
Research on Physiological Characteristics and Differential Gene Expression of Rice Hybrids and Their Parents under Salt Stress at Seedling Stage
Abstract
1. Introduction
2. Results
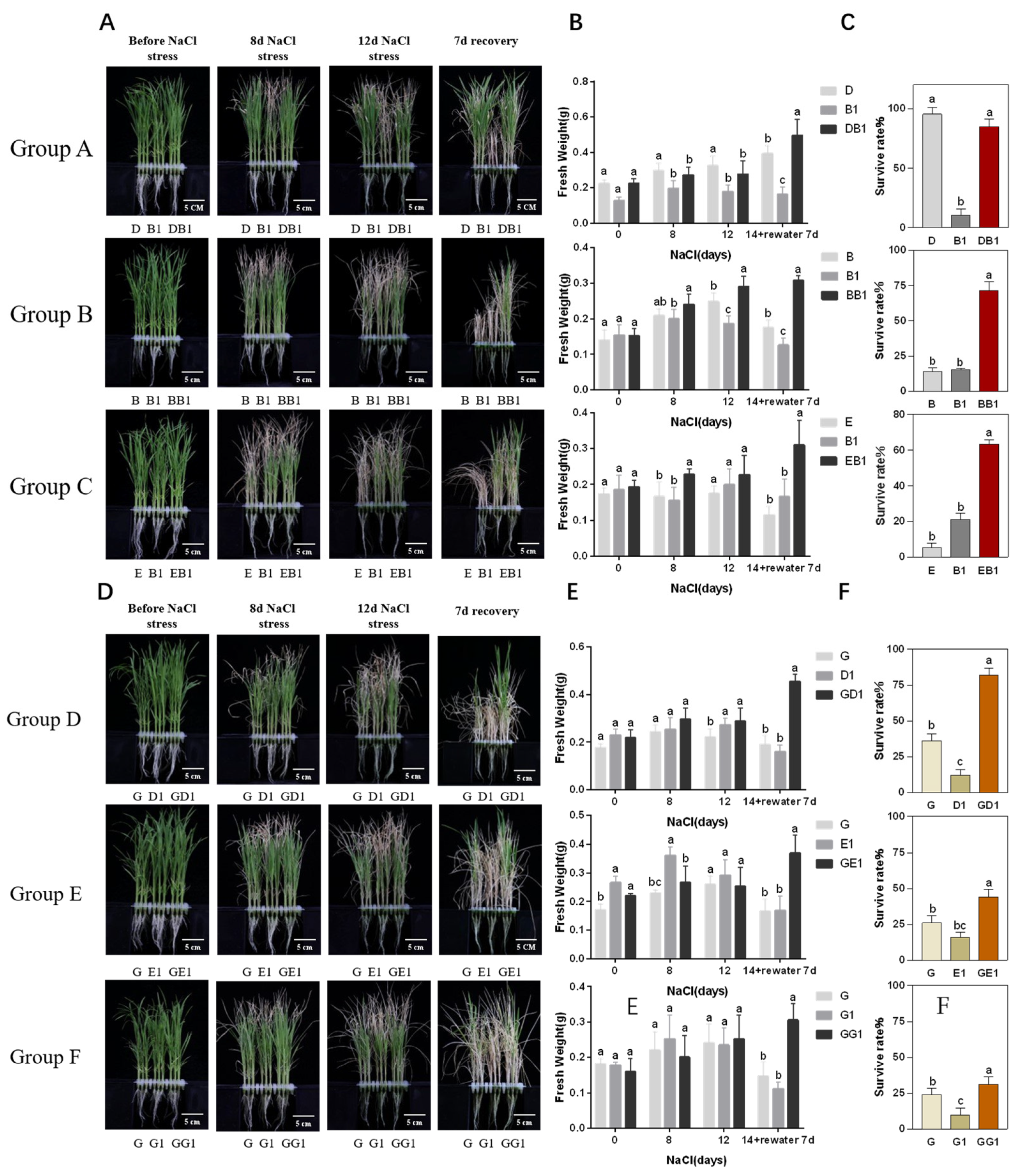
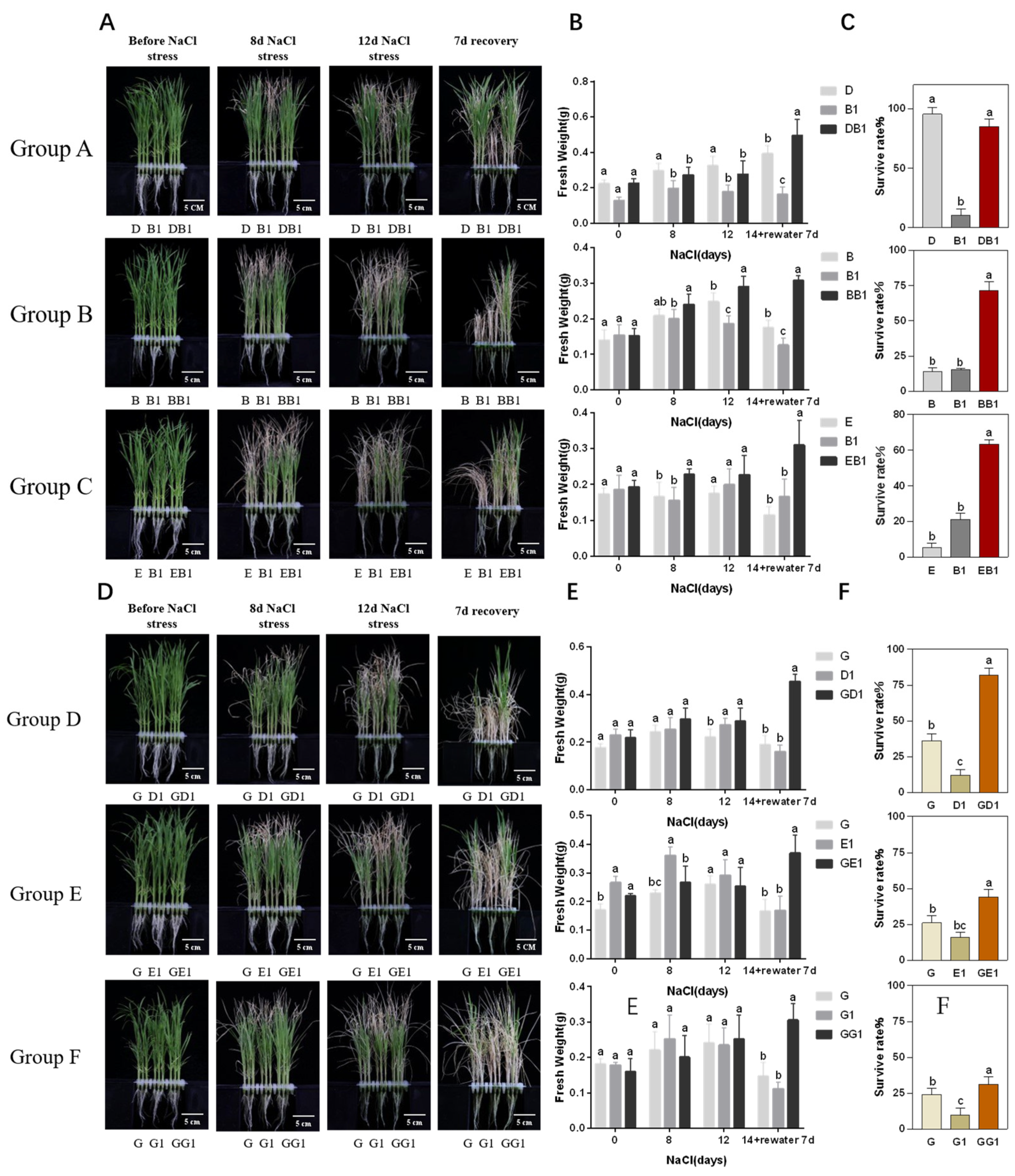
2.1. Rice Hybrids Exhibited Better Salt Tolerance Than Their Parents
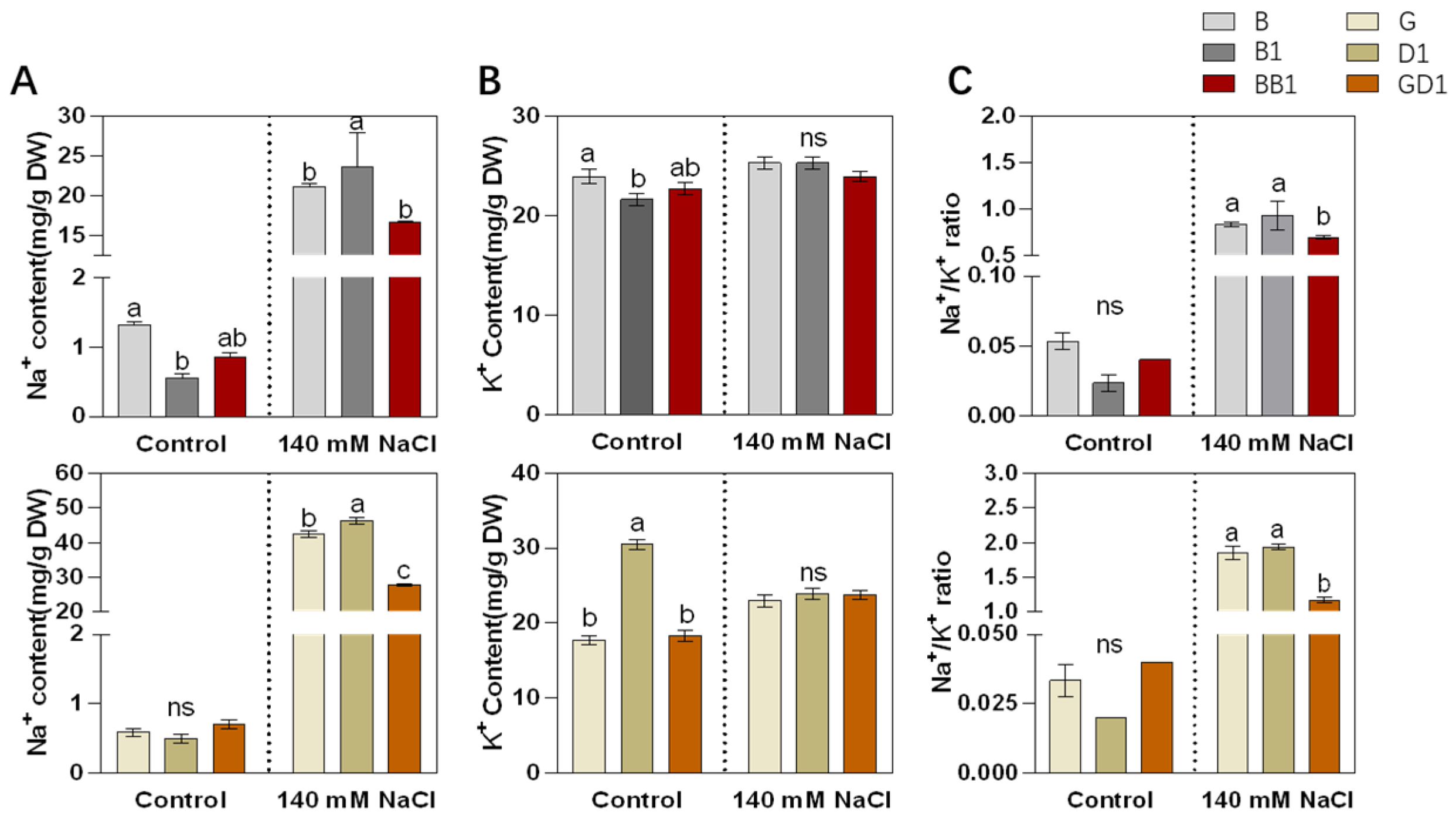
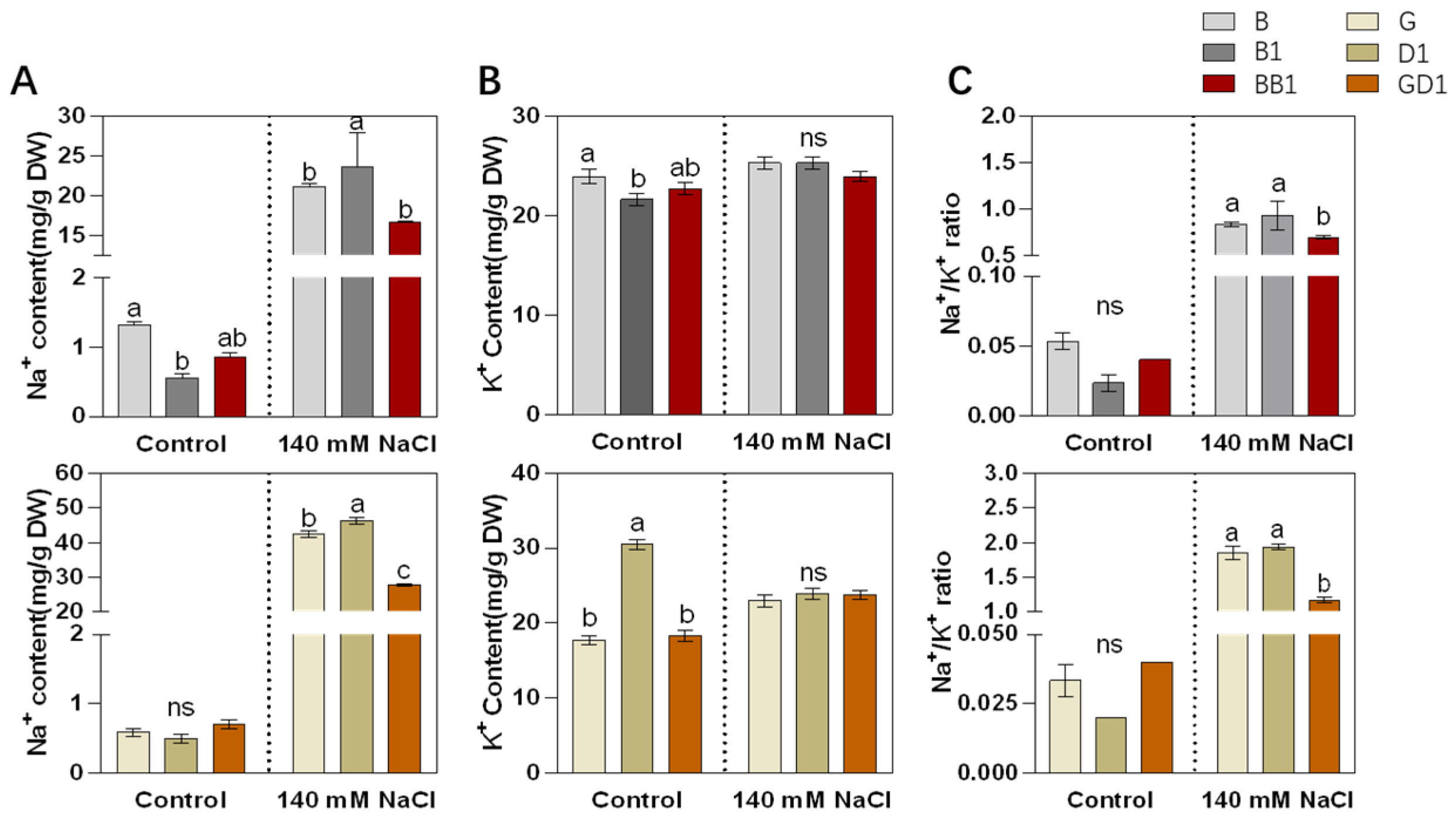
2.2. Rice Hybrids Exhibited Better Ionic Homeostasis with a Lower Na+ Content and Na+/K+ Ratio
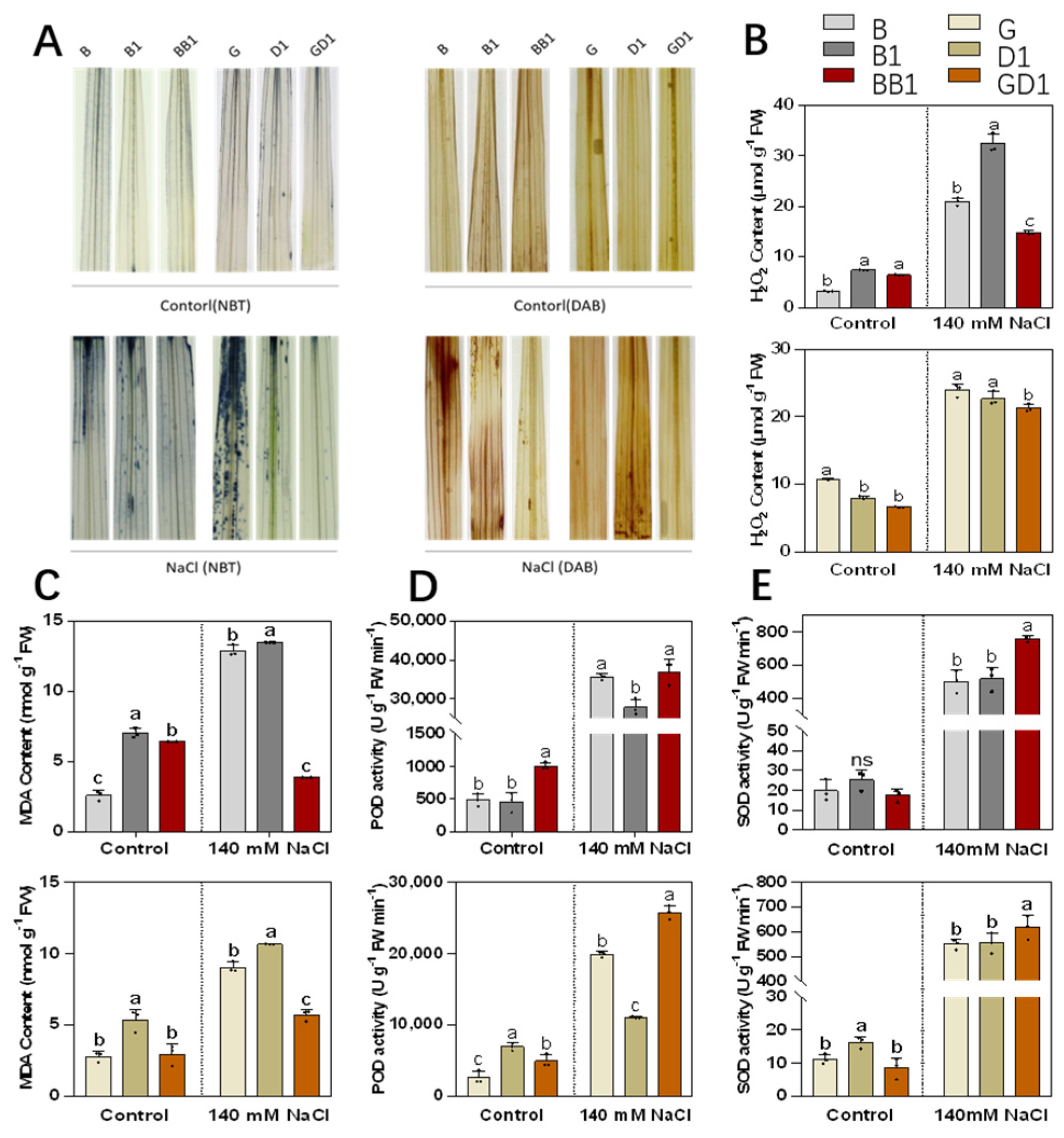
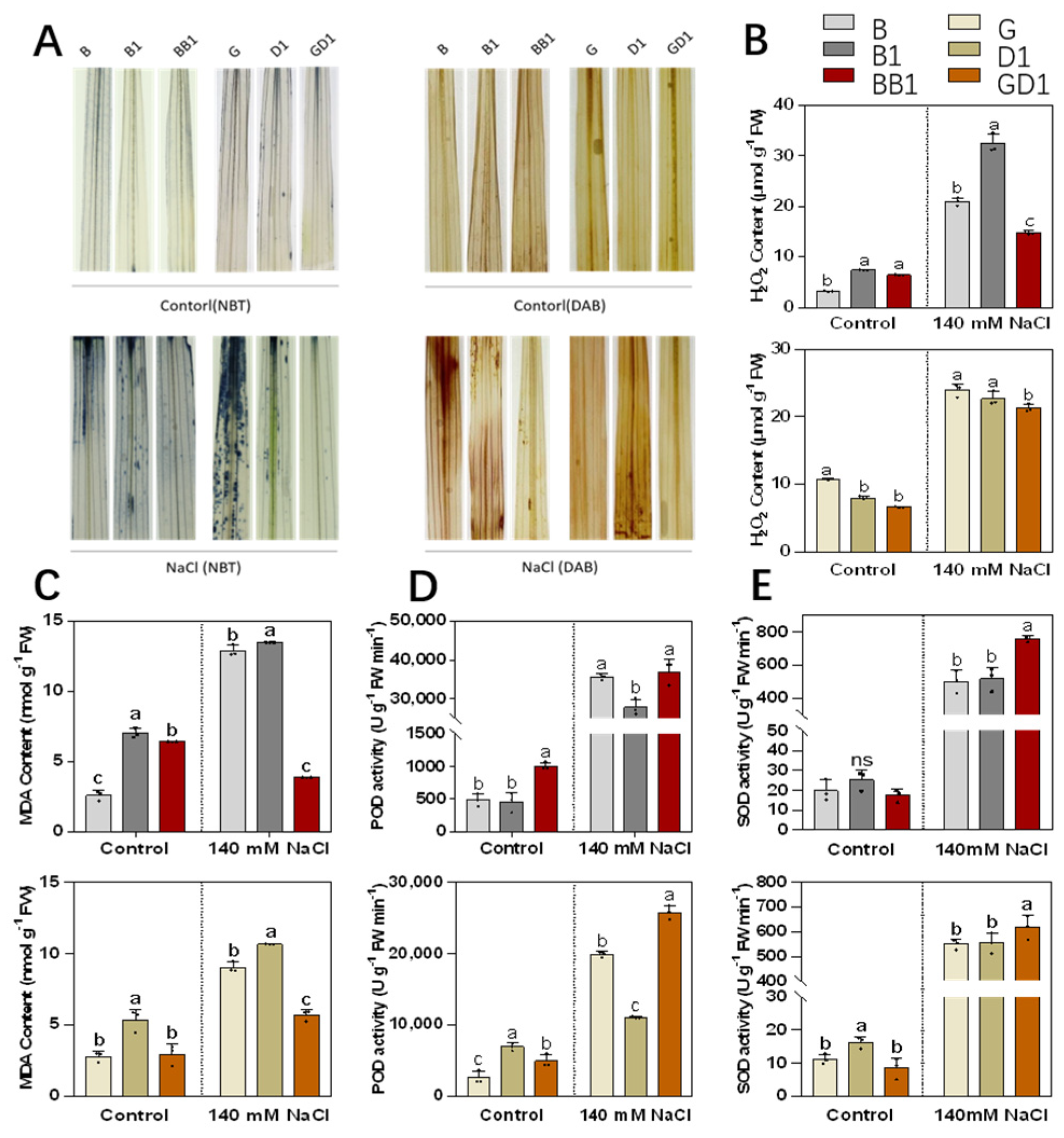
2.3. Rice Hybrids Exhibited Less ROS Accumulation under Salt Stress
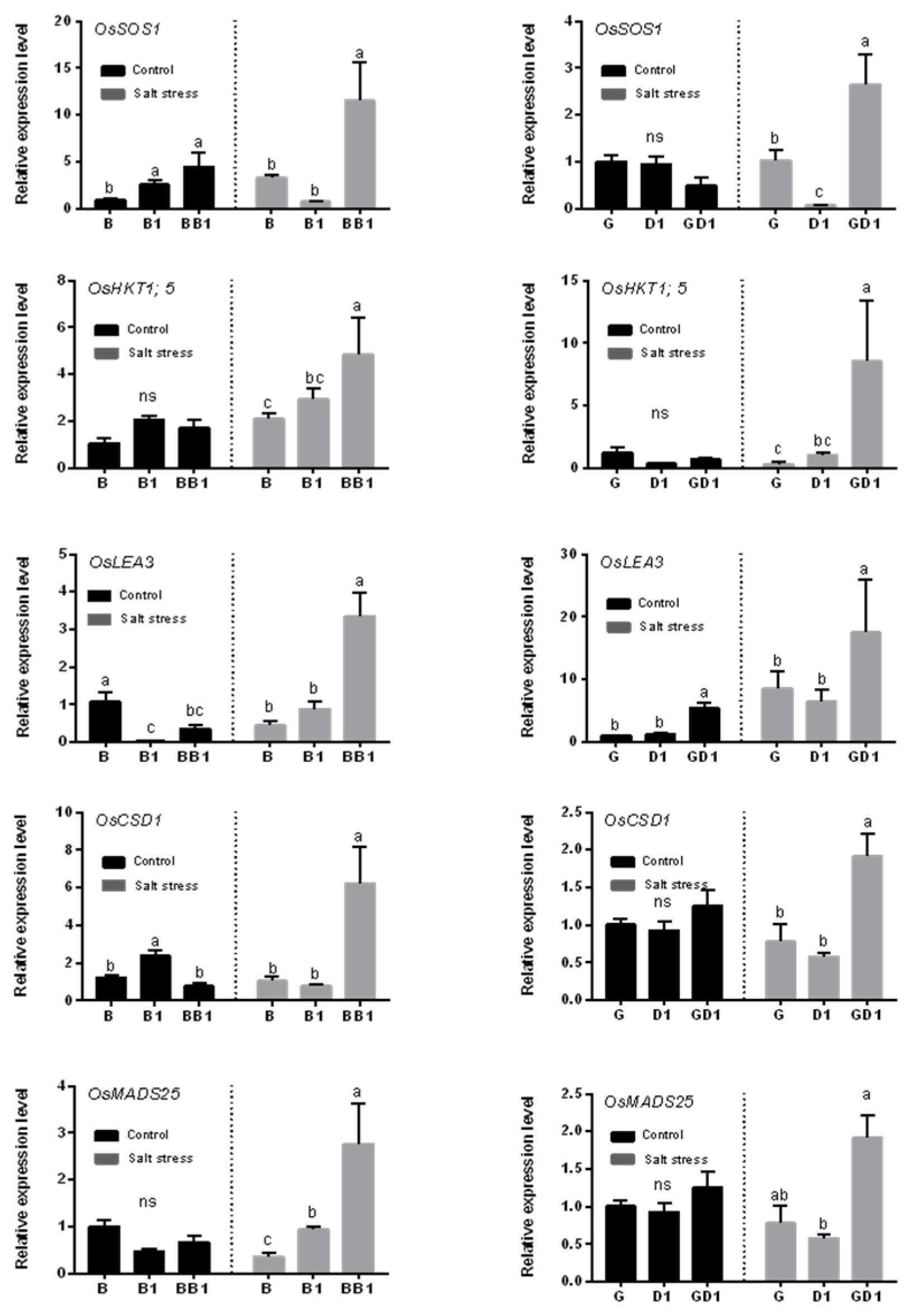
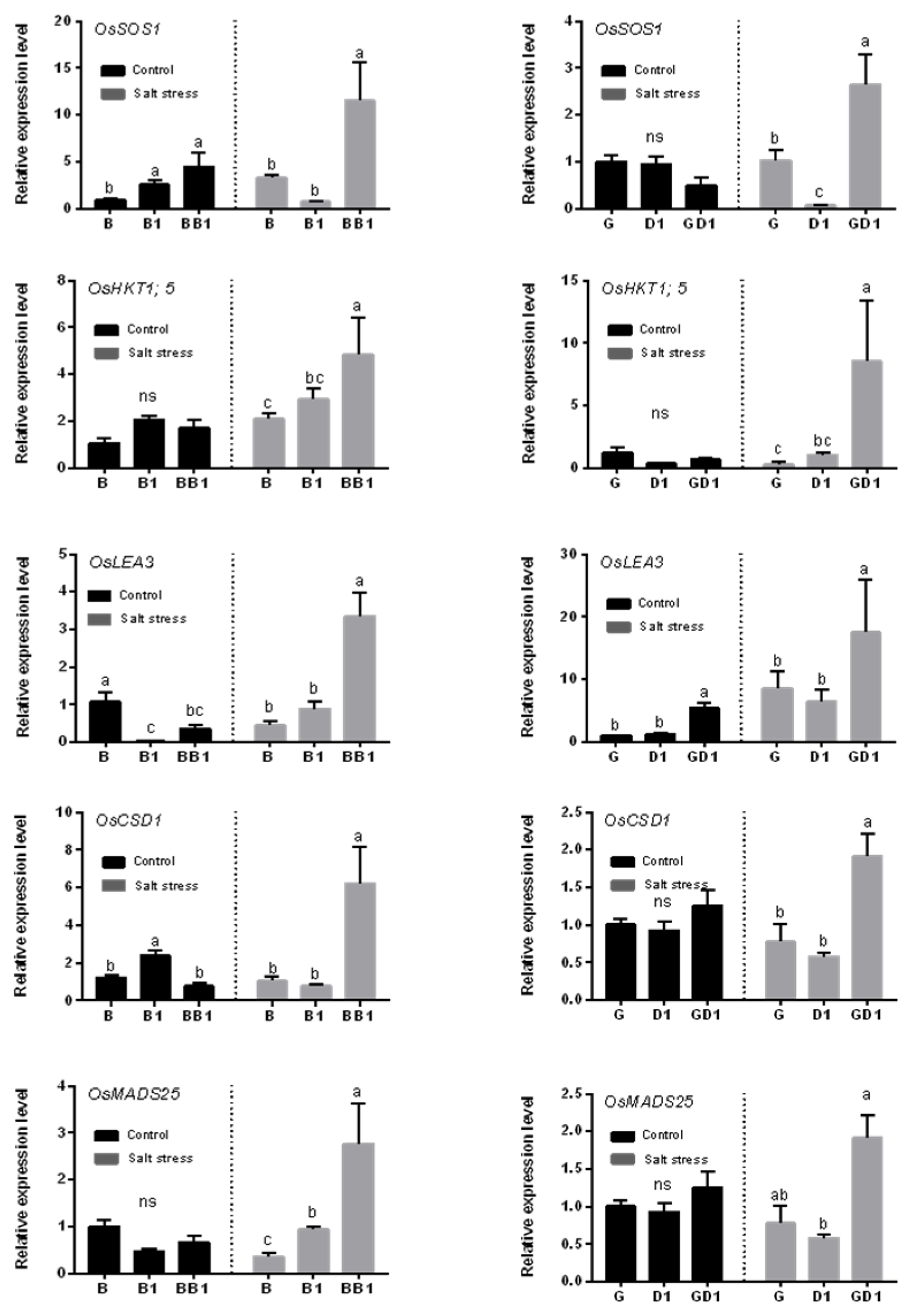
2.4. Rice Hybrids Exhibited a Higher Expression of Salt-Stress-Responsive Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Phenotype Analysis and Evaluation of Salinity Tolerance
4.3. Measurement of Na+ and K+ Contents
4.4. Measurement of Antioxidant Enzyme Activity
4.5. Measurement of the Total H2O2 and MDA Contents
4.6. Histochemical Staining
4.7. Total RNA Extraction and Gene Expression Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Cham, Switzerland, 2018; pp. 43–53. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.B.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric Oxide-Induced Salt Stress Tolerance in Plants: ROS Metabolism, Signaling, and Molecular Interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Ismail, A.M.; Heuer, S.; Thomson, M.J.; Wissuwa, M. Genetic and Genomic Approaches to Develop Rice Germplasm for Problem Soils. Plant Mol. Biol. 2007, 65, 547–570. [Google Scholar] [CrossRef]
- Citao, L.; Bigang, M.; Dingyang, Y.; Chengcai, C.; Meijuan, D. Salt Tolerance in Rice: Physiological Responses and Molecular Mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Reddy, I.N.B.L.; Kim, B.-K.; Yoon, I.-S.; Kim, K.-H.; Kwon, T.-R. Salt Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Chen, T.; Shabala, S.; Niu, Y.; Chen, Z.-H.; Shabala, L.; Meinke, H.; Venkataraman, G.; Pareek, A.; Xu, J.; Zhou, M. Molecular Mechanisms of Salinity Tolerance in Rice. Crop J. 2021, 9, 506–520. