
Proline–Nitrogen Metabolic Coordination Mediates Cold Priming-Induced Freezing Tolerance in Maize

 , ,
, ,
Abstract
1. Introduction
2. Results
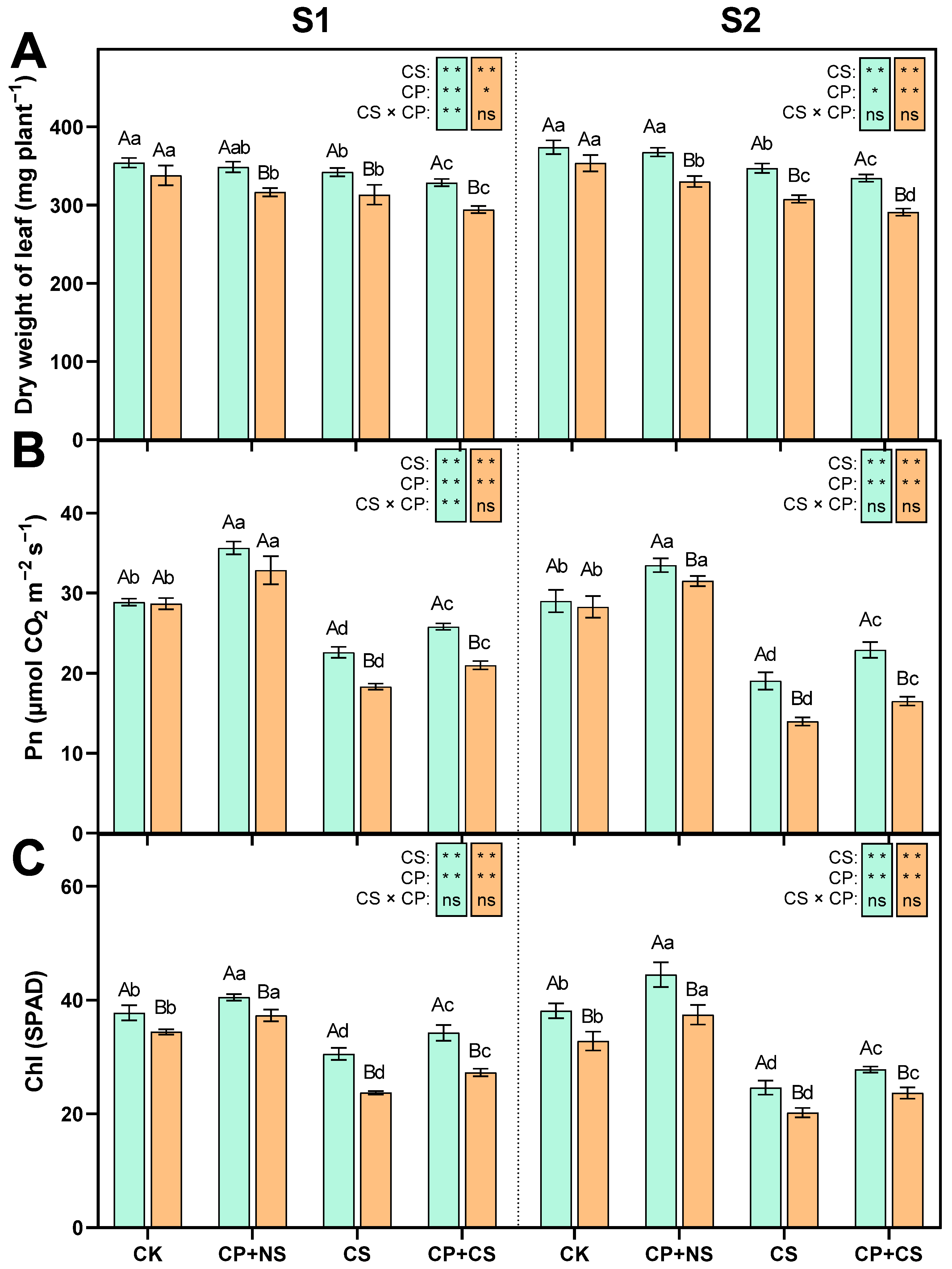
2.1. Leaf Dry Weight, Photosynthetic Rate, and Chlorophyll Content
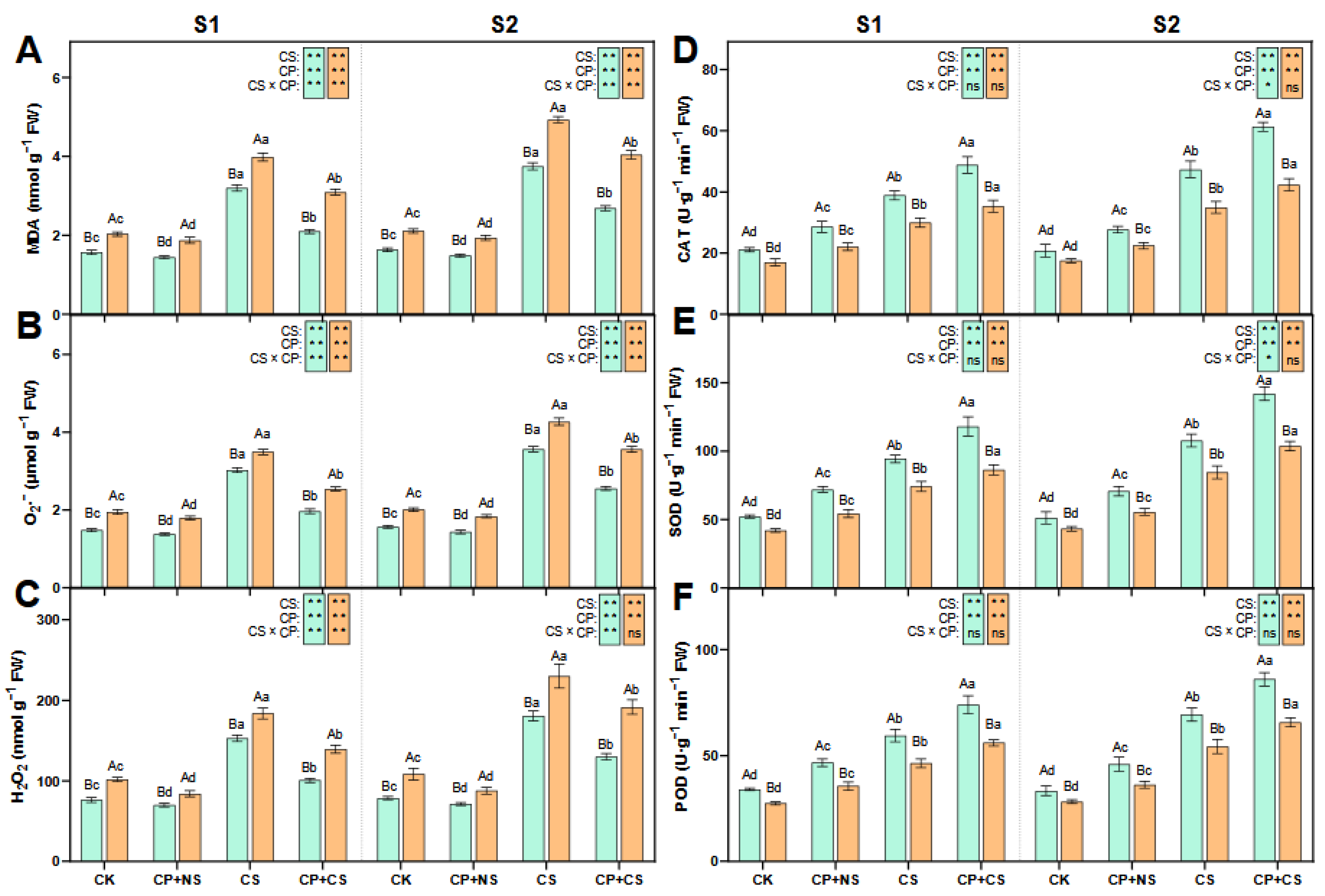
2.2. ROS Metabolism
2.3. Soluble Sugar and Amino Acid Content
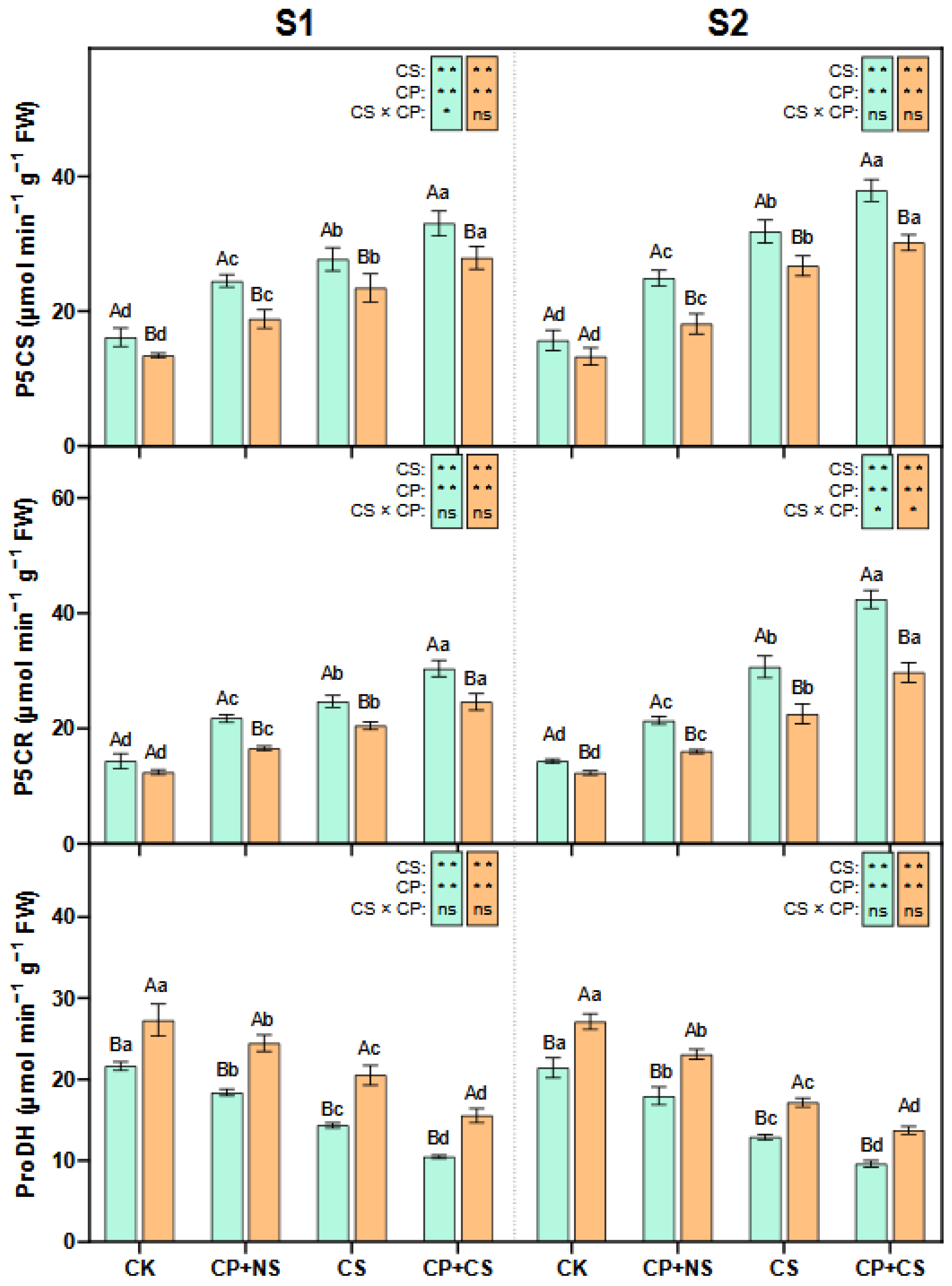
