Abstract
Flax seed is one of the richest plant sources of linolenic acid (LIN) and also contains unsaturated linoleic acid (LIO) and oleic acid (OLE). Stearoyl-ACP desaturases (SADs) and fatty acid desaturases (FADs) play key roles in the synthesis of flax fatty acids (FAs). However, there is no holistic view of which genes from the SAD and FAD families and at which developmental stages have the highest expression levels in flax seeds, as well as the influence of genotype and growth conditions on the expression profiles of these genes. We sequenced flax seed transcriptomes at 3, 7, 14, 21, and 28 days after flowering (DAF) for ten flax varieties with different oil FA compositions grown under three temperature/watering conditions. The expression levels of 25 genes of the SAD, FAD2, and FAD3 families were evaluated. FAD3b, FAD3a, FAD2b-2, SAD3-1, SAD2-1, SAD2-2, SAD3-2, FAD2a-1, and FAD2a-2 had the highest expression levels, which changed significantly during seed development. These genes probably play a key role in FA synthesis in flax seeds. High temperature and insufficient watering shifted the maximum expression levels of FAD and SAD genes to earlier developmental stages, while the opposite trend was observed for low temperature and excessive watering. Differences in the FAD and SAD expression profiles under different growth conditions may affect the FA composition of linseed oil. Stop codons in the FAD3a gene, resulting in a reduced LIN content, decreased the level of FAD3a transcript. The obtained results provide new insights into the synthesis of linseed oil.
Keywords:
Linum usitatissimum; flax; fatty acids; SAD; FAD; gene expression; transcriptome sequencing 1. Introduction
Flax (Linum usitatissimum L.) seed is one of the richest plant sources of omega-3 fatty acids, which are essential for humans and prevent the onset and progression of many diseases [1,2,3,4]. Flax varieties differ significantly in the fatty acid (FA) composition of the oil, especially in the content of linolenic acid (omega-3) [5,6,7]. Traditional flax varieties are high in linolenic acid (LIN, 50–60%) and are used in the pharmaceutical and paint industries [6,8]. Varieties with low (about 5%) and medium (30–40%) LIN contents have also been developed and are promising for use in the food industry due to the greater resistance of oil to rancidity [6,8,9]. Linoleic acid (LIO, omega-6) and oleic acid (OLE, omega-9) are also important unsaturated fatty acids in flax seeds. There is an inverse relationship between LIO and LIN levels: varieties with high LIN contents have low LIO contents, and varieties with low LIN contents have high LIO contents [7,10]. The differences in the OLE content between varieties are less pronounced and can be about 2-fold [7,10]. Varieties with higher OLE contents are promising for the production of oxidation-stable edible oil, chemical feedstock, and biodiesel [8,9].
Desaturases are known to play a key role in the synthesis of flax fatty acids by introducing double bonds into the hydrocarbon chain. Stearoyl-ACP desaturases (SAD) catalyze the conversion of stearic acid (STE) to oleic acid, fatty acid desaturases 2 (FAD2) catalyze the conversion of oleic acid to linoleic acid, and fatty acid desaturases 3 (FAD3) catalyze the conversion of linoleic acid to linolenic acid [11,12,13,14,15,16,17]. In flax, 25 genes of the SAD and FAD families were identified: SAD2-1, SAD2-2, SAD3-1, SAD3-2, FAD2a-1, FAD2a-2, FAD2b-1, FAD2b-2, FAD2c-1, FAD2c-2, FAD2d-1, FAD2d-2, FAD2e-1, FAD2e-2, FAD2f-1, FAD2f-2, FAD2g-1, FAD2g-2, FAD2h, FAD3a, FAD3b, FAD3c-1, FAD3c-2, FAD3d-1, and FAD3d-2 [18,19]. However, it is probable that not all of these genes play an important role in linseed oil synthesis; some are practically not expressed in seeds but have rather high expression levels in other organs of flax plants [18,19].
Several studies investigated changes in the expression levels of the FAD and SAD genes in flax seeds during development. As a result, the stages at which these genes were most active were identified: 20 days after flowering (DAF) for FAD3 family genes [17]; 16, 22, and 30 DAF for genes of SAD, FAD2, and FAD3 families with variations among genotypes [20]; 20 and 24 DAF for genes of SAD, FAD2, and FAD3 families [21]; and 15 and 20 DAF for genes of the FAD2 family [8]. In general, it is clear that the expression levels of FAD and SAD genes were the highest 2–4 weeks after flowering. However, data from different studies are somewhat divergent, which can be explained by differences in genotypes and growth conditions. Most importantly, in all of the above-mentioned studies, the transcript levels were evaluated for only some of the 25 FAD and SAD genes known in flax. Therefore, it is difficult to make generalizations about the expression of FAD and SAD genes in flax seeds during development.
The effect of genotype on the transcript levels of FAD and SAD genes in flax seeds was also evaluated. Missense and nonsense mutations in the FAD3a and FAD3b genes are known to result in a reduced LIN content in oil [7,16,17,22,23,24,25,26]. The association of decreased FAD3a and FAD3b transcript levels with nonsense mutations in these genes was reported [16,17,20]. However, no change in the FAD3b transcript level was observed in genotypes with the missense mutation in this gene, leading to a decrease in the LIN content [17]. Furthermore, no correlation was found between the expression levels of SAD1, SAD2, FAD2a, FAD2b, FAD3a, and FAD3b and the FA composition of linseed oil in genotypes with the same desaturase isoforms but different FA compositions [21].
In addition to the genotype, it is known that the environment has an influence on the linseed oil content and its FA composition; in particular, an elevated temperature with deficient moisture led to an increase in the OLE content, whereas a decreased temperature with excess moisture had the opposite effect, leading to an increase in the LIO and LIN contents [10,27,28,29,30]. However, it is currently unclear how growth conditions affect the expression profiles of specific FAD and SAD genes in flax seeds during development, and whether different genotypes respond similarly to growing in different environments at the level of FAD and SAD gene expression. We chose contrasting temperature and watering conditions, which led to multidirectional changes in the FA composition of linseed oil, to evaluate the maximum expression variations of FAD and SAD genes in flax seeds.
Since genes of the FAD and SAD families play a key role in determining the most important trait of linseed, namely the oil FA composition, it is necessary to understand, as much as possible, which of these genes can contribute the most to the synthesis of linseed oil fatty acids and how genotype and conditions alter their expression profiles. Finding answers to these questions was the focus of our work.
2. Results
2.1. Results of Transcriptome Sequencing of Flax Seeds
The sequencing of 266 flax seed transcriptome libraries was performed. Ten flax varieties, which differed in oil FA composition and the presence of mutations in the FAD3 genes, leading to a reduced LIN content, were used in this work (Table 1): high-LIN AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh; mid-LIN Raciol and AGT 422; and low-LIN AGT 981, AGT 1535, and Lola. The seeds were collected at 3, 7, 14, 21, and 28 days after flowering (DAF) from plants grown under conditions of 16 °C with overwatering (hereafter referred to as 16 °C), 20 °C with optimal watering (hereafter referred to as 20 °C), and 24 °C with underwatering (hereafter referred to as 24 °C). No data were obtained for the variety AGT 422 at 21 and 28 DAF at 24 °C and for the variety Lola at 28 DAF at 24 °C due to the negative impacts of elevated temperature and insufficient watering on these genotypes. Sequencing was performed in two biological replicates for the samples collected at 3, 7, 14, and 21 DAF and in one biological replicate for the samples collected at 28 DAF (because of the difficulty of obtaining high-quality RNA from seeds at 28 DAF). On average, 2 million paired-end reads were generated for each transcriptome library.

Table 1.
Characteristics of flax varieties used in this work.
To visualize the relatedness of the studied samples, they were clustered based on the expression of all identified transcripts. The samples were grouped mainly by developmental stage (i.e., DAF) and growth conditions (16 °C, 20 °C, and 24 °C) rather than by their belonging to a specific genotype (Figure S1).
