
Coconut (Cocos nucifera (L.)) Water Improves Glucose Uptake with Concomitant Modulation of Antioxidant and Purinergic Activities in Isolated Rat Psoas Muscles


Abstract
1. Introduction
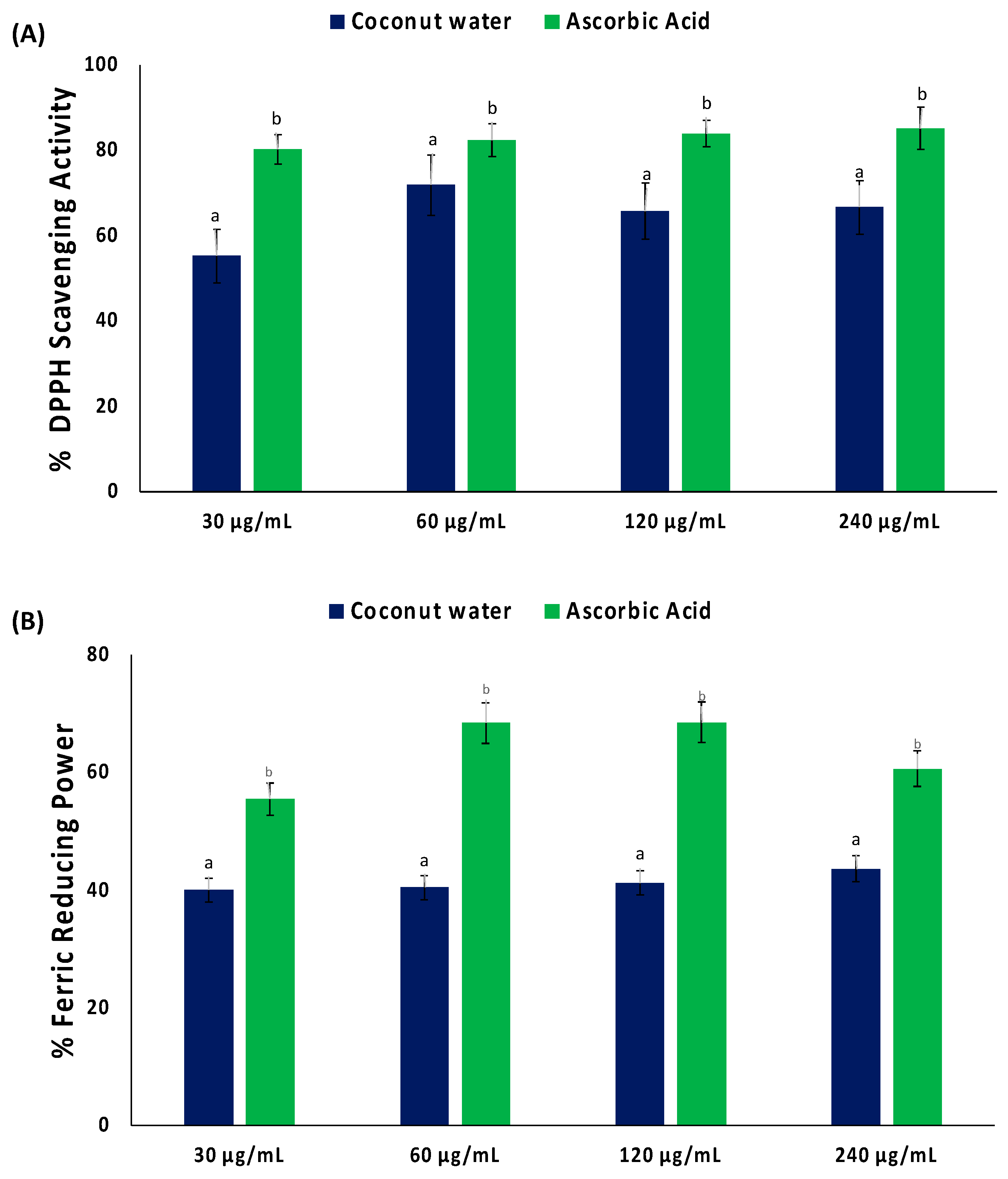
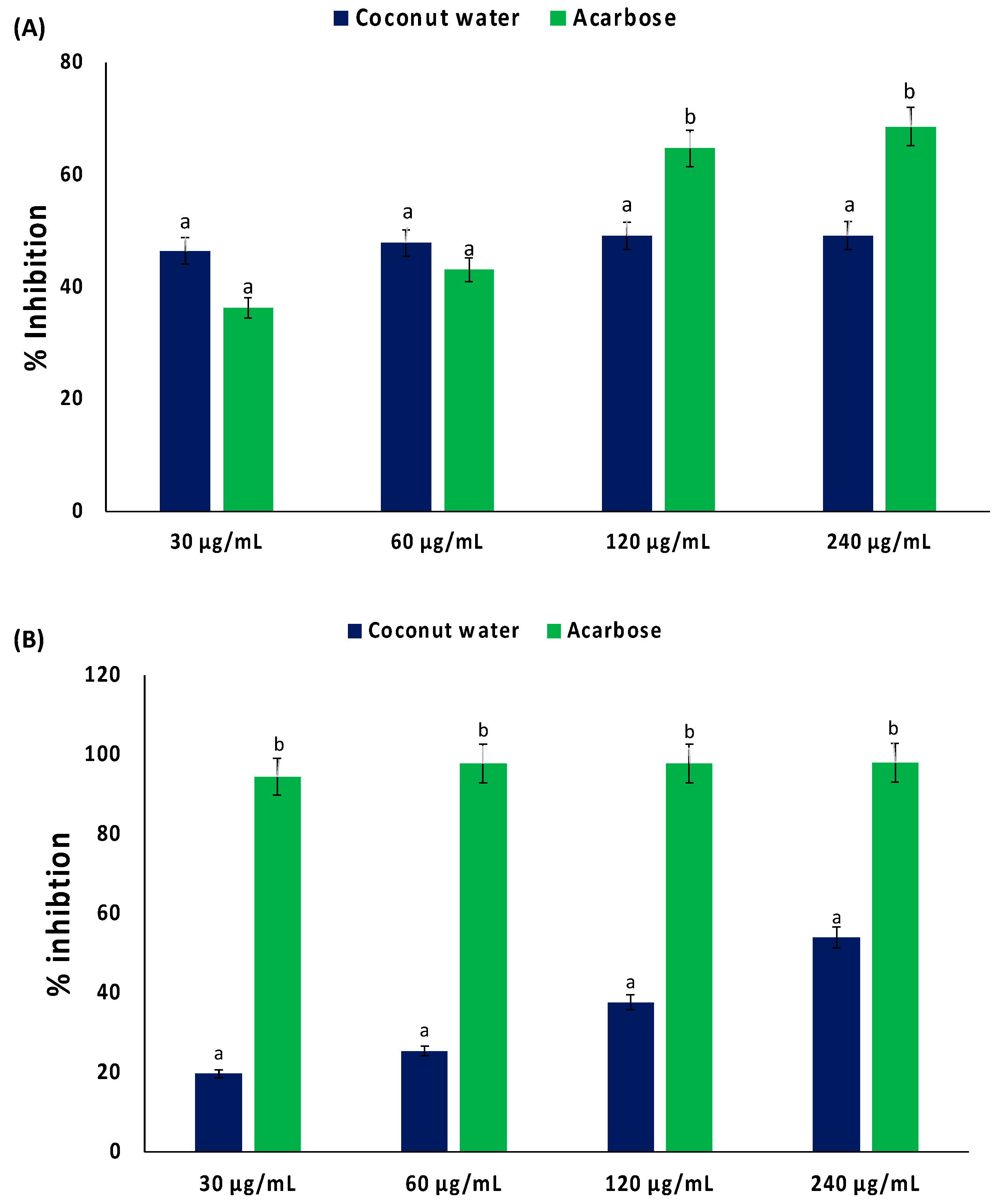
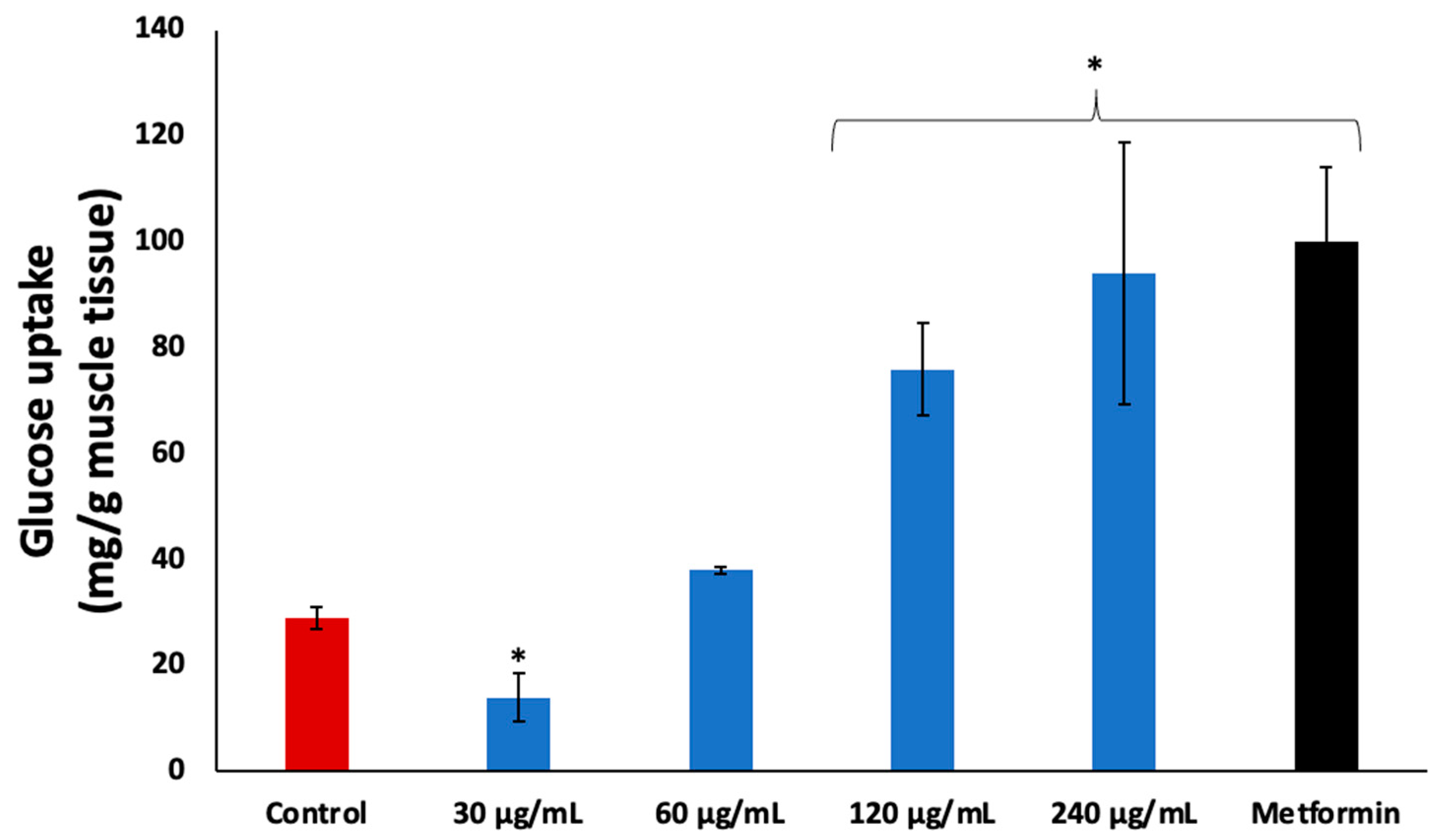
