

Research on the Species Diversity and Distribution Patterns of Wild Ribes in Northeast Asia
Abstract
1. Introduction
2. Results
2.1. Species Diversity of Wild Ribes in Northeast Asia
2.1.1. Wild Ribes Species Composition
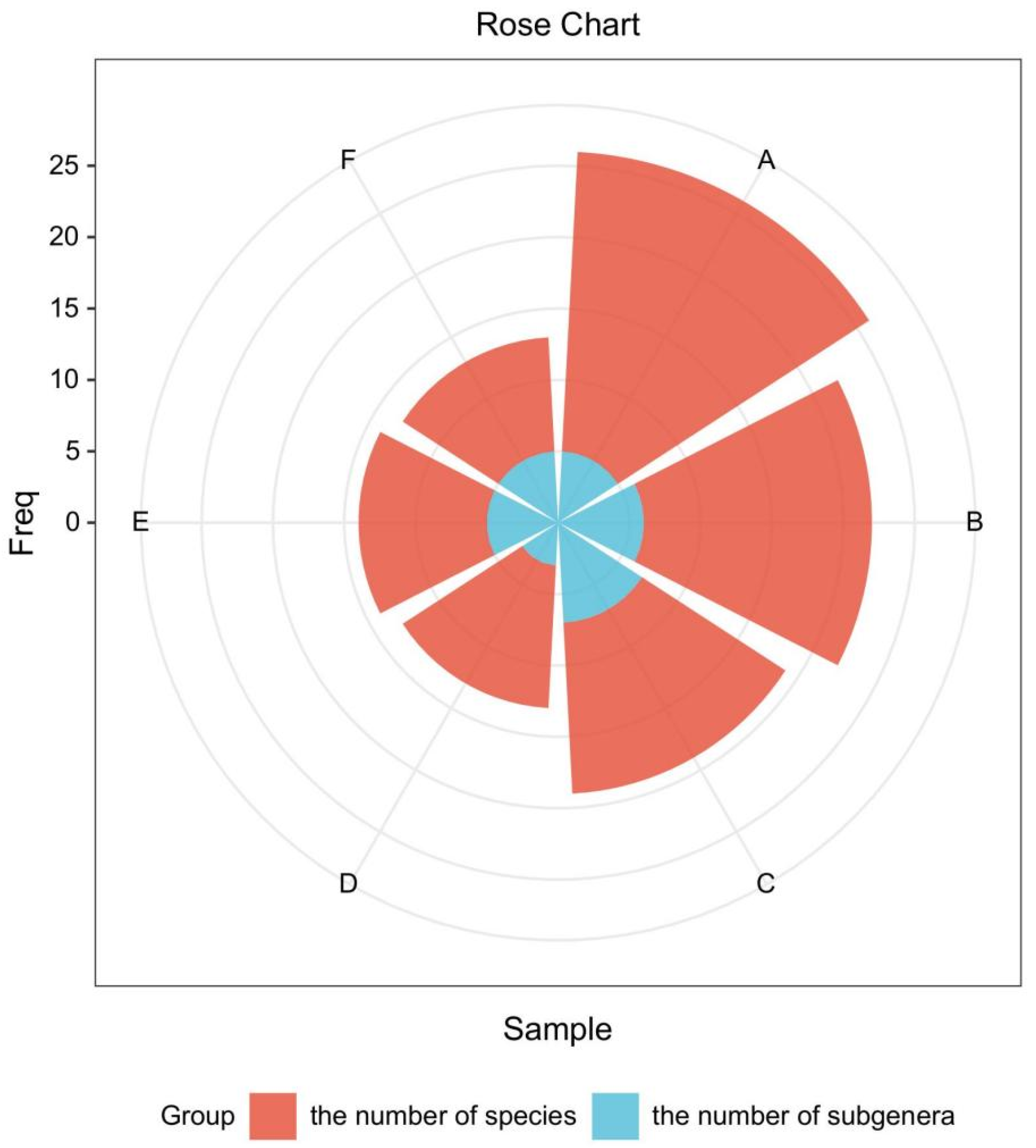
2.1.2. Analysis of Wild Ribes Species Composition in Each Country and Region of Northeast Asia
2.1.3. Distribution Center of Ribes in Northeast Asia
2.2. Distribution Patterns of Ribes in Northeast Asia and Their Correlation with Environmental Factors
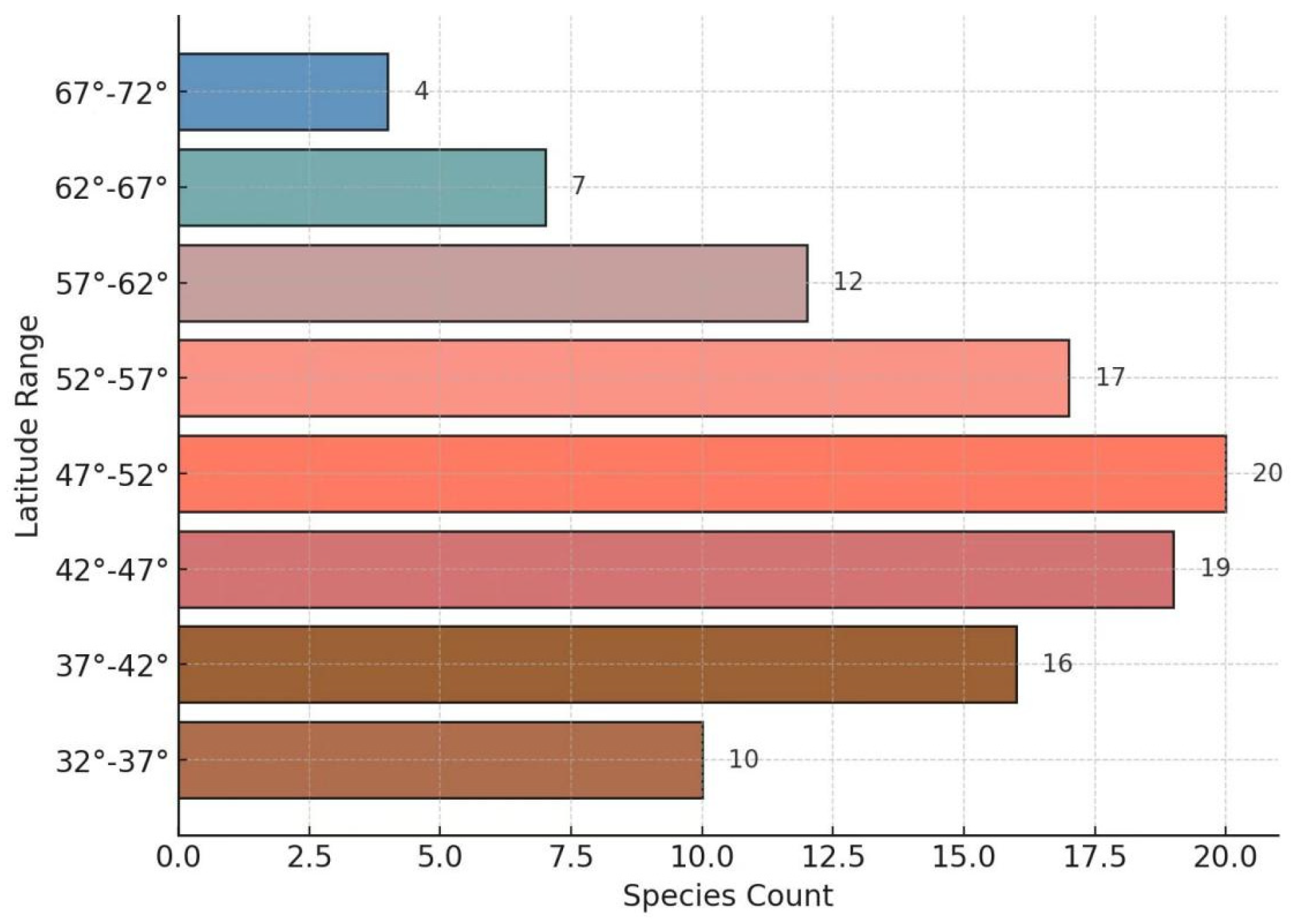
2.2.1. Spatial Distribution Pattern of Wild Ribes Species Richness in Northeast Asia
2.2.2. Relationship Between Wild Ribes Species Richness Patterns and Environmental Factors in Northeast Asia
3. Discussion
3.1. Species Diversity and Spatial Distribution Patterns of Wild Ribes Species in Northeast Asia
3.2. Temperature as the Primary Driver of Ribes Species Richness
4. Materials and Methods
4.1. Research Area
4.2. Data Collection
4.2.1. Species Distribution Data
4.2.2. Environmental Data
4.3. Data Processing
4.3.1. Species Distribution and Richness Calculation
- For analyses across the entire Northeast Asian region, both species and infraspecific taxa (e.g., varieties) were treated as separate species.