2.2. Expression of FAD and SAD Genes in Flax Seeds
The expression of FAD and SAD family genes, known to play an important role in linseed FA synthesis [14,16,18,19,23,26,31,32], was evaluated. From 25 flax genes of the SAD and FAD families (SAD2-1, SAD2-2, SAD3-1, SAD3-2, FAD2a-1, FAD2a-2, FAD2b-1, FAD2b-2, FAD2c-1, FAD2c-2, FAD2d-1, FAD2d-2, FAD2e-1, FAD2e-2, FAD2f-1, FAD2f-2, FAD2g-1, FAD2g-2, FAD2h, FAD3a, FAD3b, FAD3c-1, FAD3c-2, FAD3d-1, and FAD3d-2) [18,19,26], the highest expression levels in the seeds were found for FAD3b, FAD3a, FAD2b-2, SAD3-1, SAD2-1, SAD2-2, SAD3-2, FAD2a-1, and FAD2a-2 (Supplementary Table S1). It can be assumed that these genes play the most important roles in linseed oil synthesis among the FAD and SAD genes. For these genes, a more detailed analysis of the expression profiles in different flax varieties grown at 16 °C, 20 °C, and 24 °C was performed.
2.3. Expression Profiles of FAD3a and FAD3b Genes during Flax Seed Development under Different Temperature and Watering Conditions
FAD3 genes are known to play a key role in the conversion of LIO to LIN in flax seeds [16,17]. Among the FAD3 genes, the expression of FAD3a and FAD3b was the highest in the studied samples; it was ten times higher than the expression of FAD3c-1, FAD3c-2, FAD3d-1, and FAD3d-2. In different flax varieties, the expression of FAD3a and FAD3b generally changed in a similar manner during seed development under the same growth conditions (Figure 1 and Figure 2, respectively). The expression was minimal at 3 DAF. The maximum expression levels of the FAD3a and FAD3b genes at 16 °C were observed at 21 or 28 DAF in most varieties. At 20 °C, the maximum expression levels were reached at 14 or 21 DAF, after which a decrease was observed. At 24 °C, the maximum expression levels of the FAD3a and FAD3b genes were reached at 14 DAF, after which a decrease was observed. It is noteworthy that the maximum expression levels of the FAD3a and FAD3b genes were similar at 16 °C, 20 °C, and 24 °C. Thus, an increased temperature resulted in a shift in the maximum level of gene expression to earlier stages of seed development, while a decreased temperature resulted in a shift to later stages of seed development.
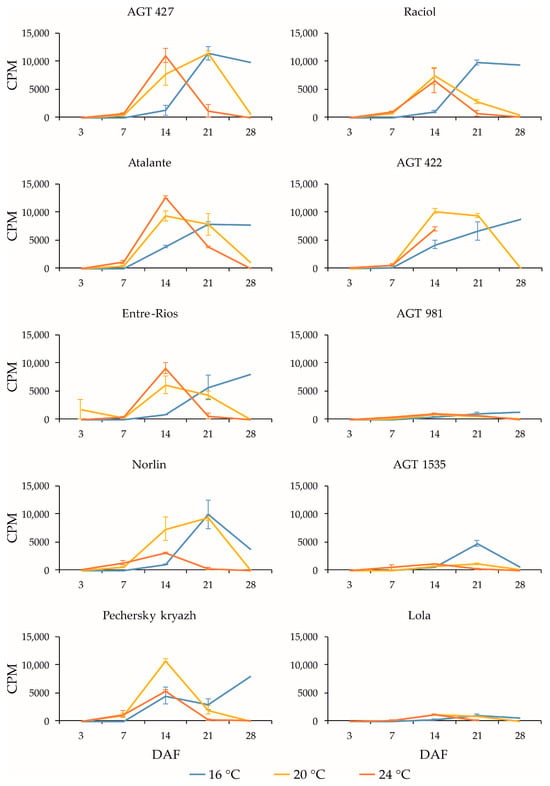
Figure 1.
Expression profiles (3, 7, 14, 21, and 28 DAF) of FAD3a gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Data are missing for 21 and 28 DAF at 24 °C for AGT 422 and 28 DAF at 24 °C for Lola. Error bars represent values obtained for two biological replicates.
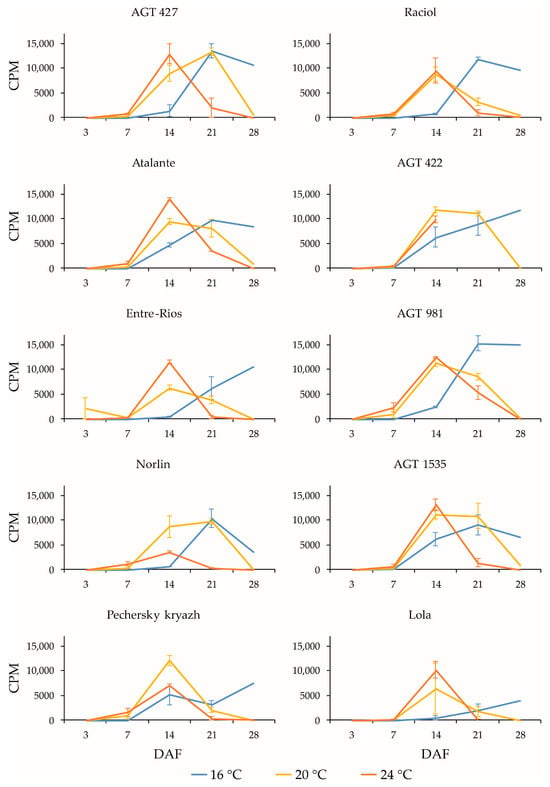
Figure 2.
Expression profiles (3, 7, 14, 21, and 28 DAF) of FAD3b gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Data are missing for 21 and 28 DAF at 24 °C for AGT 422 and 28 DAF at 24 °C for Lola. Error bars represent values obtained for two biological replicates.
The expression profiles of the FAD3a and FAD3b genes were very similar for the same variety under the same conditions (16 °C, 20 °C, and 24 °C). However, in the varieties AGT 981, AGT 1535, and Lola, the level of FAD3a transcript was drastically lower than in the other varieties, while the level of FAD3b transcript was similar to the other varieties. The varieties AGT 981, AGT 1535, and Lola are known to have mutations in the FAD3a gene, resulting in stop codons and a reduced LIN content [7]. Thus, the FAD3a transcript level was reduced in varieties with nonsense mutations in this gene. The situation was different for the FAD3b gene. The varieties AGT 422, AGT 981, AGT 1535, and Raciol, which carry the FAD3b mutation, leading to a histidine-to-tyrosine substitution in exon 2 and a reduction in the LIN content [7], had approximately the same expression level of this gene as the varieties without the mutation. It is worth noting that the expression level of FAD3b was slightly higher than that of FAD3a in all of the varieties studied.
2.4. Expression Profiles of FAD2b-2, FAD2a-1, and FAD2a-2 Genes during Flax Seed Development under Different Temperature and Watering Conditions
FAD2 genes play an important role in the synthesis of linoleic acid from oleic acid [14,15]. Among FAD2 genes, the highest expression levels in flax seeds were found for FAD2b-2, FAD2a-1, and FAD2a-2 (Supplementary Table S1). The expression level of FAD2b-2 was several times higher than those of FAD2a-1 and FAD2a-2 in all varieties studied. The expression level of FAD2a-1 was slightly higher than that of FAD2a-2. It should be noted that the FAD2a-1 and FAD2a-2 genes have very similar sequences, which differ by only seven SNPs, which could introduce some bias in the estimation of their individual expression patterns [19]. For FAD2b-2 (Figure 3), FAD2a-1 (Supplementary Figure S2), and FAD2a-2 (Supplementary Figure S3), as for FAD3a and FAD3b, cultivation at 16 °C resulted in altered expression profiles compared to those at 20 °C and 24 °C and a shift in the maximum expression levels to later stages of seed development (21 or 28 DAF). The expression profiles of FAD2a-1 and FAD2a-2 were very similar for the same variety under the same conditions, but slightly different from that of FAD2b-2.
