

Simulated Warming Reduces Biomass Accumulation in Zizania caduciflor and Sparganium stoloniferum
Abstract
1. Introduction
2. Results
2.1. Differences in Functional Traits of Z. caduciflora and S. stoloniferum Under Three Different Growth Temperatures
2.2. The Relationship Between Z. caduciflora and S. stoloniferum Functional Traits and Environmental Factors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Traits | Mean Annual Temperature/MAT | Annual Maximum Temperature/Max | ||
|---|---|---|---|---|
| Z. caduciflora | S. stoloniferum | Z. caduciflora | S. stoloniferum | |
| net photosynthetic rate | −0.852 ** | 0.315 | −0.937 *** | 0.515 |
| stomatal conductance | −0.828 ** | 0.278 | −0.937 *** | 0.490 |
| transpiration rate | −0.586 | – | −0.743 * | – |
| intercellular CO2 concentration | – | – | – | – |
| aboveground biomass | −0.727 * | – | −0.523 | – |
| interval biomass | −0.908 *** | −0.638 | −0.832 ** | −0.826 ** |
| fibrous root biomass | – | −0.437 | – | −0.705 * |
| root biomass | – | −0.406 | – | −0.641 |
| bud biomass | −0.960 *** | −0.878 ** | −0.888 *** | −0.967 ** |
| underground biomass | −0.893 *** | −0.521 | −0.820 ** | −0.761 * |
| total biomass | −0.862 ** | −0.456 | −0.724 * | −0.710 * |
| aboveground carbon comtent | −0.043 | – | 0.184 | – |
| interval carbon comtent | – | – | – | – |
| fibrous root carbon comtent | −0.558 | 0.676 * | −0.259 | 0.821 ** |
| root carbon comtent | – | – | – | – |
| bud carbon comtent | 0.898 *** | – | 0.989 *** | – |
| aboveground nitrogen comtent | −0.822 ** | −0.229 | −0.904 *** | −0.509 |
| interval nitrogen comtent | – | −0.570 | – | −0.369 |
| fibrous root nitrogen comtent | – | −0.117 | – | 0.226 |
| root nitrogen comtent | – | −0.479 | – | −0.679 * |
| bud nitrogen comtent | −0.828 ** | 0.877 ** | −0.565 | 0.666 |
| aboveground phosphorus comtent | – | – | – | – |
| interval phosphorus comtent | – | – | – | – |
| fibrous root phosphorus comtent | – | 0.201 | – | 0.493 |
| root phosphorus comtent | – | – | – | – |
| bud phosphorus comtent | – | 0.444 | – | 0.740 * |
3. Materials and Methods
3.1. Overview of the Study Area
3.2. Experimental Design
3.2.1. Construction of Artificial Simulated Warming Chambers
| Process | Temperature Variable | |||||
|---|---|---|---|---|---|---|
| MAT | Max | Min | Sat | Dat | Nat | |
| CK | 8.83 | 37.69 | −22.38 | 15.12 | 2572.79 | 281.66 |
| ET–2 | 11.00 | 46.88 | −19.13 | 17.76 | 3319.56 | 470.34 |
| ET–4 | 12.59 | 47.50 | −16.75 | 19.09 | 3877.70 | 562.04 |
3.2.2. Plant Transplantation
3.3. Measurement of Trait Parameters
3.3.1. Measurement of Photosynthetic Parameters
3.3.2. Determination of Biomass and Elemental Content
| Functional Traits | Abbreviations (Unit) | |
|---|---|---|