- In analyses at the country or region level, if both a species and its variety co-occurred in the same area, the variety was merged under the original species.
4.3.2. Correlation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soltis, S.D.E. Phylogenetic relationships in Ribes (Grossulariaceae) inferred from ITS sequence data. Taxon 2003, 52, 51–66. [Google Scholar]
- Lu, L.T. A Study on the Genus Ribes in China. J. Syst. Evol. 1995, 1, 58–75. [Google Scholar]
- Sun, Q.; Wang, N.; Xu, W.H.; Zhou, H.K. Genus Ribes Linn.(Grossulariaceae): A comprehensive review of traditional uses, phytochemistry, pharmacology and clinical applications. J. Ethnopharmacol. 2021, 276, 114166. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Wang, N.; Xu, W.H.; Zhou, H.K. Ribes himalense as potential source of natural bioactive compounds: Nutritional, phytochemical, and antioxidant properties. Food Sci. Nutr. 2021, 9, 2968–2984. [Google Scholar] [CrossRef] [PubMed]
- Cortez, R.E.; de Mejia, E.G. Blackcurrants (Ribes nigrum): A review on chemistry, processing, and health benefits. J. Food Sci. 2019, 84, 2387–2401. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Fidelis, M.; Haapakoski, M.; Dos Santos Lima, A.; Viil, J.; Hellström, J.; Rätsep, R.; Kaldmäe, H.; Bleive, U.; Azevedo, L.; et al. Enzyme-assisted extraction of anthocyanins and other phenolic compounds from blackcurrant (Ribes nigrum L.) press cake: From processing to bioactivities. Food Chem. 2022, 391, 133240. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.F.; Wang, Y.G.; Yang, G.L.; Tilyek, A.; Zhang, C.L.; Li, S.H.; Yu, B.Y.; Chai, C.Z.; Cao, Z.Y. Ribes diacanthum Pall (RDP) ameliorates UUO-induced renal fibrosis via both canonical and non-canonical TGF-β signaling pathways in mice. J. Ethnopharmacol. 2019, 231, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Park, Y.J.; Lee, B.; Park, E.; Kim, H.; Choi, C.W.; Kim, M.S. Ribes fasciculatum Ameliorates High-Fat-Diet-Induced Obesity by Elevating Peripheral Thermogenic Signaling. Molecules 2022, 27, 1649. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Jang, M.J.; Choi, Y.H.; Hwang, H.; Rhim, H.; Lee, B.; Choi, C.W.; Kim, M.S. Central administration of afzelin extracted from Ribes fasciculatum improves cognitive and memory function in a mouse model of dementia. Sci. Rep. 2021, 11, 9182. [Google Scholar] [CrossRef] [PubMed]
- Hovhannisyan, Z.; Timotina, M.; Manoyan, J.; Gabrielyan, L.; Petrosyan, M.; Kusznierewicz, B.; Bartoszek, A.; Jacob, C.; Ginovyan, M.; Trchounian, K.; et al. Ribes nigrum L. extract-mediated green synthesis and antibacterial action mechanisms of silver nanoparticles. Antibiotics 2022, 11, 1415. [Google Scholar] [CrossRef] [PubMed]
- Rizwana, H.; Alwhibi, M.S.; Al-Judaie, R.A.; Aldehaish, H.A.; Alsaggabi, N.S. Sunlight-mediated green synthesis of silver nanoparticles using the berries of Ribes rubrum (red currants): Characterisation and evaluation of their antifungal and antibacterial activities. Molecules 2022, 27, 2186. [Google Scholar] [CrossRef] [PubMed]
