

Effects of Rainfall Exclusion Treatment on Photosynthetic Characteristics of Black Locust in the Sub-Humid Region of the Loess Plateau, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Rainfall Exclusion Treatment
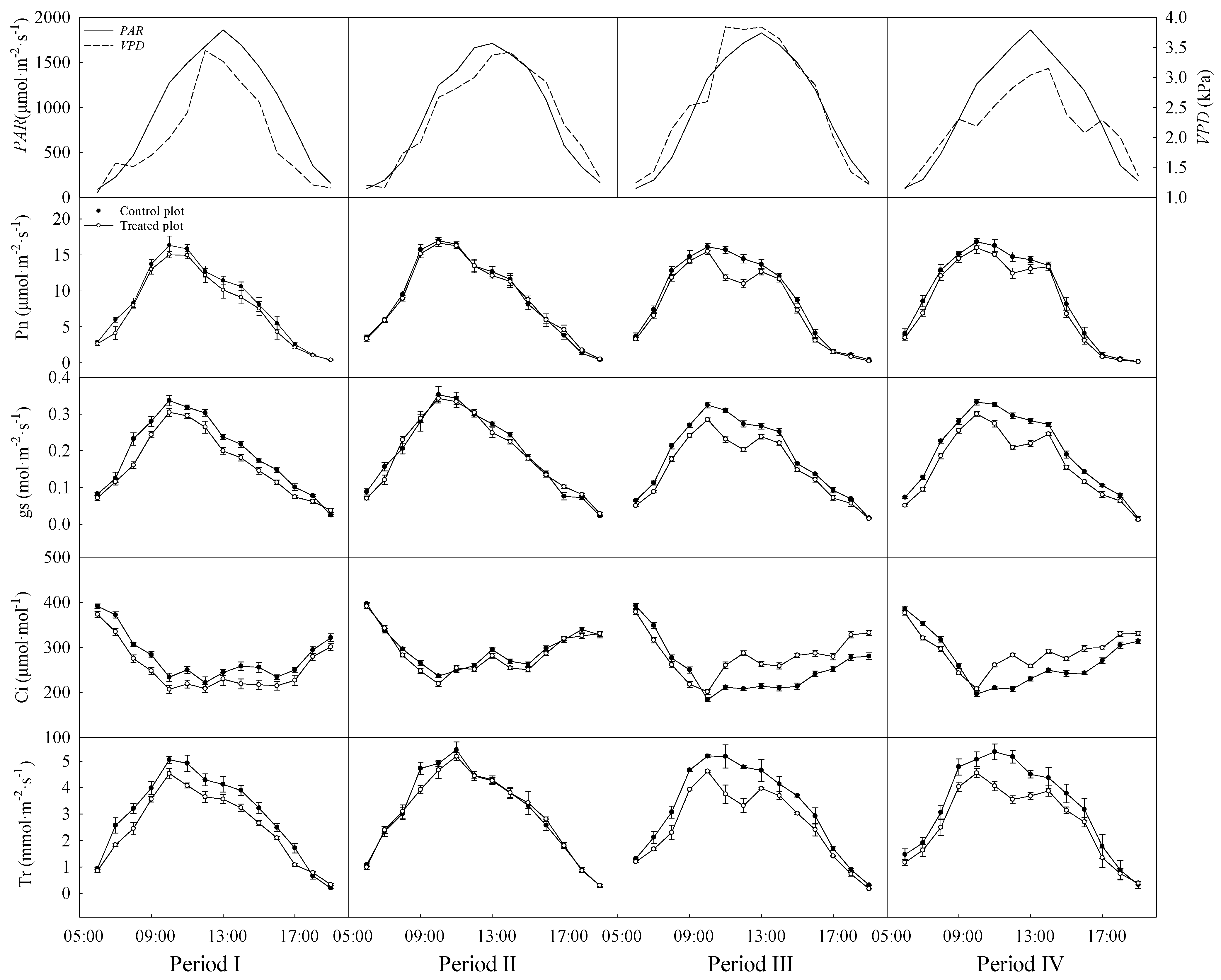
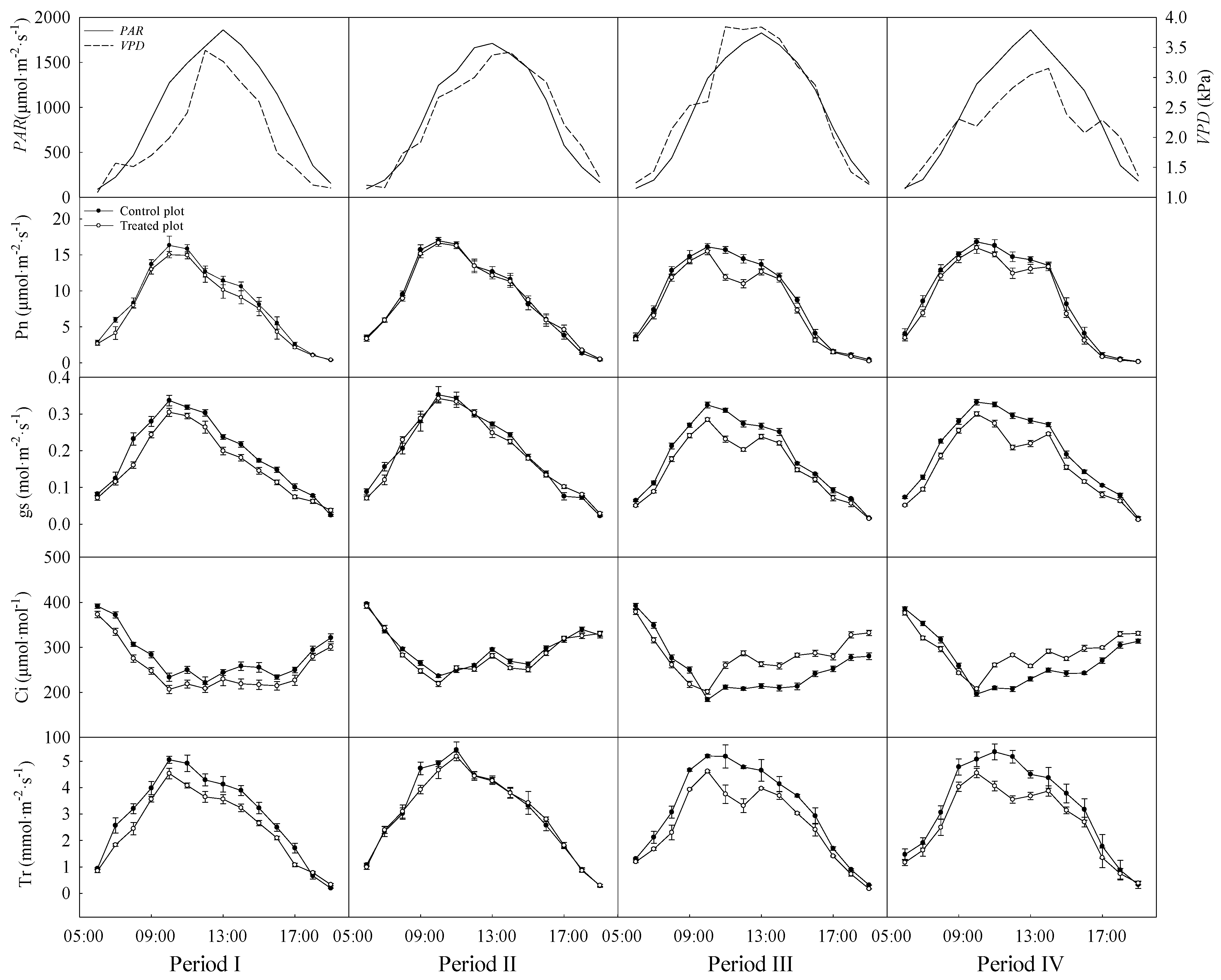
2.2. Diurnal Course Measurement of Canopy Leaf Gas Exchange
2.3. Light Response Curve
2.4. CO2 Response Curve
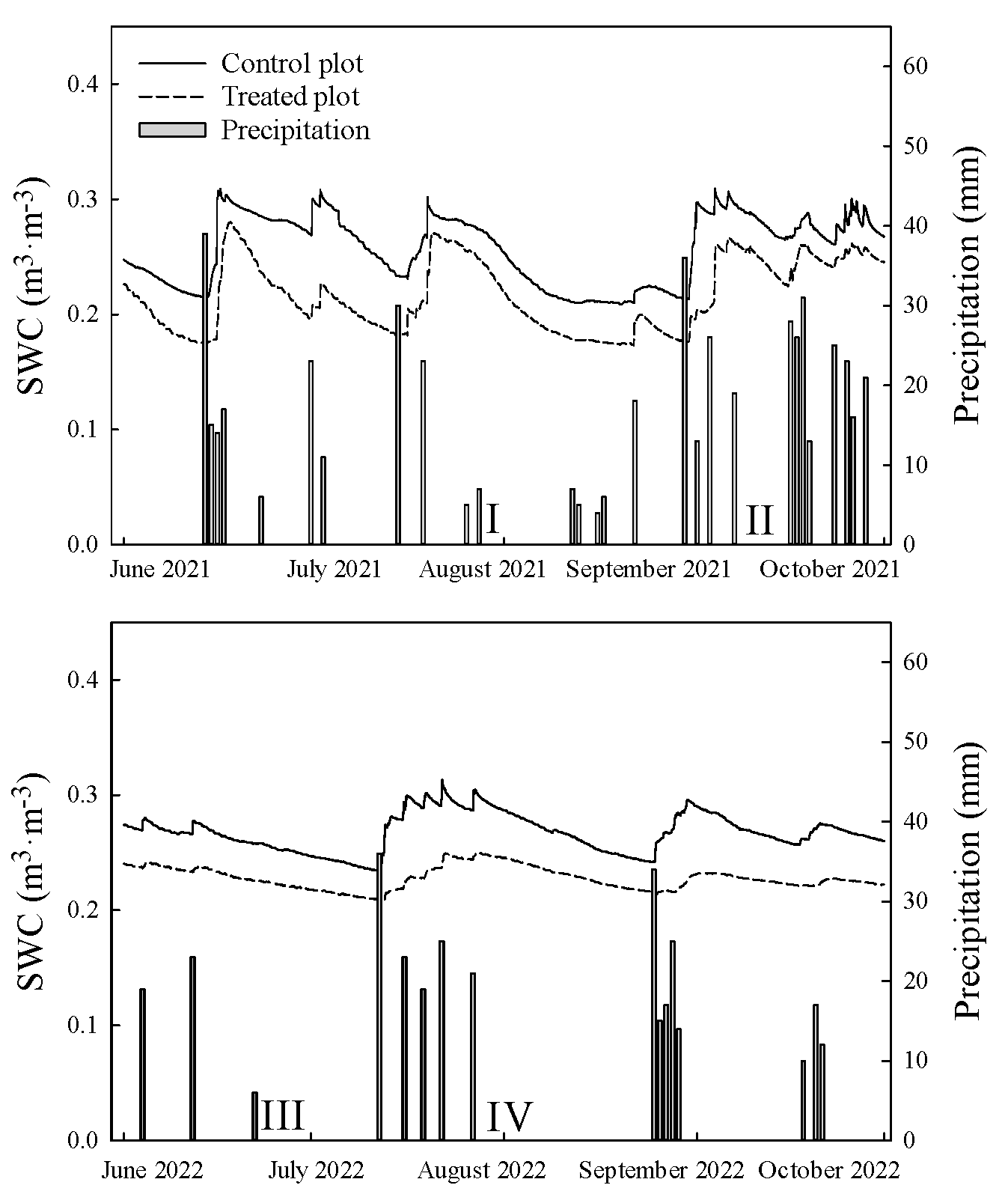
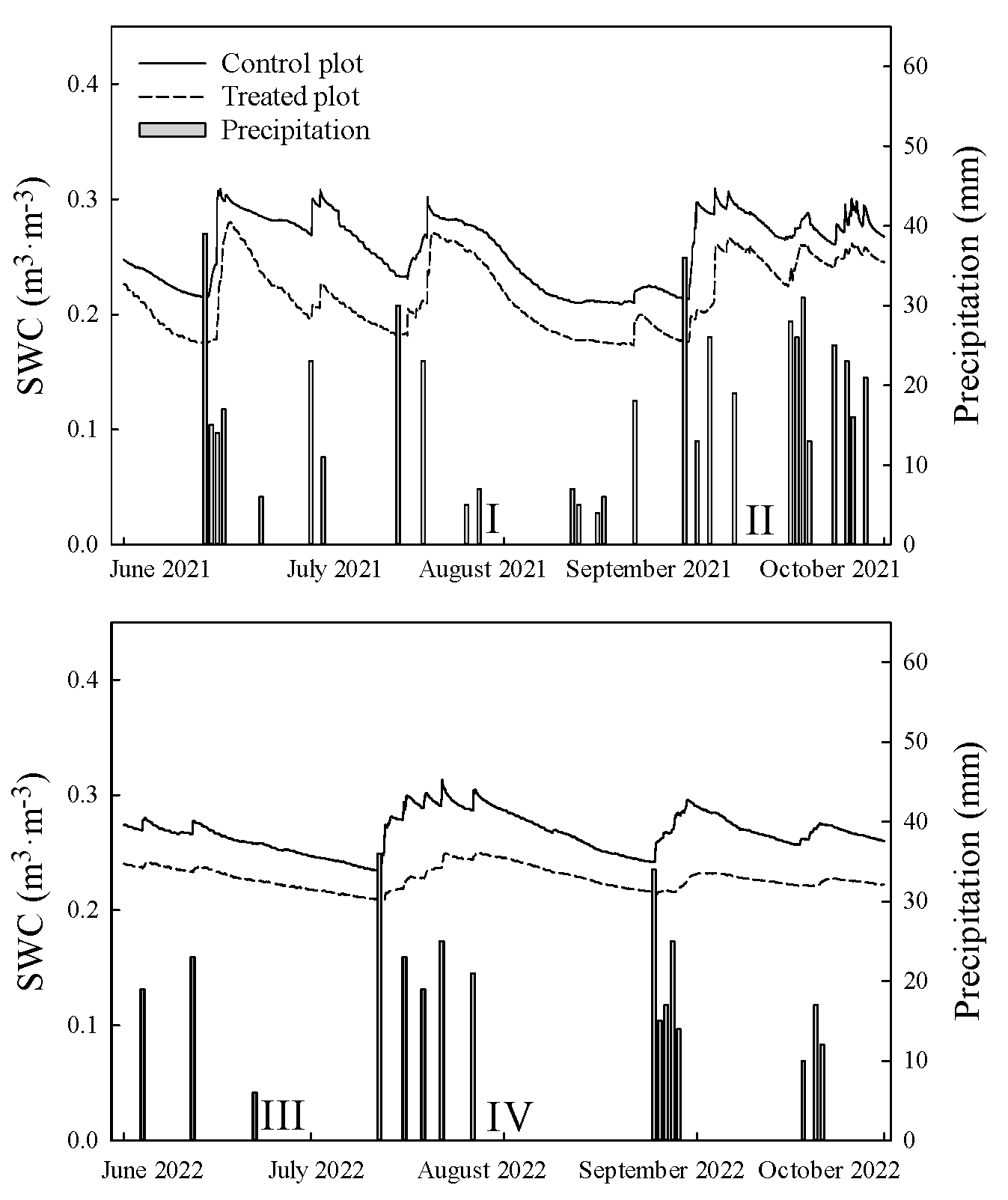
2.5. Measurement of Precipitation and Soil Water Content
2.6. Statistical Analysis
3. Results
3.1. Precipitation and Soil Water Content during the Study Periods
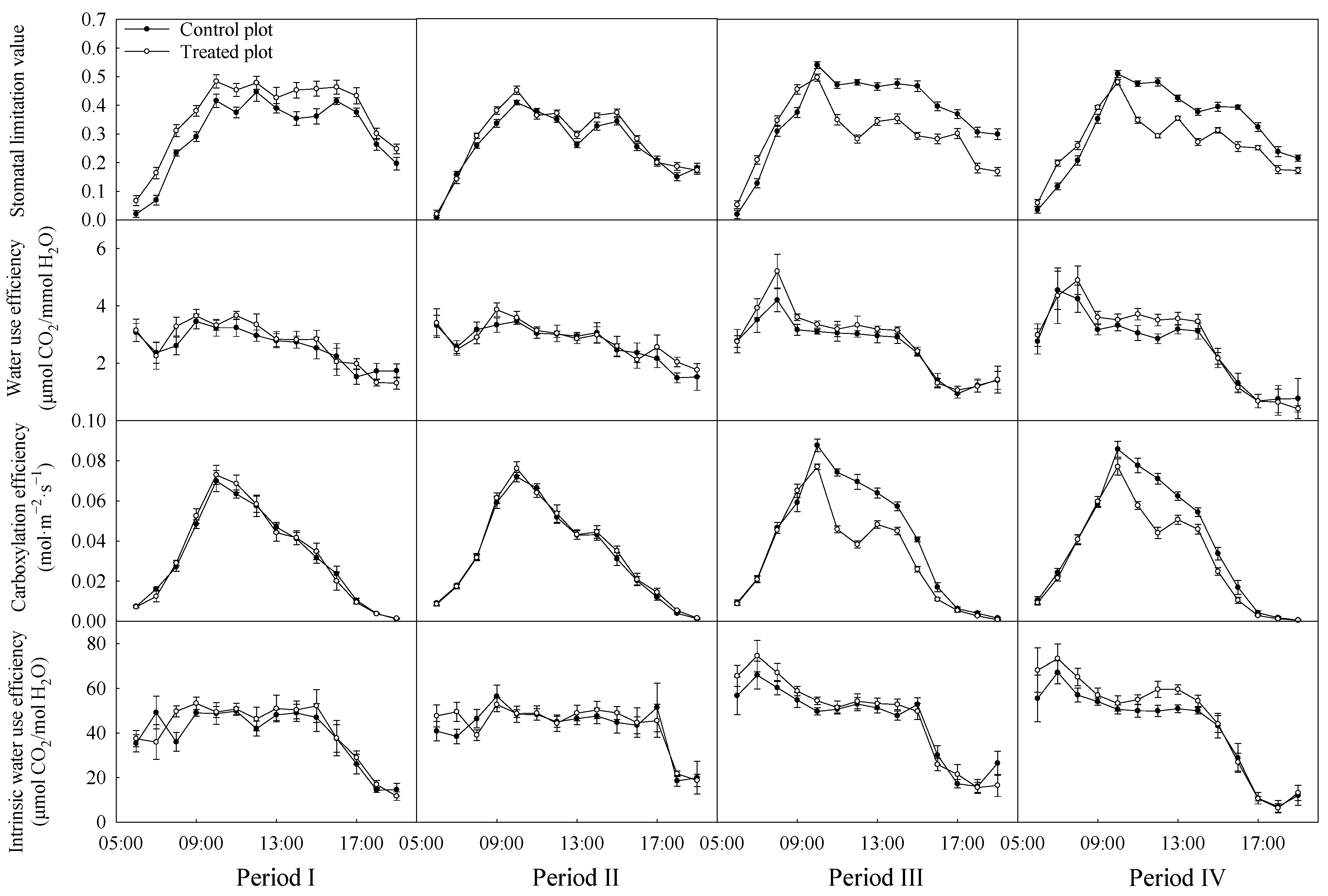
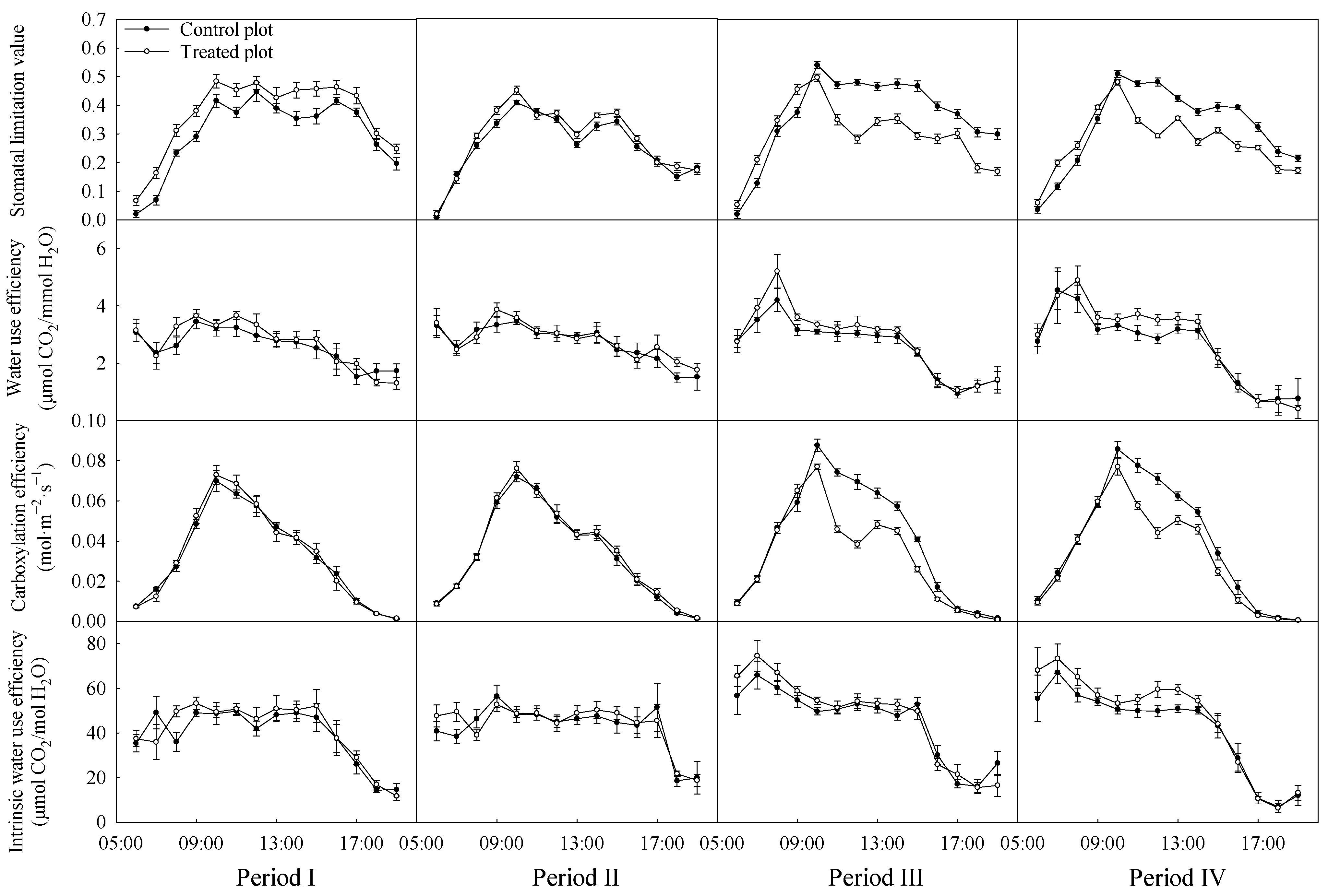
3.2. Variations in the Diurnal Courses of Photosynthetic Parameters
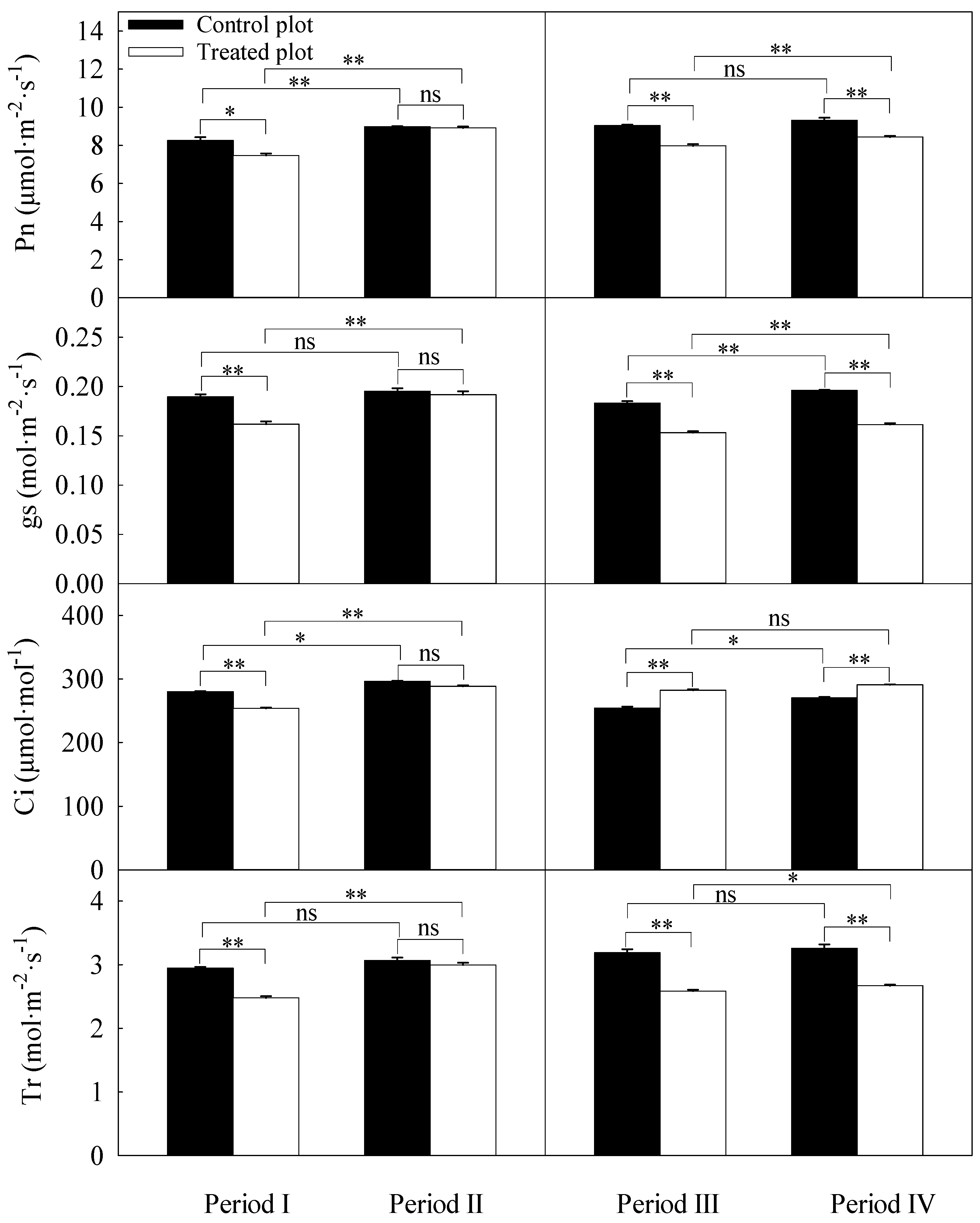
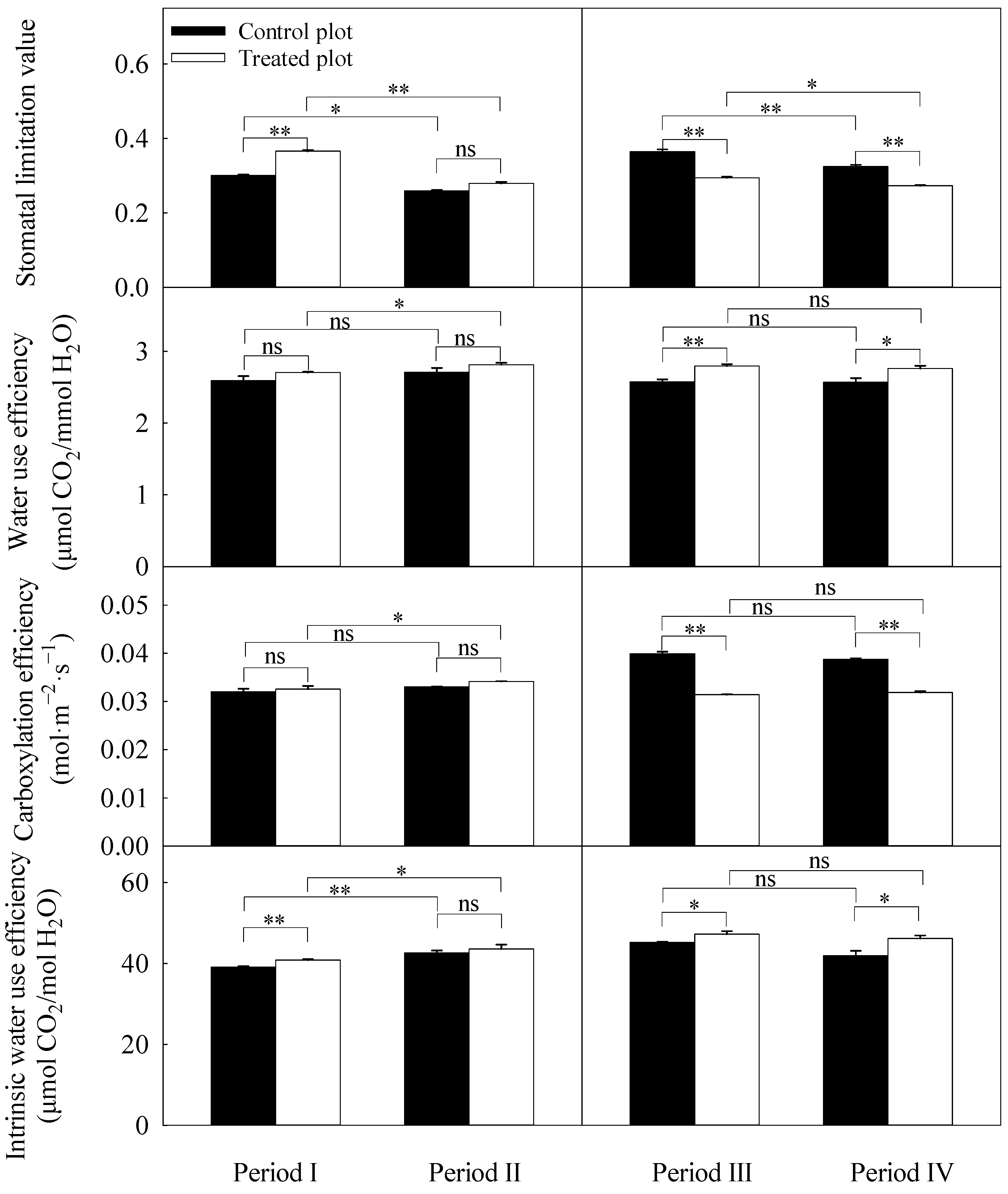
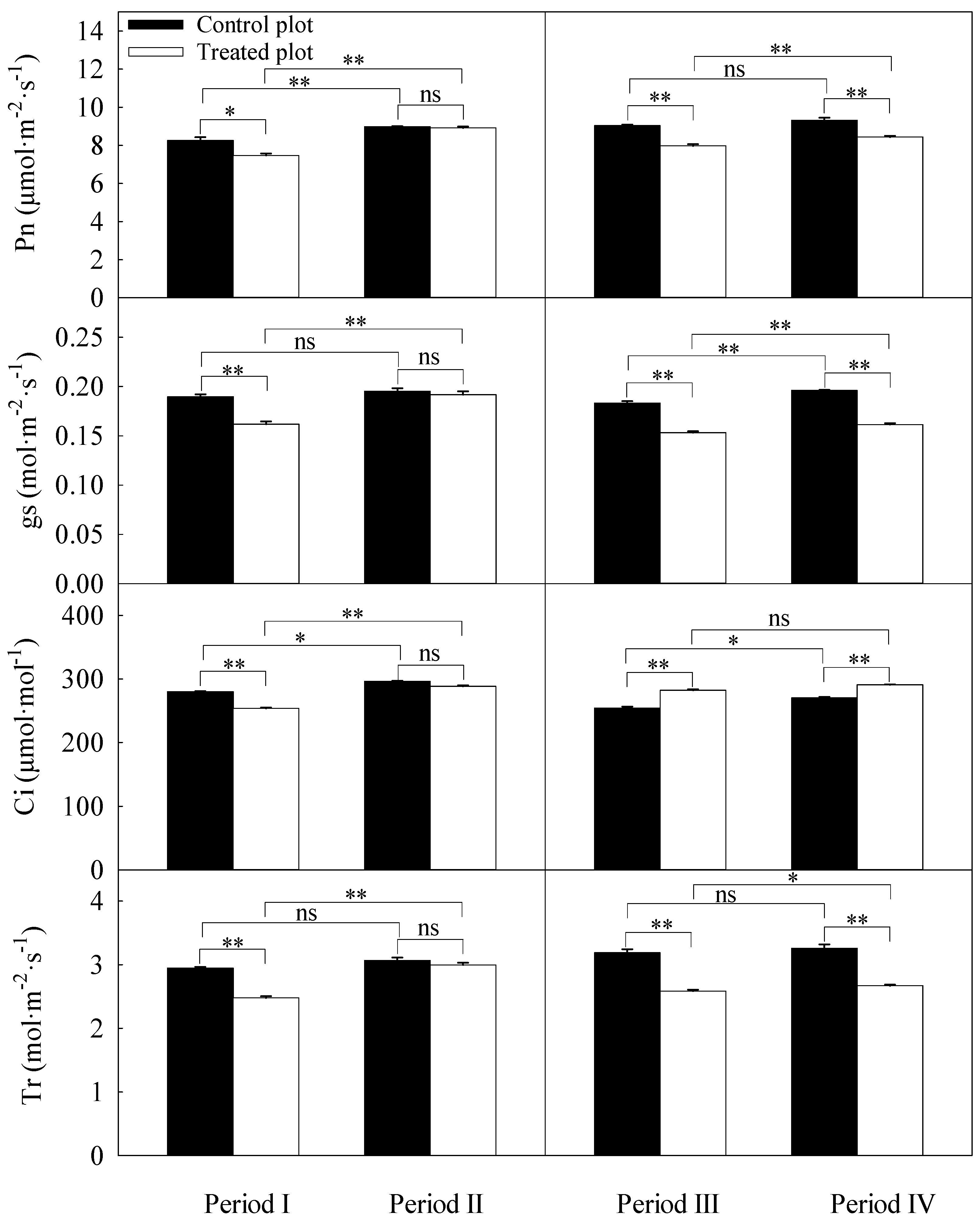
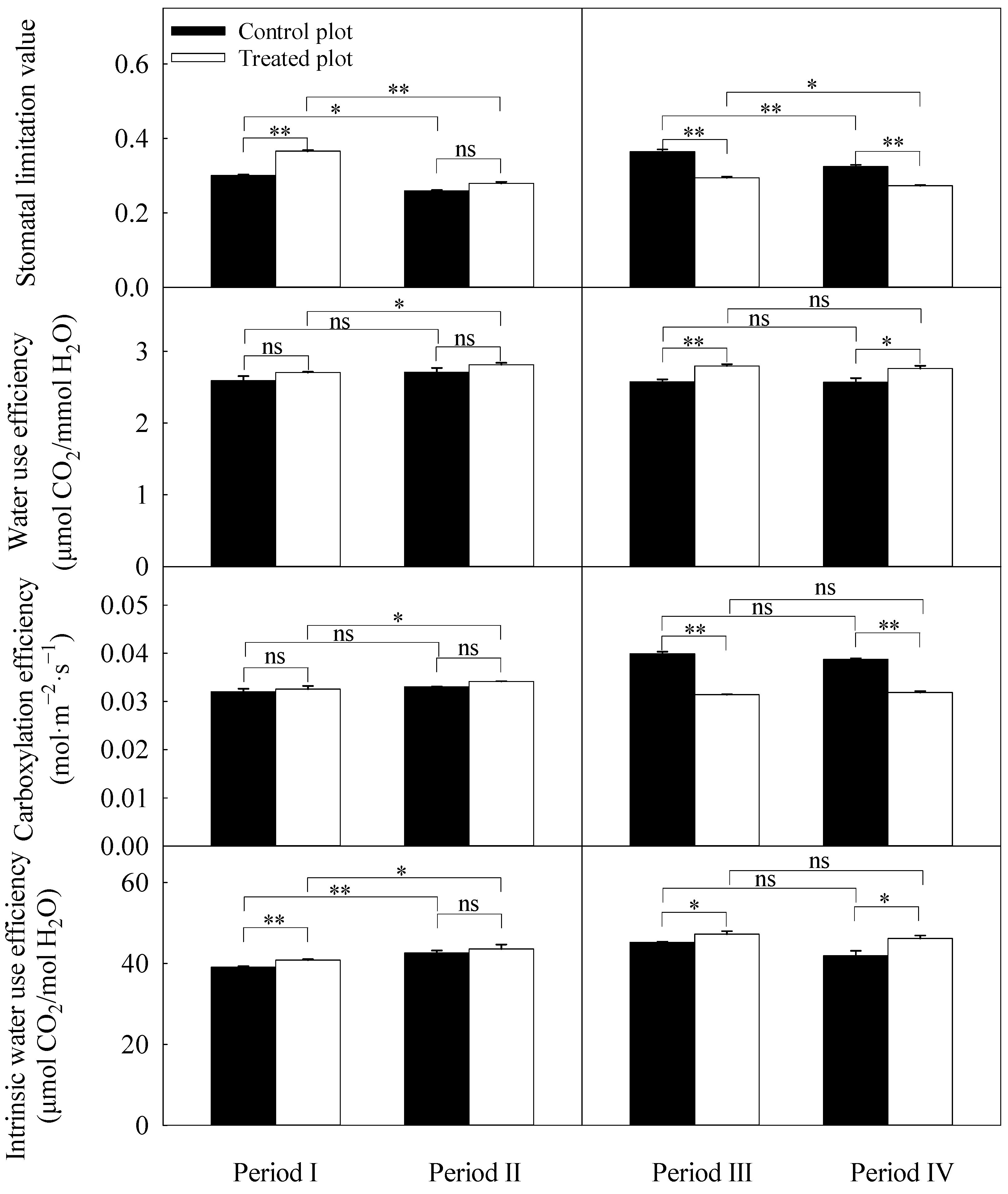
3.3. Differences in Daily Averages of the Photosynthetic Parameters
3.4. Light Response Curve Derived Parameters
3.5. CO2 Response Curve Derived Parameters
4. Discussion
4.1. Changes in Photosynthetic Characteristics under Rainfall Exclusion Treatment
4.2. Stomatal Limitation and Non-Stomatal Limitation of Black Locust
4.3. Effect of Rainfall Exclusion Treatment on Light Response Curve
4.4. The Sensitivity of Photosynthesis to CO2 Concentration Was Affected by Rainfall Exclusion Treatment
4.5. Drought Response Strategies of Black Locust under Rainfall Exclusion Treatment
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rumberg, B.; Mueller, A.; Witt, H.T. New results about the mechanism of photosynthesis. Nature 1962, 194, 854–856. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhang, G.C.; Gu, S.Y.; Xia, J.B.; Zhao, J.K. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation in semiarid loess hilly area. Photosynthetica 2010, 48, 589–595. [Google Scholar] [CrossRef]
- Catoni, R.; Gratani, L. Variations in leaf respiration and photosynthesis ratio in response to air temperature and water availability among Mediterranean evergreen species. J. Arid Environ. 2014, 102, 82–88. [Google Scholar] [CrossRef]
- Yan, W.M.; Zhong, Y.Q.W.; Shangguan, Z.P. A meta-analysis of leaf gas exchange and water status responses to drought. Sci. Rep. 2016, 6, 917–928. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.L.; Li, H.C.; Thorpe, M.R.; Hocart, C.H.; Song, X. Stomatal and mesophyll conductance are dominant limitations to photosynthesis in response to heat stress during severe drought in a temperate and a tropical tree species. Trees-Struct. Funct. 2021, 35, 1613–1626. [Google Scholar] [CrossRef]
- Yin, C.Y.; Berninger, F.; Li, C.Y. Photosynthetic responses of Populus przewalski subjected to drought stress. Photosynthetica 2006, 44, 62–68. [Google Scholar] [CrossRef]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef]
