

Growth Enhancement and Resistance of Banana Plants to Fusarium Wilt Disease as Affected by Silicate Compounds and Application Frequency

 , and
, and
Abstract
1. Introduction
2. Results
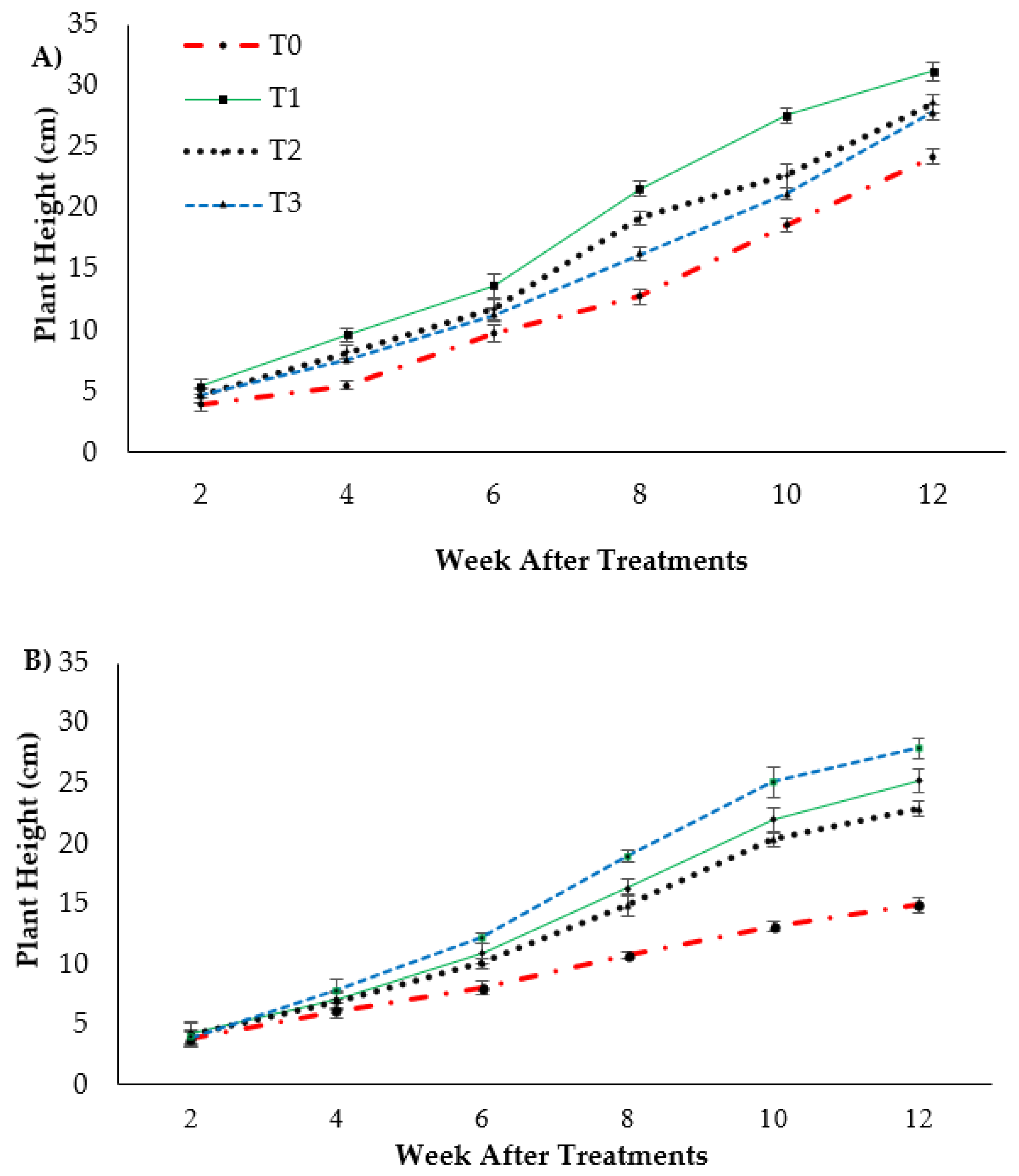