| Photosynthetic parameters | net photosynthetic rate | Pn (μmol·m−2·s−1) |
| stomatal conductance | Gs (mol·m−2·s−1) | |
| transpiration rate | Tr (μmol·mol−1) | |
| intercellular CO2 concentration | Ci (mmol·m−2·s−1) | |
| Biomass | aboveground biomass | Bioabove (g·m−2) |
| interval biomass | Biointer (g·m−2) | |
| fibrous root biomass | Biofib (g·m−2) | |
| root biomass | Bioroot (g·m−2) | |
| bud biomass | Biobud (g·m−2) | |
| underground biomass | Biounder (g·m−2) | |
| total biomass | Biototal (g·m−2) | |
| Elemental content | aboveground carbon comtent | Cabove (g·kg−1) |
| interval carbon comtent | Cinter (g·kg−1) | |
| fibrous root carbon comtent | Cfib (g·kg−1) | |
| root carbon comtent | Croot (g·kg−1) | |
| bud carbon comtent | Cbud (g·kg−1) | |
| aboveground nitrogen comtent | Nabove (g·kg−1) | |
| interval nitrogen comtent | Ninter (g·kg−1) | |
| fibrous root nitrogen comtent | Nfib (g·kg−1) | |
| root nitrogen comtent | Nroot (g·kg−1) | |
| bud nitrogen comtent | Nbud (g·kg−1) | |
| aboveground phosphorus comtent | Pabove (g·kg−1) | |
| interval phosphorus comtent | Pinter (g·kg−1) | |
| fibrous root phosphorus comtent | Pfib (g·kg−1) | |
| root phosphorus comtent | Proot (g·kg−1) | |
| bud phosphorus comtent | Pbud (g·kg−1) | |
3.4. Data Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harrison, S. Plant community diversity will decline more than increase under climatic warming. Philos. Trans. R. Soc. B 2020, 375, 20190106. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Oxford, UK, 2014. [Google Scholar]
- Li, Y.; Yu, X.B.; Liu, Y.; Zhang, G.S.; Zhang, Q.J.; Duan, H.L. Response of wetland plant functional traits to hydrological processes: A review. Chin. J. Ecol. 2018, 37, 952–959. [Google Scholar]
- Yin, L.; Dai, E.; Zheng, D.; Wang, Y.; Ma, L.; Tong, M. What drives the vegetation dynamics in the Hengduan Mountain region, southwest China: Climate change or human activity? Ecol. Indic. 2020, 112, 106013. [Google Scholar] [CrossRef]
- Kuang, X.; Jiao, J.J. Review on climate change on the Tibetan Plateau during the last half century. J. Geophys. Res. Atmos. 2016, 121, 3979–4007. [Google Scholar] [CrossRef]
- Gopal, B. Future of wetlands in tropical and subtropical Asia, especially in the face of climate change. Aquat. Sci. 2013, 75, 39–61. [Google Scholar] [CrossRef]
- Mao, X.; Wei, X.; Engel, B.; Wei, X.; Zhang, Z.; Tao, Y.; Wang, W. Network–based perspective on water–air interface GHGs flux on a cascade surface–flow constructed wetland in Qinghai–Tibet Plateau, China. Ecol. Eng. 2020, 151, 105862. [Google Scholar] [CrossRef]
- Fu, W.C.; Tian, K.; Xiao, D.R.; Li, W.; Yue, H.T.; Zhao, X.J.; Yang, H. The ecological restoration effort of degraded estuarine wetland in Northwest Yunnan Plateau, China. Acta Ecol. Sin. 2014, 34, 2187–2194. [Google Scholar]
