
Indole-3-Butyric Acid Enhances Root Formation and Alleviates Low-Temperature Stress in Sugarcane: Molecular Insights and Identification of Candidate Genes

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. IBA Promoted the Development of the Root System in Sugarcane Under Low Temperature
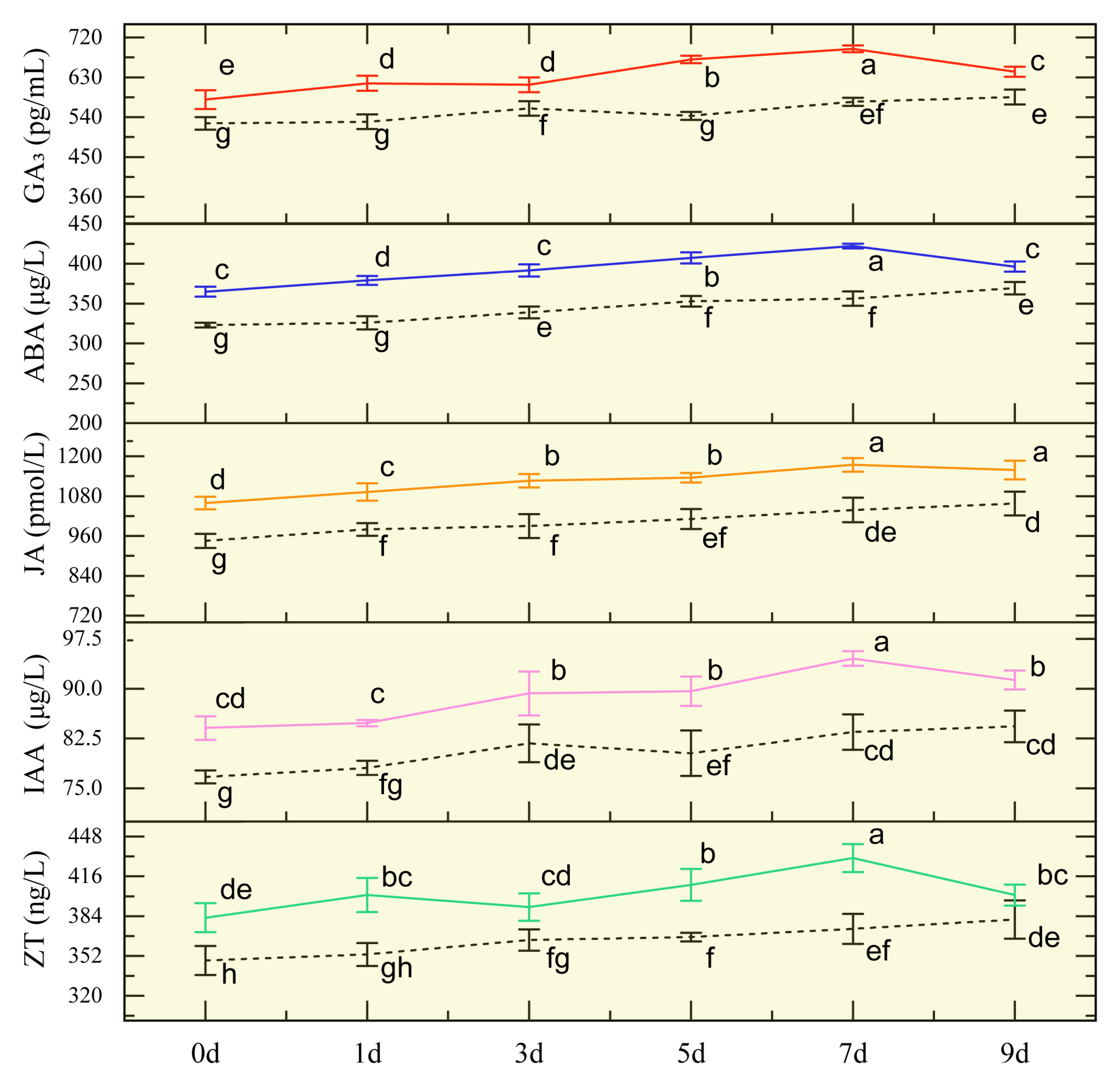
2.2. IBA Enhanced the Activities of Phytohormones in Sugarcane Roots Under Low Temperature
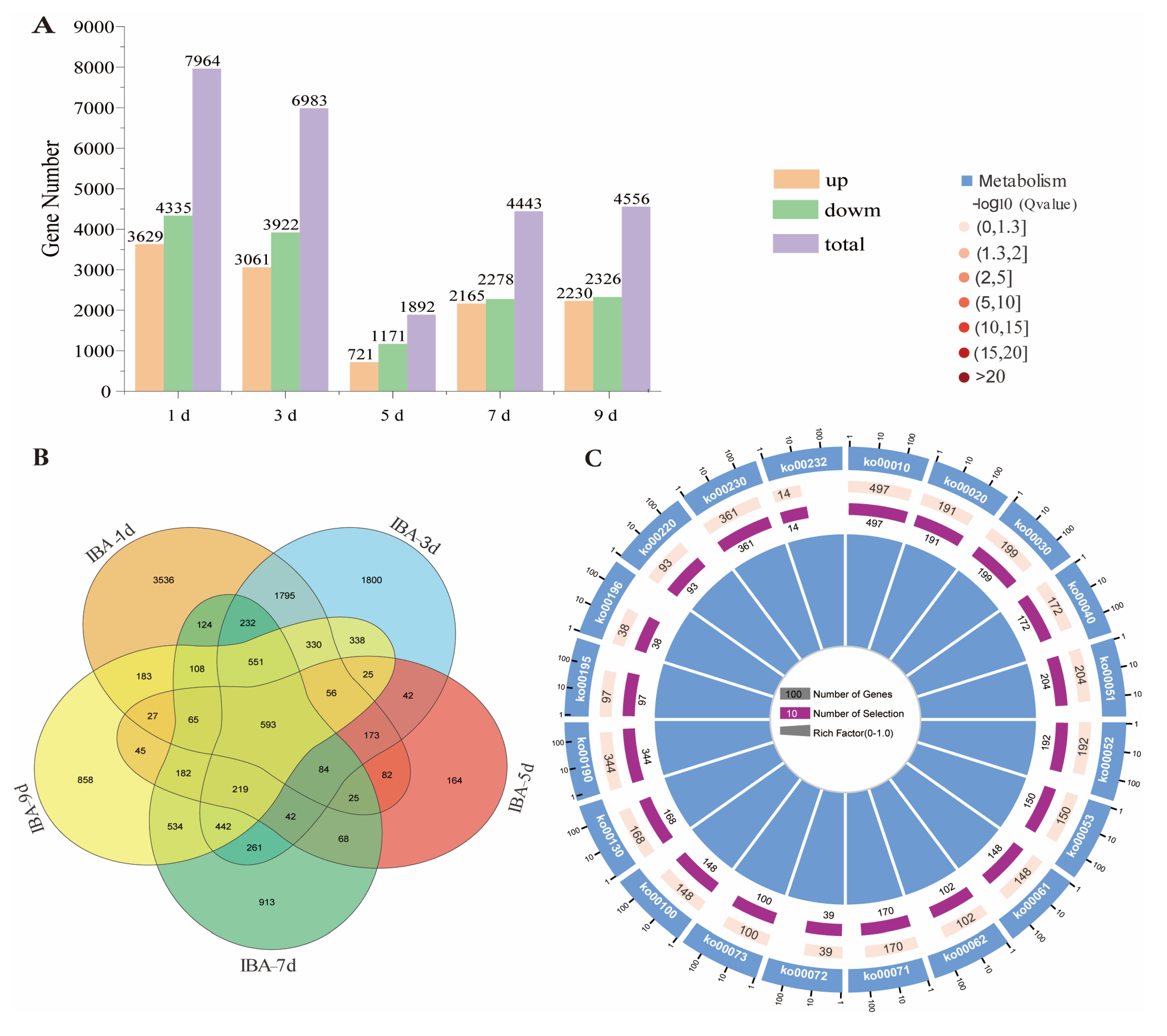
2.3. IBA Induced a Significant Number of Responsive Genes in Sugarcane Roots Under Low Temperature
2.4. Phytohormone Activities Exhibited a Strong Correlation with Gene Expression Levels in Sugarcane Roots
2.5. The Core Genes Associated with IBA Stimulation in Sugarcane Roots Under Low-Temperature Conditions Were Identified
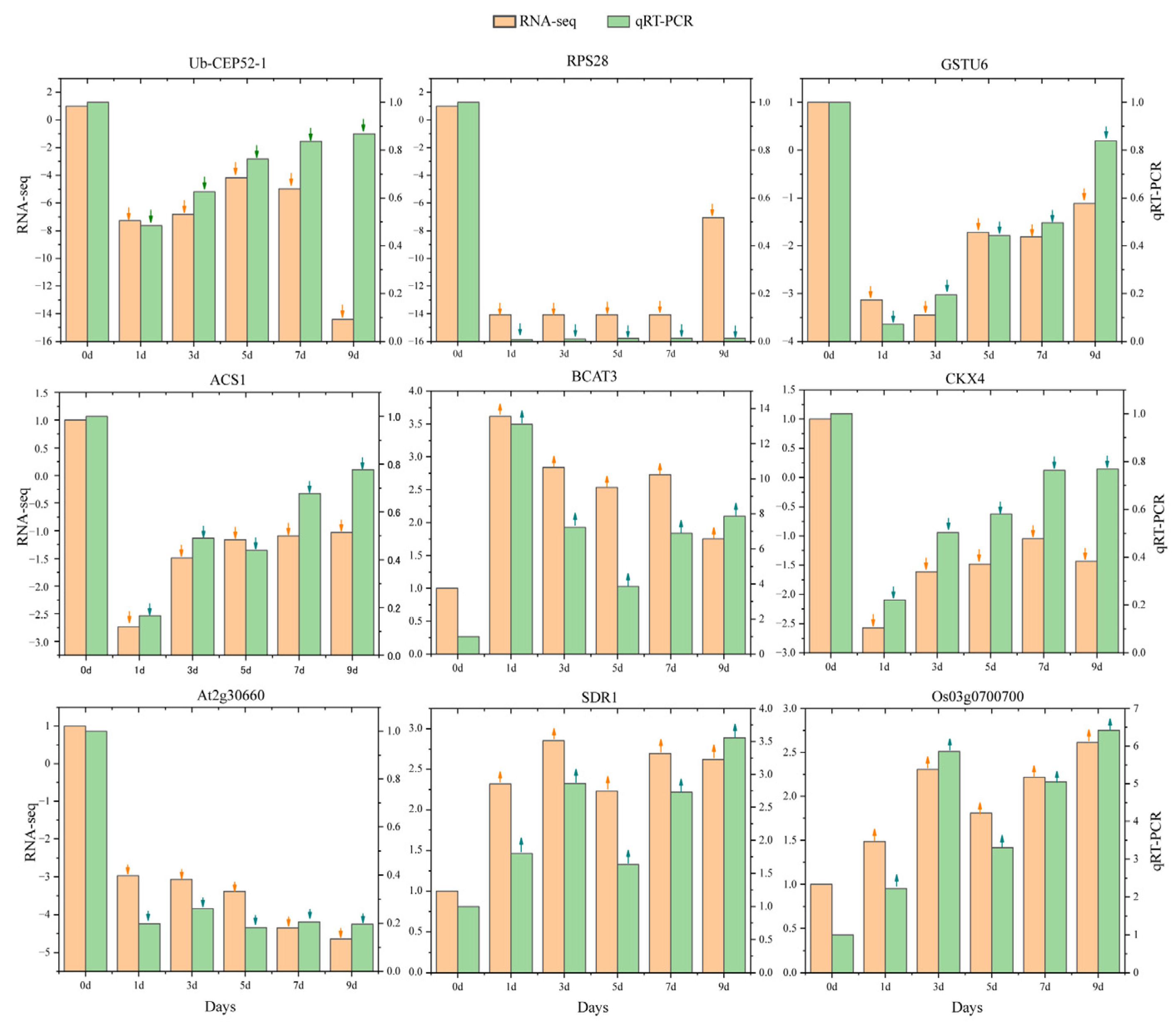
2.6. qRT-PCR Validation Confirmed the Expression Patterns of Core Genes in Response to IBA Stimulation Under Low Temperature
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Phytohormone Activity Analysis
4.3. RNA Extraction, Library Construction, and Sequencing
4.4. Sequencing Data Processing
4.5. Sequencing Data Analysis
4.6. qRT-PCR Analysis
4.7. Statistical Analysis
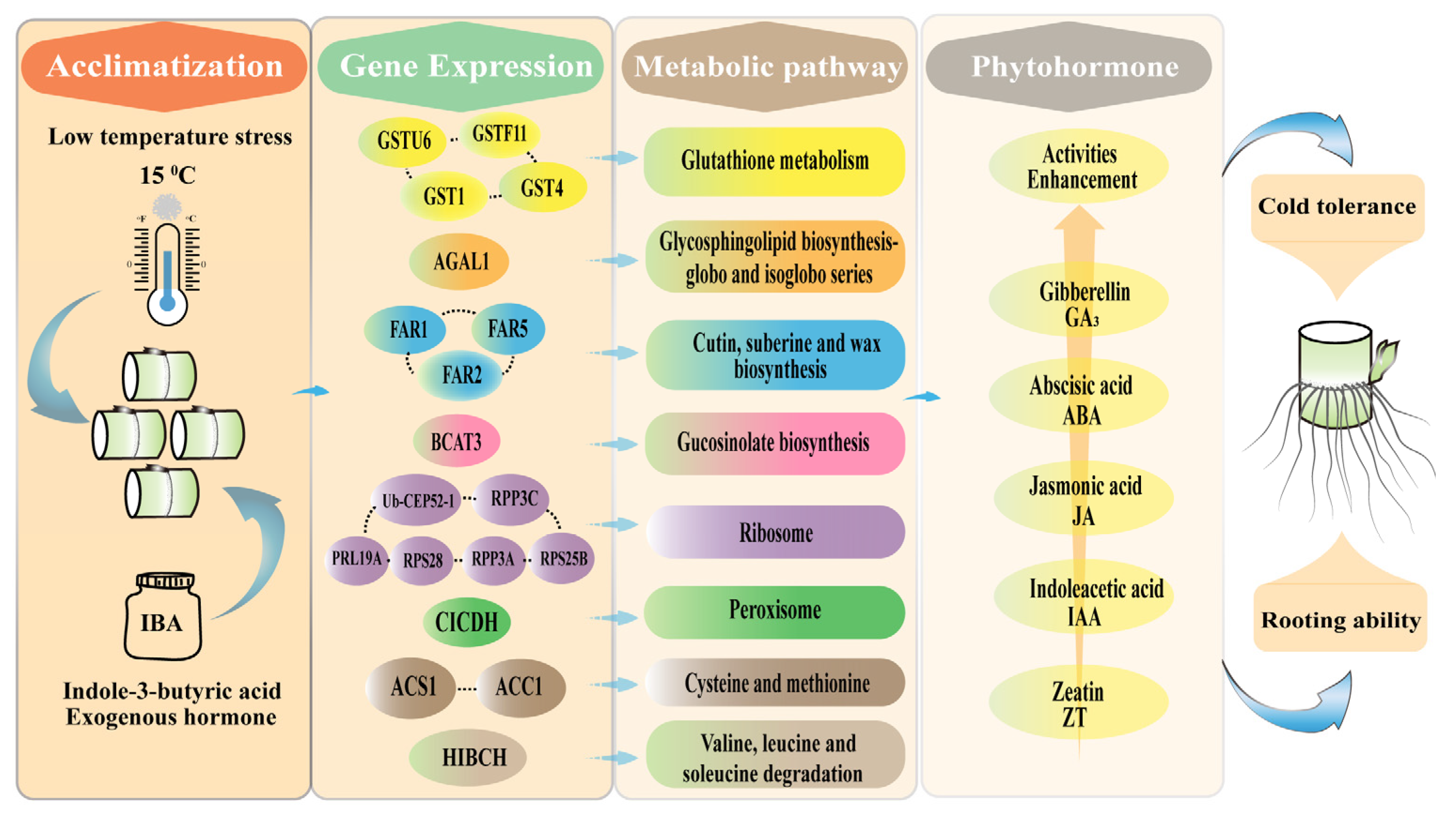
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Qi, Y.; Gao, X.; Zeng, Q.; Zheng, Z.; Wu, C.; Yang, R.; Feng, X.; Wu, Z.; Fan, L.; Huang, Z. Sugarcane Breeding, Germplasm Development and Related Molecular Research in China. Sugar Tech 2022, 24, 73–85. [Google Scholar] [CrossRef]
- Otto, R.; Silva, A.P.; Franco, H.C.J.; Oliveira, E.C.A.; Trivelin, P.C.O. High Soil Penetration Resistance Reduces Sugarcane Root System Development. Soil Tillage Res. 2011, 117, 201–210. [Google Scholar] [CrossRef]
- Lovera, L.H.; de Souza, Z.M.; Esteban, D.A.A.; de Oliveira, I.N.; Farhate, C.V.V.; Lima, E.d.S.; Panosso, A.R. Sugarcane Root System: Variation over Three Cycles under Different Soil Tillage Systems and Cover Crops. Soil Tillage Res. 2021, 208, 104866. [Google Scholar] [CrossRef]