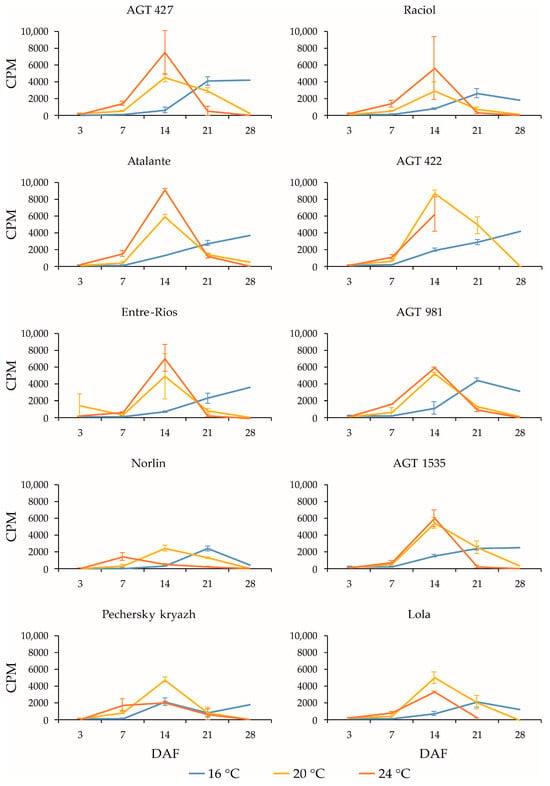
Figure 3.
Expression profiles (3, 7, 14, 21, and 28 DAF) of FAD2b-2 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Data are missing for 21 and 28 DAF at 24 °C for AGT 422 and 28 DAF at 24 °C for Lola. Error bars represent values obtained for two biological replicates.
All varieties showed drastic changes in the expression levels of FAD2b-2, FAD2a-1, and FAD2a-2 during seed development. At 3 DAF, FAD2b-2, FAD2a-1, and FAD2a-2 were barely expressed, but at 14 DAF (for 20 °C and 24 °C) or later (for 16 °C), a dramatic increase in expression levels was observed. At 28 DAF, there was a strong decrease in the expression levels of the genes for all varieties at 20 °C and 24 °C, and there were differences in the expression changes (increase, decrease, or retention compared to 21 DAF) among varieties at 16 °C. A decrease in the maximum expression level of FAD2b-2 at 16 °C compared to 20 °C and/or 24 °C could also be observed for some varieties. Among the 10 examined flax varieties, Pechersky kryazh had the highest OLE content (31.8%), which was 1.5–2.0 times higher than that in other varieties. In this variety, the expression level of FAD2b-2 was slightly lower than that in the majority of studied varieties. The expression level of FAD2b-2 was also slightly lower in Norlin, which had an increased OLE content (22.2%), and Lola, which had a decreased OLE content (12.9%). For FAD2a-1 and FAD2a-2, the differences in expression levels between Pechersky kryazh and other varieties were not very significant. At the same time, the expression profiles of FAD2b-2, FAD2a-1, and FAD2a-2 in Pechersky kryazh were slightly different from those of other varieties, and the maximum expression levels were shifted to earlier stages of seed development (this was clearly visible for FAD2a-1 and FAD2a-2 at 20 °C and for FAD2b-2 at 24 °C). Thus, the high OLE content in the variety Pechersky kryazh is likely not related to the maximum FAD2 expression levels. However, the shift in the maximum expression levels of FAD2 genes to earlier stages of seed development could play a role in increasing the OLE content in linseed oil.
2.5. Expression Levels of SAD Genes during Flax Seed Development under Different Temperature and Watering Conditions
SAD genes are involved in the conversion of stearic acid to oleic acid in flax seeds [11,12,13]. In flax, all four identified SAD genes (SAD2-1, SAD2-2, SAD3-1, and SAD3-2) were rather highly expressed (Supplementary Figure S1), with the highest level being observed for SAD3-1. The expression profiles of SAD2-1 (Figure 4) and SAD2-2 (Supplementary Figure S4) for the same variety under the same conditions were very similar. The same was true for SAD3-1 (Figure 5) and SAD3-2 (Supplementary Figure S5). Some differences in expression profiles were observed between the SAD2-1/SAD2-2 and SAD3-1/SAD3-2 pairs for the same samples, although the overall trend of expression changes was similar. During seed development, an increase in the expression levels of SAD genes was initially observed for all studied conditions. Then, at 20 °C and 24 °C, a decrease in expression levels was observed at 21 or 28 DAF, while at 16 °C, different varieties showed an increase, decrease, or retention of expression levels at 21 and 28 DAF. At the same time, at 20 °C and 24 °C, SAD3-1 and SAD3-2 reached maximum expression levels at earlier stages of seed development (14 DAF for most varieties) compared to SAD2-1 and SAD2-2 (21 DAF at 20 °C and 14 or 21 DAF at 24 °C for most varieties).
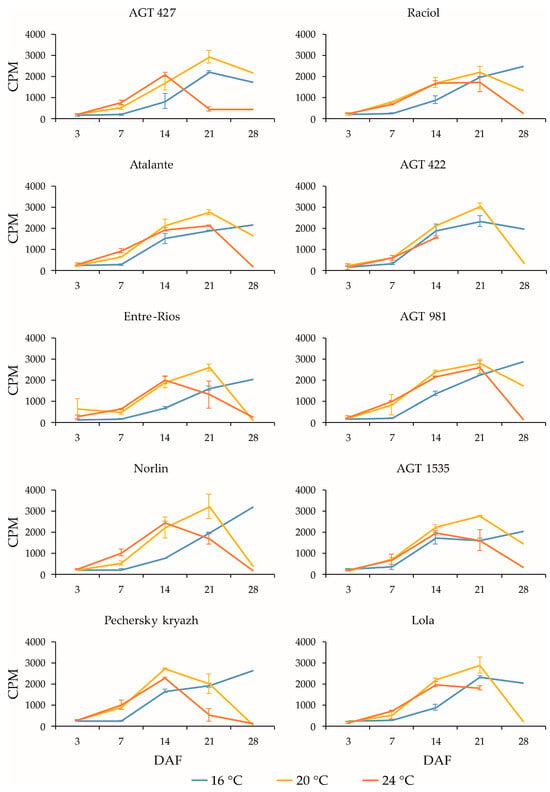
Figure 4.
Expression profiles (3, 7, 14, 21, and 28 DAF) of SAD2-1 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Data are missing for 21 and 28 DAF at 24 °C for AGT 422 and 28 DAF at 24 °C for Lola. Error bars represent values obtained for two biological replicates.
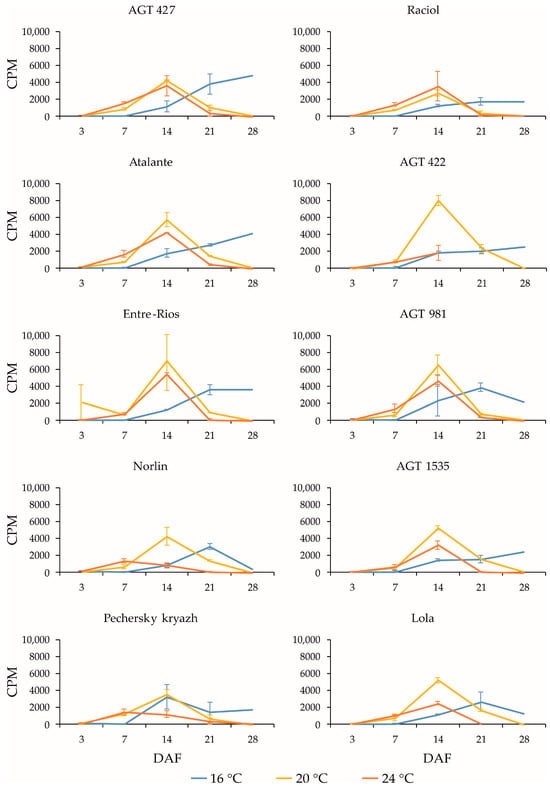
Figure 5.
Expression profiles (3, 7, 14, 21, and 28 DAF) of SAD3-1 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Data are missing for 21 and 28 DAF at 24 °C for AGT 422 and 28 DAF at 24 °C for Lola. Error bars represent values obtained for two biological replicates.