- Taghavizadeh Yazdi, M.E.; Khara, J.; Housaindokht, M.R.; Sadeghnia, H.R.; Esmaeilzadeh Bahabadi, S.; Sadegh Amiri, M.; Mosawee, H.; Taherzadeh, D.; Darroudi, M. Role of Ribes khorassanicum in the biosynthesis of AgNPs and their antibacterial properties. IET Nanobiotechnol. 2019, 13, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Wu, Y.Z.; Zhang, F.; Zheng, S.T.; Wang, L.B.; Bai, J.W.; Yang, Y. Preparation of Ribes nigrum L. polysaccharides-stabilized selenium nanoparticles for enhancement of the anti-glycation and α-glucosidase inhibitory activities. Int. J. Biol. Macromol. 2023, 253, 127122. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Zhang, M.L.; Zhou, M.; Zheng, B.J. A newly recorded plant of the genus Ribes from China. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2021, 45, 231–233. [Google Scholar]
- Zhang, Z.W.; Bai, H.C.; Liu, Z.; Li, W.G.; Yang, G.; Zheng, B.J. Species Diversity and Distribution Pattern of Wild Ribes in Northeast China. Bull. Bot. Res. 2024, 44, 192–199. [Google Scholar]
- Zhang, B.S.; Wang, M.J.; Zheng, B.J. New data of Ribes from Heilongjiang Province. J. Northeast For. Univ. 2019, 47, 113–114. [Google Scholar]
- Iwatsuki, K.; Bouffoud, D.E.; Ohba, H. Flora of Japan 2b; Kodansha: Tokyo, Japan, 2001. [Google Scholar]
- Im, R.J. Flora Coreana; Science and Technology Publishing House: Pyongyang, Republic of Korea, 2000. [Google Scholar]
- Urgamal, M.; Oyuntsetseg, B.; Nyambayar, D.; Choimaa, D. Conspectus of the Vascular Plants of Mongolia; Admon Printing: Ulaanbaatar, Mongolia, 2014. [Google Scholar]
- Lu, L.T.; Huang, S. Flora of China; Science Press: Beijing, China, 1995; pp. 278–362. [Google Scholar]
- Fu, P.Y. Clavis Plantarum Chinae Boreali-Orientalis; Science Press: Beijing, China, 1995. [Google Scholar]
- Zheng, B.J. Flora of Heilongjiang Province; Northeast Forestry University Press: Harbin, China, 2018; pp. 33–52. [Google Scholar]
- Wang, H.F.; Dong, X.Y.; Mu, L.Q. Checklist of tracheophyte in Heilongjiang Province. Biodivers. Sci. 2022, 30, 22184. [Google Scholar] [CrossRef]
- Malyschev, L.I.; Peschkova, G.A. Flora of Siberia; Science Publishers: Enfield, NH, USA, 2004; Volume 7, pp. 214–224. [Google Scholar]
- Zhang, S.M.; Zhang, Y.; Xu, L. Flora of Ligneous Plants of Liaoning; Liaoning Science and Technology Publishing House: Shenyang, China, 2019. [Google Scholar]
- Sakaguchi, S.; Oishi, M.; Takahashi, D.; Matsuo, A.; Hirota, S.K.; Suyama, Y.; Setoguchi, H. Ribes fujisanense (Grossulariaceae): A New Obligate Epiphytic Species of Gooseberry Discovered in Central Japan. Acta Phytotaxon. Geobot. 2022, 73, 49–56. [Google Scholar]
- Flora of Korea Editorial Committee. Flora of Korea. In Rosidae: Rhamnaceae to Apiaceae; National Institute of Biological Resources: Incheon, Republic of Korea, 2017; Volume 5c. [Google Scholar]
- Busch, N.A.; Fedtchenko, B.A.; Filippov, Y. Flora of Siberia and the Far East; Botanical Museum of Imperial Academy of Sciences: St. Petersburg, Russia; pp. 1913–1931.