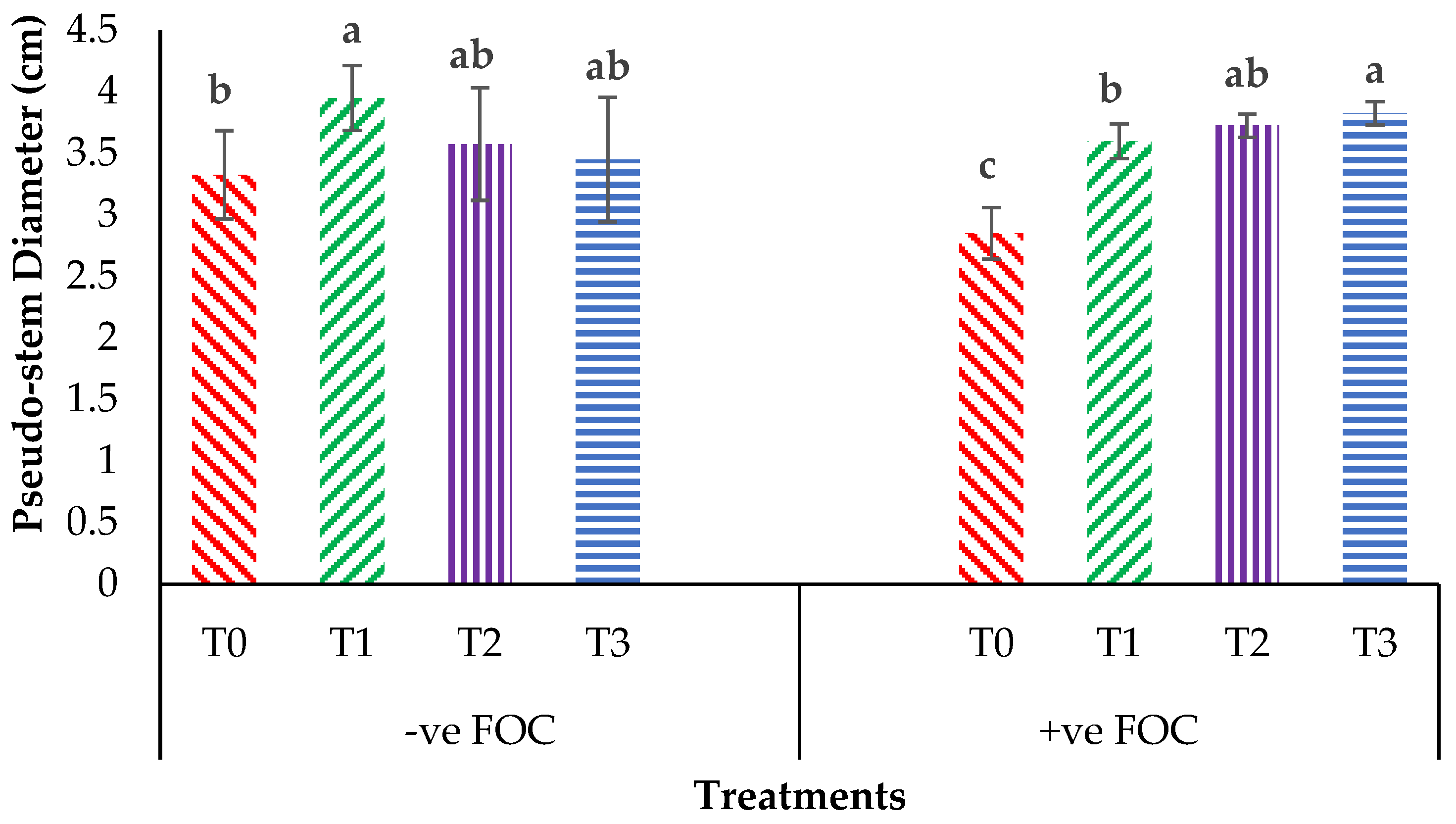
2.1. Crop Growth Performance
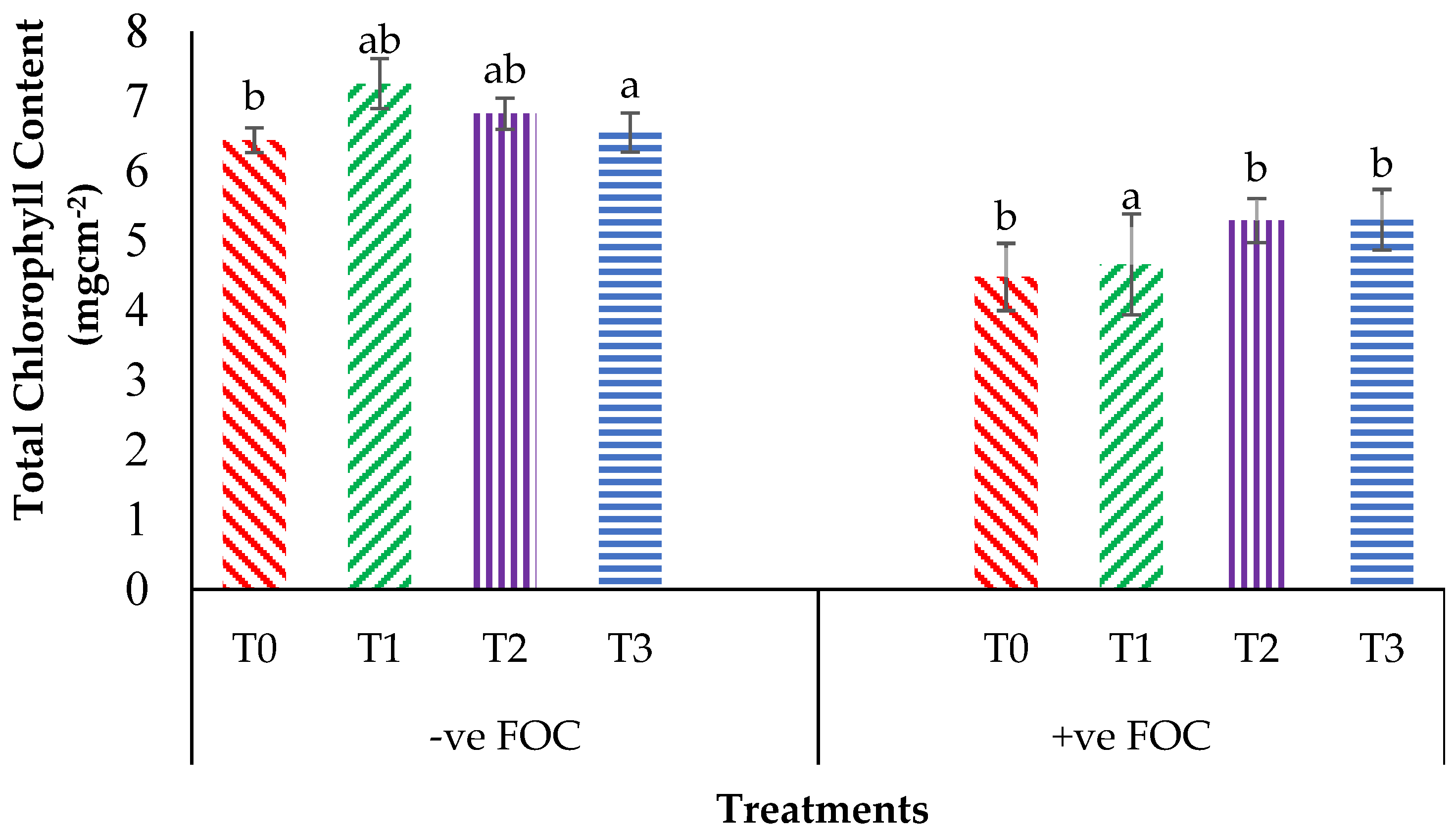
