
Enhancing Transplanting Success in Restoration of Degraded Areas Using Peat-Free Substrates

 ,
,  ,
,  ,
,  , ,
, , 
Abstract

1. Introduction
2. Results
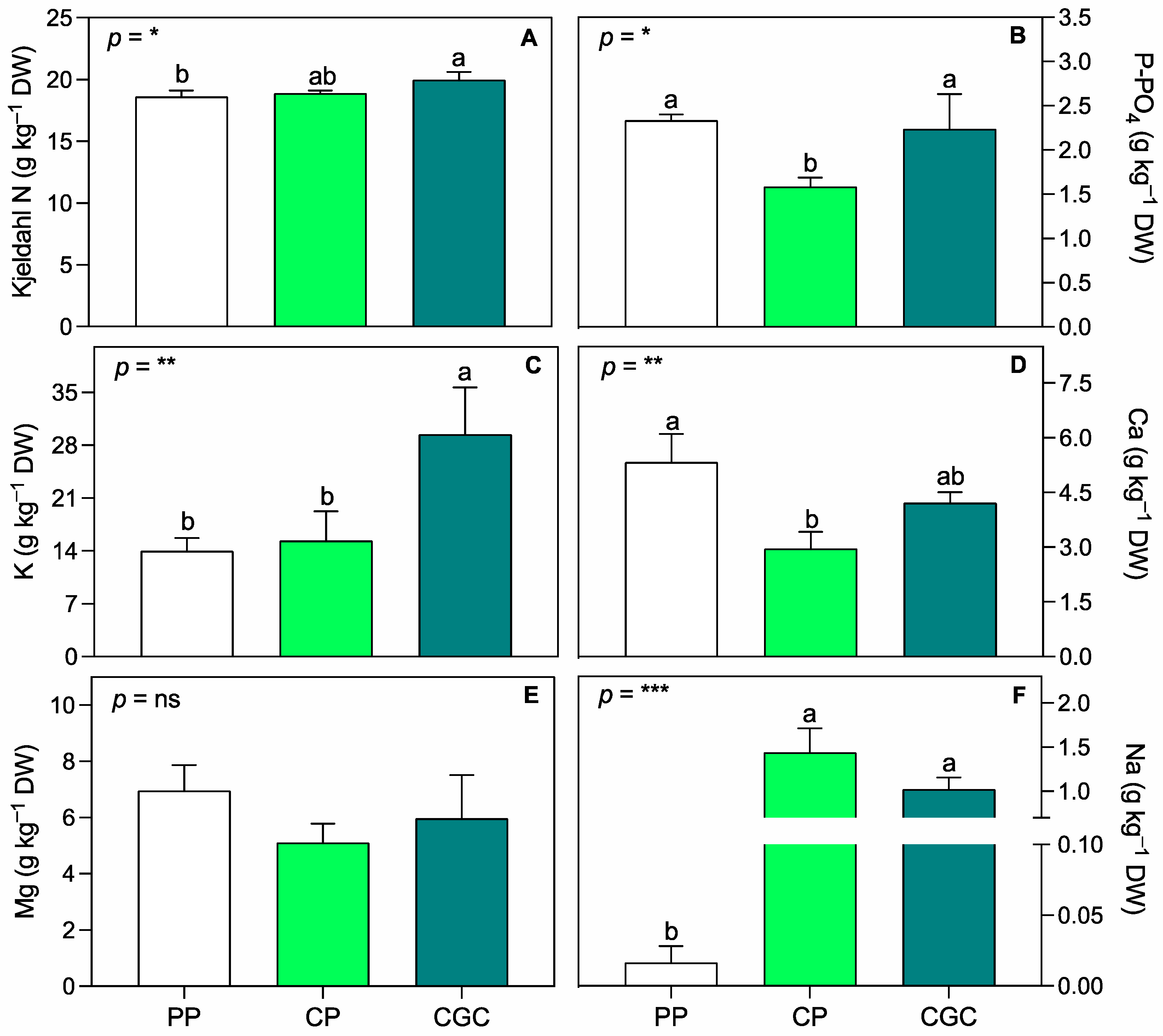
2.1. Cultivation Trial
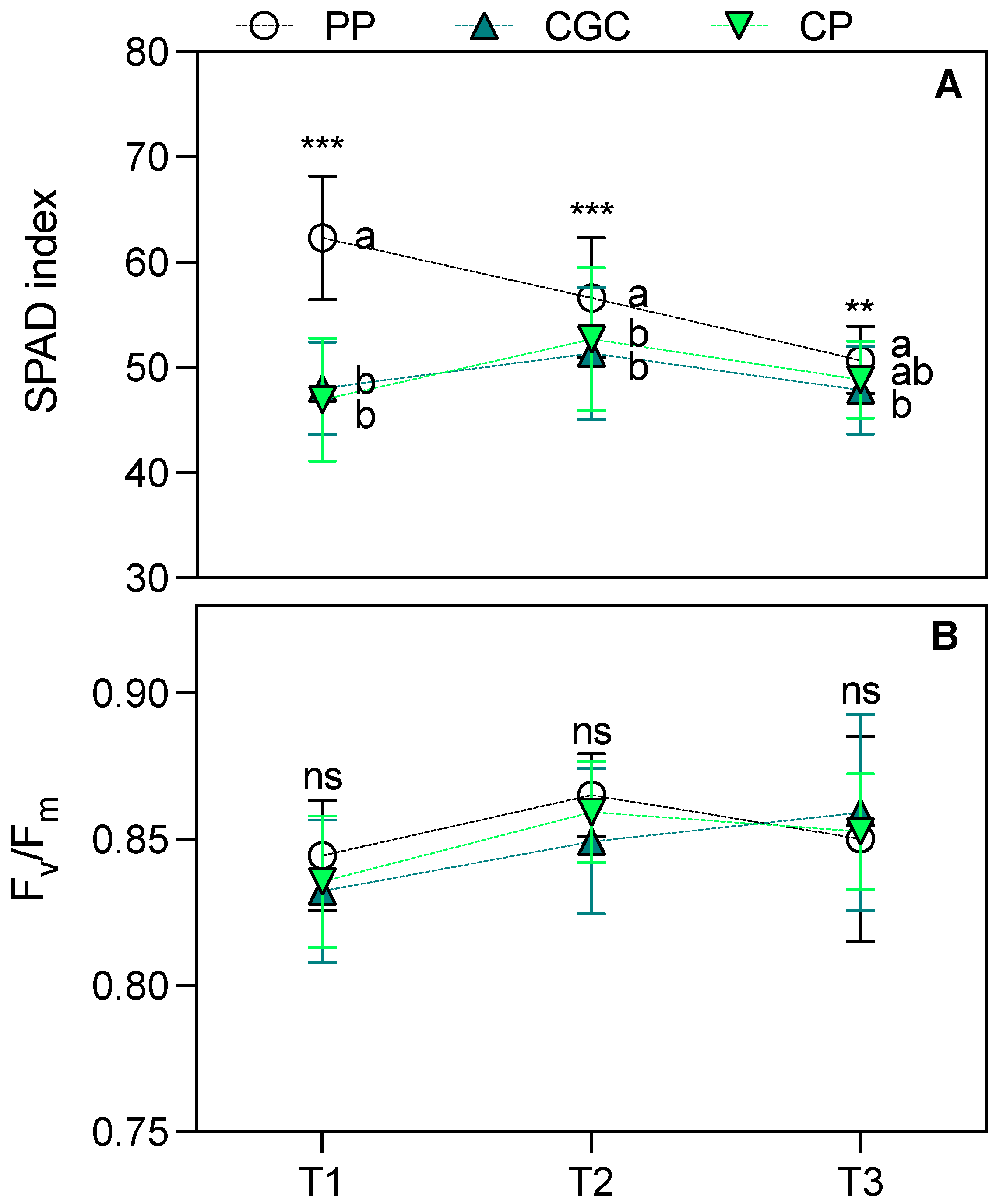
2.2. Plant Performances After Transplanting Trial in a Degraded Area
3. Discussion
4. Materials and Methods
4.1. Cultivation Trial: Growing Condition and Experimental Design
4.2. Transplanting Trial in a Degraded Area
4.3. Biometric, Eco-Physiological, and Nutrient Analyses
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Lonardo, S.; Baronti, S.; Vaccari, F.P.; Albanese, L.; Battista, P.; Miglietta, F.; Bacci, L. Biochar-based nursery substrates: The effect of peat substitution on reduced salinity. Urban For. Urban Green. 2017, 23, 27–34. [Google Scholar] [CrossRef]
- Ugolini, F.; Calzolari, C.; Lanini, G.M.; Massetti, L.; Pollaki, S.; Raschi, A.; Sabatini, F.; Tagliaferri, G.; Ungaro, F.; Massa, D.; et al. Testing decontaminated sediments as a substrate for ornamentals in field nursery plantations. J. Environ. Manag. 2017, 197, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, B.; Debode, J.; Willekens, K.; Van Delm, T. Recycling of P and K in circular horticulture through compost application in sustainable growing media for fertigated strawberry cultivation. Eur. J. Agron. 2018, 96, 131–145. [Google Scholar] [CrossRef]
- Massa, D.; Bonetti, A.; Cacini, S.; Faraloni, C.; Prisa, D.; Tuccio, L.; Petruccelli, R. Soilless tomato grown under nutritional stress increases green biomass but not yield or quality in presence of biochar as growing medium. Hortic. Environ. Biotechnol. 2019, 60, 871–881. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Younis, A.; Chen, J. Biochar or Biochar-Compost Amendment to a Peat-Based Substrate Improves Growth of Syngonium podophyllum. Agronomy 2019, 9, 460. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of organic amendments used in containerized plant production: Part 2—Non-compost-based amendments. Sci. Hortic. 2020, 260, 108855. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of organic amendments used in containerized plant production: Part 1—Compost-based amendments. Sci. Hortic. 2020, 266, 108856. [Google Scholar] [CrossRef]
- Blok, C.; Eveleens, B.; van Winkel, A. Growing media for food and quality of life in the period 2020–2050. Acta Hortic. 2021, 1305, 341–356. [Google Scholar] [CrossRef]