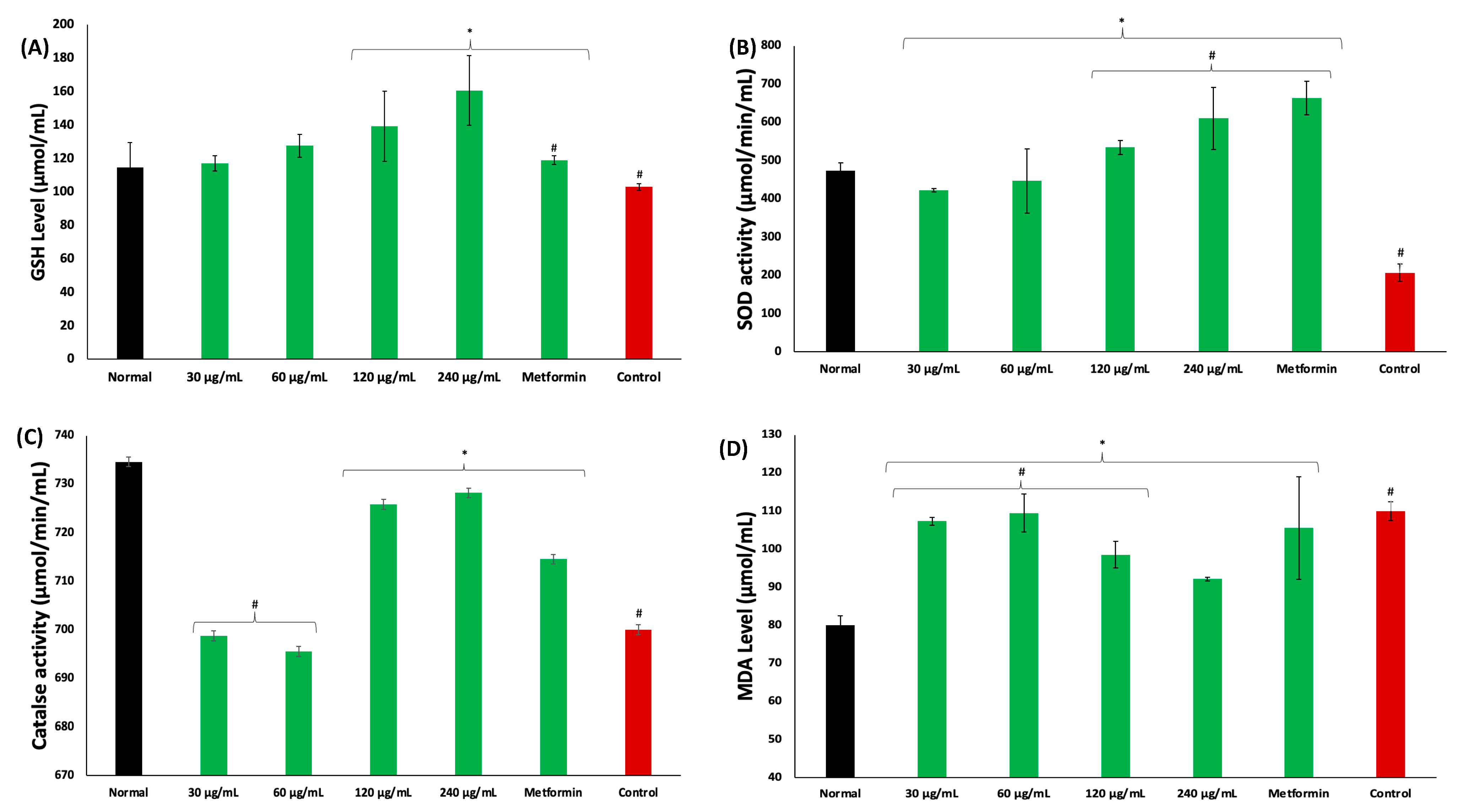
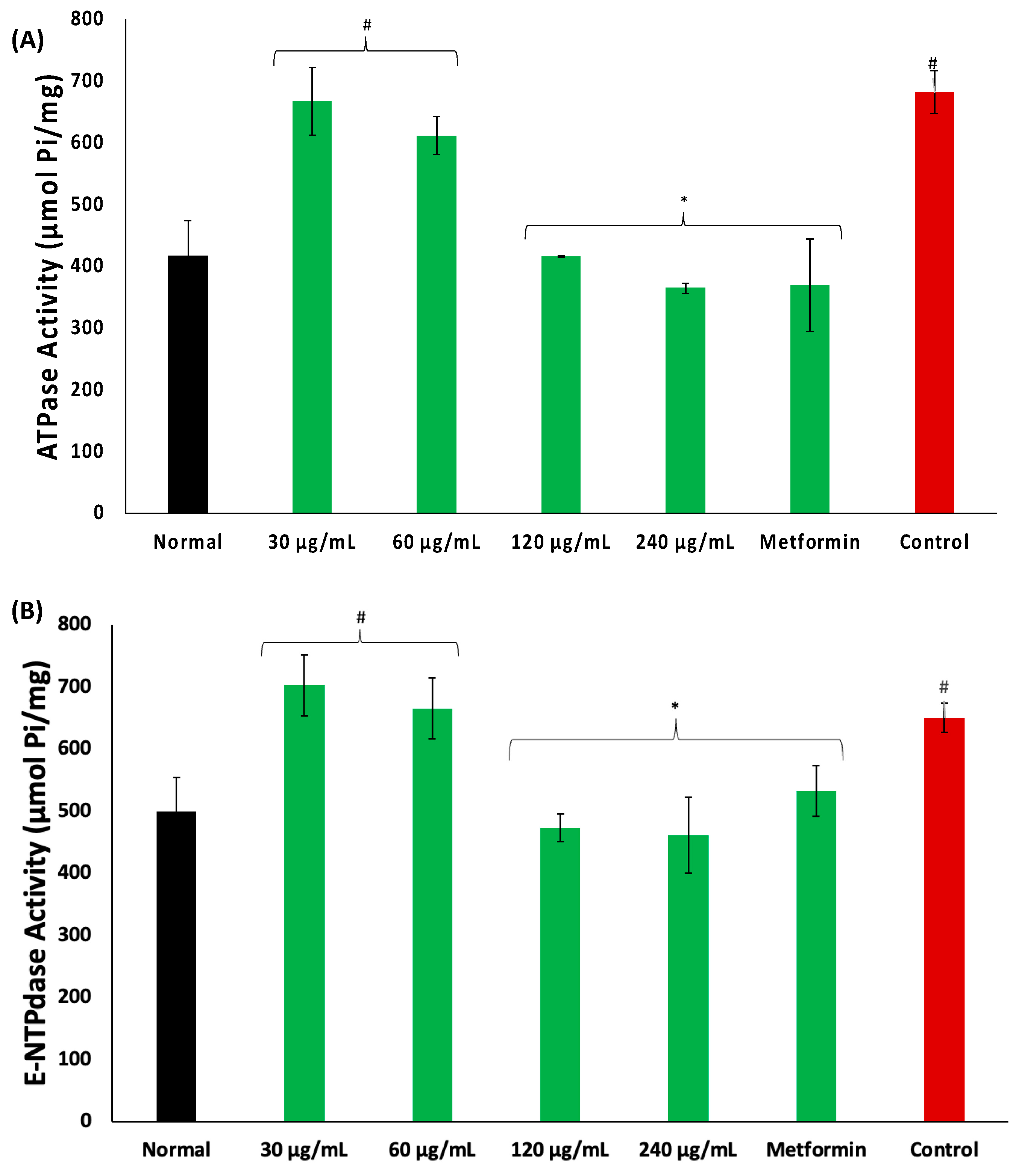
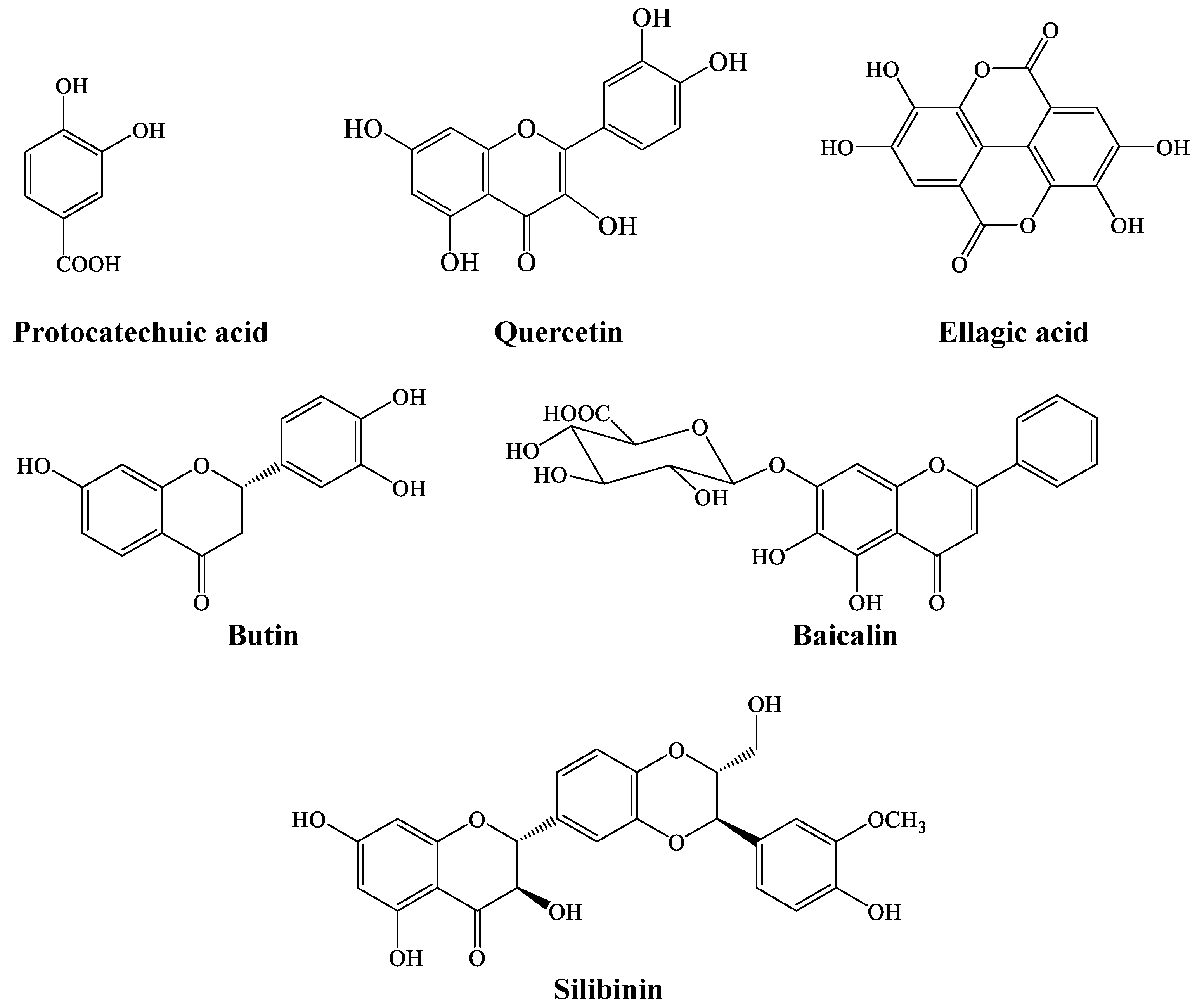
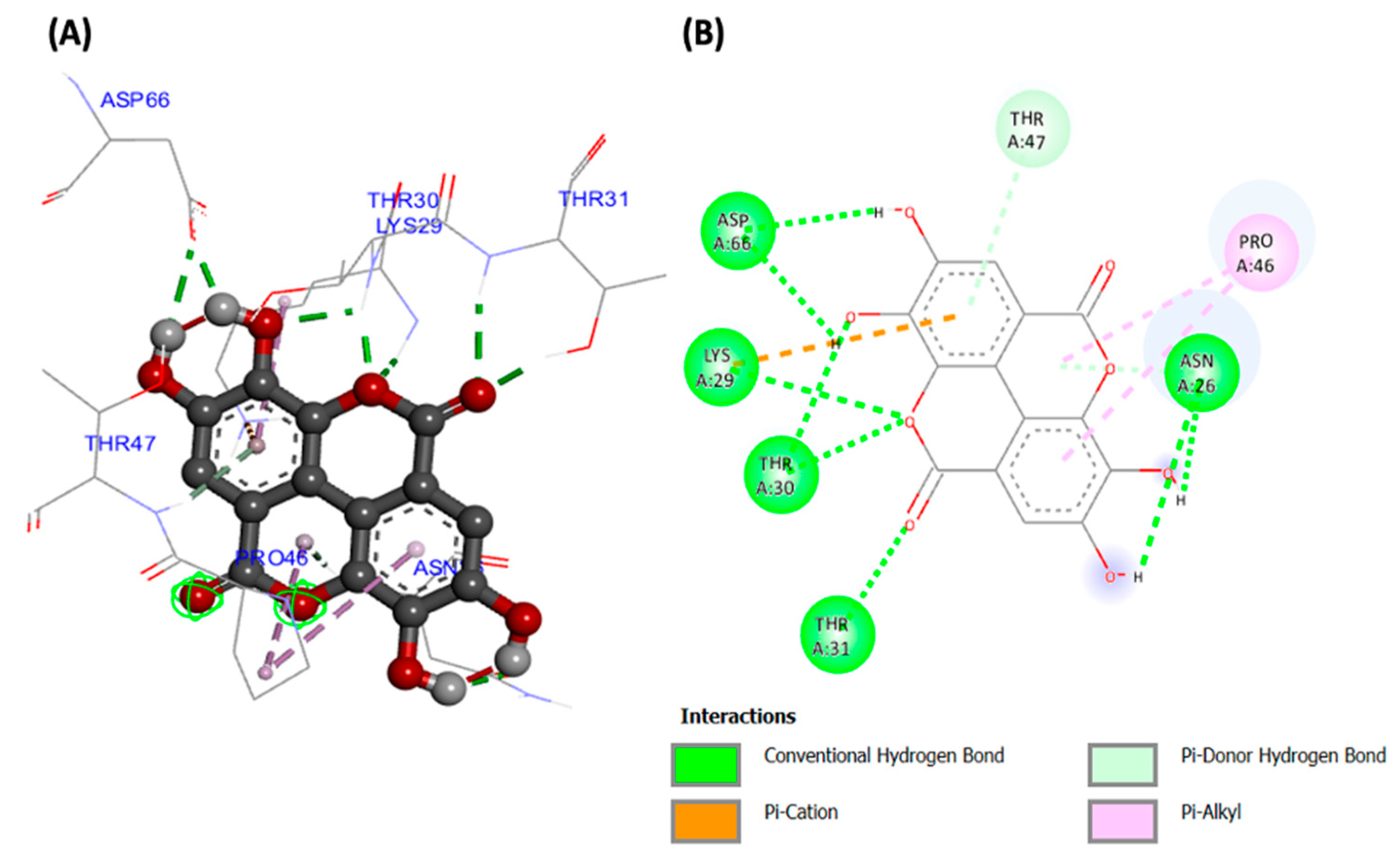
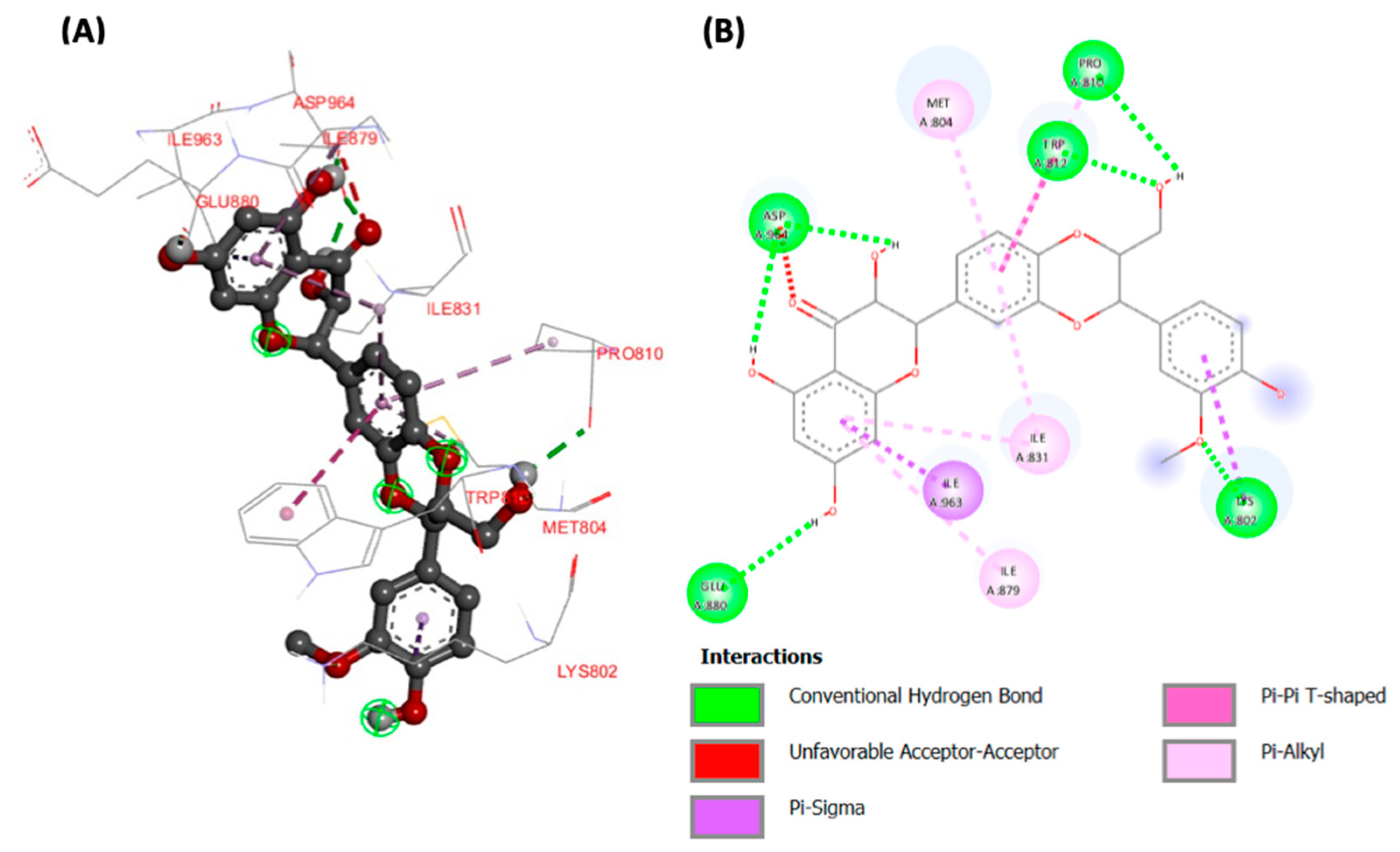
2. Results
3. Discussion
4. Materials and Methods
4.1. Coconut Water
4.2. In Vitro Antioxidant Activity
4.3. Inhibition of Carbohydrate Digestive Enzymes
- i.
- α-glucosidase activity
- ii.
- α-amylase activity
4.4. Animals for Ex Vivo Studies
4.5. Muscle Glucose Uptake
4.6. Determination of Oxidative Stress Markers
- i.
- Reduced glutathione (GSH) level
- ii.
- Superoxide dismutase (SOD) enzyme activity
- iii.
- Catalase enzyme activity
- iv.
- Malondialdehyde (MDA) level
4.7. Determination of Purinergic Enzymes Activities
- v.
- ATPase activity
- vi.
- ENTPDase activity
4.8. LC-MS Analysis of Coconut Water
4.9. Molecular Docking Analysis
- i.
- Protein Target Selection and Preparation
- ii.
- Ligand Selections and Preparations
- iii.
- Molecular Docking
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IDF. IDF Diabetes Atlas. Available online: https://diabetesatlas.org/idfawp/resource-files/2021/07/IDF_Atlas_10th_Edition_2021.pdf (accessed on 14 July 2023).