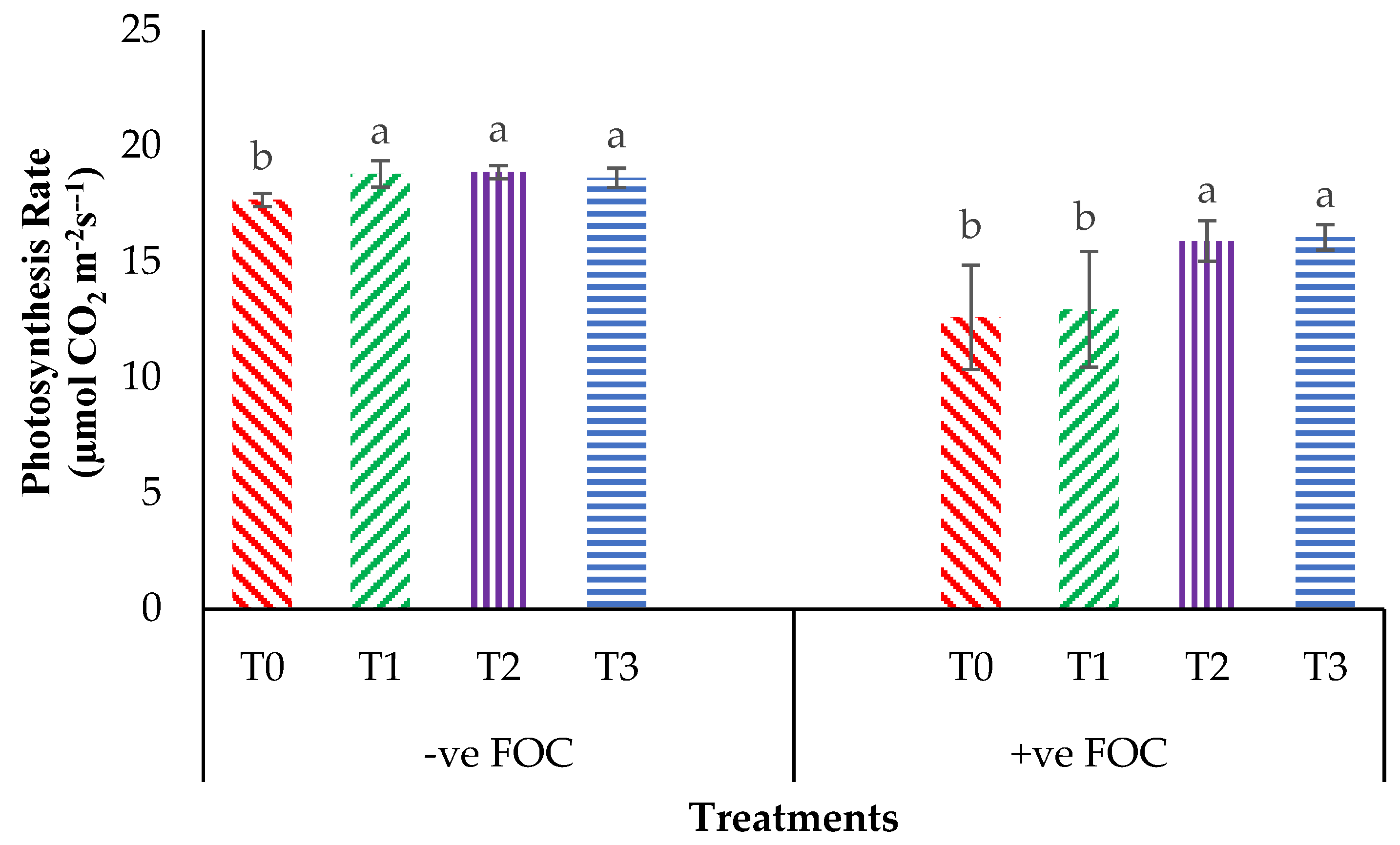
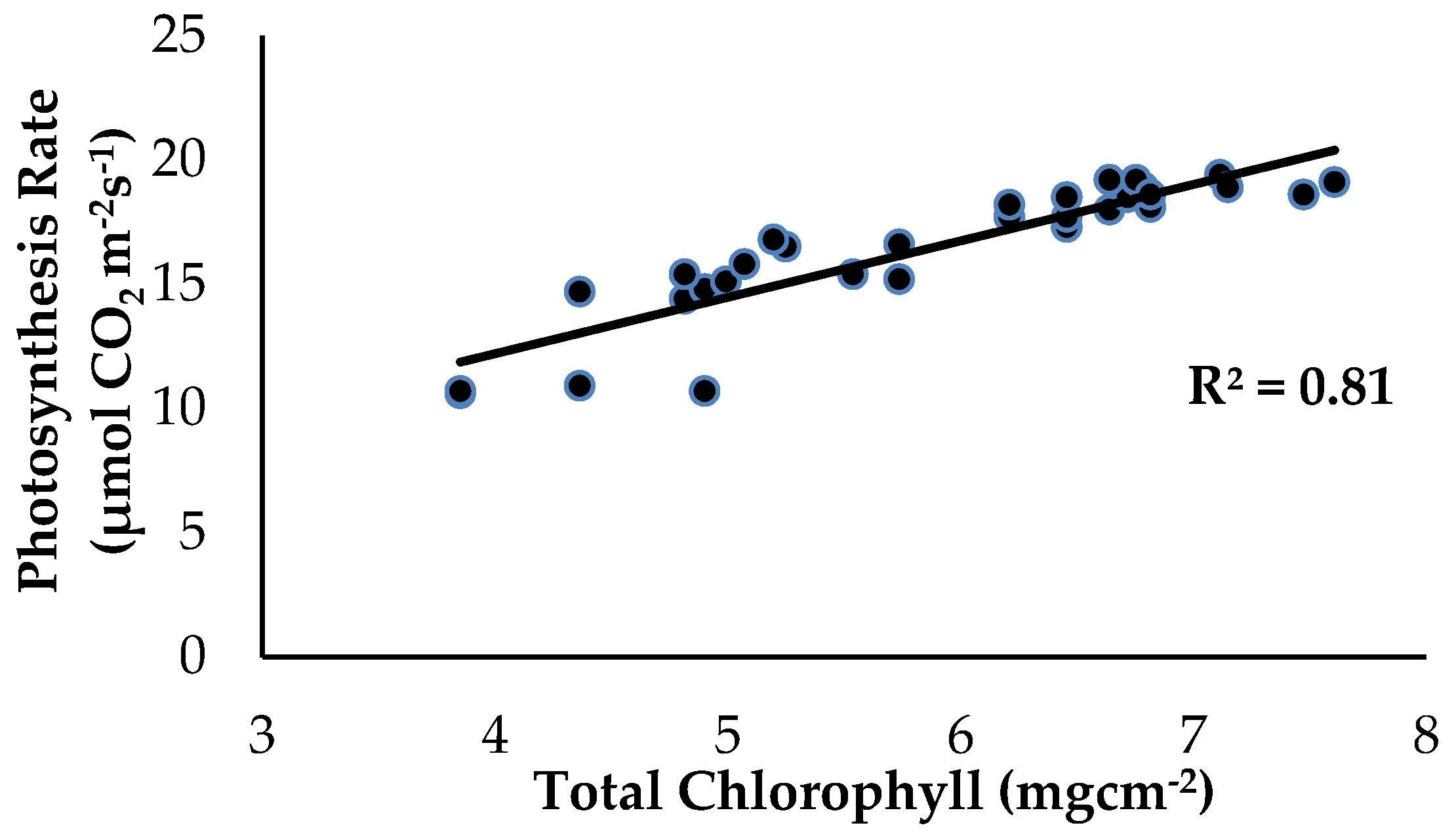