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the Molecular Mechanisms Mediating Plant Salt-Stress Responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Britto, D.T. Sodium Transport in Plants: A Critical Review. New Phytol. 2011, 189, 54–81. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Xia, Y.; Xie, Q.; Cao, Y.; Wang, Z.; Hao, G.; Song, J.; Zhou, Y.; Jiang, X. The Protein Kinase Complex CBL10-CIPK8-SOS1 Functions in Arabidopsis to Regulate Salt Tolerance. J. Exp. Bot. 2020, 71, 1801–1814. [Google Scholar] [CrossRef] [PubMed]
- Dvořák, P.; Krasylenko, Y.; Zeiner, A.; Šamaj, J.; Takáč, T. Signaling Toward Reactive Oxygen Species-Scavenging Enzymes in Plants. Front. Plant Sci. 2020, 11, 618835. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, W.; He, J.; Zhang, L.; Wei, Y.; Yang, M. Nitric Oxide Alleviates Salt Stress in Seed Germination and Early Seedling Growth of Pakchoi (Brassica chinensis L.) by Enhancing Physiological and Biochemical Parameters. Ecotoxicol. Environ. Saf. 2020, 187, 109785. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Jiang, W.; Tian, L.; Yong, F.; Tan, L.; Zhu, Z.; Sun, C.; Liu, F. Polyamine Oxidase 3 Is Involved in Salt Tolerance at the Germination Stage in Rice. J. Genet. Genom. 2022, 49, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Da Silva, J.A.T.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense Is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. [Google Scholar] [CrossRef]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt Tolerance Conferred by Overexpression of a Vacuolar Na+/H+ Antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A Rice Quantitative Trait Locus for Salt Tolerance Encodes a Sodium Transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamaji, N.; Costa, A.; Okuma, E.; Kobayashi, N.I.; Kashiwagi, T.; Katsuhara, M.; Wang, C.; Tanoi, K.; Murata, Y.; et al. OsHKT1;4-Mediated Na (+) Transport in Stems Contributes to Na (+) Exclusion from Leaf Blades of Rice at the Reproductive Growth Stage upon Salt Stress. BMC Plant Biol. 2016, 16, 22. [Google Scholar] [CrossRef]