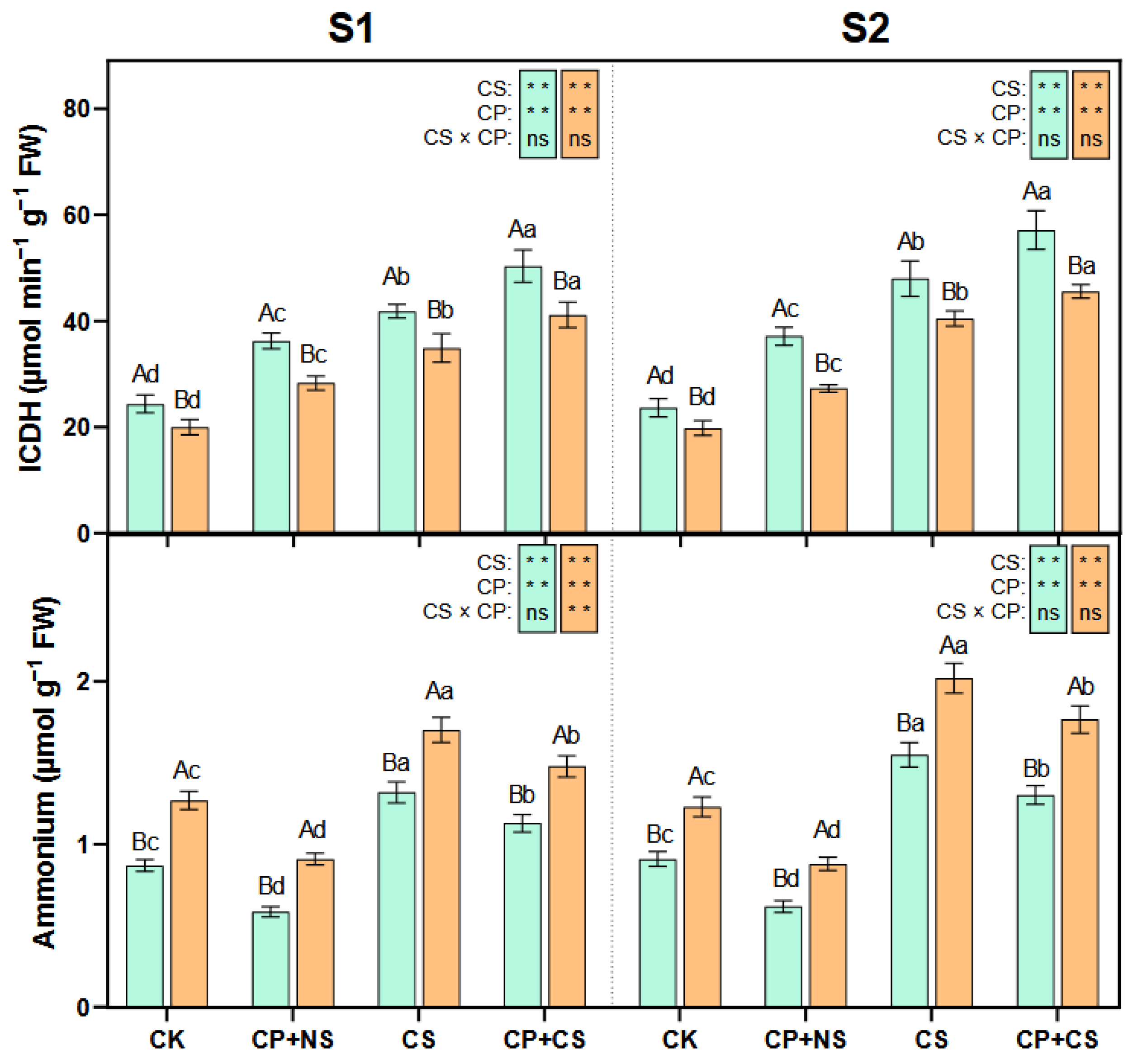
2.4. Proline Metabolism Enzymes
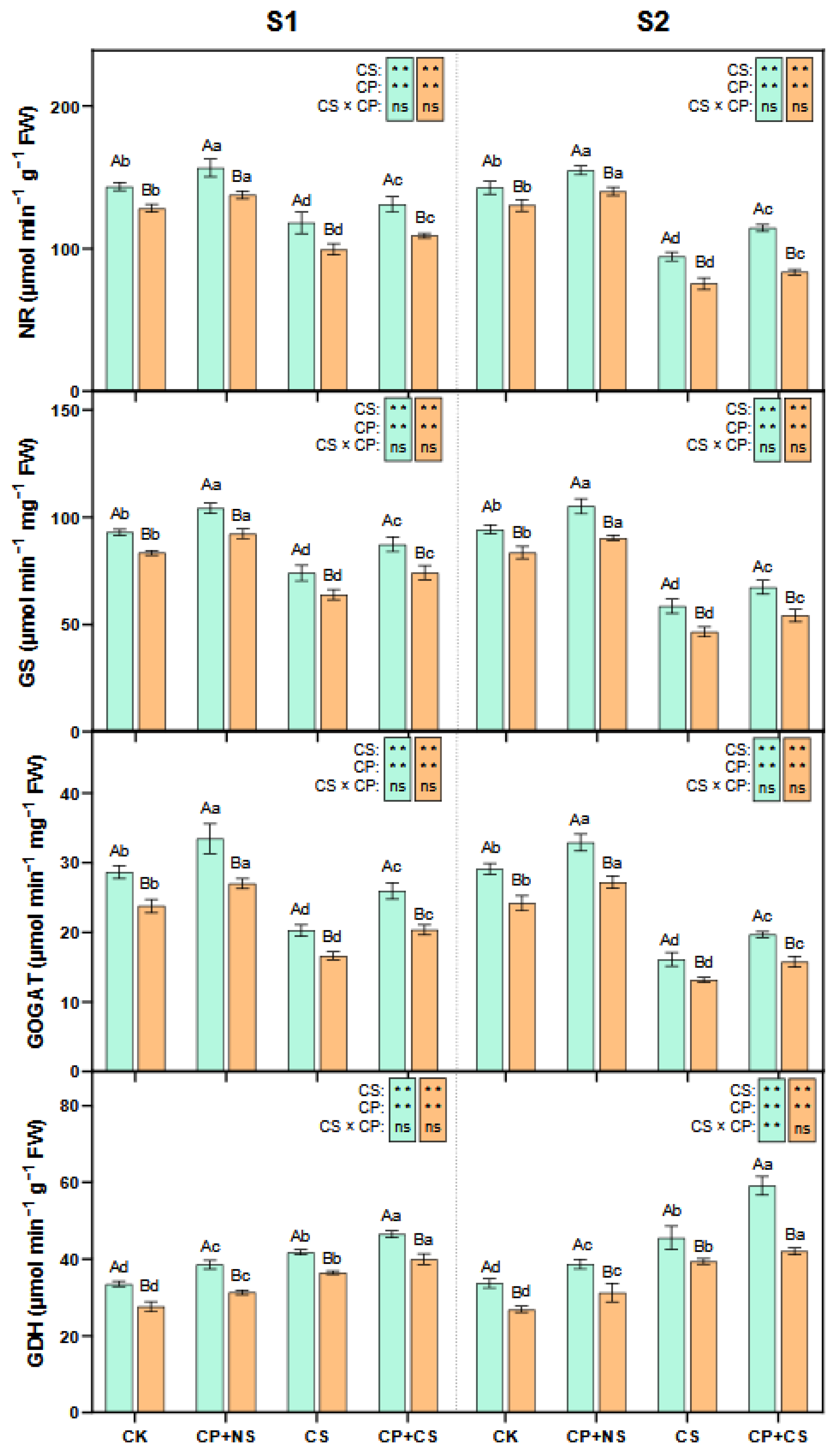
2.5. GS-GOGAT Cycle
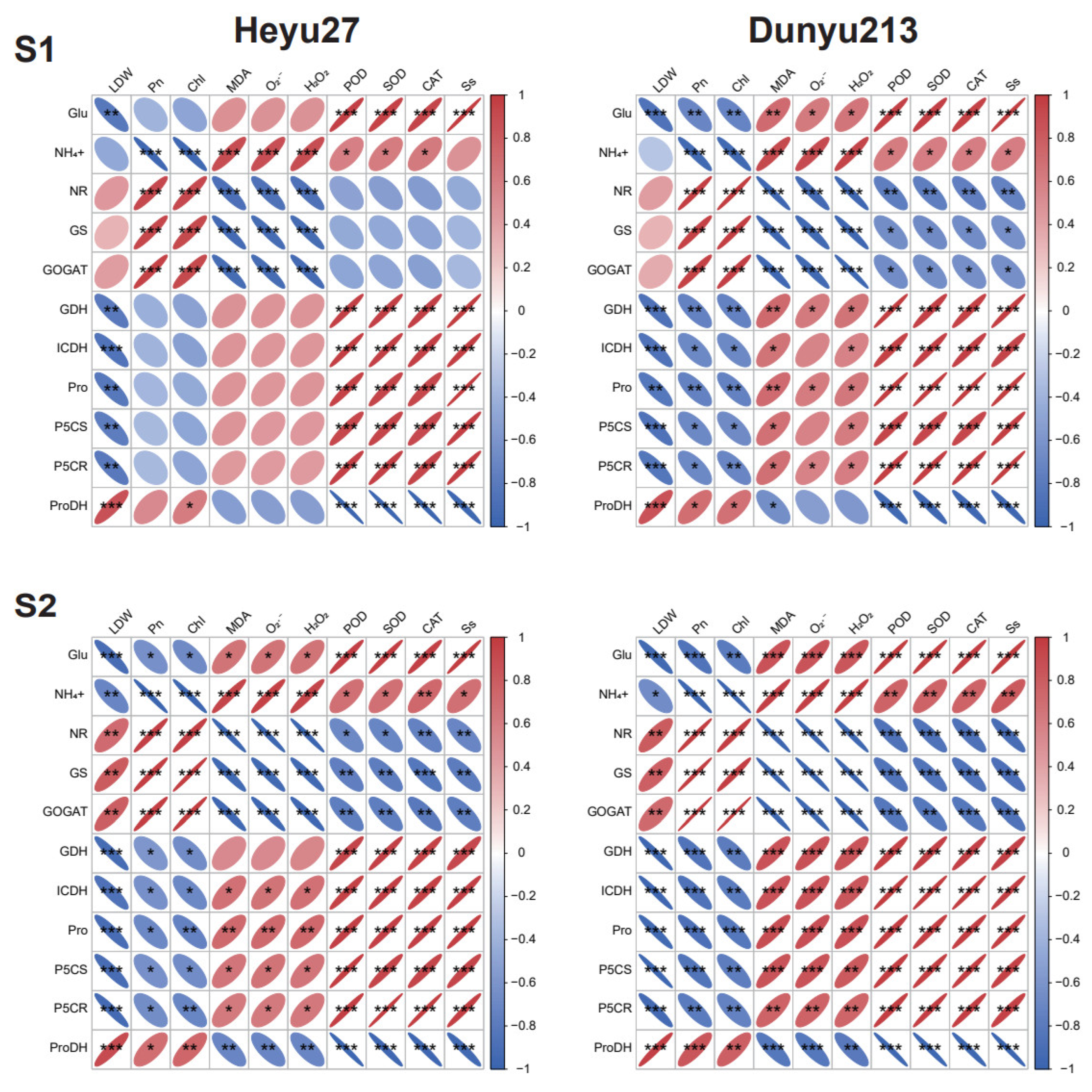
2.6. Multivariate Statistical Analysis
3. Discussion
3.1. The Impact of Cold Priming on Plant Growth and Photosynthetic System
3.2. The Impact of Cold Priming on ROS Metabolism
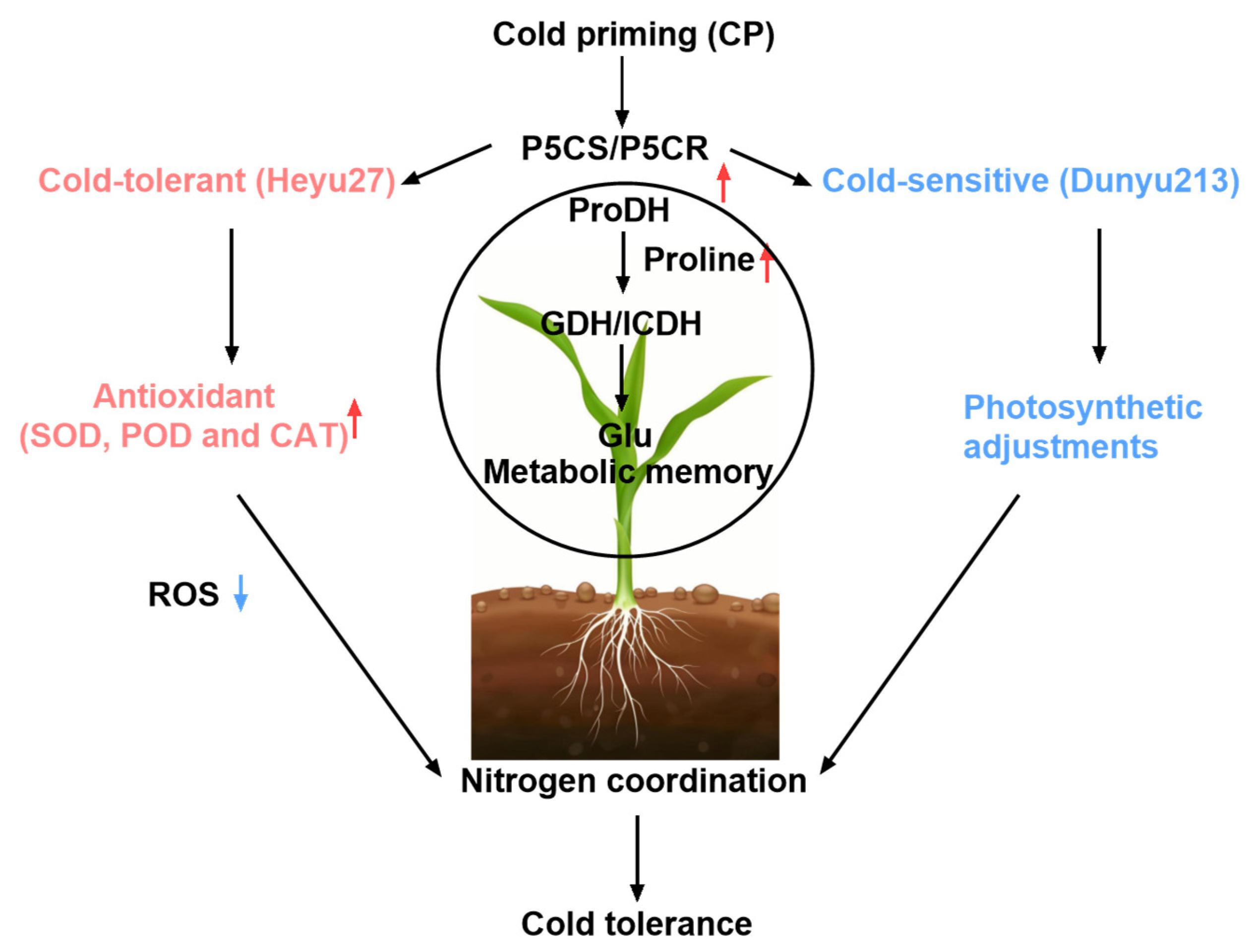