- Zhang, B.S.; Yu, Z.Y.; Xu, Z.C.; Zheng, B.J. A phylogenetic and morphological evolution study of Ribes in China using RAD-seq. Plants 2023, 12, 829. [Google Scholar] [CrossRef] [PubMed]
- Stralberg, D.; Carroll, C.; Nielsen, S.E. Toward a climate-informed North American protected areas network: Incorporating climate-change refugia and corridors in conservation planning. Conserv. Lett. 2020, 13, e12712. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.7uyxxy (accessed on 13 May 2025).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subgenera | Scientific Name |
|---|---|
| subg. Berisia | Ribes diacantha Pall. |
| Ribes dikuscha Fisch. ex Turcz. | |
| Ribes giraldii var. cuneatum F.T.Wang & S.X.Li | |
| Ribes giraldii var. polyanthum Kitag. | |
| Ribes heterotrichum C.A.Mey. | |
| Ribes komarovii Pojark. | |
| Ribes komarovii var. cuneifolium Liou | |
| Ribes maximowiczianum Kom. | |
| Ribes pulchellum Turcz. | |
| Ribes pulchellum var. manshuriense F.T.Wang & S.X.Li | |
| Ribes saxatile Pall. | |
| subg. Ribesia | Ribes altissimum Turcz. ex Pojark. |
| Ribes ambiguum Maxim. | |
| Ribes atropurpureum C.A.Mey. | |
| Ribes latifolium Jancz. | |
| Ribes mandshuricum (Maxim.) Kom. | |
| Ribes mandshuricum var. subglabrum Kom. | |
| Ribes meyeri Maxim. | |
| Ribes palczewskii (Jancz.) Pojark. | |
| Ribes pallidiflorum Pojark. | |
| Ribes triste Pall. | |
| Ribes triste var. repens (A. I. Baranov) L. T. Lu | |
| subg. Coreosma | Ribes fontaneum Bochkarn. |
| Ribes frangrans Pallas. | |
| Ribes graveolens Bunge | |
| Ribes japonicum Maxim. | |
| Ribes nigrum L. | |
| Ribes procumbens Pall. | |
| subg. Grossularia | Ribes burejense F.Schmidt |
| Ribes burejense var. villosum L.T.Lu | |
| Ribes fujisanense S.Sakag. & M.Oishi | |
| Ribes sinanense F.Maek. | |
| subg. Hemibotrya | Ribes fasciculatum Siebold & Zucc. |
| Ribes fasciculatum var. chinense Maxim. | |
| subg. Grossularioides | Ribes horridum Rupr. ex Maxim. |
| subg. Hertiera | Ribes sachalinense (F.Schmidt) Nakai |
| Country/Region | Subgenera | Scientific Name |
|---|---|---|
| Northeast Asia region of Russia | subg. Berisia | Ribes diacantha Pall. |
| subg. Berisia | Ribes dikuscha Fisch. ex Turcz. | |
| subg. Berisia | Ribes komarovii Pojark. | |
| subg. Berisia | Ribes maximowiczianum Kom. | |
| subg. Berisia | Ribes pulchellum Turcz. | |
| subg. Berisia | Ribes saxatile Pall. | |
| subg. Coreosma | Ribes fontaneum Bochkarn. | |
| subg. Coreosma | Ribes frangrans Pallas. | |
| subg. Coreosma | Ribes graveolens Bunge | |
| subg. Coreosma | Ribes nigrum L. | |
| subg. Coreosma | Ribes procumbens Pall. | |
| subg. Hertiera | Ribes sachalinense (F.Schmidt) Nakai | |
| subg. Ribesia | Ribes altissimum Turcz. ex Pojark. | |
| subg. Ribesia | Ribes atropurpureum C.A.Mey. | |