- Song, T.; Shi, Y.; Shen, L.; Cao, C.; Shen, Y.; Jing, W.; Tian, Q.; Lin, F.; Li, W.; Zhang, W. An Endoplasmic Reticulum-Localized Cytochrome b 5 Regulates High-Affinity K+ Transport in Response to Salt Stress in Rice. Proc. Natl. Acad. Sci. USA 2021, 118, e2114347118. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Liang, X.; Lu, M.; Lai, J.; Song, W.; Jiang, C. A Teosinte-Derived Allele of an HKT1 Family Sodium Transporter Improves Salt Tolerance in Maize. Plant Biotechnol. J. 2022, 21, 97–108. [Google Scholar] [CrossRef]
- Jadamba, C.; Kang, K.; Paek, N.C.; Lee, S.I.; Yoo, S.C. Overexpression of Rice Expansin7 (Osexpa7) Confers Enhanced Tolerance to Salt Stress in Rice. Int. J. Mol. Sci. 2020, 21, 454. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wei, P.; Liu, Z.; Yu, B.; Shi, H. Soybean Na+/H+ Antiporter GmsSOS1 Enhances Antioxidant Enzyme Activity and Reduces Na+ Accumulation in Arabidopsis and Yeast Cells under Salt Stress. Acta Physiol. Plant. 2017, 39, 1–11. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Yang, X.; Ju, Q.; Li, W.; Lü, S.; Tran, L.; Xu, J. The R2R3-MYB Transcription Factor AtMYB49 Modulates Salt Tolerance in Arabidopsis by Modulating the Cuticle Formation and Antioxidant Defence. Plant Cell Environ. 2020, 43, 1925–1943. [Google Scholar] [CrossRef]
- Guan, Q.; Liao, X.; He, M.; Li, X.; Wang, Z.; Ma, H.; Yu, S.; Liu, S. Tolerance Analysis of Chloroplast OsCu/Zn-SOD Overexpressing Rice under NaCl and NaHCO3 Stress. PLoS ONE 2017, 12, e0186052. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Chu, Y.; Chen, H.; Li, X.; Wu, Q.; Jin, L.; Wang, G.; Huang, J. Rice Transcription Factor OsMADS25 Modulates Root Growth and Confers Salinity Tolerance via the ABA-Mediated Regulatory Pathway and ROS Scavenging. PLoS Genet. 2018, 14, e1007662. [Google Scholar] [CrossRef] [PubMed]
- Groszmann, M.; Gonzalez-Bayon, R.; Lyons, R.L.; Dennis, E.S. Hormone-Regulated Defense and Stress Response Networks C-ontribute to Heterosis in Arabidopsis F1 Hybrids. Proc. Natl. Acad. Sci. USA 2015, 112, 6397–6406. [Google Scholar] [CrossRef]