- Zhang, Y.; Yan, J.; Cheng, X.; He, X. Wetland changes and their relation to climate change in the Pumqu Basin, Tibetan Plateau. Int. J. Environ. Res. Public Health 2021, 18, 2682. [Google Scholar] [CrossRef]
- Liu, G.; Sun, J.; Tian, K.; Xiao, D.; Yuan, X. Long–term responses of leaf litter decomposition to temperature, litter quality and litter mixing in plateau wetlands. Freshw. Biol. 2017, 62, 178–190. [Google Scholar] [CrossRef]
- Du, J.H.; Wang, Y.; Pan, Y.; Wang, J. Ecological risk assessment and ecological security pattern construction of county–level landscape in plateau valley: A case study in Yongping County, Dali Prefecture, Yunnan Province. Chin. J. Ecol. 2024, 43, 1509–1520. [Google Scholar]
- Xiao, D.R.; Zhang, C.; Tian, K.; Liu, G.; Yang, H.; An, S. Development of alpine wetland vegetation and its effect on carbon sequestration after dam construction: A case study of Lashihai in the northwestern Yunnan plateau in China. Aquat. Bot. 2015, 126, 16–24. [Google Scholar] [CrossRef]
- Walther, W.; Strauch, G.; Kersebaum, K.C.; Reinstorf, F.; Schäfer, W. About the knowledge in the field of nutrient metabolism in the drain zone and in groundwater and about their modelling. 2nd Part: The application of isotope tracer techniques and mathematical models for description of transport and turnover of plant nutrients in the water–saturated zone and the groundwater. Landnutz. Landentwickl. 2002, 43, 97–102. [Google Scholar]
- Erwin, J. Cacti: Studies on introducing a new group of ornamental plants. Acta Hortic. 2009, 813, 359–364. [Google Scholar] [CrossRef]
- Sun, M.; Feng, C.H.; Liu, Z.Y.; Tian, K. Evolutionary correlation of water–related traits between different structures of Dendrobium plants. Bot. Stud. 2020, 61, 16. [Google Scholar] [CrossRef]
- Chelli, S.; Marignani, M.; Barni, E.; Petraglia, A.; Puglielli, G.; Wellstein, C.; Acosta, A.; Bolpagni, R.; Bragazza, L.; Campetella, G.; et al. Plant–environment interactions through a functional traits perspective: A review of Italian studies. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2019, 153, 853–869. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, J.A.; Pineda, J.E.M.; Melgarejo, L.M.; Calderón, J.H.M. Functional traits of leaves and forest structure of neotropical mangroves under different salinity and nitrogen regimes. Flora 2018, 239, 52–61. [Google Scholar] [CrossRef]
- Bai, J.S.; Tang, H.R.; Lou, Y.J. Effects of water depth and nitrogen addition on functional traits of wetland plants: A review. Chin. J. Ecol. 2021, 40, 2987–2995. [Google Scholar]