- Di Lonardo, S.; Sodini, M.; Massa, D.; Nesi, B.; Orsenigo, S.; Zubani, L.; Cacini, S. The effect of different peat-free growing media and fertilization levels on the plant nutrition of Leucanthemum vulgare Lam. and Dianthus barbatus L. J. Plant Nutr. 2025, 48, 114–129. [Google Scholar] [CrossRef]
- Atzori, G.; Pane, C.; Zaccardelli, M.; Cacini, S.; Massa, D. The role of peat-free organic substrates in the sustainable management of soilless cultivations. Agronomy 2021, 11, 1236. [Google Scholar] [CrossRef]
- Toboso-Chavero, S.; Madrid-López, C.; Villalba, G.; Gabarrell Durany, X.; Hückstädt, A.B.; Finkbeiner, M.; Lehmann, A. Environmental and social life cycle assessment of growing media for urban rooftop farming. Int. J. Life Cycle Assess. 2021, 26, 2085–2102. [Google Scholar] [CrossRef]
- Young, R.E.; Gann, G.D.; Walder, B.; Liu, J.; Cui, W.; Newton, V.; Nelson, C.R.; Tashe, N.; Jasper, D.; Silveira, F.A.O.; et al. International principles and standards for the ecological restoration and recovery of mine sites. Restor. Ecol. 2022, 30, e13771. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Lazzeri, V.; Leotta, L.; Bretzel, F. How can plants used for ornamental purposes contribute to urban biodiversity? Sustainability 2025, 17, 4061. [Google Scholar] [CrossRef]
- Meli, P.; Holl, K.D.; Rey Benayas, J.M.; Jones, H.P.; Jones, P.C.; Montoya, D.; Mateos, D.M. A global review of past land use, climate, and active vs. passive restoration effects on forest recovery. PLoS ONE 2017, 12, e0171368. [Google Scholar] [CrossRef] [PubMed]
- Clewell, A.F.; Aronson, J. Ecological Restoration: Principles, Values, and Structure of an Emerging Profession; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Gómez-Aparicio, L. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 2009, 97, 1202–1214. [Google Scholar] [CrossRef]
- Tang, Y.; Shen, Y.; Feng, H.; Wu, H.; Mao, R.; Ai, W.; Wu, Z. Study on primary physicochemical characteristics and nutrient adsorption of four plant cultivation substrates. Life Sci. Space Res. 2023, 36, 78–85. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, T.; Li, J.; Zhou, H.; Zhang, H. Study on the main physicochemical characteristics of different plant cultivation substrates and their effects on standard roses. Plant Soil Environ. 2024, 70, 799–808. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; Erickson, T.E.; Martini, D.; Dixon, K.W.; Merritt, D.J. Soil physicochemical and microbiological indicators of short, medium and long term post-fire recovery in semi-arid ecosystems. Ecol. Indic. 2016, 63, 14–22. [Google Scholar] [CrossRef]