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress—A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef]
- Merz, K.E.; Thurmond, D.C. Role of skeletal muscle in insulin resistance and glucose uptake. Compr. Physiol. 2011, 10, 785–809. [Google Scholar]
- Fujimoto, B.A.; Young, M.; Nakamura, N.; Ha, H.; Carter, L.; Pitts, M.W.; Torres, D.; Noh, H.-L.; Suk, S.; Kim, J.K. Disrupted glucose homeostasis and skeletal-muscle-specific glucose uptake in an exocyst knockout mouse model. J. Biol. Chem. 2021, 296, 100482. [Google Scholar] [CrossRef]
- Hulett, N.A.; Scalzo, R.L.; Reusch, J.E. Glucose uptake by skeletal muscle within the contexts of type 2 diabetes and exercise: An integrated approach. Nutrients 2022, 14, 647. [Google Scholar] [CrossRef]
- Zhang, H.; Qi, G.; Wang, K.; Yang, J.; Shen, Y.; Yang, X.; Chen, X.; Yao, X.; Gu, X.; Qi, L. Oxidative stress: Roles in skeletal muscle atrophy. Biochem. Pharmacol. 2023, 214, 115664. [Google Scholar] [CrossRef]
- Campbell, I. Oral antidiabetic drugs: Their properties and recommended use. Prescriber 2007, 18, 56–74. [Google Scholar] [CrossRef]
- Bailey, C.; Turner, R. Metformin. N. Engl. J. Med. 1996, 334, 574–579. [Google Scholar] [CrossRef]
- Preetha, P.; Devi, V.G.; Rajamohan, T. Comparative effects of mature coconut water (Cocos nucifera) and glibenclamide on some biochemical parameters in alloxan induced diabetic rats. Rev. Bras. Farmacogn. 2013, 23, 481–487. [Google Scholar] [CrossRef]
- Preetha, P.P.; Devi, V.G.; Rajamohan, T. Effects of coconut water on carbohydrate metabolism and pancreatic pathology of alloxan induced diabetic rats. Eur. J. Integr. Med. 2013, 5, 234–240. [Google Scholar] [CrossRef]
- Pinto, I.F.; Silva, R.P.; Filho, A.d.B.C.; Dantas, L.S.; Bispo, V.S.; Matos, I.A.; Otsuka, F.A.; Santos, A.C.; Matos, H.R. Study of antiglycation, hypoglycemic, and nephroprotective activities of the green dwarf variety coconut water (Cocos nucifera L.) in alloxan-induced diabetic rats. J. Med. Food 2015, 18, 802–809. [Google Scholar] [CrossRef]
- Zhang, X.; Peng, L.; Dai, Y.; Sheng, X.; Chen, S.; Xie, Q. Effects of coconut water on retina in diabetic rats. Evid.-Based Complement. Altern. Med. 2020, 2020, 9450634. [Google Scholar] [CrossRef]
- Preetha, P.; Devi, V.G.; Rajamohan, T. Hypoglycemic and antioxidant potential of coconut water in experimental diabetes. Food Funct. 2012, 3, 753–757. [Google Scholar] [CrossRef]
- Preetha, P.P.; Devi, V.G.; Rajamohan, T. Mature coconut water exhibits antidiabetic and antithrombotic potential via L-arginine-nitric oxide pathway in alloxan induced diabetic rats. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 575–583. [Google Scholar] [CrossRef]
- Akpro, L.; Gbogouri, G.; Konan, B.; Issali, A.; Konan, K.; Brou, K.; Nemlin, G. Phytochemical compounds, antioxidant activity and non-enzymatic browning of sugars extracted from the water of immature coconut (Cocos nucifera L.). Sci. Afr. 2019, 6, e00123. [Google Scholar] [CrossRef]
- Sofowora, A. Research on medicinal plants and traditional medicine in Africa. J. Altern. Complement. Med. 1996, 2, 365–372. [Google Scholar] [CrossRef]
- Chukwuma, E.C.; Soladoye, M.O.; Feyisola, R.T. Traditional medicine and the future of medicinal Plants in Nigeria. J. Med. Plants Stud. 2015, 3, 23–29. [Google Scholar]
- Hossain, U.; Das, A.K.; Ghosh, S.; Sil, P.C. An overview on the role of bioactive α-glucosidase inhibitors in ameliorating diabetic complications. Food Chem. Toxicol. 2020, 145, 111738. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, M.; Song, F.; Wen, Y.; Li, N.; Zhang, Y. The effects of different drying methods on the structural characteristics and bioactivities in vitro of polysaccharides from coconut (Cocos nucifera L.) water. J. Food Meas. Charact. 2022, 16, 2499–2509. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef] [PubMed]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Ribeiro, D.; Freitas, M.; Fernandes, E. Flavonoids as potential agents in the management of type 2 diabetes through the modulation of α-amylase and α-glucosidase activity: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 3137–3207. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Salau, V.F.; Xiao, X.; Matsabisa, M.G.; Koorbanally, N.A.; Islam, M.S. Bioactive compounds of African star apple (Chrysophyllum albidum G. Don) and its modulatory effect on metabolic activities linked to type 2 diabetes in isolated rat psoas muscle. J. Food Biochem. 2021, 45, e13576. [Google Scholar] [CrossRef] [PubMed]
- Erukainure, O.L.; Salau, V.F.; Atolani, O.; Ravichandran, R.; Banerjee, P.; Preissner, R.; Koorbanally, N.A.; Islam, M.S. L-leucine stimulation of glucose uptake and utilization involves modulation of glucose–lipid metabolic switch and improved bioenergetic homeostasis in isolated rat psoas muscle ex vivo. Amino Acids 2021, 53, 1135–1151. [Google Scholar] [CrossRef]
- Boersma, G.J.; Johansson, E.; Pereira, M.J.; Heurling, K.; Skrtic, S.; Lau, J.; Katsogiannos, P.; Panagiotou, G.; Lubberink, M.; Kullberg, J. Altered glucose uptake in muscle, visceral adipose tissue, and brain predict whole-body insulin resistance and may contribute to the development of type 2 diabetes: A combined PET/MR study. Horm. Metab. Res. 2018, 50, 627–639. [Google Scholar]