- Damodaran, S.; Strader, L.C. Indole 3-Butyric Acid Metabolism and Transport in Arabidopsis Thaliana. Front. Plant Sci. 2019, 10, 851. [Google Scholar] [CrossRef]
- Waheed, A.; Hamid, F.S.; Ahmad, H.; Abbassi, F.M.; Aslam, S.; Shah, A.H.; Ahmad, N.; Naheed, Z.; Ali, H.; Khan, N. Effect of Indole Butyric Acid (IBA) on Early Root Formation(Tomato “ Sahil” Hybrid) Cuttings. J. Mater. Environ. Sci. 2015, 6, 272–279. [Google Scholar]
- Li, A.; Lakshmanan, P.; He, W.; Tan, H.; Liu, L.; Liu, H.; Liu, J.; Huang, D.; Chen, Z. Transcriptome Profiling Provides Molecular Insights into Auxin-Induced Adventitious Root Formation in Sugarcane (Saccharum Spp. Interspecific Hybrids) Microshoots. Plants 2020, 9, 931. [Google Scholar] [CrossRef]
- Mustafa, G.; Khan, M.S. Differential Role of Indolebutyric Acid in Sugarcane Root Development. Sugar Tech 2016, 18, 55–60. [Google Scholar] [CrossRef]
- Xu, L.; Deng, Z.-N.; Wu, K.-C.; Malviya, M.K.; Solanki, M.K.; Verma, K.K.; Pang, T.; Li, Y.-J.; Liu, X.-Y.; Kashyap, B.K.; et al. Transcriptome Analysis Reveals a Gene Expression Pattern That Contributes to Sugarcane Bud Propagation Induced by Indole-3-Butyric Acid. Front. Plant Sci. 2022, 13, 852886. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, X.; Wu, Y.; Xu, X. Indole Butyric Acid Alleviated Cadmium Toxicity in Lettuce by Increasing the Polysaccharide Contents and the Cadmium-Binding Capacity of Root Cell Walls. J. Soil Sci. Plant Nutr. 2024, 24, 5195–5207. [Google Scholar] [CrossRef]
- Demecsová, L.; Zelinová, V.; Liptáková, Ľ.; Valentovičová, K.; Tamás, L. Indole-3-Butyric Acid Priming Reduced Cadmium Toxicity in Barley Root Tip via NO Generation and Enhanced Glutathione Peroxidase Activity. Planta 2020, 252, 46. [Google Scholar] [CrossRef]