- Scotti, R.; Bonanomi, G.; Scelza, R.; Zoina, A.; Rao, M.A. Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J. Soil Sci. Plant Nutr. 2015, 15, 333–352. [Google Scholar] [CrossRef]
- Bonanomi, G.; Lorito, M.; Vinale, F.; Woo, S.L. Organic amendments, beneficial microbes, and soil microbiota: Toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 2018, 56, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, B.; Van Loo, K.; Ommeslag, S.; Vierendeels, S.; Rooseleer, M.; Vandaele, E. Sustainable growing media blends with woody green composts: Optimizing the n release with organic fertilizers and interaction with microbial biomass. Agronomy 2022, 12, 422. [Google Scholar] [CrossRef]
- Gruda, N. Increasing Sustainability of Growing Media Constituents and Stand-Alone Substrates in Soilless Culture Systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef]
- Mariotti, B.; Martini, S.; Raddi, S.; Tani, A.; Jacobs, D.F.; Oliet, J.A.; Maltoni, A. Coconut Coir as a Sustainable Nursery Growing Media for Seedling Production of the Ecologically Diverse Quercus Species. Forests 2020, 11, 522. [Google Scholar] [CrossRef]
- Eveleens, B.; van Winkel, A.; Blok, C. Wood fiber in pot plant culture; peat replacement up to 50% in volume? Acta Hortic. 2021, 1317, 165–174. [Google Scholar] [CrossRef]
- Hillova, D.; Takacsova, M.; Lichtnerova, H. Stomatal response to water stress in herbaceous perennials. In Plants Urban Areas Landscape; Slovak University of Agriculture: Nitra, Greece, 2014; pp. 52–56. [Google Scholar] [CrossRef]
- Nazemi Rafi, Z.; Kazemi, F.; Tehranifar, A. Effects of various irrigation regimes on water use efficiency and visual quality of some ornamental herbaceous plants in the field. Agric. Water Manag. 2019, 212, 78–87. [Google Scholar] [CrossRef]
- Mircea, D.-M.; Boscaiu, M.; Sestras, R.E.; Sestras, A.F.; Vicente, O. Abiotic Stress Tolerance and Invasive Potential of Ornamental Plants in the Mediterranean Area: Implications for Sustainable Landscaping. Agronomy 2024, 15, 52. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Ortuño, M.F.; Bañon, S.; Álvarez, S. Deficit irrigation as a strategy to control growth in ornamental plants and enhance their ability to adapt to drought conditions. J. Hortic. Sci. Biotechnol. 2019, 94, 137–150. [Google Scholar] [CrossRef]