- Fujimoto, R.; Uezono, K.; Ishikura, S.; Osabe, K.; Peacock, W.J.; Dennis, E.S. Recent Research on the Mechanism of Heterosis Is Important for Crop and Vegetable Breeding Systems. Breed. Sci. 2018, 68, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Labroo, M.R.; Studer, A.J.; Rutkoski, J.E. Heterosis and Hybrid Crop Breeding: A Multidisciplinary Review. Front. Genet. 2021, 12, 643761. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Li, Y.; Huang, R. Advances and Challenges in the Breeding of Salt-Tolerant Rice. Int. J. Mol. Sci. 2020, 21, 8385. [Google Scholar] [CrossRef] [PubMed]
- Cailin, W.; Yadong, Z.; Ling, Z.; Tao, C.; Qingyong, Z.; Shu, Y.; Lihui, Z.; Chunfang, Z.; Wenhua, L.; Mingfa, S.; et al. Research Status, Problems and Suggestions on Salt-Alkali Tolerant Rice. China Rice 2019, 25, 1–6. [Google Scholar] [CrossRef]
- Dongping, Y.; Jun, W.; Zhongxiao, H.; Bin, B.; Wen, Z.; Jianwu, L.; Qiyun, D. Physiological Mechanism and Breeding Strategy of Rice Saline-Alkaline Tolerance. Hybrid Rice 2019, 4, 1–7. [Google Scholar] [CrossRef]
- Singh, R.; Gregorio, G. CSR23: A New Salt-Tolerant Rice Variety for India. Int. Rice Res. Notes 2009, 31, 16–18. [Google Scholar] [CrossRef]
- Akbar, M.; Yabuno, T.; Nakao, S. Breeding for Saline-Resistant Varieties of Rice: I. Variability for Salt Tolerance among Some Rice Varietles. Breed. Sci. 1972, 22, 277–284. [Google Scholar] [CrossRef]
- Fernando, L.H. The Performance of Salt Resistant Paddy, Pokkali in Ceylon. Trop. Agric. 1949, 105, 124–126. [Google Scholar]
- Yadong, Z.; Zhen, Z.; Tao, C.; Yongqing, Z.; Shu, Y.; Lihui, Z.; Ling, Z.; Chunfang, Z.; Wenhua, L.; Kai, L.; et al. Breeding and Characteristics of the New Salt-Tolerant Rice Variety Nangengyan No.1 with Fine Quality. Barley Cereal Sci. 2023, 40, 67–70. [Google Scholar] [CrossRef]
- Shaolu, Z.; Dongying, Z.; Hongliang, Y.; Yafei, L.; Kai, L.; Bojie, W.; Jingwen, Z.; Jinying, D.; Guohong, Y.; Aimin, W.; et al. Breeding and Application of a New Salt-Tolerant Rice Cultivar Zhongkeyan No.4. Jiangsu Agric. Sci. 2022, 50, 303–308. [Google Scholar] [CrossRef]