3.3. Synergistic Regulation of Proline and Nitrogen Metabolism by Cold Priming
4. Materials and Methods
4.1. Experimental Materials and Growth Conditions
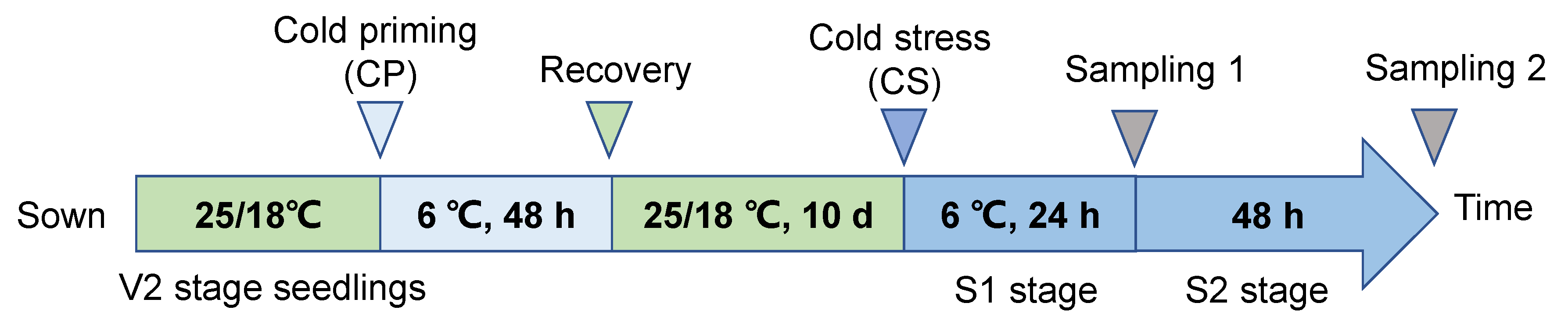
4.2. Experimental Design
4.3. Measurement of Photosynthetic Parameters and Chlorophyll Content
4.4. Measurement of Key Enzyme Activities in Nitrogen Metabolism
4.5. Measurement of ROS Contents and Key Enzyme Activities in ROS Scavenging
4.6. Determination of Amino Acids and Soluble Sugar
4.7. Measurement of Proline Metablism Enzymes
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, D.F.; Shawna, L.N.; Archie, R.P.J.; Stephen, P.M.; Stephen, P.L. Can the Cold Tolerance of C4 Photosynthesis in Miscanthus×Giganteus Relative to Zea mays be Explained by Differences in Activities and Thermal Properties of Rubisco? J. Exp. Bot. 2008, 59, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.Y.; Feng, R.; Ji, R.P.; Wang, H.B.; Zhu, Y.N.; Che, Y.S.; Zhang, Y.S. Effects of Low Temperature Stress in Seedling Stage on Growth Development and Yield for Maize. Agric. Res. Arid Areas 2013, 31, 220–226. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-GHDQ201305038.htm (accessed on 5 March 2025).