- Marenco, R.A.; Antezana-Vera, S.A.; Nascimento, H.C.S. Relationship between specific leaf area, leaf thickness, leaf water content and SPAD-502 readings in six Amazonian tree species. Photosynthetica 2009, 47, 184–190. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Zuo, G. Non-photochemical quenching (NPQ) in photoprotection: Insights into NPQ levels required to avoid photoinactivation and photoinhibition. New Phytol. 2025, 246, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Ridolfi, M.; Roupsard, O.; Garrec, J.; Dreyer, E. Effects of a calcium deficiency on stomatal conductance and photosynthetic activity of Quercus robur seedlings grown on nutrient solution. Ann. Des Sci. For. 1996, 53, 325–335. [Google Scholar] [CrossRef]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.-J.; Iba, K.; Schroeder, J.I. CO2 Sensing and CO2 Regulation of Stomatal Conductance: Advances and Open Questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.C.; Braga, R.P. Estimation of reference evapotranspiration during the irrigation season using nine temperature-based methods in a hot-summer Mediterranean climate. Agriculture 2021, 11, 124. [Google Scholar] [CrossRef]
- Cai, X.; Starman, T.; Niu, G.; Hall, C.; Lombardini, L. Response of Selected Garden Roses to Drought Stress. HortScience 2012, 47, 1050–1055. [Google Scholar] [CrossRef]
- Caser, M.; Lovisolo, C.; Scariot, V. The influence of water stress on growth, ecophysiology and ornamental quality of potted Primula vulgaris ‘Heidy’ plants. New insights to increase water use efficiency in plant production. Plant Growth Regul. 2017, 83, 361–373. [Google Scholar] [CrossRef]
- Liu, B.; Liang, J.; Tang, G.; Wang, X.; Liu, F.; Zhao, D. Drought stress affects on growth, water use efficiency, gas exchange and chlorophyll fluorescence of Juglans rootstocks. Sci. Hortic. 2019, 250, 230–235. [Google Scholar] [CrossRef]
- Di Lonardo, S.; Massa, D.; Orsenigo, S.; Zubani, L.; Rossi, G.; Fascella, G.; Cacini, S. Substitution of peat in the cultivation of two shrub species used for ecological restoration and recovery of degraded green areas. Acta Hortic. 2021, 1305, 97–102. [Google Scholar] [CrossRef]
- Di Lonardo, S.; Cacini, S.; Becucci, L.; Lenzi, A.; Orsenigo, S.; Zubani, L.; Rossi, G.; Zaccheo, P.; Massa, D. Testing new peat-free substrate mixtures for the cultivation of perennial herbaceous species: A case study on Leucanthemum vulgare Lam. Sci. Hortic. 2021, 289, 110472. [Google Scholar] [CrossRef]