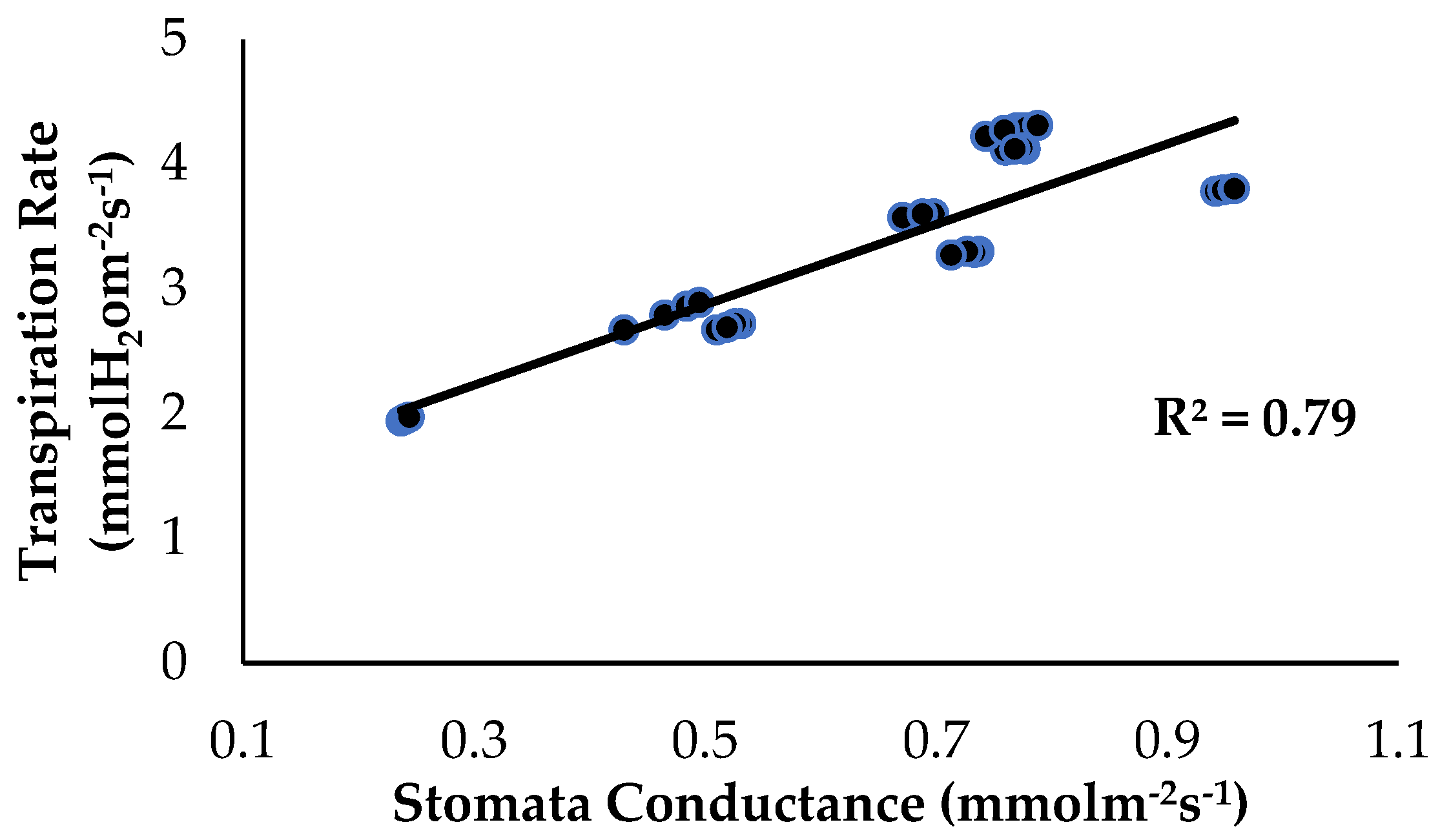
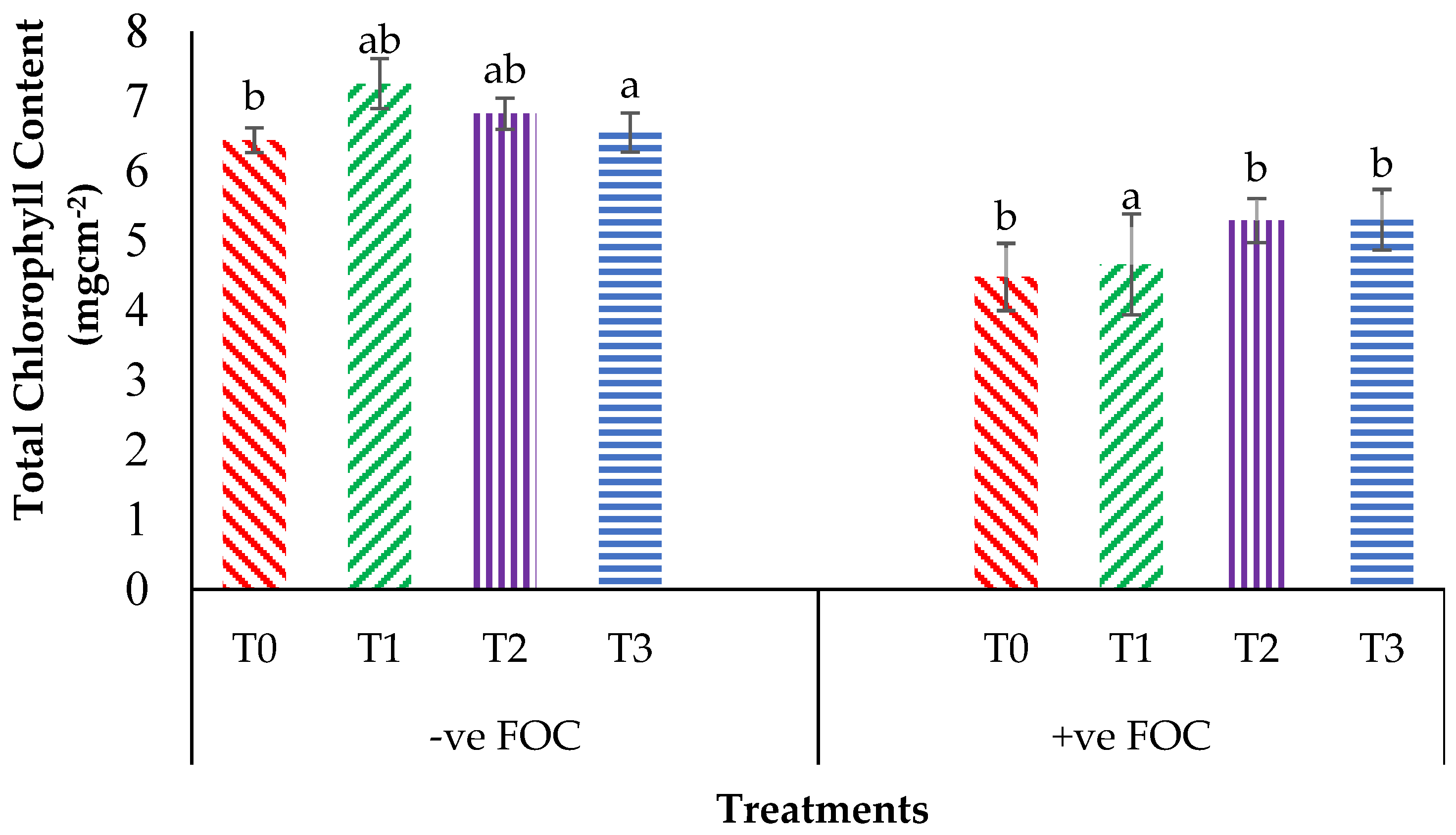
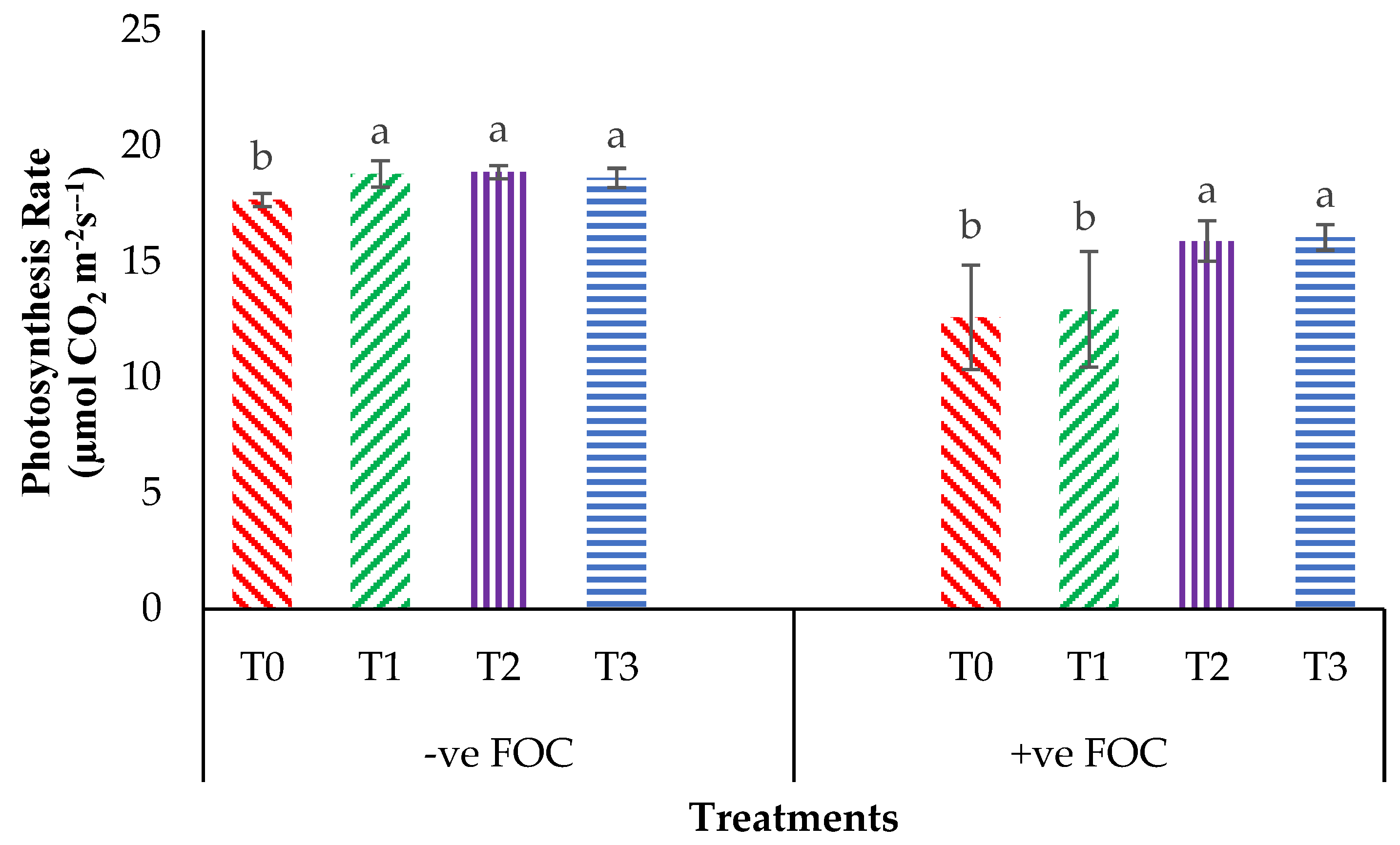