- Vaz, M.; Pereira, J.S.; Gazarini, L.C.; David, T.S.; David, J.S.; Rodrigues, A.; Maroco, J.; Chaves, M.M. Drought-induced photosynthetic inhibition and autumn recovery in two Mediterranean oak species (Quercus ilex and Quercus suber). Tree Physiol. 2010, 30, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Utkhao, W.; Yingjajaval, S. Changes in leaf gas exchange and biomass of Eucalyptus camaldulensis in response to increasing drought stress induced by polyethylene glycol. Trees-Struct. Funct. 2015, 29, 1581–1592. [Google Scholar] [CrossRef]
- Bojovic, M.; Nikolic, N.; Borisev, M.; Pajevic, S.; Zupunski, M.; Horák, R.; Pilipovic, A.; Orlonvic, S.; Stojnic, S. The diurnal time course of leaf gas exchange parameters of pedunculate oak seedlings subjected to experimental drought conditions. Balt. For. 2017, 23, 584–594. [Google Scholar]
- Young, S.S.; Harris, R. Changing patterns of global-scale vegetation photosynthesis, 1982–1999. Int. J. Remote Sens. 2005, 26, 4537–4563. [Google Scholar] [CrossRef]
- Schroth, G.; Laederach, P.; Isaac Martinez-Valle, A.; Bunn, C.; Jassogne, L. Vulnerability to climate change of cocoa in West Africa: Patterns, opportunities and limits to adaptation. Sci. Total Environ. 2016, 556, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.M.; Zhong, Y.Q.W.; Shangguan, Z.P. Responses of different physiological parameter thresholds to soil water availability in four plant species during prolonged drought. Agric. For. Meteorol. 2017, 247, 311–319. [Google Scholar] [CrossRef]
- Lu, Y.; Duursma, R.A.; Farrior, C.E.; Medlyn, B.E.; Feng, X. Optimal stomatal drought response shaped by competition for water and hydraulic risk can explain plant trait covariation. New Phytol. 2020, 225, 1206–1217. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Gao, J.; Zhao, P.; Rao, X. Species-specific transpiration and water use patterns of two pioneer dominant tree species under manipulated rainfall in a low-subtropical secondary evergreen forest. Ecohydrology 2020, 13, 2234–2245. [Google Scholar] [CrossRef]
- Inoue, Y.; Ichie, T.; Kenzo, T.; Yoneyama, A.; Kumagai, T.; Nakashizuka, T. Effects of rainfall exclusion on leaf gas exchange traits and osmotic adjustment in mature canopy trees of Dryobalanops aromatica (Dipterocarpaceae) in a Malaysian tropical rain forest. Tree Physiol. 2017, 37, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Grams, T.E.E.; Hesse, B.D.; Gebhardt, T.; Weikl, F.; Rotzer, T.; Kovacs, B.; Hikino, K.; Hafner, B.D.; Brunn, M.; Bauerle, T.; et al. The Kroof experiment: Realization and efficacy of a recurrent drought experiment plus recovery in a beech/spruce forest. Ecosphere 2021, 12, 3399–3418. [Google Scholar] [CrossRef]
- Limousin, J.M.; Bickford, C.P.; Dickman, L.T.; Pangle, R.E.; Hudson, P.J.; Boutz, A.L.; Gehres, N.; Osuna, J.L.; Pockman, W.T.; McDowell, N.G. Regulation and acclimation of leaf gas exchange in a pinon-juniper woodland exposed to three different precipitation regimes. Plant Cell Environ. 2013, 36, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.M.; Sayer, M.A.; Chambers, J.L.; Barnett, J.P. Interactive effects of fertilization and throughfall exclusion on the physiological responses and whole-tree carbon uptake of mature loblolly pine. Can. J. Bot. Rev. Can. Bot. 2004, 82, 850–861. [Google Scholar] [CrossRef]