- Li, S.-W.; Zeng, X.-Y.; Leng, Y.; Feng, L.; Kang, X.-H. Indole-3-Butyric Acid Mediates Antioxidative Defense Systems to Promote Adventitious Rooting in Mung Bean Seedlings under Cadmium and Drought Stresses. Ecotoxicol. Environ. Saf. 2018, 161, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Hernandez, J.A.; Bariat, L.; Enders, T.A.; Strader, L.C.; Reichheld, J.-P.; Belin, C. A Glutathione-Dependent Control of the Indole Butyric Acid Pathway Supports Arabidopsis Root System Adaptation to Phosphate Deprivation. J. Exp. Bot. 2020, 71, 4843–4857. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, S. Adverse Effects of Salt Stress on Rootability of Rosmarinus Officinalis Cuttings and Their Alleviation by Indole-3-Butyric Acid (IBA) and Bacillus Subtilis. Egypt. J. Hortic. 2019, 46, 279–293. [Google Scholar] [CrossRef]
- Ahsan, M.; Radicetti, E.; Jamal, A.; Ali, H.M.; Sajid, M.; Manan, A.; Bakhsh, A.; Naeem, M.; Khan, J.A.; Valipour, M. Silicon Nanoparticles and Indole Butyric Acid Positively Regulate the Growth Performance of Freesia Refracta by Ameliorating Oxidative Stress under Chromium Toxicity. Front. Plant Sci. 2024, 15, 1437276. [Google Scholar] [CrossRef]
- Heidari, P.; Reza Amerian, M.; Barcaccia, G. Hormone Profiles and Antioxidant Activity of Cultivated and Wild Tomato Seedlings under Low-Temperature Stress. Agronomy 2021, 11, 1146. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Wu, P.; Ma, Y.; Ahammed, G.J.; Hao, B.; Chen, J.; Wan, W.; Zhao, Y.; Cui, H.; Xu, W.; Cui, J.; et al. Insights into Melatonin-Induced Photosynthetic Electron Transport under Low-Temperature Stress in Cucumber. Front. Plant Sci. 2022, 13, 1029854. [Google Scholar] [CrossRef]
- Yu, J.; Cang, J.; Lu, Q.; Fan, B.; Xu, Q.; Li, W.; Wang, X. ABA Enhanced Cold Tolerance of Wheat ‘Dn1’ via Increasing ROS Scavenging System. Plant Signal Behav. 2020, 15, 1780403. [Google Scholar] [CrossRef]