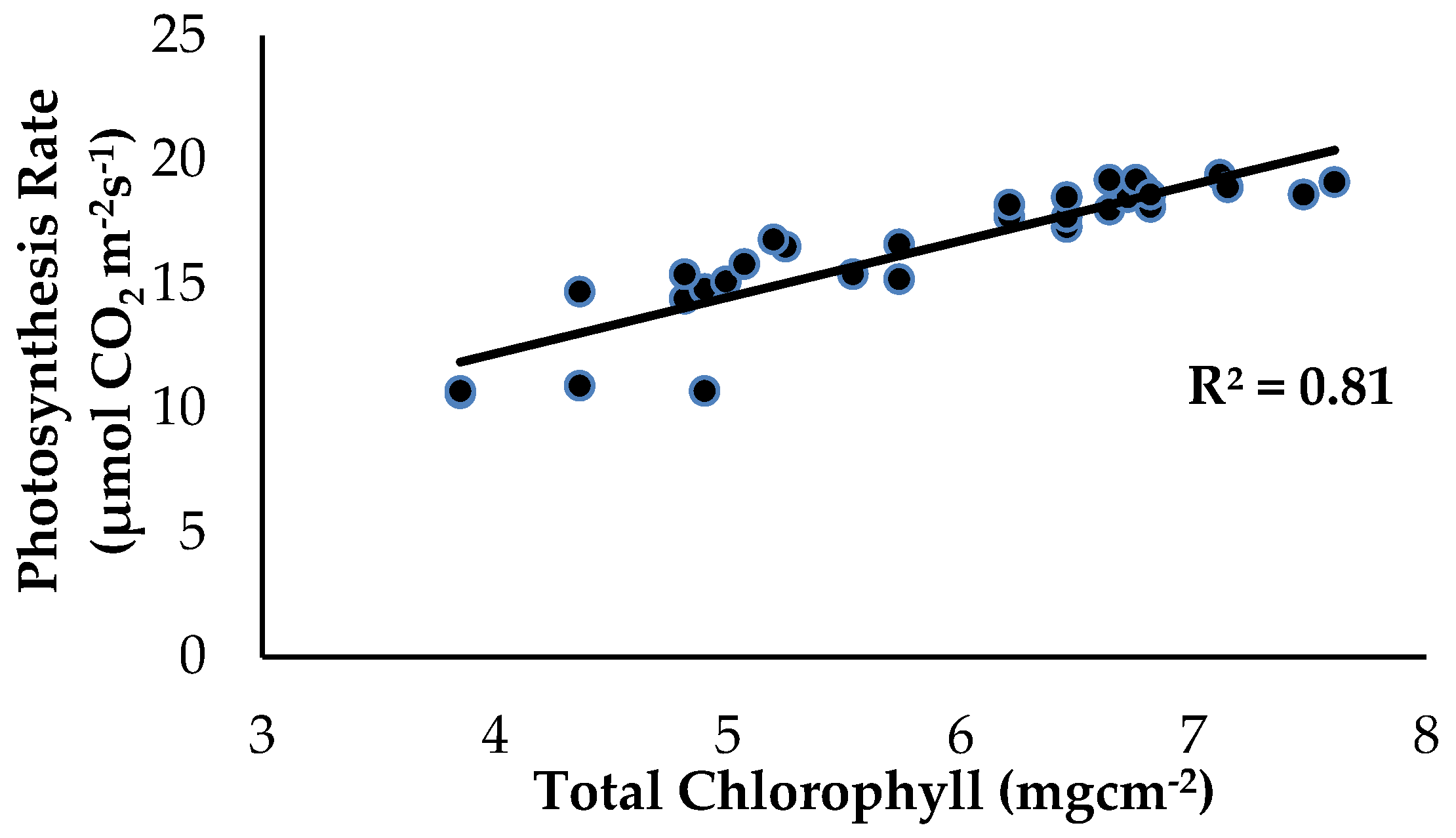
2.2. Total Chlorophyll Content and Leaf Gas Exchange
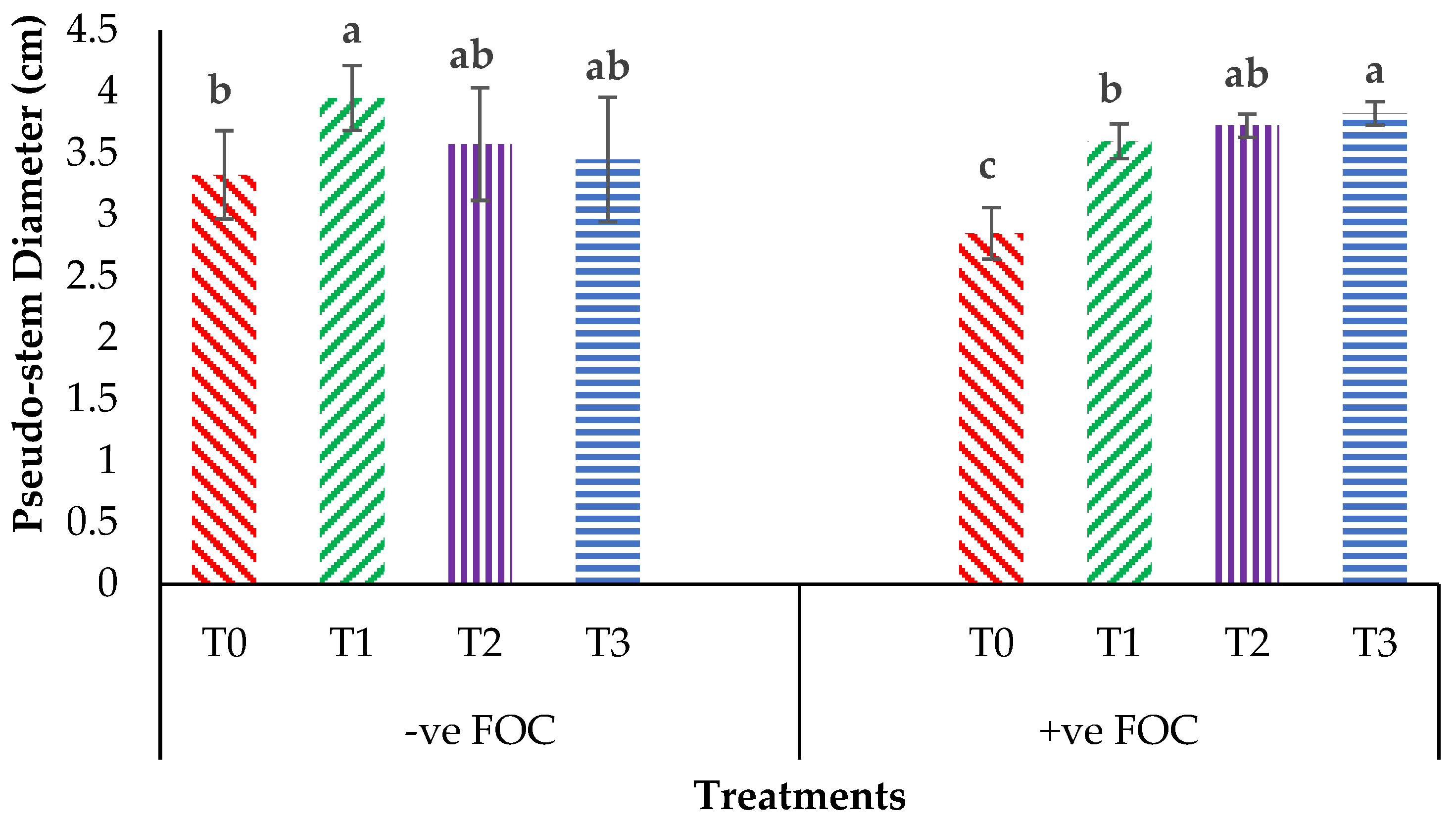