- Misson, L.; Limousin, J.M.; Rodriguez, R.; Letts, M.G. Leaf physiological responses to extreme droughts in Mediterranean Quercus ilex forest. Plant Cell Environ. 2010, 33, 1898–1910. [Google Scholar] [CrossRef]
- Limousin, J.M.; Misson, L.; Lavoir, A.V.; Martin, N.K.; Rambal, S. Do photosynthetic limitations of evergreen Quercus ilex leaves change with long-term increased drought severity? Plant Cell Environ. 2010, 33, 863–875. [Google Scholar] [CrossRef]
- Tejera-Nieves, M.; Abraha, M.; Chen, J.Q.; Hamilton, S.K.; Robertson, G.P.; James, B.W. Seasonal decline in leaf photosynthesis in perennial switchgrass explained by sink limitations and water deficit. Front. Plant Sci. 2023, 13, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.J.; Chen, L.D.; Ma, K.M.; Zhou, H.F.; Wang, J. The relationships between land use and soil conditions in the hilly area of the loess plateau in northern Shaanxi, China. Catena 2000, 39, 69–78. [Google Scholar] [CrossRef]
- Lu, Y.; Fu, B.; Feng, X.; Zeng, Y.; Liu, Y.; Chang, R.; Sun, G.; Wu, B. A policy-driven large scale ecological restoration: Quantifying ecosystem services changes in the Loess Plateau of China. PLoS ONE 2012, 7, e31782. [Google Scholar] [CrossRef]
- Sun, Q.; Miao, C.; Duan, Q.; Wang, Y. Temperature and precipitation changes over the Loess Plateau between 1961 and 2011, based on high-density gauge observations. Glob. Planet. Chang. 2015, 132, 1–10. [Google Scholar] [CrossRef]
- Jin, T.T.; Fu, B.J.; Liu, G.H.; Wang, Z. Hydrologic feasibility of artificial forestation in the semi-arid Loess Plateau of China. Hydrol. Earth Syst. Sci. 2011, 15, 2519–2530. [Google Scholar] [CrossRef]
- Mantovani, D.; Veste, M.; Freese, D. Black locust (Robinia pseudoacacia) ecophysiological and morphological adaptations to drought and their consequence on biomass production and water-use efficiency. N. Z. J. For. Sci. 2014, 44, 29–40. [Google Scholar] [CrossRef]
- He, Q.Y.; Yan, M.J.; Miyazawa, Y.; Chen, Q.W.; Du, S. Sap flow changes and climatic responses over multiple-year treatment of rainfall exclusion in a sub-humid black locust plantation. For. Ecol. Manag. 2020, 457, 730–740. [Google Scholar] [CrossRef]
- Keyimu, M.; Li, Z.; Fu, B.; Chen, W.; Wei, J.; Jiao, L.; Gao, G.; Lu, Y. Spatial differences in the radial growth responses of black locust (Robinia pseudoacacia Linn.) to climate on the Loess Plateau, China. Dendrochronologia 2021, 67, 32–43. [Google Scholar] [CrossRef]
- Lyu, J.; Li, G.; Otsuki, K.; Yamanaka, N.; Wang, Y.; Yue, M.; Du, S. Different transpiration and growth patterns of the black locust plantation and natural oak forest on China’s Loess Plateau. Ecohydrology 2023, 16, 23–34. [Google Scholar] [CrossRef]
- Du, S.; Wang, Y.L.; Kume, T.; Zhang, J.G.; Otsuki, K.; Yamanaka, N.; Liu, G.B. Sapflow characteristics and climatic responses in three forest species in the semiarid Loess Plateau region of China. Agric. For. Meteorol. 2011, 151, 1–10. [Google Scholar] [CrossRef]
- Abri, T.; Keseru, Z.; Borovics, A.; Redei, K.; Csajbok, J. Comparison of juvenile, drought tolerant black locust (Robinia pseudoacacia L.) clones with regard to plant physiology and growth characteristics in eastern hungary: Early evaluation. Forests 2022, 13, 292. [Google Scholar] [CrossRef]