- Chen, S.; Liu, H.L.; Yangzong, Z.X.; Gardea-Torresdey, J.L.; White, J.C.; Zhao, L.J. Seed Priming with Reactive Oxygen Species-Generating Nanoparticles Enhanced Maize Tolerance to Multiple Abiotic Stresses. Environ. Sci. Technol. 2023, 57, 19932–19941. [Google Scholar] [CrossRef]
- Penfield, S. Temperature Perception and Signal Transduction in Plants. New Phytol. 2008, 179, 615–628. [Google Scholar] [CrossRef]
- Li, W.Q.; Li, J.Y.; Bi, S.J.; Jin, J.Y.; Fan, Z.L.; Shang, Z.L.; Zhang, Y.F.; Wang, Y.J. Melatonin Enhances Maize Germination, Growth, and Salt Tolerance by Regulating Reactive Oxygen Species Accumulation and Antioxidant Systems. Plants 2025, 14, 296. [Google Scholar] [CrossRef]
- Leconte, J.M.L.; Marco, M.; Nicolas, B.; Gabriela, B.; Sébastien, C.; Olivier, C.; Alexis, C.; Marc, L.; Remy, M.; Nicolas, P.; et al. Multi-Scale Characterisation of Cold Response Reveals Immediate and Long-Term Impacts on Cell Physiology up to Seed Composition in Sunflower. Plant Cell Environ. 2024, 48, 2596–2614. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold Stress Regulation of Gene Expression in Plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of Proline Biosynthesis, Degradation, Uptake and Transport in Higher Plants: Its Implications in Plant Growth and Abiotic Stress Tolerance. Curr. Sci. 2005, 88, 424–438. Available online: https://www.researchgate.net/publication/242087984_Regulation_of_proline_biosynthesis_degradation_uptake_and_transport_in_higher_plants_Its_implications_in_plant_growth_and_abiotic_stress_tolerance (accessed on 5 March 2025).
- Verbruggen, N.; Hermans, C. Proline Accumulation in Plants: A Review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Renzetti, M.; Funck, D.; Trovato, M. Proline and ROS: A Unified Mechanism in Plant Development and Stress Response? Plants 2025, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.B.; Liu, H.H.; Zhang, Y.P.; Cao, Y.X.; Hu, Y.W.; Wang, Y.L.; Zheng, H.Q.; Liu, M.Z. Humic Acid Alleviates Low-Temperature Stress by Regulating Nitrogen Metabolism and Proline Synthesis in Melon (Cucumis melo L.) Seedlings. Horticulturae 2025, 11, 16. [Google Scholar] [CrossRef]
- Dong, L.; Li, L.J.; Meng, Y.; Liu, H.L.; Li, J.; Yu, Y.; Qian, C.R.; Wei, S.; Gu, W.R. Exogenous Spermidine Optimizes Nitrogen Metabolism and Improves Maize Yield under Drought Stress Conditions. Agriculture 2022, 12, 1270. [Google Scholar] [CrossRef]
- Hilker, M.; Schwachtje, J.; Baier, M.; Balazadeh, S.; Bäurle, I.; Geiselhardt, S.; Hincha, D.K.; Kunze, R.; Mueller-Roeber, B.; Rillig, M.G.; et al. Priming and Memory of Stress Responses in Organisms Lacking a Nervous System. Biol. Rev. 2016, 91, 1118–1133. [Google Scholar] [CrossRef]
- Li, X.N.; Cai, J.; Liu, F.L.; Dai, T.B.; Cao, W.X.; Jiang, D. Cold Priming Drives the Sub-Cellular Antioxidant Systems to Protect Photosynthetic Electron Transport against Subsequent Low Temperature Stress in Winter Wheat. Plant Physiol. Biochem. 2014, 82, 34–43. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Wang, Z.; Sun, Z.; Zhu, X.; Liu, S.; Song, F.; Liu, F.; Wang, Y. Cold Priming Induced Tolerance to Subsequent Low Temperature Stress is Enhanced by Melatonin Application during Recovery in Wheat. Molecules 2018, 23, 1091. [Google Scholar] [CrossRef]
- Wang, W.L.; Wang, X.; Zhang, X.Y.; Wang, Y.; Huo, Z.Y.; Huang, M.; Cai, J.; Zhou, Q.; Jiang, D. Involvement of Salicylic Acid in Cold Priming-Induced Freezing Tolerance in Wheat Plants. Plant Growth Regul. 2021, 93, 117–130. [Google Scholar] [CrossRef]
- Kaur, S.; Gupta, A.K.; Kaur, N. Effect of Osmo- and Hydro-Priming of Chickpea Seeds on Seedling Growth and Carbohydrate Metabolism under Water Deficit Stress. Plant Growth Regul. 2002, 37, 17–22. [Google Scholar] [CrossRef]
- Kaya, M.D.; Okçu, G.; Atak, M.; Çikili, Y.; Kolsarici, Ö. Seed Treatments to Overcome Salt and Drought Stress during Germination in Sunflower (Helianthus annuus L.). Eur. J. Agron. 2006, 24, 291–295. [Google Scholar] [CrossRef]
- Casenave, E.C.; Toselli, M.E. Hydropriming as a Pre-Treatment for Cotton Germination under Thermal and Water Stress Conditions. Seed Sci. Technol. 2007, 35, 88–98. [Google Scholar] [CrossRef]
- Amini, A.; Majidi, M.M.; Mokhtari, N.; Ghanavati, M. Drought Stress Memory in a Germplasm of Synthetic and Common Wheat: Antioxidant System, Physiological and Morphological Consequences. Sci. Rep. 2023, 13, 138569. [Google Scholar] [CrossRef] [PubMed]