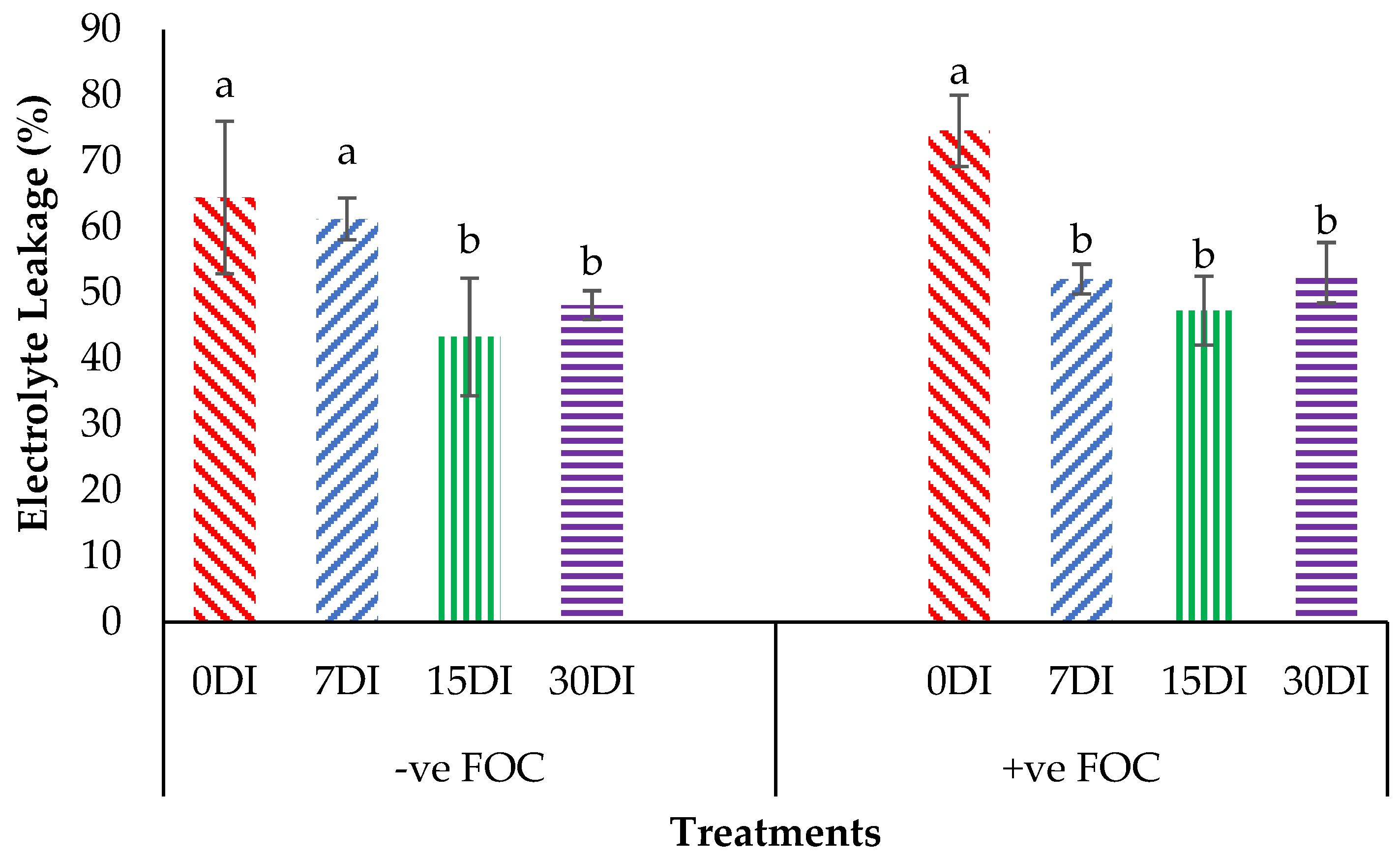
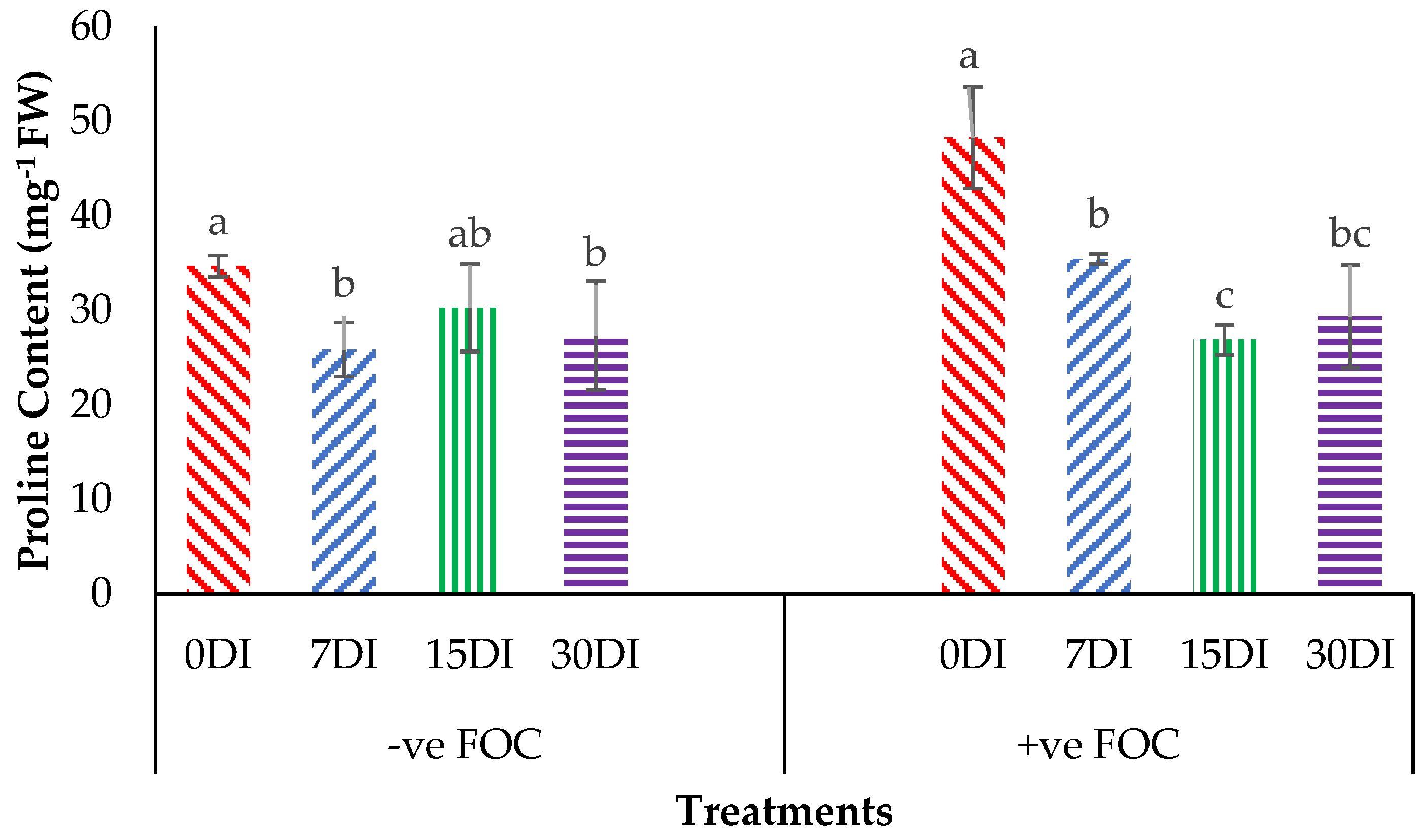