| subg. Ribesia | Ribes latifolium Jancz. | |
| subg. Ribesia | Ribes mandshuricum (Maxim.) Kom. | |
| subg. Ribesia | Ribes meyeri Maxim. | |
| subg. Ribesia | Ribes palczewskii (Jancz.) Pojark. | |
| subg. Ribesia | Ribes pallidiflorum Pojark. | |
| subg. Ribesia | Ribes triste Pall. | |
| subg. Grossularioides | Ribes horridum Rupr. ex Maxim. | |
| Northeast China | subg. Berisia | Ribes diacantha Pall. |
| subg. Berisia | Ribes giraldii var. cuneatum F.T.Wang & S.X.Li | |
| subg. Berisia | Ribes giraldii var. polyanthum Kitag. | |
| subg. Berisia | Ribes komarovii Pojark. | |
| subg. Berisia | Ribes maximowiczianum Kom. | |
| subg. Berisia | Ribes pulchellum Turcz. | |
| subg. Coreosma | Ribes frangrans Pallas. | |
| subg. Coreosma | Ribes nigrum L. | |
| subg. Coreosma | Ribes procumbens Pall. | |
| subg. Grossularia | Ribes burejense F.Schmidt | |
| subg. Hemibotrya | Ribes fasciculatum var. chinense Maxim. | |
| subg. Ribesia | Ribes altissimum Turcz. ex Pojark. | |
| subg. Ribesia | Ribes latifolium Jancz. | |
| subg. Ribesia | Ribes mandshuricum (Maxim.) Kom. | |
| subg. Ribesia | Ribes palczewskii (Jancz.) Pojark. | |
| subg. Ribesia | Ribes triste Pall. | |
| subg. Grossularioides | Ribes horridum Rupr. ex Maxim. | |
| Japan | subg. Berisia | Ribes maximowiczianum Kom. |
| subg. Coreosma | Ribes japonicum Maxim. | |
| subg. Coreosma | Ribes nigrum L. | |
| subg. Coreosma | Ribes procumbens Pall. | |
| subg. Grossularia | Ribes fujisanense S.Sakag. & M.Oishi | |
| subg. Grossularia | Ribes sinanense F.Maek. | |
| subg. Hemibotrya | Ribes fasciculatum Siebold & Zucc. | |
| subg. Hertiera | Ribes sachalinense (F.Schmidt) Nakai | |
| subg. Ribesia | Ribes ambiguum Maxim. | |
| subg. Ribesia | Ribes latifolium Jancz. | |
| subg. Ribesia | Ribes triste Pall. | |
| subg. Grossularioides | Ribes horridum Rupr. ex Maxim. | |
| Mongolia | subg. Berisia | Ribes diacantha Pall. |
| subg. Berisia | Ribes heterotrichum C.A.Mey. | |
| subg. Berisia | Ribes pulchellum Turcz. | |
| subg. Coreosma | Ribes frangrans Pallas. | |
| subg. Coreosma | Ribes graveolens Bunge | |
| subg. Coreosma | Ribes nigrum L. | |
| subg. Coreosma | Ribes procumbens Pall. | |
| subg. Ribesia | Ribes altissimum Turcz. ex Pojark. | |
| subg. Ribesia | Ribes atropurpureum C.A.Mey. | |
| subg. Ribesia | Ribes meyeri Maxim. | |
| South Korea | subg. Berisia | Ribes diacantha Pall. |
| subg. Berisia | Ribes komarovii Pojark. | |
| subg. Berisia | Ribes maximowiczianum Kom. | |
| subg. Grossularia | Ribes burejense F.Schmidt | |
| subg. Hemibotrya | Ribes fasciculatum Siebold & Zucc. | |
| subg. Ribesia | Ribes latifolium Jancz. | |
| subg. Ribesia | Ribes mandshuricum (Maxim.) Kom. | |
| subg. Ribesia | Ribes triste Pall. | |
| subg. Grossularioides | Ribes horridum Rupr. ex Maxim. | |