- Soudzilovskaia, N.A.; Elumeeva, T.G.; Onipchenko, V.G.; Shidakov, I.I.; Salpagarova, F.S.; Khubiev, A.B. Functional traits predict relationship between plant abundance dynamic and long–term climate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18180–18184. [Google Scholar] [CrossRef]
- Moles, A.T.; Perkins, S.E.; Laffan, S.W.; Flores-Moreno, H.; Awasthy, M.; Tindall, M.L.; Sack, L.; Pitman, A.; Kattge, J.; Aarssen, L.W.; et al. Which is a better predictor of plant traits: Temperature or precipitation? J. Veg. Sci. 2014, 25, 1167–1180. [Google Scholar] [CrossRef]
- Bagley, J.; Rosenthal, D.M.; Ruiz-Vera, U.M.; Siebers, M.H.; Kumar, P.; Ort, D.R.; Bernacchi, C.J. The influence of photosynthetic acclimation to rising CO2 and warmer temperatures on leaf and canopy photosynthesis models. Glob. Biogeochem. Cycles 2015, 29, 194–206. [Google Scholar] [CrossRef]
- Zhang, C.C.; Han, G.; Guan, H.D.; Bao, C.H.; Zhang, X.P. Responses of optimal photosynthesis temperature to changes in ambient temperature for Cinnamomum camphora and Osmanthus fragrans. Chin. J. Ecol. 2014, 33, 2980–2987. [Google Scholar]
- Zhou, Y.Q.; Xu, Y.S.; He, L.X.; Zhang, Y.J.; Feng, Z.Z. Combined effects of elevated ozone concentration and waring on photosynthesis of rice leaves. Acta Ecol. Sin. 2025, 45, 1–12. [Google Scholar]
- Li, L.; Feng, S.D.; Wang, J.B.; Ni, H.W.; Fu, X.L.; Yang, L.Y.; Xu, M.H. Relationship between photosynthetic characteristics and leaf functional traits of 12 plant species of marshes in Sanjiang Plain. Wetl. Sci. 2010, 8, 225–232. [Google Scholar]
- Zhao, Y.; Sun, M.; Guo, H.; Feng, C.; Liu, Z.; Xu, J. Responses of leaf hydraulic traits of Schoenoplectus tabernaemontani to increasing temperature and CO2 concentrations. Bot. Stud. 2022, 63, 2. [Google Scholar] [CrossRef]
- Wright, I.J.; Groom, P.K.; Lamont, B.B.; Poot, P.; Prior, L.D.; Reich, P.B.; Schulze, E.-D.; Veneklaas, E.J.; Westoby, M. Leaf trait relationships in Australian plant species. Funct. Plant Biol. 2004, 31, 551–558. [Google Scholar] [CrossRef]
- Roche, P.; Díaz-Burlinson, N.; Gachet, S. Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? Plant Ecol. 2004, 174, 37–48. [Google Scholar] [CrossRef]
- Qian, Y.; Tian, K.; Xiao, D.; Yin, L.; Yu, D.; Yang, Y. Ecological restoration effect of closed and half–closed degraded wetlands in Northwest Yunnan Plateau, Southwest China. Yingyong Shengtai Xuebao 2012, 23, 1520–1526. [Google Scholar]
- Niu, M.; Guo, H.; Sun, M.; Xiao, D.R.; Liu, Z.Y.; Ai, J.; Zhao, P. Response of leaf morphological traits to environmental factors of an emergent aquatic plant Sparganium stoloniferum in northwest Yunnan. J. Southwest For. Univ. 2022, 42, 84–95. [Google Scholar]