- Song, S.; Wang, T.; Li, Y.; Hu, J.; Kan, R.; Qiu, M.; Deng, Y.; Liu, P.; Zhang, L.; Dong, H.; et al. A Novel Strategy for Creating a New System of Third-Generation Hybrid Rice Technology Using a Cytoplasmic Sterility Gene and a Genic Male-Sterile Gene. Plant Biotechnol. J. 2021, 19, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Ai, Z.; Tan, Y.; Hu, Y.; Guo, X.; Liu, X.; Sun, Z.; Yu, D.; Chen, J.; Tang, N.; et al. Novel Salinity-Tolerant Third-Generation Hybrid Rice Developed via CRISPR/Cas9-Mediated Gene Editing. Int. J. Mol. Sci. 2023, 24, 8025. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Joshi, M.; Joshi, V. Transcriptomic Analysis of Short-Term Salt Stress Response in Watermelon Seedlings. Int. J. Mol. Sci. 2020, 21, 6036. [Google Scholar] [CrossRef] [PubMed]
- Chunthaburee, S.; Dongsansuk, A.; Sanitchon, J.; Pattanagul, W.; Theerakullpisut, P. Physiological and Biochemical Parameters for Evaluation and Clustering of Rice Cultivars Differing in Salt Tolerance at Seedling Stage. Saudi J. Biol. Sci. 2016, 23, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Alkahtani, J.; Dwiningsih, Y. Analysis of Morphological, Physiological, and Biochemical Traits of Salt Stress Tolerance in Asian Rice Cultivars at Seedling and Early Vegetative Stages. Stresses 2023, 3, 717–735. [Google Scholar] [CrossRef]
- Singh, M.B.; Singh, D.; Rai, M. Assessment of Growth, Physiological and Biochemical Parameters and Activities of Antioxidative Enzymes in Salinity Tolerant and Sensitive Basmati Rice Varieties. J. Agron. Crop Sci. 2007, 193, 398–412. [Google Scholar] [CrossRef]
- Ying, T.; Dingyang, Y.; Zhixiang, G.; Taoli, L.; Xiabing, S.; Huiqun, W.; Yanning, T. Screening and Physiological Analysis of Salt-Tolerant Male Sterile Lines in Indica Rice. Hybrid Rice 2023, 38, 25–31. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Sugawara, M.; Okada, T.; Taira, K.; Nakayama, P.; Katsuhara, M.; Shinmyo, A.; Nakayama, H. Rice Sodium-Insensitive Potassium Transporter, OsHAK5, Confers Increased Salt Tolerance in Tobacco BY2 Cells. J. Biosci. Bioeng. 2011, 111, 346–356. [Google Scholar] [CrossRef]