- Sintaha, M.; Man, C.K.; Yung, W.S.; Duan, S.; Li, M.W.; Lam, H.M. Drought Stress Priming Improved the Drought Tolerance of Soybean. Plants 2022, 11, 2954. [Google Scholar] [CrossRef] [PubMed]
- Rossatto, T.; Souza, G.M.; Amaral, M.N.D.; Auler, P.A.; Perez-Alonso, N.M.M.; Pollmann, S.; Braga, E.J.B. Cross-Stress Memory: Salt Priming at Vegetative Growth Stages Improves Tolerance to Drought Stress during Grain-Filling in Rice Plants. Environ. Exp. Bot. 2023, 206, 105187. [Google Scholar] [CrossRef]
- Arunyanark, A.; Jogloy, S.; Akkasaeng, C.; Vorasoot, N.; Kesmala, T.; Rao, R.C.N.; Wright, G.C.; Patanothai, A. Chlorophyll Stability is an Indicator of Drought Tolerance in Peanut. J. Agron. Crop Sci. 2008, 194, 113–125. [Google Scholar] [CrossRef]
- Schwachtje, J.; Whitcomb, S.J.; Firmino, A.A.P.; Zuther, E.; Hincha, D.K.; Kopka, J. Induced, Imprinted, and Primed Responses to Changing Environments: Does Metabolism Store and Process Information? Front. Plant Sci. 2019, 10, 106. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Advances and Challenges in Uncovering Cold Tolerance Regulatory Mechanisms in Plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef]
- David, S.K.; Susanne, V.C.; Robert, T.F.; Rowan, F.S. C4 Photosynthesis at Low Temperature. A Study Using Transgenic Plants with Reduced Amounts of Rubisco. Plant Physiol. 2003, 132, 1577–1585. [Google Scholar] [CrossRef]
- Long, S.P. Environmental Responses. In C4 Plant Biology; Sage, R.F., Monson, R.K., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 215–249. [Google Scholar]
- Sage, R.F. Why C4 Photosynthesis? In C4 Plant Biology; Sage, R.F., Monson, R.K., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 3–16. [Google Scholar]
- Osmond, C.B.; Winter, K.; Ziegler, H. Functional significance of different pathways of CO2 fixation in photosynthesis. In Physiological Plant Ecology II. Encyclopedia of Plant Physiology; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer-Verlag: Berlin, Germany, 1982; pp. 480–547. [Google Scholar]
- Singha, A.; Soothar, R.K.; Wang, C.; Marín, E.E.T.; Tankari, M.; Hao, W.P.; Wang, Y.S. Drought Priming Alleviated Salinity Stress and Improved Water Use Efficiency of Wheat Plants. Plant Growth Regul. 2022, 96, 357–368. [Google Scholar] [CrossRef]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.H.; Zhu, J.K. Methods and Concepts in Quantifying Resistance to Drought, Salt and Freezing, Abiotic Stresses that Affect Plant Water Status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef]
- Sun, L.; Yang, Y.B.; Wang, R.; Li, S.; Qiu, Y.B.; Lei, P.; Gao, J.; Xu, H.; Zhang, F.L.; Lv, Y.F. Effects of Exopolysaccharide Derived From Pantoea alhagi NX-11 on Drought Resistance of Rice and its Efficient Fermentation Preparation. Int. J. Biol. Macromol. 2020, 162, 946–955. [Google Scholar] [CrossRef]
- Zhang, J.L.; Poorter, L.; Cao, K.F. Productive Leaf Functional Traits of Chinese Savanna Species. Plant Ecol. 2012, 213, 1449–1460. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS Signaling: The New Wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Matsuoka, D.; Nanmori, T. Membrane Rigidification Functions Upstream of The MEKK1-MKK2-MPK4 Cascade during Cold Acclimation in Arabidopsis thaliana. FEBS Lett. 2014, 588, 2025–2030. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl Radical Scavenging Activity of Compatible Solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.C.; Li, Z.B.; Song, J.Q.; Liu, Y.J.; Qiu, Y.Z.; Zhang, Y.; Li, X. Atmospheric NO2 Enhances Tolerance to Low Temperature by Promoting Nitrogen and Carbon Metabolism in Tobacco. Environ. Exp. Bot. 2024, 225, 105860. [Google Scholar] [CrossRef]
- Zuo, S.Q.; Li, F.Y.; Gu, X.; Wei, Z.P.; Qiao, L.K.; Du, C.Y.; Chi, Y.Z.; Liu, R.Z.; Wang, P. Effects of Low Molecular Weight Polysaccharides from Ulva Prolifera on the Tolerance of Triticum Aestivum to Osmotic Stress. Int. J. Biol. Macromol. 2021, 183, 12–22. [Google Scholar] [CrossRef]
- Sage, R.F. The Evolution of C4 Photosynthesis. New Phytol. 2004, 161, 341–370. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of Photosynthesis, Nitrogen and Proline Metabolism to Salinity Stress in Solanum Lycopersicum under Different Levels of Nitrogen Supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.B.; Yuan, Y.Z.; Zhang, C.F.; Ou, J.Q.; Zhou, W.; Lin, Q.H. Modulation of Key Enzymes Involved in Ammonium Assimilation and Carbon Metabolism by Low Temperature in Rice (Oryza sativa L.) Roots. Plant Sci. 2005, 169, 295–302. [Google Scholar] [CrossRef]