It can be noted that, for SAD2-1/SAD2-2, some expression was observed even at 3 DAF, in contrast to the SAD3-1/SAD3-2 and FAD genes, for which expression was practically absent at this stage. It is also interesting that the expression changes during seed development (most notably at 20 °C) were more similar between SAD3-1/SAD3-2 and FAD2b-2 (reaching maximum levels at earlier stages) and between SAD2-1/SAD2-2 and FAD2a-1/FAD2a-2, FAD3a, and FAD3b (reaching maximum levels at later stages). In Pechersky kryazh with a high OLE content in the oil, characteristics similar to those described above for the FAD2 genes were observed for the SAD genes. Thus, the expression levels of SAD genes in the high-OLE variety Pechersky kryazh were close to those in the low-OLE variety Lola. However, in Pechersky kryazh, the SAD expression profiles were slightly different from those of other varieties, and the maximum expression levels were shifted to earlier stages of seed development (this was particularly noticeable for SAD2-1 and SAD2-2 at 20 °C and for SAD3-1 and SAD3-2 at 24 °C). Thus, the high OLE content in the variety Pechersky kryazh could be related not to its differences from other varieties in the maximum expression levels of SAD and FAD2 genes, but to the shift in the maximum expression levels of these genes to earlier stages of seed development.
3. Discussion
It is known that the key role in the synthesis of unsaturated fatty acids of linseed oil is played by genes of the SAD, FAD2, and FAD3 families [11,12,13,14,15,16,17]. We analyzed the expression of these genes during seed development in a representative set of 10 flax varieties with different oil FA compositions grown under three different temperature and watering conditions. In contrast to most previous works evaluating SAD and FAD gene expression in flax [8,16,17,18,20,21], we obtained individual expression data for all 25 known SAD and FAD genes rather than the expression levels for only a few genes from each family or common expression patterns for pairs of homologous genes. Due to the large number of works devoted to conducting transcriptome analyses of different flax organs and tissues [33,34,35,36,37,38,39,40,41,42,43,44], we were previously able to determine which of the 25 SAD and FAD genes are expressed at high levels in flax seeds and to estimate the expression levels of these genes in other organs and tissues [19]. However, in that work, because of the lack of transcriptomic data, we were unable to assess the dynamics of expression of these genes during seed development; thus, this was carried out in the present work for a representative set of flax varieties grown under different conditions affecting the FA composition of linseed oil.
The highest expression levels were detected for FAD3b, FAD3a, FAD2b-2, SAD3-1, SAD2-1, SAD2-2, SAD3-2, FAD2a-1, and FAD2a-2. In addition, the expression levels of these genes changed tens and hundreds of times during seed development. In the early stages of development, the expression of these genes was minimal, and then a drastic increase was observed, but the dynamics of rise depended significantly on the growth conditions. The strongest increase was observed at 20 °C and 24 °C, with a maximum reached at 14 or 21 DAF in most varieties, followed by a drastic decrease. At 24 °C, compared to 20 °C, a shift in the maximum expression levels to earlier developmental stages was observed for the majority of the highly expressed FAD and SAD genes. At 16 °C, the maximum expression levels of the analyzed genes were observed at later stages of seed development (mainly at 21 and 28 DAF), and the differences between varieties were quite pronounced. It is likely that, among the FAD and SAD families, FAD3b, FAD3a, FAD2b-2, SAD3-1, SAD2-1, SAD2-2, SAD3-2, FAD2a-1, and FAD2a-2 genes with high maximum expression levels and drastic expression changes during seed development play a key role in the synthesis of the FA of linseed oil.
It is also possible to hypothesize how variations in FAD and SAD expression are reflected in the composition of linseed oil when plants are grown under different temperature and moisture conditions. Wet and cold summers are known to increase the LIN + LIO content in linseed oil, whereas hot and dry summers are known to increase the OLE content [10]. In our study, the decrease in FAD2a-1 and FAD2a-2 expression at elevated temperature (24 °C) occurred at earlier stages of seed development compared to 20 °C and, especially, 16 °C. This trend was less clear for FAD2b-2, but it was also present in some genotypes. Under conditions of elevated temperature and insufficient watering, the FAD2 genes probably do not have sufficient time to desaturate oleic acid to linoleic acid to the same extent as at reduced temperature, resulting in increased OLE and decreased LIO+LIN contents. FAD genes are known to be involved in the responses to a variety of stresses, including high and low temperatures [45]. The effects of increased and/or decreased temperatures on the expression of FAD genes were reported for banana [46], maize [47], Gossypium [48], cucumber [49], and some genotypes of soybean [50]. The effect of temperature on the production of LIN and LIO in olive was also shown [51]. For flax, we observed that temperature affects the expression profiles of FAD and SAD genes, which could be reflected in the oil FA composition.
We did not find a correlation between the expression levels of FAD2 or SAD genes and the OLE content. Thus, among the genotypes we studied, there was the high-OLE variety Pechersky kryazh, and expression levels of FAD2 and SAD genes in it were close to those of the low-OLE variety Lola. At the same time, a shift in the maximum expression levels of FAD and SAD genes to earlier stages of seed development was observed in Pechersky kryazh compared to the other varieties studied. This peculiarity could lead to a higher content of OLE in the seeds of this variety. The strongest expression of FAD and SAD genes in this variety may occur in hotter summer periods than in other varieties, which could further enhance the effects of an increasing OLE content and a decreasing LIO content. We grew flax plants under controlled conditions at constant temperatures, so differences in the flowering time did not bias the evaluation of the expression levels. However, when plants are grown under field conditions, the contribution of differences in flowering time between genotypes could also be reflected in the oil FA composition, as seed maturation may occur under different temperature/watering conditions.
Among the SAD genes, the highest expression level was found for SAD3-1. The expression level of its homolog, SAD3-2, was, on average, two times lower. The expression levels of the homologous genes SAD2-1 and SAD2-2 were quite high, not more than 30% lower than that of SAD3-1. The expression level of SAD2-1 was slightly higher than that of SAD2-2. It can be assumed that the SAD3-1 gene plays the major role in the conversion of STE to OLE in flax seeds, but SAD2-1, SAD2-2, and SAD3-2 also contribute significantly to this process.
Among the FAD2 genes, FAD2b-2 had the highest expression level in most samples. However, the expression level of its homolog, FAD2b-1, was tens of times lower. For the homologous genes FAD2a-1 and FAD2a-2, the expression levels were quite similar, with a slightly higher level of FAD2a-1. FAD2b-2 is probably the key gene in the desaturation of oleic acid to linoleic acid in flax seeds, but FAD2a-1 and FAD2a-2 also contribute to this process. The role of FAD2b-1 in linoleic acid synthesis in linseed oil is probably insignificant, which is also characteristic of the other FAD2 genes studied, namely FAD2c-1, FAD2c-2, FAD2d-1, FAD2d-2, FAD2e-1, FAD2e-2, FAD2f-1, FAD2f-2, FAD2g-1, FAD2g-2, and FAD2h.
Among the FAD3 genes, FAD3a and FAD3b were characterized by the highest expression levels. The expression levels of these genes were similar, but in the varieties AGT 981, AGT 1535, and Lola with nonsense mutations in the FAD3a gene, leading to a reduced LIN content, the level of the FAD3a transcript was ten times lower than the level of the FAD3b transcript. The low level of the FAD3a transcript in the flax genotype with a mutation in this gene, leading to a stop codon, was previously reported and explained by nonsense-mediated mRNA decay [17]. No decrease in the FAD3b transcript level was detected in varieties with a missense mutation in FAD3b, leading to a reduction in the LIN content. The fact that a missense mutation in the FAD3b gene does not lead to a change in its expression compared to other varieties was also reported previously [17]. It was also shown that several FAD3a and FAD3b gene isoforms carrying mutations encode non-functional enzymes that are unable to convert LIO to LIN [26]. The contribution of FAD3a and FAD3b to the formation of LIN from LIO is probably close, but the expression level of FAD3b was slightly higher than that of FAD3a, so the role of FAD3b in the synthesis of linolenic acid might be slightly higher. This assumption is supported by the higher LIN content in flax varieties with inactivating mutations in the FAD3a gene compared to varieties with inactivating mutations in the FAD3b gene [7].