- Olis, C.A.; Yong, M.-T.; Zhou, M.; Venkataraman, G.; Shabala, L.; Holford, P.; Shabala, S.; Chen, Z.-H. Evolutionary Significance of NHX Family and NHX1 in Salinity Stress Adaptation in the Genus Oryza. Int. J. Mol. Sci. 2022, 23, 2092. [Google Scholar] [CrossRef]
- Cuin, T.A.; Bose, J.; Stefano, G.; Jha, D.; Tester, M.; Mancuso, S.; Shabala, S. Assessing the Role of Root Plasma Membrane and Tonoplast Na+/H+ Exchangers in Salinity Tolerance in Wheat: In Planta Quantification Methods. Plant Cell Environ. 2011, 34, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, D.; Wu, T.; Yuanzhu, Y.; Cong, L.; Lu, Y.; Dongying, T.; Xiaoying, Z.; Yonghua, Z.; Jianzhong, L.; et al. LRRK1, a receptor-like cytoplasmic kinase, regulates leaf rolling through modulating bulliform cell development in rice. Mol. Breed. 2018, 38, 48. [Google Scholar] [CrossRef]
- Stupar, R.M.; Springer, N.M. Cis-Transcriptional Variation in Maize Inbred Lines B73 and Mo17 Leads to Additive Expression Patterns in the F1 Hybrid. Genetics 2006, 173, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Ni, Z.; Zhang, Y.; Chen, Y.; Ding, Y.; Han, Z.; Liu, Z.; Sun, Q. Identification of Differentially Expressed Genes in Leaf and Root between Wheat Hybrid and Its Parental Inbreds Using PCR-Based cDNA Subtraction. Plant Mol. Biol. 2005, 58, 367–384. [Google Scholar] [CrossRef]
- Martínez-Atienza, J.; Jiang, X.; Garciadeblas, B.; Mendoza, I.; Zhu, J.-K.; Pardo, J.M.; Quintero, F.J. Conservation of the Salt Overly Sensitive Pathway in Rice. Plant Physiol. 2007, 143, 1001–1012. [Google Scholar] [CrossRef]
- El Mahi, H.; Pérez-Hormaeche, J.; De Luca, A.; Villalta, I.; Espartero, J.; Gámez-Arjona, F.; Fernández, J.L.; Bundó, M.; Mendoza, I.; Mieulet, D.; et al. A Critical Role of Sodium Flux via the Plasma Membrane Na+/H+ Exchanger SOS1 in the Salt Tolerance of Rice. Plant Physiol. 2019, 180, 1046–1065. [Google Scholar] [CrossRef]
- Shohan, M.U.S.; Sinha, S.; Nabila, F.H.; Dastidar, S.G.; Seraj, Z.I. HKT1; 5 Transporter Gene Expression and Association of Amino Acid Substitutions with Salt Tolerance across Rice Genotypes. Front. Plant Sci. 2019, 10, 01420. [Google Scholar] [CrossRef]