- Nova, F.S.; Chasani, S.; Hussanna, A.; Zulaikhah, S.T. Tender Coconut Water Inhibits the Process of Lipid Peroxidation, Reduce Glucose Levels and Increase Plasma Insulin in Pregnant Diabetic Rats. Pharmacogn. J. 2020, 12, 162–167. [Google Scholar] [CrossRef]
- Mastrocola, R.; Reffo, P.; Penna, F.; Tomasinelli, C.E.; Boccuzzi, G.; Baccino, F.M.; Aragno, M.; Costelli, P. Muscle wasting in diabetic and in tumor-bearing rats: Role of oxidative stress. Free Radic. Biol. Med. 2008, 44, 584–593. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Ibeji, C.U.; Koorbanally, N.A.; Islam, M.S. Umbelliferone stimulates glucose uptake; modulates gluconeogenic and nucleotide-hydrolyzing enzymes activities, and dysregulated lipid metabolic pathways in isolated psoas muscle. J. Funct. Foods 2020, 67, 103847. [Google Scholar] [CrossRef]
- Halim, M.; Halim, A. The effects of inflammation, aging and oxidative stress on the pathogenesis of diabetes mellitus (type 2 diabetes). Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1165–1172. [Google Scholar] [CrossRef]
- Lima, J.E.; Moreira, N.C.; Sakamoto-Hojo, E.T. Mechanisms underlying the pathophysiology of type 2 diabetes: From risk factors to oxidative stress, metabolic dysfunction, and hyperglycemia. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2022, 874, 503437. [Google Scholar] [CrossRef]
- Maduka, O.O.; Jacob, O.I.; Uzochukwu, O.I.; Chukwuemeka, A.E.; Nwadiogbu, O.V. Effect of Coconut Water on Lipid Peroxidation and Some Antioxidant Status of Diabetic Rats. Int. J. Diabetes Metab. Disord. 2022, 7, 151–158. [Google Scholar]
- Santos, J.L.; Bispo, V.S.; BC FILHO, A.; Pinto, I.F.; Dantas, L.S.; Vasconcelos, D.F.; Abreu, F.F.; Melo, D.A.; Matos, I.A.; Freitas, F.P. Evaluation of chemical constituents and antioxidant activity of coconut water (Cocus nucifera L.) and caffeic acid in cell culture. An. Acad. Bras. Ciências 2013, 85, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Yegutkin, G.; Novak, I. ATP release, generation and hydrolysis in exocrine pancreatic duct cells. Purinergic Signal. 2015, 11, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. History of ectonucleotidases and their role in purinergic signaling. Biochem. Pharmacol. 2021, 187, 114322. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Arnett, T.R.; Orriss, I.R. Purinergic signalling in the musculoskeletal system. Purinergic Signal. 2013, 9, 541–572. [Google Scholar] [CrossRef] [PubMed]
- Fatima, N.; Hafizur, R.M.; Hameed, A.; Ahmed, S.; Nisar, M.; Kabir, N. Ellagic acid in Emblica officinalis exerts anti-diabetic activity through the action on β-cells of pancreas. Eur. J. Nutr. 2017, 56, 591–601. [Google Scholar] [CrossRef]
- Zhang, R.; Kang, K.A.; Piao, M.J.; Chang, W.Y.; Maeng, Y.H.; Chae, S.; Lee, I.K.; Kim, B.J.; Hyun, J.W. Butin reduces oxidative stress-induced mitochondrial dysfunction via scavenging of reactive oxygen species. Food Chem. Toxicol. 2010, 48, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Abdelmoaty, M.A.; Ibrahim, M.; Ahmed, N.; Abdelaziz, M. Confirmatory studies on the antioxidant and antidiabetic effect of quercetin in rats. Indian J. Clin. Biochem. 2010, 25, 188–192. [Google Scholar] [CrossRef]
- Harini, R.; Pugalendi, K.V. Antioxidant and antihyperlipidaemic activity of protocatechuic acid on streptozotocindiabetic rats. Redox Rep. 2010, 15, 71–80. [Google Scholar] [CrossRef]
- Szkudelski, T.; Szkudelska, K. The Anti-Diabetic Potential of Baicalin: Evidence from Rodent Studies. Int. J. Mol. Sci. 2023, 25, 431. [Google Scholar] [CrossRef]
- El-Far, M.; Negm, A.; El-Azim, A.; Wahdan, M. Antioxidant therapeutic actions of medicinal phytochemicals, silymarin, and silibinin, on streptozotocin diabetic rats: First novel comparative assessment of structural recoveries of histological and ultrastructural changes on islets of Langerhans, beta cells, mitochondria and nucleus. Int. J. Pharm. Pharm. Sci 2016, 8, 1–8. [Google Scholar]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Ng, Y.; Ramm, G.; Lopez, J.A.; James, D.E. Rapid activation of Akt2 is sufficient to stimulate GLUT4 translocation in 3T3-L1 adipocytes. Cell Metab. 2008, 7, 348–356. [Google Scholar] [CrossRef]
- Sakamoto, K.; Arnolds, D.E.; Fujii, N.; Kramer, H.F.; Hirshman, M.F.; Goodyear, L.J. Role of Akt2 in contraction-stimulated cell signaling and glucose uptake in skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2006, 291, E1031–E1037. [Google Scholar] [CrossRef]
- Świderska, E.; Strycharz, J.; Wroblewski, A. Role of PI3K/AKT Pathway in Insulin-Mediated Glucose Uptake. In Blood Glucose Levels; IntechOpen: London, UK, 2018. [Google Scholar]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Braca, A.; Sortino, C.; Politi, M.; Morelli, I.; Mendez, J. Antioxidant activity of flavonoids from Licania licaniaeflora. J. Ethnopharmacol. 2002, 79, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademosun, A.O. Shaddock peels (Citrus maxima) phenolic extracts inhibit α-amylase, α-glucosidase and angiotensin I-converting enzyme activities: A nutraceutical approach to diabetes management. Diabetes Metab. Syndr. Clin. Res. Rev. 2011, 5, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Chukwuma, C.I.; Islam, M.S. Effects of xylitol on carbohydrate digesting enzymes activity, intestinal glucose absorption and muscle glucose uptake: A multi-mode study. Food Funct. 2015, 6, 955–962. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, P.; Das, B.; Viswanathan, P. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar] [PubMed]
