 9.55 | 0.16 |
| QKl4A.3 | E7 | 581.9098 | AX-110526171-AX-111830901 | QKl6D.6 | 400.4398 | AX-108821954-AX-110689037 | 3.22 | 9.48 | 0.16 |
| QKw2B | E10 | 50.06 | AX-109364692-AX-111111325 | QKw4A.1 | 71.91 | AX-111163489-AX-111591351 | 3.1 | 7.74 | 0.1 |
| QKw2D.1 | E10 | 276.75 | AX-108885026-AX-111837110 | QKw4A.3 | 321.91 | AX-111591351-AX-94881516 | 3.43 | 7.66 | −0.09 |
| QKdr5B.1 | E4 | 310.3226 | AX-94627936-AX-109846736 | QKdr7B.2 | 720.0621 | AX-109274013-AX-109295053 | 3.98 | 10.36 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Liu, W.; Yan, G.; Dong, Y.; Guan, C.; Zhang, Z.; Zhao, C.; Xia, L.; Zhu, D.; Zhao, C.; et al. Conditional QTL Analysis and Fine Mapping for Thousand-Kernel Weight in Common Wheat. Plants 2025, 14, 1848. https://doi.org/10.3390/plants14121848
Guo H, Liu W, Yan G, Dong Y, Guan C, Zhang Z, Zhao C, Xia L, Zhu D, Zhao C, et al. Conditional QTL Analysis and Fine Mapping for Thousand-Kernel Weight in Common Wheat. Plants. 2025; 14(12):1848. https://doi.org/10.3390/plants14121848
Chicago/Turabian StyleGuo, Haoru, Wei Liu, Geling Yan, Yifan Dong, Chongshuo Guan, Zhiyan Zhang, Changhao Zhao, Linxuan Xia, Da Zhu, Chunhua Zhao, and et al. 2025. "Conditional QTL Analysis and Fine Mapping for Thousand-Kernel Weight in Common Wheat" Plants 14, no. 12: 1848. https://doi.org/10.3390/plants14121848
APA StyleGuo, H., Liu, W., Yan, G., Dong, Y., Guan, C., Zhang, Z., Zhao, C., Xia, L., Zhu, D., Zhao, C., Sun, H., Wu, Y., Wu, J., Qin, R., & Cui, F. (2025). Conditional QTL Analysis and Fine Mapping for Thousand-Kernel Weight in Common Wheat. Plants, 14(12), 1848. https://doi.org/10.3390/plants14121848