We observed similar expression profiles for SAD3-1, SAD3-2, and FAD2b-2. The expression profiles of SAD2-1, SAD2-2, FAD2a-1, FAD2a-2, FAD3a, and FAD3b were also similar. For the first group of genes, maximum expression levels were reached at earlier stages of seed development compared to the second group. There may be common regulatory mechanisms for the genes in each group, and these groups of genes may contribute somewhat differently to the synthesis of linseed oil at different stages of seed development.
Thus, by analyzing more than two hundred cDNA libraries of flax seeds at five stages of development for a representative set of ten flax varieties grown under three different environmental conditions, we were able to identify the key genes and stages of the FA synthesis of linseed oil and evaluate the influence of environmental conditions and genotype on the expression of these genes. FAD and SAD genes with the highest expression in flax seeds were identified, and these genes probably play important roles in the synthesis of OLE, LIO, and LIN in flax seeds. SAD3-1, SAD2-1, SAD2-2, and SAD3-2 contribute to the synthesis of OLE from STE. Among the fifteen FAD2 genes, the FAD2b-2 gene is probably the key gene in the desaturation of OLE to LIO, but FAD2a-1 and FAD2a-2 also contribute to this process. Among the six FAD3 genes, FAD3a and FAD3b play major roles in the conversion of LIO to LIN. The obtained data, in addition to their basic significance, are important for the selection of FAD and SAD genes for editing to obtain flax varieties with modified FA compositions to meet the requirements of different economic sectors. For the FAD3a gene, a correlation was found between the transcript level and the LIN content, which was associated with nonsense mutations in this gene. This feature could be used in the marker-assisted selection of flax to develop varieties with reduced linolenic acid contents for the food industry. The analyzed FAD and SAD genes showed dramatic changes in the expression levels during seed development, and the expression profiles of these genes were quite similar during seed development under the same growth conditions. At the same time, the growth conditions contributed significantly to the expression profiles: under elevated temperature and insufficient watering, maximum expression levels were reached at earlier stages, and an earlier decrease in expression during seed development was observed, while reduced temperature and excessive watering shifted the maximum expression levels to later stages of seed development. Thus, 14 DAF was the key for FA synthesis in flax seeds at 24 °C with underwatering, and 14 and 21 DAF were the keys at 20 °C with optimal watering, while 21 and 28 DAF turned out to be the most important at 16 °C with overwatering. These expression changes could be reflected in the FA composition of linseed oil. In addition, genotype-dependent features of FAD and SAD expression profiles were found, which were more pronounced at 16 °C with overwatering. Moreover, the shift in expression profiles to earlier stages of seed development in the variety Pechersky kryazh compared to the other varieties could result in a high OLE content in its linseed oil under certain conditions. Thus, our work is important for understanding the contribution of genotype and environment to the expression of FAD and SAD genes, which can be used to develop recommendations for optimal conditions of linseed cultivation to obtain oil with a specific FA composition.
4. Materials and Methods
4.1. Flax Varieties
We used plants of 10 flax varieties: AGT 422, AGT 427, AGT 981, AGT 1535, Atalante, Entre-Rios, Lola, Norlin, Pechersky kryazh (namely, l. 1-2 from k-2889 according to the catalog of the Federal Research Center for Bast Fiber Crops, selected from Pechersky kryazh, originating from N.I. Vavilov All-Russian Institute of Plant Genetic Resources (VIR)), and Raciol. AGT 981, AGT 1535, and Lola had low LIN contents; Raciol and AGT 422 had medium LIN contents; and AGT 427, Entre-Rios, Norlin, Atalante, and Pechersky kryazh had high LIN contents (Table 1). Low and medium LIN levels were associated with mutations in the FAD3a and FAD3b genes [7]; the data are shown in Table 1.
4.2. Growth Conditions
Flax plants were grown in 15 L pots with soil pre-treated with fungicide. pH was maintained at about 5.5, which is the optimum level for flax. Thirty-two seeds were sown per pot, and three pots were planted for each variety. All plants were maintained under the same conditions for one month after planting. The plants were then transferred to three climate chambers with different temperature regimes for growth. In the first chamber, the temperature was maintained at 16 °C and watering was carried out every day. In the second chamber, the temperature was 20 °C and watering was carried out every two days. In the third chamber, the temperature was 24 °C and watering was carried out every three days.
4.3. Collection of Plant Material and RNA Isolation
The flower was marked with the date of the day it opened (day of flowering) and the data were recorded in a table. Seeds were collected at 3, 7, 14, 21, and 28 days after flowering (DAF). For each variety, at least 10 samples were collected for each temperature/watering condition and each developmental stage. Seeds collected from individual capsules were placed in tubes and immediately frozen in liquid nitrogen. Plant material was placed in a low-temperature freezer at −70 °C.
For RNA isolation, seeds were first ground to a fine powder using disposable homogenization pestles (Helicon, Moscow, Russia) inserted into a DeWalt DCD701D2 cordless drill (DeWalt, Towson, MD, USA) at a speed of 1200–1500 rpm in 1.5 mL tubes in liquid nitrogen without thawing the sample. RNA isolation was performed according to the protocol of Wang et al. [52] with some modifications. Briefly, 1 mL CTAB lysis buffer (2% CTAB (neoFroxx GmbH, Einhausen, Germany), 2% PVP K30 (PanReac AppliChem, Darmstadt, Germany), 100 mM Tris HCl pH 8.0 (Thermo Fisher Scientific, Waltham, MA, USA), 25 mM EDTA (Thermo Fisher Scientific), 2 M NaCl (Scharlab, Barcelona, Spain), and 2% β-mercaptoethanol (Bio-Rad, Hercules, CA, USA)) pre-warmed to 65 °C were added to the homogenized sample. The homogenate was incubated in a TDB-120 thermostat (Biosan, Riga, Latvia) at 65 °C for 30 min with mixing every 10 min. Then, five identical samples (same variety, growth conditions, and developmental stage) were mixed as follows: 350 μL of homogenate was taken from each sample using a 1000 mL wide-bore pipette tip and added to a 2 mL tube. Next, 500 μL of homogenate was taken from the pool (the remainder was frozen in liquid nitrogen), and 500 μL of CTAB lysis buffer pre-warmed to 65 °C was added. In the next step, an equal volume of cold chloroform (Acros Organics, Geel, Belgium) was added to the homogenate, vortexed for 30 s, and then centrifuged for 20 min at 10,000× g and 4 °C in a 5418R microcentrifuge (Eppendorf, Hamburg, Germany). The aqueous phase was transferred to clean tubes, and then an equal volume of cold chloroform was added, vortexed for 30 s, and centrifuged for 10 min at 10,000× g and 4 °C. The aqueous phase was transferred to clean tubes, and 1/2 volume of 96% ethanol was added and mixed until homogeneous. Total RNA was then purified using the CleanRNA Standard Kit (Evrogen, Moscow, Russia) according to the manufacturer’s protocol with a DNAase I treatment step from the RNase-Free DNase Set (Qiagen, Chatsworth, CA, USA). RNA quality and concentration were evaluated using a gel electrophoresis, 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA), and Qubit 4 fluorometer (Thermo Fisher Scientific). Only non-degraded RNA samples with a concentration of at least 20 ng/μL were used for further work.
4.4. Preparation and Sequencing of cDNA Libraries
cDNA libraries for sequencing of flax seed transcriptomes were prepared using the QIAseq Stranded mRNA Select Kit (Qiagen) according to the manufacturer’s protocol. RNA from seeds of 10 flax varieties (AGT 422, AGT 427, AGT 981, AGT 1535, Atalante, Entre-Rios, Lola, Norlin, Pechersky kryazh, and Raciol) collected at 3, 7, 14, 21, and 28 DAF from plants grown under three different temperature/watering conditions were used. For 3, 7, 14, and 21 DAF, cDNA libraries were prepared in 2 replicates, and for 28 DAF, cDNA libraries were prepared in 1 replicate (because of the difficulty of obtaining RNA of high quality and sufficient quantity for seeds of late developmental stages). The quality of the obtained cDNA libraries (agreement of the length of the obtained libraries with the expected ones and the absence of adapter dimers) was evaluated on a 2100 Bioanalyzer (Agilent Technologies), and the concentration was evaluated on a Qubit 4 fluorometer (Thermo Fisher Scientific).