- Lyu, J.L.; He, Q.Y.; Chen, Q.W.; Cheng, R.R.; Li, G.Q.; Otsuki, K.; Yamanaka, N.; Du, S. Distinct transpiration characteristics of black locust plantations acclimated to semiarid and subhumid sites in the Loess Plateau, China. Agric. Water Manag. 2022, 262, 107402. [Google Scholar] [CrossRef]
- Ye, Z.P.; Suggett, D.J.; Robakowski, P.; Kang, H.J. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Ye, Z.P.; Robakowski, P.; Suggett, D.J. A mechanistic model for the light response of photosynthetic electron transport rate based on light harvesting properties of photosynthetic pigment molecules. Planta 2013, 237, 837–847. [Google Scholar] [CrossRef]
- Wilson, K.B.; Hanson, P.J.; Mulholland, P.J.; Baldocchi, D.D.; Wullschleger, S.D. A comparison of methods for determining forest evapotranspiration and its components: Sap-flow, soil water budget, eddy covariance and catchment water balance. Agric. For. Meteorol. 2001, 106, 153–168. [Google Scholar] [CrossRef]
- Wang, L.; Dai, Y.X.; Sun, J.Z.; Wan, X.C. Differential hydric deficit responses of Robinia pseudoacacia and Platycladus orientalis in pure and mixed stands in northern China and the species interactions under drought. Trees-Struct. Funct. 2017, 31, 2011–2021. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Q.Y.; Song, M.X.; Wang, N.; Fan, P.X.; Wu, P.; Cui, K.N.; Zheng, P.M.; Du, N.; Wang, H.; et al. Physiological responses of Robinia pseudoacacia and Quercus acutissima seedlings to repeated drought-rewatering under different planting methods. Front. Plant Sci. 2021, 12, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Staniak, M.; Bojarszczuk, J.; Kraska, P.; Kwiatkowski, C.; Harasim, E. Prolonged drought stress induced changes in yield and physiological processes of Trifolium repens and Festulolium braunii. Biol. Plant. 2020, 64, 701–709. [Google Scholar] [CrossRef]
- Gao, H.; Bai, N.; Zhang, Y.; Zhang, X.H.; Zhang, Y.J.; Wang, L.; Wang, E.J.; Tian, Y.Y.; Guo, Y.Y.; Yan, F.; et al. Drought stress alters gas exchange, chlorophyll fluorescence, and antioxidant enzyme activities in Glycyrrhiza uralensis in the Hexi Corridor, China. Russ. J. Plant Physiol. 2022, 69, 69–81. [Google Scholar] [CrossRef]
- Niu, M.G.; Zhao, T.R.; Xu, D.; Liu, C.S.; Liu, Y.; Sun, M.T.; Xie, H.C.; Li, J.H. Physiological responses of chionanthus retusus seedlings to drought and waterlogging stresses. Forests 2023, 14, 429. [Google Scholar] [CrossRef]
- Sim, Y.S.; Yim, S.H.; Choo, Y.S. Photosynthetic and physiological characteristics of the evergreen Ligustrum japonicum and the deciduous Cornus officinalis. J. Plant Biol. 2021, 64, 73–85. [Google Scholar] [CrossRef]
- Song, Y.; Liu, T.; Wang, J.; Lu, Y.; Guo, J.; Dong, Z.; Wen, Y.; Pei, Z. The responses of three dominant species to increased rainfall under different grazing systems in a desert steppe. Hydrol. Process. 2022, 36, 32–44. [Google Scholar] [CrossRef]