- Wang, Z.B.; Tian, K.; Guan, D.X.; Li, H.; Zhang, Y.; Feng, C.; Sun, M. Responses of seed reproduction traits of a wetland dominant plant Schoenoplectus tabernaemontani to environmental changes in Hengduan Mountains. J. Northeast For. Univ. 2018, 46, 12–15. [Google Scholar]
- Liu, Z.Y.; Zhang, X.N.; Li, L.P.; Wang, H.; Zhang, Y.; Sun, M.; Xiao, D. Influence of simulated warming on light and CO2 utilization capacities of lakeside dominant plants in a typical plateau wetland in northwestern Yunnan. Acta Ecol. Sin. 2017, 37, 7821–7832. [Google Scholar]
- Xiao, D.R.; Tian, K.; Zhang, L.Q. Relationship between plant diversity and soil fertility in Napahai wetland of Northwestern Yunnan Plateau. Acta Ecol. Sin. 2008, 28, 3116–3124. [Google Scholar]
- Liu, Z.Y.; Zhao, Y.; Yu, H.Y.; Zhao, Y.; Guo, H.; Sun, M. Response of the functional traits of Schoenoplectus tabernaemontani to simulated warming in the Napahai wetland of northwestern Yunnan, China. Front. Ecol. Evol. 2024, 12, 1399584. [Google Scholar] [CrossRef]
- Yu, H.Y.; Sun, M.; Feng, C.H.; Xu, J.P.; Chen, H.Y.; Liu, Z.Y. Responses of leaf economic traits of Scirpus validus and Typha orientalis to simulated warming and CO2 concentration multiplication. Guihaia 2023, 43, 1588–1599. [Google Scholar]
- Kim, K.; Portis, A.R., Jr. Temperature dependence of photosynthesis in Arabidopsis plants with modifications in Rubisco activase and membrane fluidity. Plant Cell Physiol. 2005, 46, 522–530. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Robinson, S.P.; Portis, A.R., Jr. Adenosine triphosphate hydrolysis by purified Rubisco activase. Arch. Biochem. Biophys. 1989, 268, 93–99. [Google Scholar] [CrossRef]
- Kumar, A.; Li, C.; Portis, A.R. Arabidopsis thaliana expressing a thermostable chimeric Rubisco activase exhibits enhanced growth and higher rates of photosynthesis at moderately high temperatures. Photosynth. Res. 2009, 100, 143–153. [Google Scholar] [CrossRef]
- Ibañez, C.; Poeschl, Y.; Peterson, T.; Bellstädt, J.; Denk, K.; Gogol-Döring, A.; Marcel, Q.; Delker, C. Ambient temperature and genotype differentially affect developmental and phenotypic plasticity in Arabidopsis thaliana. BMC Plant Biol. 2017, 17, 114. [Google Scholar] [CrossRef]
- Frei, E.R.; Ghazoul, J.; Pluess, A.R. Plastic responses to elevated temperature in low and high elevation populations of three grassland species. PLoS ONE 2014, 9, e98677. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of wild and cultivated plants under global warming conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Huang, M.; Wang, W.Y.; Li, Z.H.; Zhang, T.; Ma, L.; Bai, Y.F.; Wang, Y.L.; Shi, J.J.; Long, R.J.; et al. Variation characteristics of plant community diversity and above–ground biomass in alpine degraded slopes along altitude gradients in the headwaters region of three–river on Tibetan plateau. Acta Ecol. Sin. 2022, 42, 3640–3655. [Google Scholar]