The resulting cDNA libraries (266 libraries in total, as the libraries for the variety AGT 422 at 21 and 28 DAF at 24 °C and the variety Lola at 28 DAF at 24 °C were excluded from the analysis because of their low quality) were mixed equimolarly and sequenced on a NextSeq 2000 instrument (Illumina, San Diego, CA, USA) using the NextSeq 2000 P3 Reagents (100 Cycles) kit (Illumina) in 51 + 51 nucleotide format.
4.5. Expression Analysis
Raw reads were processed using Trimmomatic 0.38 [53]. The data were checked for the presence of adapter sequences, trimmed for quality (TRAILING:24 and SLIDINGWINDOW:4:14), and filtered for length (MINLEN:40). The PPline tool was then used for expression analysis [54]. Transcriptomic reads were aligned to the variety Atlant genome (GCA_014858635.1 in NCBI Genome) [55], and counts per million (CPM) values were calculated for 25 genes: SAD2-1 (Atlant transcript H1233_031351), SAD2-2 (H1233_038408), SAD3-1 (H1233_054424), SAD3-2 (H1233_039970), FAD2a-1 (H1233_058938), FAD2a-2 (H1233_061927), FAD2b-1 (H1233_075799), FAD2b-2 (H1233_078572), FAD2c-1 (H1233_075801), FAD2c-2 (H1233_078571), FAD2d-1 (H1233_041596), FAD2d-2 (H1233_078570), FAD2e-1 (H1233_075803), FAD2e-2 (H1233_078569), FAD2f-1 (H1233_075804), FAD2f-2 (H1233_078567), FAD2g-1 (H1233_075805), FAD2g-2 (H1233_078566), FAD2h (H1233_078565), FAD3a (H1233_027729), FAD3b (H1233_038272), FAD3c-1 (H1233_054813), FAD3c-2 (H1233_039845), FAD3d-1 (H1233_041596), and FAD3d-2 (H1233_041890).
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/plants13070956/s1, Figure S1: Clustering of seed samples collected at five time points (3 (3d), 7 (7d), 14 (14d), 21 (21d), and 28 (28d) days after flowering) for ten flax varieties (AGT 422 (422), AGT 427 (427), AGT 981 (981), AGT 1535 (1535), Atalante (Ata), Entre-Rios (Ent), Lola (Lol), Norlin (Nor), Pechersky kryazh (Pech), and Raciol (Rac)) grown under three different temperature/watering conditions (16 °C and overwatering (16C), 20 °C and optimal watering (20C), 24 °C and underwatering (24C)) based on gene expression profiles. Figure S2: Expression profiles (3, 7, 14, 21, and 28 DAF) of FAD2a-1 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Figure S3: Expression profiles (3, 7, 14, 21, and 28 DAF) of FAD2a-2 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Figure S4: Expression profiles (3, 7, 14, 21, and 28 DAF) of SAD2-2 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C). Figure S5: Expression profiles (3, 7, 14, 21, and 28 DAF) of SAD3-2 gene in high-LIN (AGT 427, Atalante, Entre-Rios, Norlin, and Pechersky kryazh), mid-LIN (Raciol and AGT 422), and low-LIN (AGT 981, AGT 1535, and Lola) flax varieties grown at 16 °C and overwatered (16 °C), at 20 °C and optimally watered (20 °C), and at 24 °C and underwatered (24 °C); Table S1: Expression levels of SAD (SAD2-1, SAD2-2, SAD3-1, and SAD3-2), FAD2 (FAD2a-1, FAD2a-2, FAD2b-1, FAD2b-2, FAD2c-1, FAD2c-2, FAD2d-1, FAD2d-2, FAD2e-1, FAD2e-2, FAD2f-1, FAD2f-2, and FAD2g-2), and FAD3 (FAD3a, FAD3b, FAD3c-1, FAD3c-2, FAD3d-1, and FAD3d-2) genes in seeds (3, 7, 14, 21, and 28 DAF) of flax plants of 10 varieties (AGT 422 (422), AGT 427 (427), AGT 981 (981), AGT 1535 (1535), Atalante (Ata), Entre-Rios (Ent), Lola (Lol), Norlin (Nor), Pechersky kryazh (Pech), and Raciol (Rac)) grown under different temperature and watering conditions (16 °C, 20 °C, and 24 °C).
Author Contributions
Conceptualization, T.A.R., A.A.D. and N.V.M.; performing experiments, E.N.P., L.V.P., R.O.N., T.A.R., A.A.G., D.A.Z., E.V.B., A.A.T., A.G.Y. and N.L.B.; data analysis, E.M.D., E.A.S., A.A.D. and N.V.M.; writing, E.N.P., E.M.D., A.A.D. and N.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Russian Science Foundation, grant 21-16-00111.
Data Availability Statement
The raw sequencing data have been deposited in the NCBI Sequence Read Archive (SRA) under the BioProject accession number PRJNA1039849.
Acknowledgments
We thank the Center for Precision Genome Editing and Genetic Technologies for Biomedicine, EIMB RAS for providing the computing power and techniques for the data analysis. This work was performed using the equipment of the EIMB RAS “Genome” center (http://www.eimb.ru/ru1/ckp/ccu_genome_ce.php (accessed on 4 December 2023)).