- Tambussi, E.A.; Bartoli, C.G.; Guiamet, J.J.; Beltrano, J.; Araus, J.L. Oxidative Stress and Photodamage at Low Temperatures in Soybean (Glycine max L. Merr.) Leaves. Plant Sci. 2004, 167, 19–26. [Google Scholar] [CrossRef]
- Aslam, M.; Huffaker, R.C.; Rains, D.W. Early Effects of Salinity on Nitrate Assimilation in Barley Seedlings. Plant Physiol. 1984, 76, 321–325. [Google Scholar] [CrossRef]
- Liang, C.G.; Chen, L.P.; Wang, Y.; Liu, J.; Xu, G.; Li, T. High Temperature at Grain-filling Stage Affects Nitrogen Metabolism Enzyme Activities in Grains and Grain Nutritional Quality in Rice. Rice Sci. 2011, 18, 210–216. [Google Scholar] [CrossRef]
- Singh, R.P.; Srivastava, H. Regulation of glutamate dehydrogenase activity by amino acids in maize seedlings. Physiol. Plant. 1983, 57, 549–554. [Google Scholar] [CrossRef]
- Zhao, S.; Li, D. Experiment Guide of Modern Plant Physiology; Science Press: Beijing, China, 1999. [Google Scholar]
- Xu, J.; Zhu, Y.; Ge, Q.; Li, Y.; Sun, J.; Zhang, Y.; Liu, X. Comparative Physiological Responses of Solanum Nigrum and Solanum Torvum to Cadmium Stress. New Phytol. 2012, 196, 125–138. [Google Scholar] [CrossRef]
- He, J.; Ma, C.; Ma, Y.; Li, H.; Kang, J.; Liu, T.; Polle, A.; Peng, C.; Luo, Z. Cadmium Tolerance in Six Poplar Species. Environ. Sci. Pollut. Res. 2013, 20, 163–174. [Google Scholar] [CrossRef]
- Pompeu, G.B.; Vilhena, M.B.; Gratao, P.L.; Carvalho, R.F.; Rossi, M.L.; Martinelli, A.P.; Azevedo, R.A. Abscisic Acid-Deficient Sit Tomato Mutant Responses to Cadmium-Induced Stress. Protoplasma 2017, 254, 771–783. [Google Scholar] [CrossRef]
- Fimognari, L.; Dolker, R.; Kaselyte, G.; Jensen, C.N.G.; Akhtar, S.S.; Grosskinsky, D.K.; Roitsch, T. Simple Semi-High Throughput Determination of Activity Signatures of Key Antioxidant Enzymes for Physiological Phenotyping. Plant Methods 2020, 16, 42. [Google Scholar] [CrossRef]
- Di Martino, C.; Delfine, S.; Alvino, A.; Loreto, F. Photorespiration Rate in Spinach Leaves under Moderate Nacl Stress. Photosynthetica 1999, 36, 233–242. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Quan, R.D.; Shang, M.; Zhang, H.; Zhao, Y.X.; Zhang, J.R. Improved Chilling Tolerance by Transformation with Beta Gene for the Enhancement of Glycinebetaine Synthesis in Maize. Plant Sci. 2004, 166, 141–149. [Google Scholar] [CrossRef]
- Yang, H.C. The Effect of Drought Stress Influence in the Proline Metabolism Pathways of Seedling Period of Sugar Beet. Master’s Thesis, Harbin Institute of Technology, Harbin, China, 2016. [Google Scholar]
- Bao, G.G.; Ashraf, U.; Wang, C.L.; He, L.X.; Wei, X.S.; Zheng, A.; Mo, Z.W.; Tang, X.R. Molecular Basis for Increased 2-Acetyl-1-Pyrroline Contents Under Alternate Wetting and Drying (AWD) Conditions in Fragrant Rice. Plant Physiol. Biochem. 2018, 133, 149–157. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total N (g/kg) | Available N (mg/kg) | Total P (g/kg) | Available P (mg/kg) | Total K (g/kg) | Available K (mg/kg) | Organic Matter (g/kg) | pH |
|---|---|---|---|---|---|---|---|
| 2.87 ± 0.11 | 148.65 ± 4.65 | 1.65 ± 0.09 | 32.21 ± 3.23 | 29.53 ± 2.78 | 132.43 ± 4.21 | 32.43 ± 2.91 | 7.08 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gai, Z.; Liu, L.; Zhang, N.; Liu, J.; Cai, L.; Yang, X.; Zhang, A.; Zhang, P.; Ding, J.; Zhang, Y. Proline–Nitrogen Metabolic Coordination Mediates Cold Priming-Induced Freezing Tolerance in Maize. Plants 2025, 14, 1415. https://doi.org/10.3390/plants14101415
Gai Z, Liu L, Zhang N, Liu J, Cai L, Yang X, Zhang A, Zhang P, Ding J, Zhang Y. Proline–Nitrogen Metabolic Coordination Mediates Cold Priming-Induced Freezing Tolerance in Maize. Plants. 2025; 14(10):1415. https://doi.org/10.3390/plants14101415
Chicago/Turabian StyleGai, Zhijia, Lei Liu, Na Zhang, Jingqi Liu, Lijun Cai, Xu Yang, Ao Zhang, Pengfei Zhang, Junjie Ding, and Yifei Zhang. 2025. "Proline–Nitrogen Metabolic Coordination Mediates Cold Priming-Induced Freezing Tolerance in Maize" Plants 14, no. 10: 1415. https://doi.org/10.3390/plants14101415
APA StyleGai, Z., Liu, L., Zhang, N., Liu, J., Cai, L., Yang, X., Zhang, A., Zhang, P., Ding, J., & Zhang, Y. (2025). Proline–Nitrogen Metabolic Coordination Mediates Cold Priming-Induced Freezing Tolerance in Maize. Plants, 14(10), 1415. https://doi.org/10.3390/plants14101415