- Chao, I.-L.; Cho, C.-L.; Chen, L.-M.; Liu, Z.-H. Effect of Indole-3-Butyric Acid on the Endogenous Indole-3-Acetic Acid and Lignin Contents in Soybean Hypocotyl during Adventitious Root Formation. J. Plant Physiol. 2001, 158, 1257–1262. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in Plants: An Integrated Overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.-Y.; Wu, L.-Y.; Zhang, C.-C.; Li, H.-L.; Tan, L.-Q.; Cao, H.-L.; Cheng, H. Transcriptome Analysis of Indole-3-Butyric Acid-Induced Adventitious Root Formation in Nodal Cuttings of Camellia Sinensis (L.). PLoS ONE 2014, 9, e107201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ni, C.; Dong, Y.; Jiang, X.; Liu, C.; Wang, W.; Zhao, C.; Li, G.; Xu, K.; Huo, Z. The Role of the Ascorbic Acid–Glutathione Cycle in Young Wheat Ears’ Response to Spring Freezing Stress. Plants 2023, 12, 4170. [Google Scholar] [CrossRef] [PubMed]
- Rowland, O.; Domergue, F. Plant Fatty Acyl Reductases: Enzymes Generating Fatty Alcohols for Protective Layers with Potential for Industrial Applications. Plant Sci. 2012, 193–194, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, Y.; You, Q.; Luo, W.; Wang, C.; Zhao, S.; Chai, G.; Li, T.; Shi, X.; Li, C.; et al. Three Fatty Acyl-Coenzyme A Reductases, BdFAR1, BdFAR2 and BdFAR3, Are Involved in Cuticular Wax Primary Alcohol Biosynthesis in Brachypodium Distachyon. Plant Cell Physiol. 2018, 59, 527–543. [Google Scholar] [CrossRef]
- Guan, L.; Xia, D.; Hu, N.; Zhang, H.; Wu, H.; Jiang, Q.; Li, X.; Sun, Y.; Wang, Y.; Wang, Z. OsFAR1 Is Involved in Primary Fatty Alcohol Biosynthesis and Promotes Drought Tolerance in Rice. Planta 2023, 258, 24. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, J.; He, Z.; Hu, N.; Luo, W.; Liu, X.; Shi, X.; Liu, T.; Jiang, Q.; An, P.; et al. BdFAR4, a Root-specific Fatty Acyl-coenzyme A Reductase, Is Involved in Fatty Alcohol Synthesis of Root Suberin Polyester in Brachypodium Distachyon. Plant J. 2021, 106, 1468–1483. [Google Scholar] [CrossRef]
- Chen, C.; Naveed, H.; Chen, K. Research Progress on Branched-Chain Amino Acid Aminotransferases. Front. Genet. 2023, 14, 1233669. [Google Scholar] [CrossRef]