- Pan, Y.; Cieraad, E.; Armstrong, J.; Armstrong, W.; Clarkson, B.R.; Colmer, T.D.; Pedersen, O.; Visser, E.J.W.; Voesenek, L.A.C.J.; van Bodegom, P.M. Global patterns of the leaf economics spectrum in wetlands. Nat. Commun. 2020, 11, 4519. [Google Scholar] [CrossRef]
- Gardner, R.C.; Finlayson, C. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People; Ramsar Convention Secretariat: Gland, Switzerland, 2018. [Google Scholar]
- Sun, M.; Tian, K.; Yue, H.T.; Liu, Q.; Wang, Z.B. Correlated evolution between leaf nutrients and leaf biomass across epiphytic Dendrobium species. Ecol. Sci. 2019, 38, 1–8. [Google Scholar]
- Goraya, G.K.; Kaur, B.; Asthir, B.; Bala, S.; Kaur, G.; Farooq, M. Rapid injuries of high temperature in plants. J. Plant Biol. 2017, 60, 298–305. [Google Scholar] [CrossRef]
- Xalxo, R.; Yadu, B.; Chandra, J.; Chandrakar, V.; Keshavkant, S. Alteration in carbohydrate metabolism modulates thermotolerance of plant under heat stress. In Heat Stress Tolerance in Plants: Physiological, Molecular and Genetic Perspectives; Wiley: New York, NY, USA, 2020; pp. 77–115. [Google Scholar]
- Zahra, N.; Hafeez, M.B.; Ghaffar, A.; Kausar, A.; Al Zeidi, M.; Siddique, K.H.; Farooq, M. Plant photosynthesis under heat stress: Effects and management. Environ. Exp. Bot. 2023, 206, 105178. [Google Scholar] [CrossRef]
- Schreiber, U.; Berry, J.A. Heat–induced changes of chlorophyll fluorescence in intact leaves correlated with damage of the photosynthetic apparatus. Planta 1977, 136, 233–238. [Google Scholar] [CrossRef]
- Bilger, H.W.; Schreiber, U.; Lange, O.L. Determination of leaf heat resistance: Comparative investigation of chlorophyll fluorescence changes and tissue necrosis methods. Oecologia 1984, 63, 256–262. [Google Scholar] [CrossRef]
- Schymanski, S.J.; Or, D.; Zwieniecki, M. Stomatal control and leaf thermal and hydraulic capacitances under rapid environmental fluctuations. PLoS ONE 2013, 8, e54231. [Google Scholar] [CrossRef]
- Hueve, K.; Bichele, I.; Rasulov, B.; Niinemets, Ü.L.O. When it is too hot for photosynthesis: Heat–induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H2O2 formation. Plant Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef]
- Marchin, R.M.; Backes, D.; Ossola, A.; Leishman, M.R.; Tjoelker, M.G.; Ellsworth, D.S. Extreme heat increases stomatal conductance and drought–induced mortality risk in vulnerable plant species. Glob. Change Biol. 2022, 28, 1133–1146. [Google Scholar] [CrossRef] [PubMed]