- Xie, K.; Lu, Z.; Pan, Y.; Gao, L.; Hu, P.; Wang, M.; Guo, S. Leaf photosynthesis is mediated by the coordination of nitrogen and potassium: The importance of anatomical-determined mesophyll conductance to CO2 and carboxylation capacity. Plant Sci. 2020, 290, 110267. [Google Scholar] [CrossRef]
- Sanchez, C.A. Phosphorus. In Handbook of Plant Nutrition; CRC Press: Boca Raton, FL, USA, 2007; pp. 51–90. [Google Scholar]
- Taylor, M.D.; Locascio, S.J. Blossom-End Rot: A Calcium Deficiency. J. Plant Nutr. 2004, 27, 123–139. [Google Scholar] [CrossRef]
- Gorham, J. Sodium. In Handbook of Plant Nutrition; CRC Press: Boca Raton, FL, USA, 2007; pp. 585–600. [Google Scholar]
- Allen, K.S.; Harper, R.W.; Bayer, A.; Brazee, N.J. A review of nursery production systems and their influence on urban tree survival. Urban For. Urban Green. 2017, 21, 183–191. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, K.R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata [L.] Walp.) under drought. J. Photochem. Photobiol. B Biol. 2011, 105, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Pirasteh-Anosheh, H.; Saed-Moucheshi, A.; Pakniyat, H.; Pessarakli, M. Stomatal Responses to Drought Stress. In Water Stress and Crop Plants; Ahmad, P., Ed.; Wiley: Hoboken, NJ, USA, 2016; pp. 24–40. [Google Scholar] [CrossRef]
- Hentschel, R.; Hommel, R.; Poschenrieder, W.; Grote, R.; Holst, J.; Biernath, C.; Gessler, A.; Priesack, E. Stomatal conductance and intrinsic water use efficiency in the drought year 2003: A case study of European beech. Trees 2016, 30, 153–174. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Milinković, M.; Lalević, B.; Jovičić-Petrović, J.; Golubović-Ćurguz, V.; Kljujev, I.; Raičević, V. Biopotential of compost and compost products derived from horticultural waste—Effect on plant growth and plant pathogens’ suppression. Process Saf. Environ. Prot. 2019, 121, 299–306. [Google Scholar] [CrossRef]
- Aydi, S.; Sassi Aydi, S.; Rahmani, R.; Bouaziz, F.; Souchard, J.P.; Merah, O.; Abdelly, C. Date-palm compost as soilless substrate improves plant growth, photosynthesis, yield and phytochemical quality of greenhouse melon (Cucumis melo L.). Agronomy 2023, 13, 212. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, X.; Chen, L.; Jia, G. Whole-plant instantaneous and short-term water-use efficiency in response to soil water content and CO2 concentration. Plant Soil 2019, 444, 281–298. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.-M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef]