- Hadwan, M.H.; Abed, H.N. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief 2016, 6, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, P.; Soulsby, M. Lipid peroxidation in rat brain is increased by simulated weightlessness and decreased by a soy-protein diet. Ann. Clin. Lab. Sci. 2002, 32, 188–192. [Google Scholar] [PubMed]
- Adewoye, O.; Bolarinwa, A.; Olorunsogo, O. Ca++, Mg++-ATPase activity in insulin-dependent and non-insulin dependent diabetic Nigerians. Afr. J. Med. Med. Sci. 2000, 29, 195–199. [Google Scholar] [PubMed]
- Erukainure, O.L.; Mopuri, R.; Oyebode, O.A.; Koorbanally, N.A.; Islam, M.S. Dacryodes edulis enhances antioxidant activities, suppresses DNA fragmentation in oxidative pancreatic and hepatic injuries; and inhibits carbohydrate digestive enzymes linked to type 2 diabetes. Biomed. Pharmacother. 2017, 96, 37–47. [Google Scholar] [CrossRef]
- Schetinger, M.R.C.; Morsch, V.M.; Bonan, C.D.; Wyse, A.T. NTPDase and 5′-nucleotidase activities in physiological and disease conditions: New perspectives for human health. Biofactors 2007, 31, 77–98. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Ogunsuyi, O.B.; Oboh, G. Alkaloid extracts from Jimson weed (Datura stramonium L.) modulate purinergic enzymes in rat brain. Neurotoxicology 2016, 56, 107–117. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Oyebode, O.A.; Sokhela, M.K.; Koorbanally, N.A.; Islam, M.S. Caffeine–rich infusion from Cola nitida (kola nut) inhibits major carbohydrate catabolic enzymes; abates redox imbalance; and modulates oxidative dysregulated metabolic pathways and metabolites in Fe2+-induced hepatic toxicity. Biomed. Pharmacother. 2017, 96, 1065–1074. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Activities | Coconut Water | Ascorbic Water |
|---|---|---|
| (μg/mL) | ||
| DPPH | 34.09 | 0.09 |
| FRAP | >1000 | 0.23 |
| α-glucosidase | 338.27 | 70.58 |
| α-amylase | 219.73 | 0.02 |
| Protein | Compounds | Estimated Free Binding Energy (kcal/mol) | Estimated Inhibition Constant, Ki in nM | Hydrogen Bonds (Distance Å) | Electrostatic Interaction | Electrostatic Interaction |
|---|---|---|---|---|---|---|
| 3DOE | Ellagic acid | −8.99 | 257.83 | LYS29 (2.92223) THR30 (1.83044) THR30 (2.10013) THR31 (2.21939) THR31 (1.77066) ASP66 (1.74398) ASP66 (2.20146) ASN26 (2.19584) ASN26 (2.12805) ASN26 (2.24013) THR47 (3.12677) | LYS29 | LYS29 PRO46 (x2) |
| Butin | −8.35 | 760.49 | ALA27 (2.46832) GLY28 (2.19884) LYS29 (2.25839) LYS29 (1.96312) THR30 (2.9807) GLN70 (1.91152) GLN70 (2.55934) THR47 (1.75311) | PRO46 | ||
| Quercetin | −7.5 | 3.16 | GLY69 (2.12109) GLN70 (2.4799) LYS126 (2.15314) THR31 (1.97452) THR47 (2.13694) ASP25 (2.51937) GLY68 (3.52814) ASN26 (2.54491) THR47 (3.0409) | LYS29 | PRO46 (x2) | |
| Protocatechuic acid | −6.13 | 32.35 | GLY28 (2.08006) LYS29 (2.21339) LYS29 (1.89555) THR31 (1.93536) LEU24 (2.32277) ASP25 (3.15522) | |||
| Baicalin | −8.45 | 635.2 | ALA27 (2.22248) GLY28 (2.09824) LYS29 (2.244) LYS29 (1.75644) LYS29 (1.7697) THR30 (2.91587) THR31 (2.25074) THR30 (3.00405) VAL160 (3.07995) | LYS126 VAL160 ALA159 | ||
| Silibinin | −8.48 | 605.62 | THR43 (2.6353) ILE44 (2.08981) GLN70 (2.45455) ILE44 (1.98277) ILE41 (2.11285) LEU24 (2.59707) ASP25 (3.12523) | LYS126 | ASN26 ALA27 PRO46 (x2) ILE44 | |
| 7JWZ | Ellagic acid | −8.55 | 543.33 | VAL882 (2.04853) ALA885 (2.11578) ASP964 (1.81248) ASP964 (2.2236) | MET953 (x2) ILE963 (x2) TRP812 TYR867 VAL882 ALA885 ILE831 | |
| Butin | −7.07 | 6.55 | VAL882 (1.79667) GLU880 (1.98775) GLU880 (2.09533) | ILE831 MET804 MET953 PRO810 ILE831 ILE963 | ||
| Quercetin | −7.25 | 4.88 | VAL882 (1.74091) ASP964 (2.09705) GLU880 (2.10031) ASP964 (2.12019) | ILE879 ILE963 (x2) MET953 (x2) TRP812 TYR867 ILE831 | ||
| Protocatechuic acid | −5.17 | 162.78 | ASP964 (2.07765) GLU880 (2.25826) ASP964 (2.00965) ASP964 (1.87388) | ALE879 TYR867 ILE831 ILE963 | ||
| Baicalin | −8.46 | 630.16 | ALA885 (2.80904) ALA885 (1.89965) | MET953 (x3) TRP812 (x2) TYR867 ALA885 VAL882 ILE963 | ||
| Silibinin | −9.03 | 240.83 | LYS802 (1.80046) TRP812 (2.2991) ASP964 (1.7375) ASP964 (1.85756) GLU880 (2.2275) PRO810 (2.2449) | LYS802 ILE963 TRP812 MET804 PRO810 ILE831(x2) ILE879 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erukainure, O.L.; Chukwuma, C.I. Coconut (Cocos nucifera (L.)) Water Improves Glucose Uptake with Concomitant Modulation of Antioxidant and Purinergic Activities in Isolated Rat Psoas Muscles. Plants 2024, 13, 665. https://doi.org/10.3390/plants13050665
Erukainure OL, Chukwuma CI. Coconut (Cocos nucifera (L.)) Water Improves Glucose Uptake with Concomitant Modulation of Antioxidant and Purinergic Activities in Isolated Rat Psoas Muscles. Plants. 2024; 13(5):665. https://doi.org/10.3390/plants13050665
Chicago/Turabian StyleErukainure, Ochuko L., and Chika I. Chukwuma. 2024. "Coconut (Cocos nucifera (L.)) Water Improves Glucose Uptake with Concomitant Modulation of Antioxidant and Purinergic Activities in Isolated Rat Psoas Muscles" Plants 13, no. 5: 665. https://doi.org/10.3390/plants13050665
APA StyleErukainure, O. L., & Chukwuma, C. I. (2024). Coconut (Cocos nucifera (L.)) Water Improves Glucose Uptake with Concomitant Modulation of Antioxidant and Purinergic Activities in Isolated Rat Psoas Muscles. Plants, 13(5), 665. https://doi.org/10.3390/plants13050665