- Liu, X.P.; Fan, Y.Y.; Long, J.X.; Wei, R.F.; Kjelgren, R.; Gong, C.M.; Zhao, J. Effects of soil water and nitrogen availability on photosynthesis and water use efficiency of Robinia pseudoacacia seedlings. J. Environ. Sci. 2013, 25, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, M.T.; Maksymowicz, A.; Hura, K.; Dziurka, K.; Ostrowska, A.; Grzesiak, S. Separate or combined effects of soil compaction and/or drought on gas exchange, chlorophyll fluorescence and physiological traits of maize (Zea mays L.) hybrids. J. Agron. Crop Sci. 2023, 209, 689–704. [Google Scholar] [CrossRef]
- Yan, W.M.; Zheng, S.X.; Zhong, Y.Q.W.; Shangguan, Z.P. Contrasting dynamics of leaf potential and gas exchange during progressive drought cycles and recovery in Amorpha fruticosa and Robinia pseudoacacia. Sci. Rep. 2017, 7, 4470–4481. [Google Scholar] [CrossRef]
- Duan, H.L.; Wang, D.F.; Zhao, N.; Huang, G.M.; de Dios, V.R.; Tissue, D.T. Limited hydraulic recovery in seedlings of six tree species with contrasting leaf habits in subtropical China. Front. Plant Sci. 2022, 13, 187–205. [Google Scholar] [CrossRef]
- Xie, T.; Su, P. Canopy and leaf photosynthetic characteristics and water use efficiency of sweet sorghum under drought stress. Russ. J. Plant Physiol. 2012, 59, 224–234. [Google Scholar] [CrossRef]
- Kenzo, T.; Yoneda, R.; Ninomiya, I. Seasonal changes in photosynthesis and starch content in Japanese fir (Abies firma Sieb. et Zucc.) saplings under different levels of irradiance. Trees-Struct. Funct. 2018, 32, 429–439. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 99–110. [Google Scholar] [CrossRef]
- Ghotbi-Ravandi, A.A.; Shahbazi, M.; Shariati, M.; Mulo, P. Effects of mild and severe drought stress on photosynthetic efficiency in tolerant and susceptible barley (Hordeum vulgare L.) genotypes. J. Agron. Crop Sci. 2014, 200, 403–415. [Google Scholar] [CrossRef]
- Feng, S.L.; Sikdar, A.; Wang, J.X.; Li, B.Y.; Lv, G.L.; Ma, X. Response of gas exchange and water-use efficiency in Platycladus orientalis and Amorpha fruticosa to drought episode and rewatering. Pak. J. Agric. Sci. 2020, 57, 1237–1249. [Google Scholar] [CrossRef]
- Fenta, B.A.; Driscoll, S.P.; Kunert, K.J.; Foyer, C.H. Characterization of drought-tolerance traits in nodulated soya beans: The importance of maintaining photosynthesis and shoot biomass under drought-induced limitations on nitrogen metabolism. J. Agron. Crop Sci. 2012, 198, 92–103. [Google Scholar] [CrossRef]
- Benesova, M.; Hola, D.; Fischer, L.; Jedelsky, P.L.; Hnilicka, F.; Wilhelmova, N.; Rothova, O.; Kocova, M.; Prochazkova, D.; Honnerova, J.; et al. The physiology and proteomics of drought tolerance in maize: Early stomatal closure as a cause of lower tolerance to short-term dehydration? PLoS ONE 2012, 7, e38017. [Google Scholar] [CrossRef] [PubMed]
- Pepe, M.; Crescente, M.F.; Varone, L. Effect of water stress on physiological and morphological leaf traits: A comparison among the three widely-spread invasive alien species Ailanthus altissima, Phytolacca americana, and Robinia pseudoacacia. Plants 2022, 11, 899. [Google Scholar] [CrossRef] [PubMed]
- Rapacz, M.; Koscieiniak, J.; Jurczyk, B.; Adamska, A.; Wojcik, M. Different patterns of physiological and molecular response to drought in seedlings of malt- and feed-type barleys (Hordeum vulgare). J. Agron. Crop Sci. 2010, 196, 9–19. [Google Scholar] [CrossRef]