- Mitreiter, S.; Gigolashvili, T. Regulation of Glucosinolate Biosynthesis. J. Exp. Bot. 2021, 72, 70–91. [Google Scholar] [CrossRef]
- Vik, D.; Mitarai, N.; Wulff, N.; Halkier, B.A.; Burow, M. Dynamic Modeling of Indole Glucosinolate Hydrolysis and Its Impact on Auxin Signaling. Front. Plant Sci. 2018, 9, 550. [Google Scholar] [CrossRef]
- Corredor-Moreno, P.; Minter, F.; Davey, P.E.; Wegel, E.; Kular, B.; Brett, P.; Lewis, C.M.; Morgan, Y.M.L.; Macías Pérez, L.A.; Korolev, A.V.; et al. The Branched-Chain Amino Acid Aminotransferase TaBCAT1 Modulates Amino Acid Metabolism and Positively Regulates Wheat Rust Susceptibility. Plant Cell 2021, 33, 1728–1747. [Google Scholar] [CrossRef]
- Malatrasi, M.; Corradi, M.; Svensson, J.T.; Close, T.J.; Gulli, M.; Marmiroli, N. A Branched-Chain Amino Acid Aminotransferase Gene Isolated from Hordeum Vulgare Is Differentially Regulated by Drought Stress. Theor. Appl. Genet. 2006, 113, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.S.; Jeong, H.I.; Bang, S.W.; Jung, S.E.; Kim, G.; Kim, Y.S.; Redillas, M.C.F.R.; Oh, S.-J.; Seo, J.S.; Kim, J.-K. DROUGHT-INDUCED BRANCHED-CHAIN AMINO ACID AMINOTRANSFERASE Enhances Drought Tolerance in Rice. Plant Physiol. 2023, 191, 1435–1447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-F.; Gong, Z.-H.; Wu, M.-B.; Chan, H.; Yuan, Y.-J.; Tang, N.; Zhang, Q.; Miao, M.-J.; Chang, W.; Li, Z.; et al. Integrative Comparative Analyses of Metabolite and Transcript Profiles Uncovers Complex Regulatory Network in Tomato (Solanum Lycopersicum L.) Fruit Undergoing Chilling Injury. Sci. Rep. 2019, 9, 4470. [Google Scholar] [CrossRef]
- Kissen, R.; Eberl, F.; Winge, P.; Uleberg, E.; Martinussen, I.; Bones, A.M. Effect of Growth Temperature on Glucosinolate Profiles in Arabidopsis Thaliana Accessions. Phytochemistry 2016, 130, 106–118. [Google Scholar] [CrossRef]
- Steindal, A.L.H.; Rødven, R.; Hansen, E.; Mølmann, J. Effects of Photoperiod, Growth Temperature and Cold Acclimatisation on Glucosinolates, Sugars and Fatty Acids in Kale. Food Chem. 2015, 174, 44–51. [Google Scholar] [CrossRef]