| North Korea | subg. Berisia | Ribes komarovii Pojark. |
| subg. Berisia | Ribes maximowiczianum Kom. | |
| subg. Grossularia | Ribes burejense F.Schmidt | |
| subg. Hemibotrya | Ribes fasciculatum Siebold & Zucc. | |
| subg. Ribesia | Ribes latifolium Jancz. | |
| subg. Ribesia | Ribes mandshuricum (Maxim.) Kom. | |
| subg. Ribesia | Ribes triste Pall. | |
| subg. Grossularioides | Ribes horridum Rupr. ex Maxim. |
| Species | Northeast Asia Region of Russia | Northeast China | Japan | Mongolia | South Korea | North Korea |
|---|---|---|---|---|---|---|
| Ribes sachalinense (F.Schmidt) Nakai | 1 | 0 | 1 | 0 | 0 | 0 |
| Ribes altissimum Turcz. ex Pojark. | 1 | 1 | 0 | 1 | 0 | 0 |
| Ribes atropurpureum C.A.Mey. | 1 | 0 | 0 | 1 | 0 | 0 |
| Ribes diacantha Pall. | 1 | 1 | 0 | 1 | 1 | 0 |
| Ribes dikuscha Fisch. ex Turcz. | 1 | 0 | 0 | 0 | 0 | 0 |
| Ribes fontaneum Bochkarn. | 1 | 0 | 0 | 0 | 0 | 0 |
| Ribes frangrans Pallas. | 1 | 1 | 0 | 1 | 0 | 0 |
| Ribes graveolens Bunge | 1 | 0 | 0 | 1 | 0 | 0 |
| Ribes horridum Rupr. ex Maxim. | 1 | 1 | 1 | 0 | 1 | 1 |
| Ribes komarovii Pojark. | 1 | 1 | 0 | 0 | 1 | 1 |
| Ribes latifolium Jancz. | 1 | 1 | 1 | 0 | 1 | 1 |
| Ribes mandshuricum (Maxim.) Kom. | 1 | 1 | 0 | 0 | 1 | 1 |
| Ribes maximowiczianum Kom. | 1 | 1 | 1 | 0 | 1 | 1 |
| Ribes meyeri Maxim. | 1 | 0 | 0 | 1 | 0 | 0 |
| Ribes nigrum L. | 1 | 1 | 1 | 1 | 0 | 0 |
| Ribes palczewskii (Jancz.) Pojark. | 1 | 1 | 0 | 0 | 0 | 0 |
| Ribes pallidiflorum Pojark. | 1 | 0 | 0 | 0 | 0 | 0 |
| Ribes procumbens Pall. | 1 | 1 | 1 | 1 | 0 | 0 |
| Ribes pulchellum Turcz. | 1 | 1 | 0 | 1 | 0 | 0 |
| Ribes saxatile Pall. | 1 | 0 | 0 | 0 | 0 | 0 |
| Ribes triste Pall. | 1 | 1 | 1 | 0 | 1 | 1 |
| Ribes burejense F.Schmidt | 0 | 1 | 0 | 0 | 1 | 1 |
| Ribes fasciculatum var. chinense Maxim. | 0 | 1 | 0 | 0 | 0 | 0 |
| Ribes giraldii Jancz. | 0 | 1 | 0 | 0 | 0 | 0 |
| Ribes ambiguum Maxim. | 0 | 0 | 1 | 0 | 0 | 0 |
| Ribes fasciculatum Siebold & Zucc. | 0 | 0 | 1 | 0 | 1 | 1 |
| Ribes fujisanense S.Sakag. & M.Oishi | 0 | 0 | 1 | 0 | 0 | 0 |
| Ribes japonicum Maxim. | 0 | 0 | 1 | 0 | 0 | 0 |
| Ribes sinanense F.Maek. | 0 | 0 | 1 | 0 | 0 | 0 |
| Ribes heterotrichum C.A.Mey. | 0 | 0 | 0 | 1 | 0 | 0 |
| Variable Type | Variable | Variable Description | [Max, Min] | Mean Value | Unit | T-Value Correlation Coefficient | p-Value |
|---|---|---|---|---|---|---|---|
| Climate | BIO1 | Annual Mean Temperature | [−17.101, 21.843] | −1.906 ± 7.515 | °C | 11.81 | <2 × 10−16 *** |
| BIO2 | Mean Diurnal Range | [1.628, 18.229] | 10.795 ± 2.523 | - | 7.039 | 2.01 × 10−12 *** | |
| BIO3 | Isothermality | [12.878, 80.903] | 23.246 ± 5.584 | - | 13.59 | <2 × 10−16 *** | |
| BIO4 | Temperature Seasonality | [212.407, 2310.871] | 1364.185 ± 347.707 | - | −9.213 | <2 × 10−16 *** | |