- Snider, J.L.; Collins, G.D.; Whitaker, J.; Perry, C.D.; Chastain, D.R. Electron transport through photosystem II is not limited by a wide range of water deficit conditions in field-grown Gossypium hirsutum. J. Agron. Crop Sci. 2014, 200, 77–82. [Google Scholar] [CrossRef]
- Drake, J.E.; Power, S.A.; Duursma, R.A.; Medlyn, B.E.; Aspinwall, M.J.; Choat, B.; Creek, D.; Eamus, D.; Maier, C.; Pfautsch, S.; et al. Stomatal and non-stomatal limitations of photosynthesis for four tree species under drought: A comparison of model formulations. Agric. For. Meteorol. 2017, 247, 454–466. [Google Scholar] [CrossRef]
- Yang, M.; Gao, X.D.; Wang, S.F.; Zhang, X.; Cai, Y.H.; Song, X.L.; Siddique, K.H.M.; Zhao, X.N. Photosynthetic response to deep soil water deficit in a semiarid apple tree plantation. J. Hydrol. 2023, 617, 161–171. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gao, H.; Li, Y.H.; Wang, L.; Kong, D.S.; Guo, Y.Y.; Yan, F.; Wang, Y.W.; Lu, K.; Tian, J.W.; et al. Effect of water stress on photosynthesis, chlorophyll fluorescence parameters and water use efficiency of common reed in the Hexi Corridor. Russ. J. Plant Physiol. 2019, 66, 556–563. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Xia, J.; Xing, X.; Zhang, S. Critical effects on the photosynthetic efficiency and stem sap flow of poplar in the Yellow River Delta in response to soil water. J. For. Res. 2021, 32, 2485–2498. [Google Scholar] [CrossRef]
- Fan, Z.; Quan, Q.; Li, Y.; Jun, Y.; Wang, S. Exploring the best model for describing light-response curves in two Epimedium species. Technol. Health Care 2015, 23, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Toda, M.; Fukuzawa, K.; Nakamura, M.; Miyata, R.; Wang, X.; Doi, K.; Tabata, A.; Shibata, H.; Yoshida, T.; Hara, T. Photosynthetically distinct responses of an early-successional tree, Betula ermanii, following a defoliating disturbance: Observational results of a manipulated typhoon-mimic experiment. Trees-Struct. Funct. 2018, 32, 1789–1799. [Google Scholar] [CrossRef]
- Liu, J.W.; Zhang, R.H.; Zhang, G.C.; Guo, J.; Dong, Z. Effects of soil drought on photosynthetic traits and antioxidant enzyme activities in Hippophae rhamnoides seedlings. J. For. Res. 2017, 28, 255–263. [Google Scholar] [CrossRef]
- Xia, J.B.; Zhao, Z.G.; Sun, J.K.; Liu, J.T.; Zhao, Y.Y. Response of stem sap flow and leaf photosynthesis in Tamarix chinensis to soil moisture in the Yellow River Delta, China. Photosynthetica 2017, 55, 368–377. [Google Scholar] [CrossRef]
- Leon-Burgos, A.F.; Unigarro, C.; Balaguera-Lopez, H.E. Can prolonged conditions of water deficit alter photosynthetic performance and water relations of coffee plants in central-west Colombia? S. Afr. J. Bot. 2022, 149, 366–375. [Google Scholar] [CrossRef]
- Wu, T.; Tan, N.; Tissue, D.T.; Huang, J.; Duan, H.; Su, W.; Song, Y.; Liu, X.; Liu, Y.; Li, X.; et al. Physiological traits and response strategies of four subtropical tree species exposed to drought. Environ. Exp. Bot. 2022, 203, 5046–5057. [Google Scholar] [CrossRef]