- Zang, Y.; Ge, J.; Huang, L.; Gao, F.; Lv, X.; Zheng, W.; Hong, S.; Zhu, Z. Leaf and Root Glucosinolate Profiles of Chinese Cabbage (Brassica Rapa Ssp. Pekinensis) as a Systemic Response to Methyl Jasmonate and Salicylic Acid Elicitation. J. Zhejiang Univ.-Sci. B 2015, 16, 696–708. [Google Scholar] [CrossRef]
- Dong, C.-H.; Zolman, B.K.; Bartel, B.; Lee, B.; Stevenson, B.; Agarwal, M.; Zhu, J.-K. Disruption of Arabidopsis CHY1 Reveals an Important Role of Metabolic Status in Plant Cold Stress Signaling. Mol. Plant 2009, 2, 59–72. [Google Scholar] [CrossRef]
- Gipson, A.B.; Morton, K.J.; Rhee, R.J.; Simo, S.; Clayton, J.A.; Perrett, M.E.; Binkley, C.G.; Jensen, E.L.; Oakes, D.L.; Rouhier, M.F.; et al. Disruptions in Valine Degradation Affect Seed Development and Germination in Arabidopsis. Plant J. 2017, 90, 1029–1039. [Google Scholar] [CrossRef]
- Zamani, M.; diCenzo, G.C.; Milunovic, B.; Finan, T.M. A Putative 3-hydroxyisobutyryl-CoA Hydrolase Is Required for Efficient Symbiotic Nitrogen Fixation in Sinorhizobium Meliloti and Sinorhizobium Fredii NGR234. Environ. Microbiol. 2017, 19, 218–236. [Google Scholar] [CrossRef]
- Yang, Y.; Saand, M.A.; Abdelaal, W.B.; Zhang, J.; Wu, Y.; Li, J.; Fan, H.; Wang, F. ITRAQ-Based Comparative Proteomic Analysis of Two Coconut Varieties Reveals Aromatic Coconut Cold-Sensitive in Response to Low Temperature. J. Proteomics 2020, 220, 103766. [Google Scholar] [CrossRef]
- Kobayashi, M.; Takada, Y. Effects of the Combined Substitutions of Amino Acid Residues on Thermal Properties of Cold-Adapted Monomeric Isocitrate Dehydrogenases from Psychrophilic Bacteria. Extremophiles 2014, 18, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Yuan, Y.; Zhang, C.; Ou, J.; Zhou, W.; Lin, Q. Modulation of Key Enzymes Involved in Ammonium Assimilation and Carbon Metabolism by Low Temperature in Rice (Oryza Sativa L.) Roots. Plant Sci. 2005, 169, 295–302. [Google Scholar] [CrossRef]