| BIO5 | Max Temperature of Warmest Month | [−0.315, 37.688] | 22.19 ± 5.379 | °C | 7.195 | 6.52 × 10−13 *** | |
| BIO6 | Min Temperature of Coldest Month | [−48.566, 10.161] | −25.251 ± 10.684 | °C | 10.10 | <2 × 10−16 *** | |
| BIO7 | Temperature Annual Range | [6.886, 71.383] | 47.441 ± 9.942 | - | −6.805 | <2 × 10−16 *** | |
| BIO8 | Mean Temperature of Wettest Quarter | [−5.28, 28.312] | 13.837 ± 5.506 | °C | 7.965 | 1.76 × 10−15 *** | |
| BIO9 | Mean Temperature of Driest Quarter | [−38.054, 22.096] | −14.75 ± 10.108 | °C | 9.729 | <2 × 10−16 *** | |
| BIO10 | Mean Temperature of Warmest Quarter | [−2.344, 28.312] | 14.643 ± 5.192 | °C | 8.744 | <2 × 10−16 *** | |
| BIO11 | Mean Temperature of Coldest Quarter | [−42.866, 14.825] | −18.707 ± 10.512 | °C | 11.39 | <2 × 10−16 *** | |
| BIO12 | Annual Precipitation | [13.653, 3238.4] | 469.173 ± 275.078 | mm | 1.698 | 0.0896 ns | |
| BIO13 | Precipitation of Wettest Month | [4.592, 538] | 82.711 ± 49.824 | mm | 8.924 | <2 × 10−16 *** | |
| BIO14 | Precipitation of Driest Month | [0, 138.8] | 13.942 ± 12.228 | mm | −1.224 | 0.221 ns | |
| BIO15 | Precipitation Seasonality | [15.385, 140.815] | 62.705 ± 29.082 | 8.141 | 4.21 × 10−16 *** | ||
| BIO16 | Precipitation of Wettest Quarter | [11.122, 1274.2] | 215.826 ± 125.478 | mm | 4.968 | 6.82 × 10−7 *** | |
| BIO17 | Precipitation of Driest Quarter | [0, 470.6] | 48.234 ± 40.823 | mm | 20.438 | 0.621 ns | |
| BIO18 | Precipitation of Warmest Quarter | [11.122, 964.6] | 210.241 ± 116.819 | mm | 4.800 | 1.6 × 10−6 *** | |
| BIO19 | Precipitation of Coldest Quarter | [0, 503.486] | 57.579 ± 50.016 | mm | 14.012 | <2 × 10−16 *** | |
| Topography | ELE | Mean Elevation | [−29, 5936.245] | 683.056 ± 974.05 | m | 8.502 | 0.00821 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Zhao, D.; Ge, X.; Zhang, Y.; Du, Y.; Liu, J.; Liu, Y.; Wang, H.; Zheng, B. Research on the Species Diversity and Distribution Patterns of Wild Ribes in Northeast Asia. Plants 2025, 14, 1780. https://doi.org/10.3390/plants14121780
Zhao X, Zhao D, Ge X, Zhang Y, Du Y, Liu J, Liu Y, Wang H, Zheng B. Research on the Species Diversity and Distribution Patterns of Wild Ribes in Northeast Asia. Plants. 2025; 14(12):1780. https://doi.org/10.3390/plants14121780
Chicago/Turabian StyleZhao, Ximing, Dandan Zhao, Xinrui Ge, Yin Zhang, Yuxiao Du, Jingguo Liu, Yuning Liu, Hongfeng Wang, and Baojiang Zheng. 2025. "Research on the Species Diversity and Distribution Patterns of Wild Ribes in Northeast Asia" Plants 14, no. 12: 1780. https://doi.org/10.3390/plants14121780
APA StyleZhao, X., Zhao, D., Ge, X., Zhang, Y., Du, Y., Liu, J., Liu, Y., Wang, H., & Zheng, B. (2025). Research on the Species Diversity and Distribution Patterns of Wild Ribes in Northeast Asia. Plants, 14(12), 1780. https://doi.org/10.3390/plants14121780