- Sheshshayee, M.S.; Shivashankar, K.S.; Ramaswamy, G.S.; Devendra, R.; Shankar, A.G.; Prasad, T.G.; Udayakumar, M. A new method to generate diffferent CO2 concentrations for developing a CO2-response curve. Curr. Sci. 1992, 62, 538–540. [Google Scholar]
- Zhao, X.Y.; Li, Y.; Zheng, M.; Bian, X.Y.; Liu, M.R.; Sun, Y.S.; Jiang, J.; Wang, F.W.; Li, S.C.; Cui, Y.H.; et al. Comparative analysis of growth and photosynthetic characteristics of (Populus simonii × P-nigra) × (P-nigra × P-simonii) hybrid clones of different ploidides. PLoS ONE 2015, 10, e0119259. [Google Scholar] [CrossRef]
- Gao, S.; Yan, Q.; Chen, L.; Song, Y.; Li, J.; Fu, C.; Dong, M. Effects of ploidy level and haplotype on variation of photosynthetic traits: Novel evidence from two Fragaria species. PLoS ONE 2017, 12, e0179899. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, T.; Li, C.R.; Xie, H.B.; Zhang, G.C. Photosynthetic CO2 response to soil water and its simulation using different models in leaves of two species. Photosynthetica 2020, 58, 790–798. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, W.Z.; Wen, Z.J. Photosynthetic response of two shrubs to rainfall pulses in desert regions of northwestern China. Photosynthetica 2012, 50, 109–119. [Google Scholar] [CrossRef]
- Kosugi, Y.; Takanashi, S.; Ueyama, M.; Ohkubo, S.; Tanaka, H.; Matsumoto, K.; Yoshifuji, N.; Ataka, M.; Sakabe, A. Determination of the gas exchange phenology in an evergreen coniferous forest from 7 years of eddy covariance flux data using an extended big-leaf analysis. Ecol. Res. 2013, 28, 373–385. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Kane, M.B.; Markewitz, D.; Teskey, R.O.; Akers, M.K.; Stokes, T.A.; Pell, C.J.; Qi, J. Fertilization increased leaf water use efficiency and growth of Pinus taeda subjected to five years of throughfall reduction. Can. J. For. Res. 2018, 48, 227–236. [Google Scholar] [CrossRef]
- Lapa, G.; Morandini, F.; Ferrat, L. Sap flow and photosynthetic response to climate and drought of Pinus nigra in a Mediterranean natural forest. Trees-Struct. Funct. 2017, 31, 1711–1721. [Google Scholar] [CrossRef]
- Nadal-Sala, D.; Sabaté, S.; Sánchez-Costa, E.; Poblador, S.; Sabater, F.; Gracia, C. Growth and water use performance of four co-occurring riparian tree species in a Mediterranean riparian forest. For. Ecol. Manag. 2017, 396, 132–142. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Before the Two Weeks SWC | Measurement Period SWC | ||
|---|---|---|---|---|
| Control Plot | Treated Plot | Control Plot | Treated Plot | |
| Period I (28–30 July 2021) | 0.267 ± 0.020 | 0.234 ± 0.033 | 0.268 ± 0.004 | 0.239 ± 0.006 |
| Period II (9–11 September 2021) | 0.265 ± 0.037 | 0.216 ± 0.035 | 0.284 ± 0.003 | 0.252 ± 0.004 |
| Period III (23–25 June 2022) | 0.265 ± 0.006 | 0.232 ± 0.004 | 0.255 ± 0.002 | 0.224 ± 0.001 |
| Period IV (1–3 August 2022) | 0.294 ± 0.005 | 0.243 ± 0.007 | 0.283 ± 0.002 | 0.244 ± 0.001 |
| Source of Variation | df | Pn | gs | Ci | Tr |
|---|---|---|---|---|---|
| Treatment | 1 | 0.001 | <0.001 | 0.328 | <0.001 |
| Period | 3 | 0.016 | 0.005 | <0.001 | 0.015 |
| Treatment × Period | 3 | 0.014 | 0.005 | 0.001 | 0.012 |