- Moons, A.; Bauw, G.; Prinsen, E.; Van Montagu, M.; Van der Straeten, D. Molecular and Physiological Responses to Abscisic Acid and Salts in Roots of Salt-Sensitive and Salt-Tolerant Indica Rice Varieties. Plant Physiol. 1995, 107, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S. Laboratory Manual for Physiological Studies of Rice. Agric. Food Sci. 1971, 23, 61–66. [Google Scholar]
- Roshdy, A.E.-D.; Alebidi, A.; Almutairi, K.; Al-Obeed, R.; Elsabagh, A. The Effect of Salicylic Acid on the Performances of Salt Stressed Strawberry Plants, Enzymes Activity, and Salt Tolerance Index. Agronomy 2021, 11, 775. [Google Scholar] [CrossRef]
- Spitz, D.R.; Oberley, L.W. An Assay for Superoxide Dismutase Activity in Mammalian Tissue Homogenates. Anal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Reuveni, R.; Shimoni, M.; Karchi, Z.; Kuc, J. Peroxidase Activity as a Biochemical Marker for Resistance of Muskmelon (Cucumis melo) to Pseudoperonospora Cubensis. Phytopathology 1992, 82, 749–753. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Cultivars | Letter |
|---|---|---|
| A | Guang-Xiang-24 (♀) | D |
| Hua-Zhan (♂) | B1 | |
| Xiang-Liang-You-Hua-Zhan (F1) | DB1 | |
| B | Guang-Ba-A (♀) | B |
| Hua-Zhan (♂) | B1 | |
| Guang-Ba-You-Hua-Zhan (F1) | BB1 | |
| C | Jing-4155S (♀) | E |
| Hua-Zhan (♂) | B1 | |
| Jing-Liang-You-Hua-Zhan (F1) | EB1 | |
| D | Y58S (♀) | G |
| R900 (♂) | D1 | |
| Y-Liang-You-900 (F1) | GD1 | |
| E | Y58S (♀) | G |
| R9311 (♂) | E1 | |
| Y-Liang-You-1 (F1) | GE1 | |
| F | Y58S (♀) | G |
| Yuan-Hui-2 (♂) | G1 | |
| Y-Liang-You-2 (F1) | GG1 |
| Groups | Cultivars | Dry Weight | Salt Tolerant Index (%) | Salt Tolerance Level | Overparent Heterosis (%) |
|---|---|---|---|---|---|
| Group A | CK-D | 0.07 ± 0.01 | 79 | 3 | −18 |
| Salt-D | 0.06 ± 0.01 | ||||
| CK-B1 | 0.08 ± 0.02 | 52 | 5 | ||
| Salt-B1 | 0.04 ± 0.01 | ||||
| CK-DB1 | 0.11 ± 0.01 | 64 | 3 | ||
| Salt-DB1 | 0.07 ± 0.01 | ||||
| Group B | CK-B | 0.07 ± 0.02 | 68 | 3 | 21 |
| Salt-B | 0.05 ± 0.01 | ||||
| CK-B1 | 0.08 ± 0.01 | 59 | 5 | ||
| Salt-B1 | 0.04 ± 0.01 | ||||
| CK-BB1 | 0.02 ± 0.03 | 83 | 1 | ||
| Salt-BB1 | 0.07 ± 0.01 | ||||