Conflicts of Interest
The authors declare no conflicts of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Parikh, M.; Maddaford, T.G.; Austria, J.A.; Aliani, M.; Netticadan, T.; Pierce, G.N. Dietary Flaxseed as a Strategy for Improving Human Health. Nutrients 2019, 11, 1171. [Google Scholar] [CrossRef]
- Saini, R.K.; Prasad, P.; Sreedhar, R.V.; Akhilender Naidu, K.; Shang, X.; Keum, Y.S. Omega-3 Polyunsaturated Fatty Acids (PUFAs): Emerging Plant and Microbial Sources, Oxidative Stability, Bioavailability, and Health Benefits-A Review. Antioxidants 2021, 10, 1627. [Google Scholar] [CrossRef]
- Al-Madhagy, S.; Ashmawy, N.S.; Mamdouh, A.; Eldahshan, O.A.; Farag, M.A. A comprehensive review of the health benefits of flaxseed oil in relation to its chemical composition and comparison with other omega-3-rich oils. Eur. J. Med. Res. 2023, 28, 240. [Google Scholar] [CrossRef]
- Campos, J.R.; Severino, P.; Ferreira, C.S.; Zielinska, A.; Santini, A.; Souto, S.B.; Souto, E.B. Linseed Essential Oil-Source of Lipids as Active Ingredients for Pharmaceuticals and Nutraceuticals. Curr. Med. Chem. 2019, 26, 4537–4558. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, S.; Ragupathy, R.; Niu, Z.; Duguid, S. SSR-based linkage map of flax (Linum usitatissimum L.) and mapping of QTLs underlying fatty acid composition traits. Mol. Breed. 2011, 28, 437–451. [Google Scholar] [CrossRef]
- Muir, A.D.; Westcott, N.D. Flax: The Genus Linum; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Dmitriev, A.A.; Kezimana, P.; Rozhmina, T.A.; Zhuchenko, A.A.; Povkhova, L.V.; Pushkova, E.N.; Novakovskiy, R.O.; Pavelek, M.; Vladimirov, G.N.; Nikolaev, E.N.; et al. Genetic diversity of SAD and FAD genes responsible for the fatty acid composition in flax cultivars and lines. BMC Plant Biol. 2020, 20, 301. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, X.R.; Zhang, Z.J.; Dribnenki, P.; Singh, S.; Green, A. Development of high oleic oil crop platform in flax through RNAi-mediated multiple FAD2 gene silencing. Plant Cell Rep. 2015, 34, 643–653. [Google Scholar] [CrossRef]
- Brutch, N.; Porokhoviniva, E.; Shelenga, T. Perspectives of the creation of oil flax varieties for the specialized purpose. Agrar. Report. South-East 2016, 1, 50–52. [Google Scholar]
- Porokhovinova, E.; Shelenga, T.; Kosykh, L.; Sanin, A.; Kazarina, A.; Kutuzova, S.; Pavlov, A.; Brutch, N. Biochemical diversity of fatty acid composition in flax from VIR genetic collection and effect of environment on its development. Ecol. Genet. 2016, 14, 13. [Google Scholar] [CrossRef][Green Version]
- Jain, R.K.; Thompson, R.G.; Taylor, D.C.; MacKenzie, S.L.; McHughen, A.; Rowland, G.G.; Tenaschuk, D.; Coffey, M. Isolation and characterization of two promoters from linseed for genetic engineering. Crop Sci. 1999, 39, 1696–1701. [Google Scholar] [CrossRef]
- Singh, S.; McKinney, S.; Green, A. Sequence of a cDNA from Linum usitatissimum encoding the stearoyl-acyl carrier protein desaturase. Plant Physiol. 1994, 104, 1075. [Google Scholar] [CrossRef][Green Version]
- Fofana, B.; Duguid, S.; Cloutier, S. Cloning of fatty acid biosynthetic genes β-ketoacyl CoA synthase, fatty acid elongase, stearoyl-ACP desaturase, and fatty acid desaturase and analysis of expression in the early developmental stages of flax (Linum usitatissimum L.) seeds. Plant Sci. 2004, 166, 1487–1496. [Google Scholar] [CrossRef]
- Khadake, R.M.; Ranjekar, P.K.; Harsulkar, A.M. Cloning of a novel omega-6 desaturase from flax (Linum usitatissimum L.) and its functional analysis in Saccharomyces cerevisiae. Mol. Biotechnol. 2009, 42, 168–174. [Google Scholar] [CrossRef]
- Krasowska, A.; Dziadkowiec, D.; Polinceusz, A.; Plonka, A.; Łukaszewicz, M. Cloning of flax oleic fatty acid desaturase and its expression in yeast. J. Am. Oil Chem. Soc. 2007, 84, 809–816. [Google Scholar] [CrossRef]
- Vrinten, P.; Hu, Z.; Munchinsky, M.A.; Rowland, G.; Qiu, X. Two FAD3 desaturase genes control the level of linolenic acid in flax seed. Plant Physiol. 2005, 139, 79–87. [Google Scholar] [CrossRef]
- Banik, M.; Duguid, S.; Cloutier, S. Transcript profiling and gene characterization of three fatty acid desaturase genes in high, moderate, and low linolenic acid genotypes of flax (Linum usitatissimum L.) and their role in linolenic acid accumulation. Genome 2011, 54, 471–483. [Google Scholar] [CrossRef]
- You, F.M.; Li, P.; Kumar, S.; Ragupathy, R.; Li, Z.; Fu, Y.-B.; Cloutier, S. Genome-wide identification and characterization of the gene families controlling fatty acid biosynthesis in flax (Linum usitatissimum L). J. Proteom. Bioinform. 2014, 7, 310–326. [Google Scholar]
- Dvorianinova, E.M.; Zinovieva, O.L.; Pushkova, E.N.; Zhernova, D.A.; Rozhmina, T.A.; Povkhova, L.V.; Novakovskiy, R.O.; Sigova, E.A.; Turba, A.A.; Borkhert, E.V.; et al. Key FAD2, FAD3, and SAD Genes Involved in the Fatty Acid Synthesis in Flax Identified Based on Genomic and Transcriptomic Data. Int. J. Mol. Sci. 2023, 24, 14885. [Google Scholar] [CrossRef]
- Rajwade, A.V.; Kadoo, N.Y.; Borikar, S.P.; Harsulkar, A.M.; Ghorpade, P.B.; Gupta, V.S. Differential transcriptional activity of SAD, FAD2 and FAD3 desaturase genes in developing seeds of linseed contributes to varietal variation in alpha-linolenic acid content. Phytochemistry 2014, 98, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, D.; Cloutier, S. Fatty acid composition and desaturase gene expression in flax (Linum usitatissimum L.). J. Appl. Genet. 2014, 55, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Rajwade, A.V.; Joshi, R.S.; Kadoo, N.Y.; Gupta, V.S. Sequence characterization and in silico structure prediction of fatty acid desaturases in linseed varieties with differential fatty acid composition. J. Sci. Food Agric. 2016, 96, 4896–4906. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, D.; Duguid, S.; Loewen, E.; Rowland, G.; Booker, H.; You, F.M.; Cloutier, S. Genetic variation of six desaturase genes in flax and their impact on fatty acid composition. TAG. Theor. Appl. Genet. Theor. Und Angew. Genet. 2013, 126, 2627–2641. [Google Scholar] [CrossRef] [PubMed]
- Porokhovinova, E.A.; Shelenga, T.V.; Matveeva, T.V.; Pavlov, A.V.; Grigorieva, E.A.; Brutch, N.B. Polymorphism of genes controlling low level of linolenic acid in lines from VIR flax genetic collection. Ecol. Genet. 2019, 17, 5–19. [Google Scholar] [CrossRef]
- Povkhova, L.V.; Pushkova, E.N.; Rozhmina, T.A.; Zhuchenko, A.A.; Frykin, R.I.; Novakovskiy, R.O.; Dvorianinova, E.M.; Gryzunov, A.A.; Borkhert, E.V.; Sigova, E.A.; et al. Development and Complex Application of Methods for the Identification of Mutations in the FAD3A and FAD3B Genes Resulting in the Reduced Content of Linolenic Acid in Flax Oil. Plants 2022, 12, 95. [Google Scholar] [CrossRef] [PubMed]
- Radovanovic, N.; Thambugala, D.; Duguid, S.; Loewen, E.; Cloutier, S. Functional characterization of flax fatty acid desaturase FAD2 and FAD3 isoforms expressed in yeast reveals a broad diversity in activity. Mol. Biotechnol. 2014, 56, 609–620. [Google Scholar] [CrossRef]
- Xie, D.; Dai, Z.; Yang, Z.; Tang, Q.; Deng, C.; Xu, Y.; Wang, J.; Chen, J.; Zhao, D.; Zhang, S.; et al. Combined genome-wide association analysis and transcriptome sequencing to identify candidate genes for flax seed fatty acid metabolism. Plant Sci. Int. J. Exp. Plant Biol. 2019, 286, 98–107. [Google Scholar] [CrossRef] [PubMed]