| Source of Variation | df | Ls | WUE | CE | WUEi |
|---|---|---|---|---|---|
| Treatment | 1 | 0.328 | <0.001 | <0.001 | <0.001 |
| Period | 3 | <0.001 | 0.341 | 0.046 | 0.046 |
| Treatment × Period | 3 | 0.001 | 0.597 | 0.021 | 0.021 |
| Study Period | Plot | LSP | LCP | AQE | Pmax | Rd |
|---|---|---|---|---|---|---|
| Period I | Control | 1726.12 ± 35.20 a | 21.58 ± 3.28 a | 0.06 ± 0.003 a | 16.89 ± 0.11 a | 1.07 ± 0.19 a |
| Treated | 1663.20 ± 22.91 b | 28.25 ± 2.78 b | 0.05 ± 0.003 b | 16.53 ± 0.14 b | 1.40 ± 0.12 b | |
| Period II | Control | 1881.37 ± 107.30 a | 17.21 ± 1.87 a | 0.07 ± 0.003 a | 16.98 ± 0.20 a | 1.03 ± 0.10 a |
| Treated | 1823.86 ± 79.81 a | 18.38 ± 2.43 a | 0.07 ± 0.004 a | 16.91 ± 0.11 a | 1.15 ± 0.20 a | |
| Periods I and II | pControl | 0.022 | 0.032 | 0.001 | 0.414 | 0.685 |
| Periods I and II | pTreated | 0.006 | 0.001 | 0.001 | 0.001 | 0.044 |
| Period III | Control | 1707.53 ± 32.69 a | 22.69 ± 0.79 a | 0.06 ± 0.001 a | 16.80 ± 0.03 a | 1.27 ± 0.05 a |
| Treated | 1648.55 ± 19.14 b | 31.70 ± 2.33 b | 0.05 ± 0.003 b | 16.37 ± 0.09 b | 1.58 ± 0.05 b | |
| Period IV | Control | 1868.64 ± 24.25 a | 18.30 ± 1.07 a | 0.07 ± 0.002 a | 17.02 ± 0.07 a | 1.12 ± 0.04 a |
| Treated | 1751.47 ± 25.95 b | 30.76 ± 0.79 b | 0.05 ± 0.001 b | 16.52 ± 0.07 b | 1.44 ± 0.07 b | |
| Periods III and IV | pControl | 0.009 | 0.011 | 0.019 | 0.04 | 0.027 |
| Periods III and IV | pTreated | 0.011 | 0.633 | 0.384 | 0.155 | 0.09 |
| Study Period | Plot | CSP | CCP | Rp | Pmax | CE |
|---|---|---|---|---|---|---|
| Period III | Control | 1297.84 ± 27.18 a | 60.35 ± 0.35 a | 4.28 ± 0.02 a | 27.15 ± 0.11 a | 0.08 ± 0.0004 a |
| Treated | 1198.60 ± 10.36 b | 66.33 ± 0.70 b | 4.57 ± 0.07 b | 26.31 ± 0.12 b | 0.07 ± 0.0004 b | |
| Period IV | Control | 1255.56 ± 18.63 a | 57.09 ± 0.43 a | 4.09 ± 0.04 a | 27.52 ± 0.39 a | 0.08 ± 0.0001 a |
| Treated | 1198.90 ± 12.53 b | 61.17 ± 0.44 b | 4.28 ± 0.03 b | 26.41 ± 0.26 b | 0.07 ± 0.0002 b | |
| Period III and IV | pControl | 0.153 | 0.001 | 0.014 | 0.307 | 0.37 |
| Period III and IV | pTreated | 0.98 | 0.002 | 0.005 | 0.676 | 0.079 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Wang, Y.; Li, G.; Du, S. Effects of Rainfall Exclusion Treatment on Photosynthetic Characteristics of Black Locust in the Sub-Humid Region of the Loess Plateau, China. Plants 2024, 13, 704. https://doi.org/10.3390/plants13050704
Guo H, Wang Y, Li G, Du S. Effects of Rainfall Exclusion Treatment on Photosynthetic Characteristics of Black Locust in the Sub-Humid Region of the Loess Plateau, China. Plants. 2024; 13(5):704. https://doi.org/10.3390/plants13050704
Chicago/Turabian StyleGuo, Haining, Yiran Wang, Guoqing Li, and Sheng Du. 2024. "Effects of Rainfall Exclusion Treatment on Photosynthetic Characteristics of Black Locust in the Sub-Humid Region of the Loess Plateau, China" Plants 13, no. 5: 704. https://doi.org/10.3390/plants13050704
APA StyleGuo, H., Wang, Y., Li, G., & Du, S. (2024). Effects of Rainfall Exclusion Treatment on Photosynthetic Characteristics of Black Locust in the Sub-Humid Region of the Loess Plateau, China. Plants, 13(5), 704. https://doi.org/10.3390/plants13050704