| Group C | CK-E | 0.13 ± 0.02 | 40 | 7 | 6 |
| Salt-E | 0.05 ± 0.01 | ||||
| CK-B1 | 0.07 ± 0.01 | 48 | 5 | ||
| Salt-B1 | 0.03 ± 0.00 | ||||
| CK-EB1 | 0.09 ± 0.01 | 51 | 5 | ||
| Salt-EB1 | 0.04 ± 0.01 | ||||
| Group D | CK-G | 0.10 ± 0.01 | 59 | 5 | 26 |
| Salt-G | 0.06 ± 0.01 | ||||
| CK-D1 | 0.10 ± 0.01 | 62 | 3 | ||
| Salt-D1 | 0.06 ± 0.01 | ||||
| CK-GD1 | 0.07 ± 0.01 | 78 | 3 | ||
| Salt-GD1 | 0.05 ± 0.01 | ||||
| Group E | CK-G | 0.12 ± 0.01 | 57 | 5 | 15 |
| Salt-G | 0.06 ± 0.01 | ||||
| CK-E1 | 0.11 ± 0.03 | 34 | 7 | ||
| Salt-E1 | 0.04 ± 0.01 | ||||
| CK-GE1 | 0.05 ± 0.01 | 66 | 3 | ||
| Salt-GE1 | 0.04 ± 0.01 | ||||
| Group F | CK-G | 0.10 ± 0.03 | 48 | 5 | 11 |
| Salt-G | 0.05 ± 0.01 | ||||
| CK-G1 | 0.08 ± 0.01 | 55 | 5 | ||
| Salt-G1 | 0.04 ± 0.01 | ||||
| CK-GG1 | 0.07 ± 0.01 | 62 | 3 | ||
| Salt-GG1 | 0.04 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Hu, Y.; Li, R.; Tang, L.; Mo, L.; Pan, Y.; Mao, B.; Shao, Y.; Zhao, B.; Lei, D. Research on Physiological Characteristics and Differential Gene Expression of Rice Hybrids and Their Parents under Salt Stress at Seedling Stage. Plants 2024, 13, 744. https://doi.org/10.3390/plants13050744
Zhang D, Hu Y, Li R, Tang L, Mo L, Pan Y, Mao B, Shao Y, Zhao B, Lei D. Research on Physiological Characteristics and Differential Gene Expression of Rice Hybrids and Their Parents under Salt Stress at Seedling Stage. Plants. 2024; 13(5):744. https://doi.org/10.3390/plants13050744
Chicago/Turabian StyleZhang, Dan, Yuanyi Hu, Ruopeng Li, Li Tang, Lin Mo, Yinlin Pan, Bigang Mao, Ye Shao, Bingran Zhao, and Dongyang Lei. 2024. "Research on Physiological Characteristics and Differential Gene Expression of Rice Hybrids and Their Parents under Salt Stress at Seedling Stage" Plants 13, no. 5: 744. https://doi.org/10.3390/plants13050744
APA StyleZhang, D., Hu, Y., Li, R., Tang, L., Mo, L., Pan, Y., Mao, B., Shao, Y., Zhao, B., & Lei, D. (2024). Research on Physiological Characteristics and Differential Gene Expression of Rice Hybrids and Their Parents under Salt Stress at Seedling Stage. Plants, 13(5), 744. https://doi.org/10.3390/plants13050744