- Gutschick, V.P. Leaf energy balance: Basics, and modeling from leaves to canopies. In Canopy Photosynthesis: From Basics to Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 23–58. [Google Scholar]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.H.; Cao, Z.; Li, X.G.; Scholten, T.; Kühn, P.; Wang, L.; Yu, R.P.; He, J.S. Soil phosphorus availability mediates the effects of nitrogen addition on community and species level phosphorus acquisition strategies in alpine grasslands. Sci. Total Environ. 2024, 906, 167630. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.L.; Zhang, S.H.; Wang, L.J.; Mao, Z.P.; Ma, X.Y. Numerical simulation of water purification effects of aquatic plant in artificial surface flow wetland. J. Hydraul. Eng. 2020, 51, 675–684. [Google Scholar]
- Tian, H.; Jin, H.T.; Sun, Q.F.; Liang, X.; Liu, Q.; Li, X. Ecolgical evaluation on Panjin wetland based on analytic hierarchy process. Geol. Resour. 2018, 27, 268–271. [Google Scholar]
- Manzoni, S.; Vico, G.; Katul, G.; Fay, P.A.; Polley, W.; Palmroth, S.; Porporato, A. Optimizing stomatal conductance for maximum carbon gain under water stress: A meta–analysis across plant functional types and climates. Funct. Ecol. 2011, 25, 456–467. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
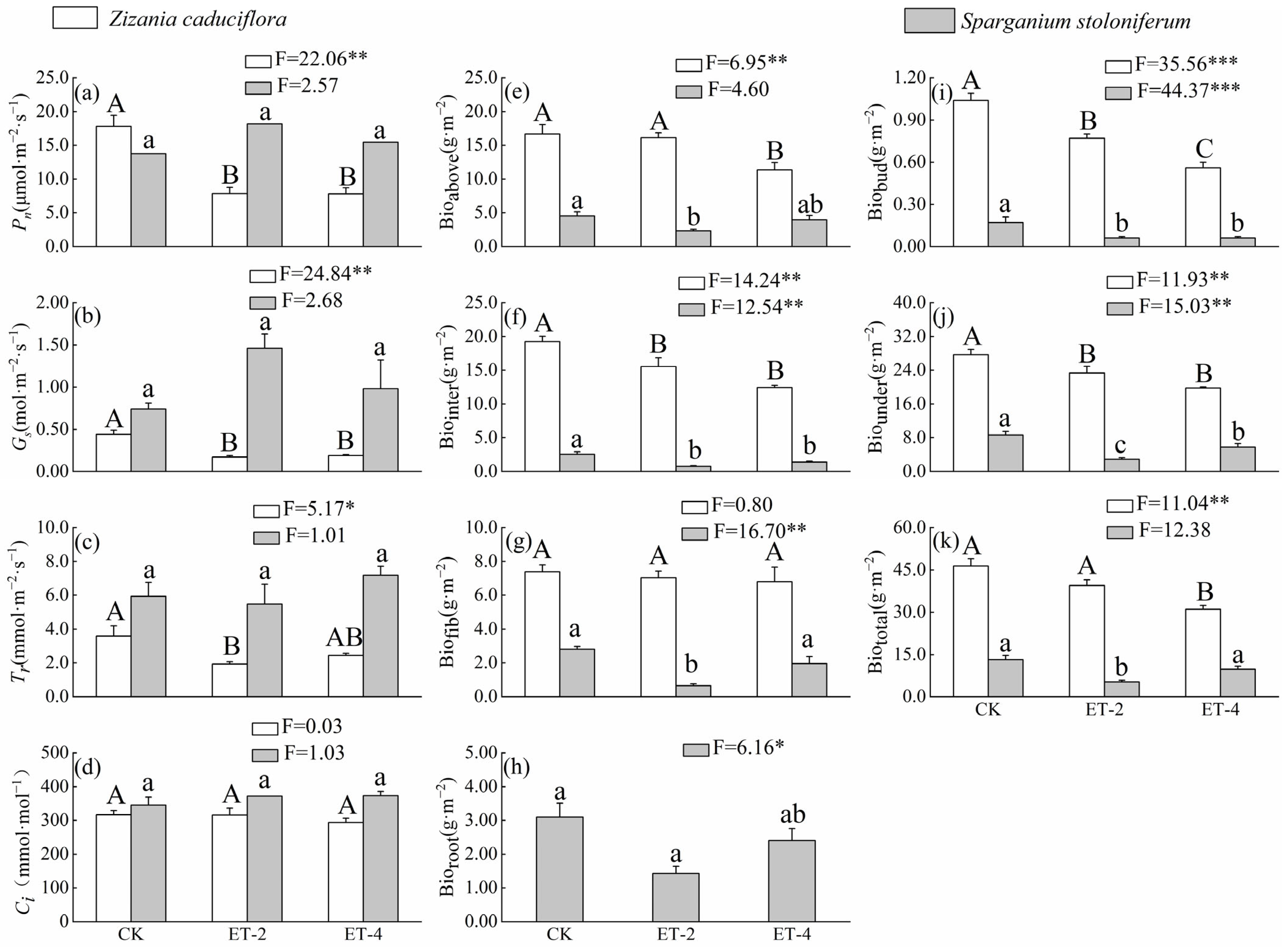
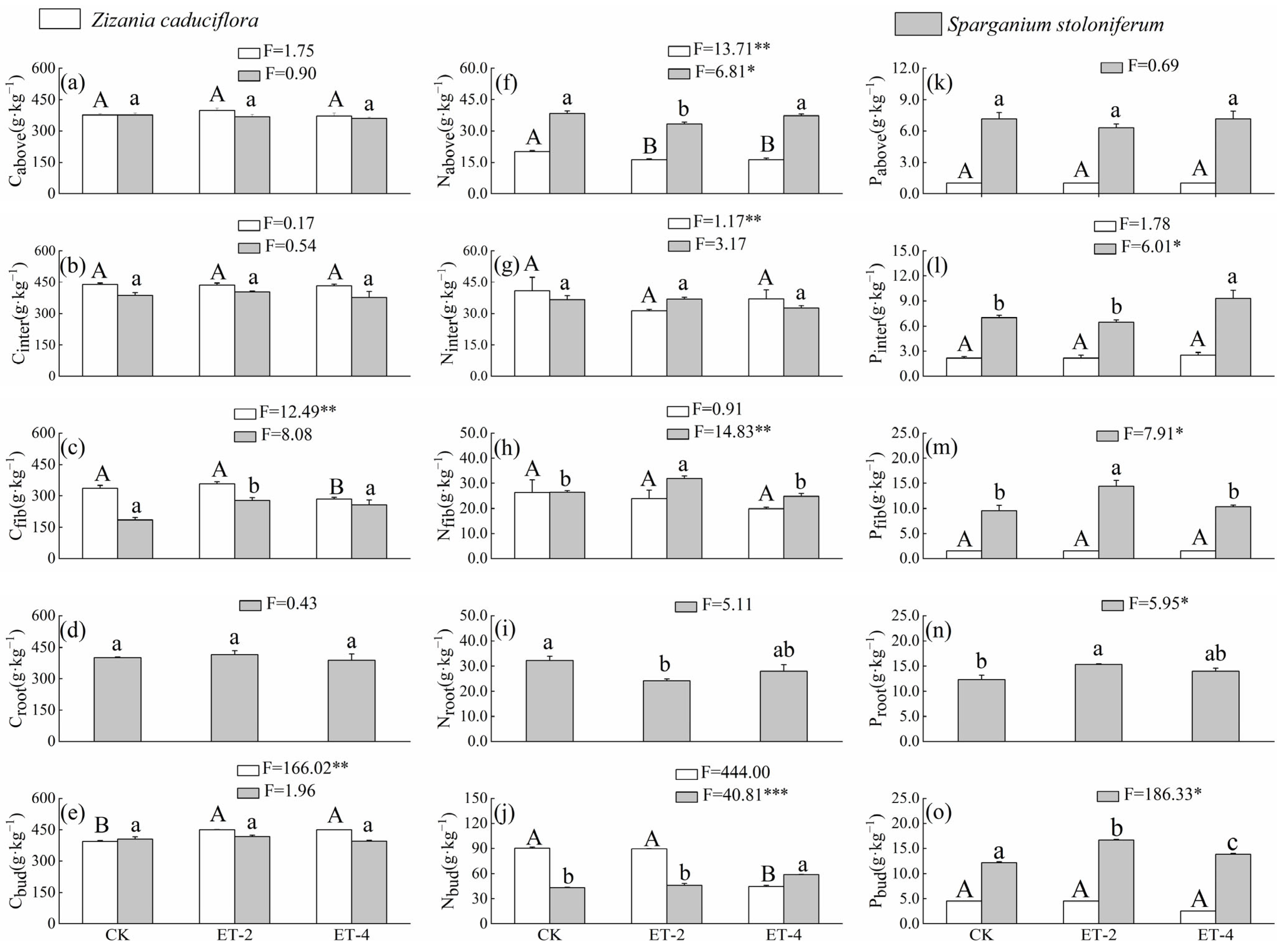


| Traits | Variable | Coefficient | R2 | p | Traits | Variable | Coefficient | R2 | p |
|---|---|---|---|---|---|---|---|---|---|
| Pn | max | −1.05 | 0.89 | 0.000 | Cabove | MAT | −19.90 | 0.37 | 0.253 |
| Constant | 57.19 | max | 7.16 | ||||||
| Constant | 282.04 | ||||||||
| Gs | max | −0.03 | 0.88 | 0.000 | Cinter | none | |||
| Constant | 1.46 | ||||||||
| Tr | max | −0.14 | 0.55 | 0.022 | Cfib | MAT | −50.66 | 0.81 | 0.007 |
| Constant | 9.01 | max | 14.24 | ||||||
| Constant | 247.12 | ||||||||
| Ci | none | Cbud | max | 5.76 | 0.98 | <0.001 | |||
| Constant | 177.19 | ||||||||
| Bioabove | MAT | −3.27 | 0.69 | 0.027 | Nabove | max | −0.42 | 0.82 | 0.001 |
| max | 0.71 | Constant | 35.98 | ||||||
| Constant | 18.75 | ||||||||
| Biointer | MAT | −1.81 | 0.82 | 0.001 | Ninter | none | |||
| Constant | 35.34 | ||||||||
| Biofib | MAT | −0.16 | 0.21 | 0.215 | Nfib | MAT | −1.69 | 0.22 | 0.205 |
| Constant | 8.77 | Constant | 41.66 | ||||||
| Biobud | MAT | −0.13 | 0.92 | 3.974 × 10−5 | Nbud | MAT | −29.02 | 0.99 | <0.001 |
| Constant | 2.18 | max | 6.73 | ||||||
| Constant | 93.03 | ||||||||
| Biounder | MAT | −2.10 | 0.80 | 0.001 | Pabove | none | |||
| Constant | 46.31 | ||||||||
| Biototal | MAT | −3.45 | 0.74 | 0.003 | Pinter | MAT | 0.17 | 0.25 | 0.171 |
| Constant | 75.57 | Constant | 0.59 | ||||||
| Pfib | none | ||||||||
| Pbud | none |
| Traits | Variable | Coefficient | R2 | p | Traits | Variable | Coefficient | R2 | p |
|---|---|---|---|---|---|---|---|---|---|
| Pn | MAT | −2.09 | 0.46 | 0.156 | Cfib | max | 8.64 | 0.67 | 0.007 |