- Cui, X.; Zheng, Y.; Lu, Y.; Issakidis-Bourguet, E.; Zhou, D.-X. Metabolic Control of Histone Demethylase Activity Involved in Plant Response to High Temperature. Plant Physiol. 2021, 185, 1813–1828. [Google Scholar] [CrossRef] [PubMed]
- Arunraj, R.; Skori, L.; Kumar, A.; Hickerson, N.M.N.; Shoma, N.M.V.; Samuel, M.A. Spatial Regulation of Alpha-Galactosidase Activity and Its Influence on Raffinose Family Oligosaccharides during Seed Maturation and Germination in Cicer Arietinum. Plant Signal Behav. 2020, 15, 1709707. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Vepachedu, R.; Stushnoff, C.; Jones, M.L. Expression of an α-Galactosidase Gene in Petunia Is Upregulated during Low-Temperature Deacclimation. J. Am. Soc. Hortic. Sci. 2004, 129, 491–496. [Google Scholar] [CrossRef]
- González-Torralva, F.; Norsworthy, J.K. Overexpression of Acetyl CoA Carboxylase 1 and 3 (ACCase1 and ACCase3), and CYP81A21 Were Related to Cyhalofop Resistance in a Barnyardgrass Accession from Arkansas. Plant Signal Behav. 2023, 18, 2172517. [Google Scholar] [CrossRef]
- Khan, S.; Alvi, A.F.; Saify, S.; Iqbal, N.; Khan, N.A. The Ethylene Biosynthetic Enzymes, 1-Aminocyclopropane-1-Carboxylate (ACC) Synthase (ACS) and ACC Oxidase (ACO): The Less Explored Players in Abiotic Stress Tolerance. Biomolecules 2024, 14, 90. [Google Scholar] [CrossRef]
- Mei, J.; Yang, S.; Linghu, Y.; Gao, Y.; Hu, Y.; Nie, W.; Zhang, Y.; Peng, L.; Wu, Y.; Ding, Y.; et al. Unveiling the Role of MicroRNAs in Nonhost Resistance to Sclerotinia Sclerotiorum: Rice-specific MicroRNAs Attack the Pathogen via Cross-kingdom RNAi. J. Integr. Plant Biol. 2025, 67, 1179–1195. [Google Scholar] [CrossRef]
- Siodmak, A.; Martinez-Seidel, F.; Rayapuram, N.; Bazin, J.; Alhoraibi, H.; Gentry-Torfer, D.; Tabassum, N.; Sheikh, A.H.; Kise, J.K.G.; Blilou, I.; et al. Dynamics of Ribosome Composition and Ribosomal Protein Phosphorylation in Immune Signaling in Arabidopsis Thaliana. Nucleic Acids Res. 2023, 51, 11876–11892. [Google Scholar] [CrossRef]