- You, F.M.; Xiao, J.; Li, P.; Yao, Z.; Jia, G.; He, L.; Kumar, S.; Soto-Cerda, B.; Duguid, S.D.; Booker, H.M.; et al. Genome-Wide Association Study and Selection Signatures Detect Genomic Regions Associated with Seed Yield and Oil Quality in Flax. Int. J. Mol. Sci. 2018, 19, 2303. [Google Scholar] [CrossRef] [PubMed]
- Marketta, S.; Juha-Matti, P.; Merja, E.; Ari, K.; Lauri, J.; Veli, H. Yield, SDG lignan, cadmium, lead, oil and protein contents of linseed (Linum usitatissimum L.) cultivated in trials and at different farm conditions in the south-western part of Finland. Agric. Food Sci. 2013, 22, 296–306. [Google Scholar] [CrossRef]
- Dybing, C.D.; Zimmerman, D.C. Fatty Acid accumulation in maturing flaxseeds as influenced by environment. Plant Physiol. 1966, 41, 1465–1470. [Google Scholar] [CrossRef]
- Fofana, B.; Cloutier, S.; Duguid, S.; Ching, J.; Rampitsch, C. Gene expression of stearoyl-ACP desaturase and delta12 fatty acid desaturase 2 is modulated during seed development of flax (Linum usitatissimum). Lipids 2006, 41, 705–712. [Google Scholar] [CrossRef]
- Khadake, R.; Khonde, V.; Mhaske, V.; Ranjekar, P.; Harsulkar, A. Functional and bioinformatic characterisation of sequence variants of Fad3 gene from flax. J. Sci. Food Agric. 2011, 91, 2689–2696. [Google Scholar] [CrossRef]
- Dash, P.K.; Rai, R.; Mahato, A.K.; Gaikwad, K.; Singh, N.K. Transcriptome Landscape at Different Developmental Stages of a Drought Tolerant Cultivar of Flax (Linum usitatissimum). Front. Chem. 2017, 5, 82. [Google Scholar] [CrossRef]
- Dmitriev, A.A.; Kudryavtseva, A.V.; Krasnov, G.S.; Koroban, N.V.; Speranskaya, A.S.; Krinitsina, A.A.; Belenikin, M.S.; Snezhkina, A.V.; Sadritdinova, A.F.; Kishlyan, N.V.; et al. Gene expression profiling of flax (Linum usitatissimum L.) under edaphic stress. BMC Plant Biol. 2016, 16, 237. [Google Scholar] [CrossRef]
- Galindo-Gonzalez, L.; Deyholos, M.K. RNA-seq Transcriptome Response of Flax (Linum usitatissimum L.) to the Pathogenic Fungus Fusarium oxysporum f. sp. lini. Front. Plant Sci. 2016, 7, 1766. [Google Scholar] [CrossRef]
- Dmitriev, A.A.; Krasnov, G.S.; Rozhmina, T.A.; Novakovskiy, R.O.; Snezhkina, A.V.; Fedorova, M.S.; Yurkevich, O.Y.; Muravenko, O.V.; Bolsheva, N.L.; Kudryavtseva, A.V.; et al. Differential gene expression in response to Fusarium oxysporum infection in resistant and susceptible genotypes of flax (Linum usitatissimum L.). BMC Plant Biol. 2017, 17, 253. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, A.A.; Novakovskiy, R.O.; Pushkova, E.N.; Rozhmina, T.A.; Zhuchenko, A.A.; Bolsheva, N.L.; Beniaminov, A.D.; Mitkevich, V.A.; Povkhova, L.V.; Dvorianinova, E.M.; et al. Transcriptomes of Different Tissues of Flax (Linum usitatissimum L.) Cultivars With Diverse Characteristics. Front. Genet. 2020, 11, 565146. [Google Scholar] [CrossRef]
- Dmitriev, A.A.; Krasnov, G.S.; Rozhmina, T.A.; Zyablitsin, A.V.; Snezhkina, A.V.; Fedorova, M.S.; Pushkova, E.N.; Kezimana, P.; Novakovskiy, R.O.; Povkhova, L.V.; et al. Flax (Linum usitatissimum L.) response to non-optimal soil acidity and zinc deficiency. BMC Plant Biol. 2019, 19, 54. [Google Scholar] [CrossRef]
- Petrova, A.; Kozlova, L.; Gorshkov, O.; Nazipova, A.; Ageeva, M.; Gorshkova, T. Cell Wall Layer Induced in Xylem Fibers of Flax Upon Gravistimulation Is Similar to Constitutively Formed Cell Walls of Bast Fibers. Front. Plant Sci. 2021, 12, 660375. [Google Scholar] [CrossRef]
- Gorshkov, O.; Chernova, T.; Mokshina, N.; Gogoleva, N.; Suslov, D.; Tkachenko, A.; Gorshkova, T. Intrusive Growth of Phloem Fibers in Flax Stem: Integrated Analysis of miRNA and mRNA Expression Profiles. Plants 2019, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Gorshkova, T.; Chernova, T.; Mokshina, N.; Gorshkov, V.; Kozlova, L.; Gorshkov, O. Transcriptome Analysis of Intrusively Growing Flax Fibers Isolated by Laser Microdissection. Sci. Rep. 2018, 8, 14570. [Google Scholar] [CrossRef] [PubMed]
- Gorshkov, O.; Mokshina, N.; Ibragimova, N.; Ageeva, M.; Gogoleva, N.; Gorshkova, T. Phloem fibres as motors of gravitropic behaviour of flax plants: Level of transcriptome. Funct. Plant Biol. 2018, 45, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Gorshkov, O.; Mokshina, N.; Gorshkov, V.; Chemikosova, S.; Gogolev, Y.; Gorshkova, T. Transcriptome portrait of cellulose-enriched flax fibres at advanced stage of specialization. Plant Mol. Biol. 2017, 93, 431–449. [Google Scholar] [CrossRef]
- Gao, P.; Qiu, S.; Ma, X.; Parkin, I.A.P.; Xiang, D.; Datla, R. Spatiotemporal Transcriptomic Atlas of Developing Embryos and Vegetative Tissues in Flax. Plants 2022, 11, 2031. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zou, Y.; Guo, X.; Li, H.; Lu, H. Fatty acid desaturases (FADs) modulate multiple lipid metabolism pathways to improve plant resistance. Mol. Biol. Rep. 2022, 49, 9997–10011. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Liu, F.; Sun, X.; Wang, B.; Liu, J.; Ni, X.; Hu, C.; Deng, G.; Tong, Z.; Zhang, Y.; et al. Genome-wide identification of FAD gene family and their contributions to the temperature stresses and mutualistic and parasitic fungi colonization responses in banana. Int. J. Biol. Macromol. 2022, 204, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wei, J.; He, L.; Zhang, Y.; Zhao, Y.; Xu, X.; Wei, Y.; Ge, S.; Ding, D.; Liu, M.; et al. Identification of Fatty Acid Desaturases in Maize and Their Differential Responses to Low and High Temperature. Genes 2019, 10, 445. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, O.P.; Park, S.; Ilut, D.C.; Inmon, J.J.; Millhollon, J.C.; Liechty, Z.; Page, J.T.; Jenks, M.A.; Chapman, K.D.; Udall, J.A.; et al. Genome-wide analysis of the omega-3 fatty acid desaturase gene family in Gossypium. BMC Plant Biol. 2014, 14, 312. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.J.; Cao, N.; Zhang, Z.G.; Shang, Q.M. Characterization of the Fatty Acid Desaturase Genes in Cucumber: Structure, Phylogeny, and Expression Patterns. PLoS ONE 2016, 11, e0149917. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Zoong-Lwe, Z.S.; Gandhi, N.; Welti, R.; Fallen, B.; Smith, J.R.; Rustgi, S. Comparative Lipidomic Analysis Reveals Heat Stress Responses of Two Soybean Genotypes Differing in Temperature Sensitivity. Plants 2020, 9, 457. [Google Scholar] [CrossRef]
- D’Angeli, S.; Altamura, M.M. Unsaturated Lipids Change in Olive Tree Drupe and Seed during Fruit Development and in Response to Cold-Stress and Acclimation. Int. J. Mol. Sci. 2016, 17, 1889. [Google Scholar] [CrossRef]
- Wang, L.; Stegemann, J.P. Extraction of high quality RNA from polysaccharide matrices using cetyltrimethylammonium bromide. Biomaterials 2010, 31, 1612–1618. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Krasnov, G.S.; Dmitriev, A.A.; Kudryavtseva, A.V.; Shargunov, A.V.; Karpov, D.S.; Uroshlev, L.A.; Melnikova, N.V.; Blinov, V.M.; Poverennaya, E.V.; Archakov, A.I.; et al. PPLine: An Automated Pipeline for SNP, SAP, and Splice Variant Detection in the Context of Proteogenomics. J. Proteome Res. 2015, 14, 3729–3737. [Google Scholar] [CrossRef]
- Dmitriev, A.A.; Pushkova, E.N.; Novakovskiy, R.O.; Beniaminov, A.D.; Rozhmina, T.A.; Zhuchenko, A.A.; Bolsheva, N.L.; Muravenko, O.V.; Povkhova, L.V.; Dvorianinova, E.M.; et al. Genome Sequencing of Fiber Flax Cultivar Atlant Using Oxford Nanopore and Illumina Platforms. Front. Genet. 2020, 11, 590282. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).