- Massa, D.; Prisa, D.; Montoneri, E.; Battaglini, D.; Ginepro, M.; Negre, M.; Burchi, G. Application of municipal biowaste derived products in Hibiscus cultivation: Effect on leaf gaseous exchange activity and plant biomass accumulation and quality. Sci. Hortic. 2016, 205, 59–69. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Plant Height | Collar Diameter | Shoot FW | Shoot DW FW−1 | Leaf DW | SLA | Root/Shoot |
|---|---|---|---|---|---|---|---|
| cm Plant−1 | mm Plant−1 | g Plant−1 | % | g Plant−1 | cm2 g DW−1 | ||
| PP | 90.0 ± 17.9 | 22.1 ± 5.5 | 365.4 ± 86.2 | 35.9 ± 1.5 a | 59.8 ± 16.7 | 81.1 ± 11.6 c | 0.24 ± 0.05 |
| CP | 90.5 ± 15.3 | 19.3 ± 4.7 | 423.7 ± 154.5 | 35.3 ± 3.0 ab | 70.4 ± 27.3 | 113.0 ± 13.3 b | 0.27 ± 0.06 |
| CGC | 92.8 ± 17.9 | 18.4 ± 4.0 | 403.6 ± 62.3 | 33.4 ± 1.5 b | 59.1 ± 16.0 | 141.8 ± 28.4 a | 0.32 ± 0.13 |
| ANOVA | ns | ns | ns | * | ns | *** | ns |
| Substrate | Leaf FW | Shoot DW | Shoot DW FW−1 | Necrotic LA | Stem DW Shoot DW−1 |
|---|---|---|---|---|---|
| g Plant−1 | g Plant−1 | % Plant−1 | % Plant−1 | % Plant−1 | |
| PP | 36.6 ± 18.4 b | 80.3 ± 20.9 | 46.1 ± 3.0 | 14.6 ± 10.7 a | 83.1 ± 12.0 a |
| CP | 56.8 ± 20.1 ab | 98.1 ± 35.4 | 44.7 ± 3.7 | 16.6 ± 8.9 a | 76.9 ± 8.7 ab |
| CGC | 66.6 ± 21.6 a | 89.4 ± 20.2 | 44.9 ± 2.9 | 4.8 ± 1.6 b | 72.2 ± 7.9 b |
| ANOVA | * | ns | ns | *** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Traversari, S.; Di Lonardo, S.; Orsenigo, S.; Massa, D.; Nesi, B.; Zubani, L.; Cacini, S. Enhancing Transplanting Success in Restoration of Degraded Areas Using Peat-Free Substrates. Plants 2025, 14, 1450. https://doi.org/10.3390/plants14101450
Traversari S, Di Lonardo S, Orsenigo S, Massa D, Nesi B, Zubani L, Cacini S. Enhancing Transplanting Success in Restoration of Degraded Areas Using Peat-Free Substrates. Plants. 2025; 14(10):1450. https://doi.org/10.3390/plants14101450
Chicago/Turabian StyleTraversari, Silvia, Sara Di Lonardo, Simone Orsenigo, Daniele Massa, Beatrice Nesi, Lino Zubani, and Sonia Cacini. 2025. "Enhancing Transplanting Success in Restoration of Degraded Areas Using Peat-Free Substrates" Plants 14, no. 10: 1450. https://doi.org/10.3390/plants14101450
APA StyleTraversari, S., Di Lonardo, S., Orsenigo, S., Massa, D., Nesi, B., Zubani, L., & Cacini, S. (2025). Enhancing Transplanting Success in Restoration of Degraded Areas Using Peat-Free Substrates. Plants, 14(10), 1450. https://doi.org/10.3390/plants14101450