| max | 0.97 | Constant | −140.33 | ||||||
| Constant | −4.54 | ||||||||
| Gs | MAT | −0.37 | 0.47 | 0.148 | Croot | none | |||
| max | 0.16 | ||||||||
| Constant | −2.24 | ||||||||
| Tr | none | Cbud | MAT | −15.74 | 0.39 | 0.222 | |||
| max | 4.99 | ||||||||
| Constant | 356.57 | 0.81 | 0.007 | ||||||
| Ci | max | 2.82 | 0.26 | 0.165 | Nabove | MAT | 3.01 | 0.69 | 0.029 |
| Constant | 239.34 | Max | −1.25 | ||||||
| Constant | 59.05 | ||||||||
| Bioabove | MAT | 1.22 | 0.61 | 0.062 | Ninter | MAT | −2.90 | 0.51 | 0.115 |
| max | −0.53 | max | 0.70 | ||||||
| Constant | 13.62 | Constant | 35.700 | ||||||
| Biointer | MAT | 0.51 | 0.81 | 0.007 | Nfib | MAT | −5.23 | 0.83 | 0.005 |
| max | −0.31 | max | 1.84 | ||||||
| Constant | 9.82 | Constant | 3.29 | ||||||
| Biofib | MAT | 1.00 | 0.85 | 0.004 | Nroot | MAT | 3.04 | 0.63 | 0.051 |
| max | −0.47 | max | −1.61 | ||||||
| Constant | 11.73 | Constant | 66.04 | ||||||
| Bioroot | MAT | 0.75 | 0.67 | 0.035 | Nbud | MAT | 8.64 | 0.93 | 0.000 |
| max | −0.36 | max | −1.71 | ||||||
| Constant | 9.98 | Constant | 31.35 | ||||||
| Biobud | max | −0.01 | 0.94 | 2.092 × 10−5 | Pabove | none | |||
| Constant | 0.63 | ||||||||
| Biounder | MAT | 2.27 | 0.834 | 0.005 | Pinter | MAT | 2.01 | 0.67 | 0.037 |
| max | −1.16 | max | −0.54 | ||||||
| Constant | 32.24 | Constant | 9.44 | ||||||
| Biototal | MAT | 3.49 | 0.81 | 0.007 | Pfib | MAT | −3.07 | 0.725 | 0.021 |
| max | −1.68 | max | 1.26 | ||||||
| Constant | 45.87 | Constant | −10.68 | ||||||
| Cabove | MAT | −4.20 | 0.23 | 0.195 | Proot | MAT | −1.06 | 0.67 | 0.038 |
| Constant | 413.78 | max | 0.58 | ||||||
| Constant | −0.04 | ||||||||
| Cinter | none | Pbud | MAT | −2.17 | 0.98 | <0.001 | |||
| max | 1.00 | ||||||||
| Constant | −6.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Yu, J.; Zhang, Y.; Tian, K.; Zhu, X.; Sun, M.; Liu, Z. Simulated Warming Reduces Biomass Accumulation in Zizania caduciflor and Sparganium stoloniferum. Plants 2025, 14, 1414. https://doi.org/10.3390/plants14101414
Wang T, Yu J, Zhang Y, Tian K, Zhu X, Sun M, Liu Z. Simulated Warming Reduces Biomass Accumulation in Zizania caduciflor and Sparganium stoloniferum. Plants. 2025; 14(10):1414. https://doi.org/10.3390/plants14101414
Chicago/Turabian StyleWang, Tingfeng, Junbao Yu, Yun Zhang, Kun Tian, Xiangyu Zhu, Mei Sun, and Zhenya Liu. 2025. "Simulated Warming Reduces Biomass Accumulation in Zizania caduciflor and Sparganium stoloniferum" Plants 14, no. 10: 1414. https://doi.org/10.3390/plants14101414
APA StyleWang, T., Yu, J., Zhang, Y., Tian, K., Zhu, X., Sun, M., & Liu, Z. (2025). Simulated Warming Reduces Biomass Accumulation in Zizania caduciflor and Sparganium stoloniferum. Plants, 14(10), 1414. https://doi.org/10.3390/plants14101414