- Yuan, L.; Dang, J.; Zhang, J.; Wang, L.; Zheng, H.; Li, G.; Li, J.; Zhou, F.; Khan, A.; Zhang, Z.; et al. A Glutathione S-Transferase Regulates Lignin Biosynthesis and Enhances Salt Tolerance in Tomato. Plant Physiol. 2024, 196, 2989–3006. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING V10: Protein–Protein Interaction Networks, Integrated over the Tree of Life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.-Q.; Liang, Y.-J.; Song, X.-P.; Yan, M.-X.; Tang, L.-Q.; Qin, Z.-Q.; Huang, Y.-X.; Li, D.-W.; Huang, D.-M.; Shi, Z.-S.; et al. Indole-3-Butyric Acid Enhances Root Formation and Alleviates Low-Temperature Stress in Sugarcane: Molecular Insights and Identification of Candidate Genes. Plants 2025, 14, 1502. https://doi.org/10.3390/plants14101502
Zhang X-Q, Liang Y-J, Song X-P, Yan M-X, Tang L-Q, Qin Z-Q, Huang Y-X, Li D-W, Huang D-M, Shi Z-S, et al. Indole-3-Butyric Acid Enhances Root Formation and Alleviates Low-Temperature Stress in Sugarcane: Molecular Insights and Identification of Candidate Genes. Plants. 2025; 14(10):1502. https://doi.org/10.3390/plants14101502
Chicago/Turabian StyleZhang, Xiao-Qiu, Yong-Jian Liang, Xiu-Peng Song, Mei-Xin Yan, Li-Qiu Tang, Zhen-Qiang Qin, Yu-Xin Huang, De-Wei Li, Dong-Mei Huang, Ze-Sheng Shi, and et al. 2025. "Indole-3-Butyric Acid Enhances Root Formation and Alleviates Low-Temperature Stress in Sugarcane: Molecular Insights and Identification of Candidate Genes" Plants 14, no. 10: 1502. https://doi.org/10.3390/plants14101502
APA StyleZhang, X.-Q., Liang, Y.-J., Song, X.-P., Yan, M.-X., Tang, L.-Q., Qin, Z.-Q., Huang, Y.-X., Li, D.-W., Huang, D.-M., Shi, Z.-S., Zhang, B.-Q., & Huang, D.-L. (2025). Indole-3-Butyric Acid Enhances Root Formation and Alleviates Low-Temperature Stress in Sugarcane: Molecular Insights and Identification of Candidate Genes. Plants, 14(10), 1502. https://doi.org/10.3390/plants14101502