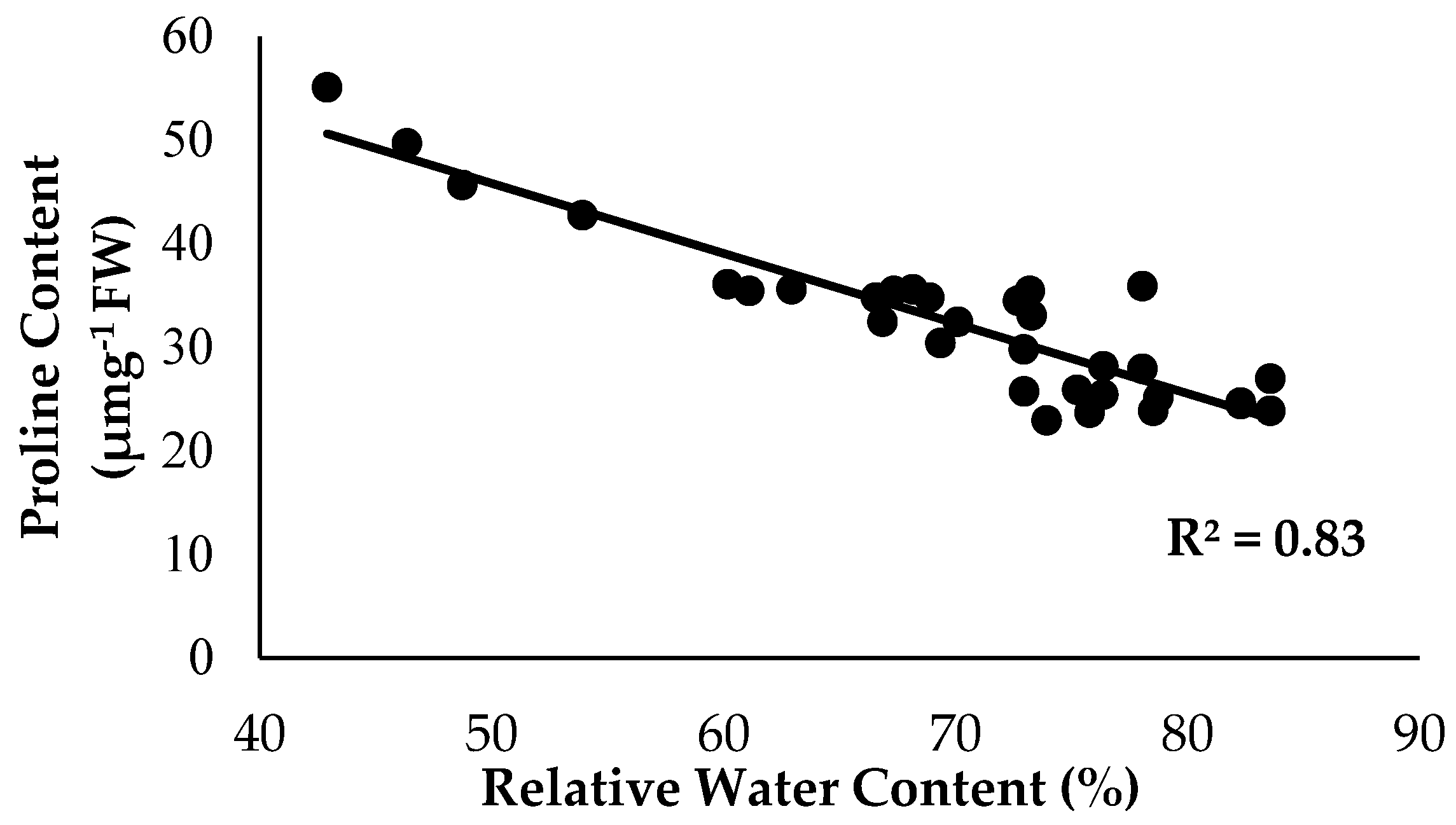
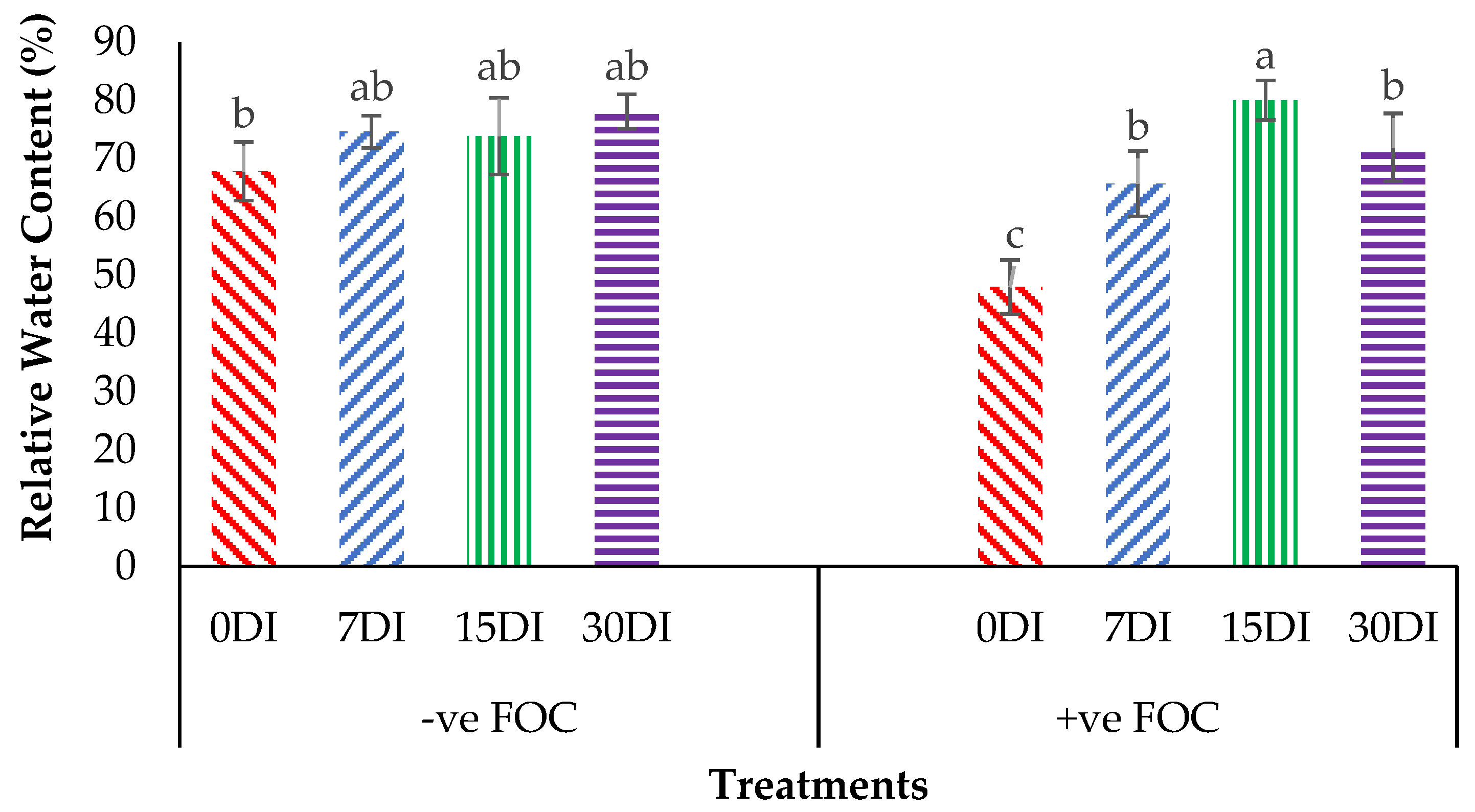
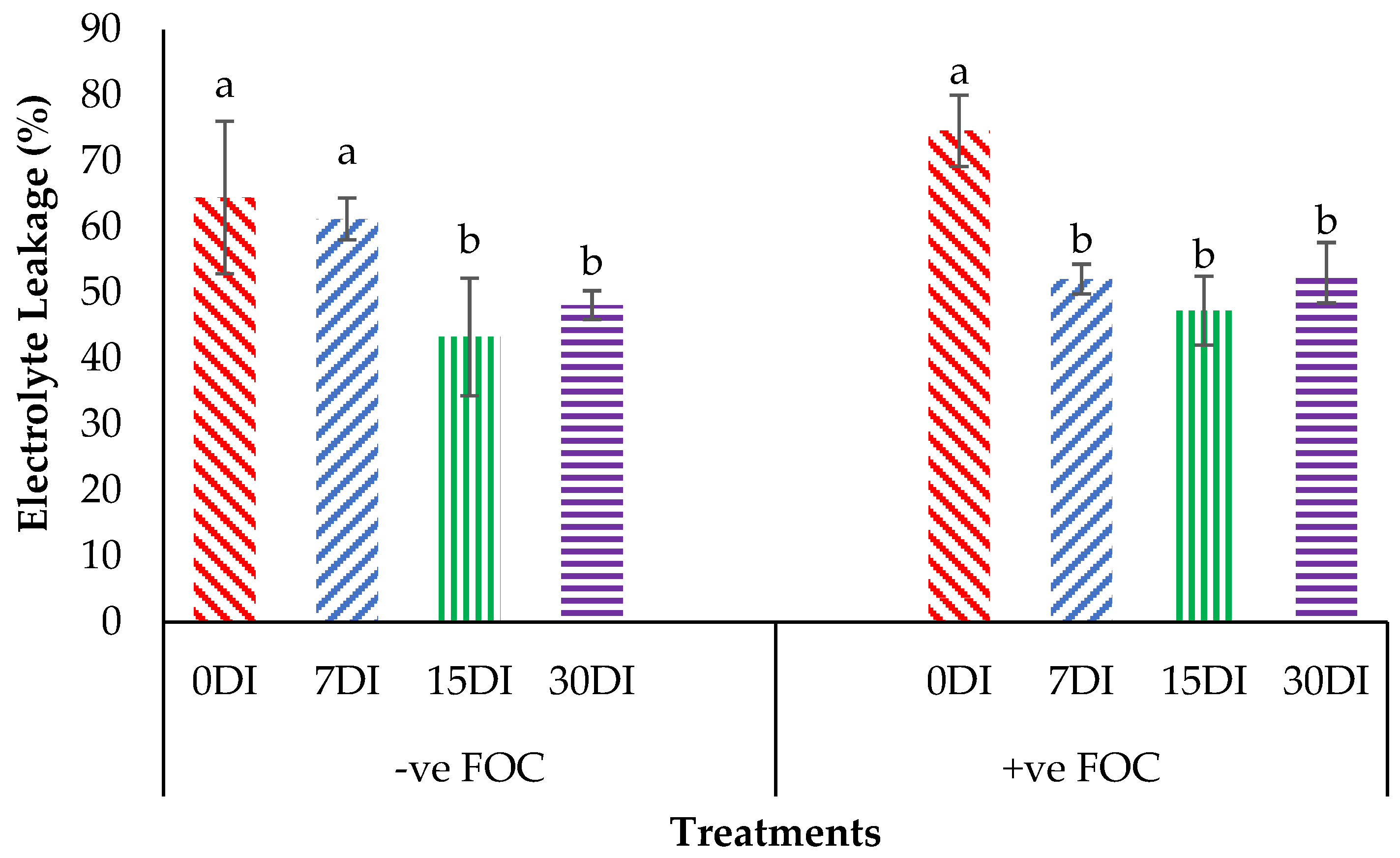
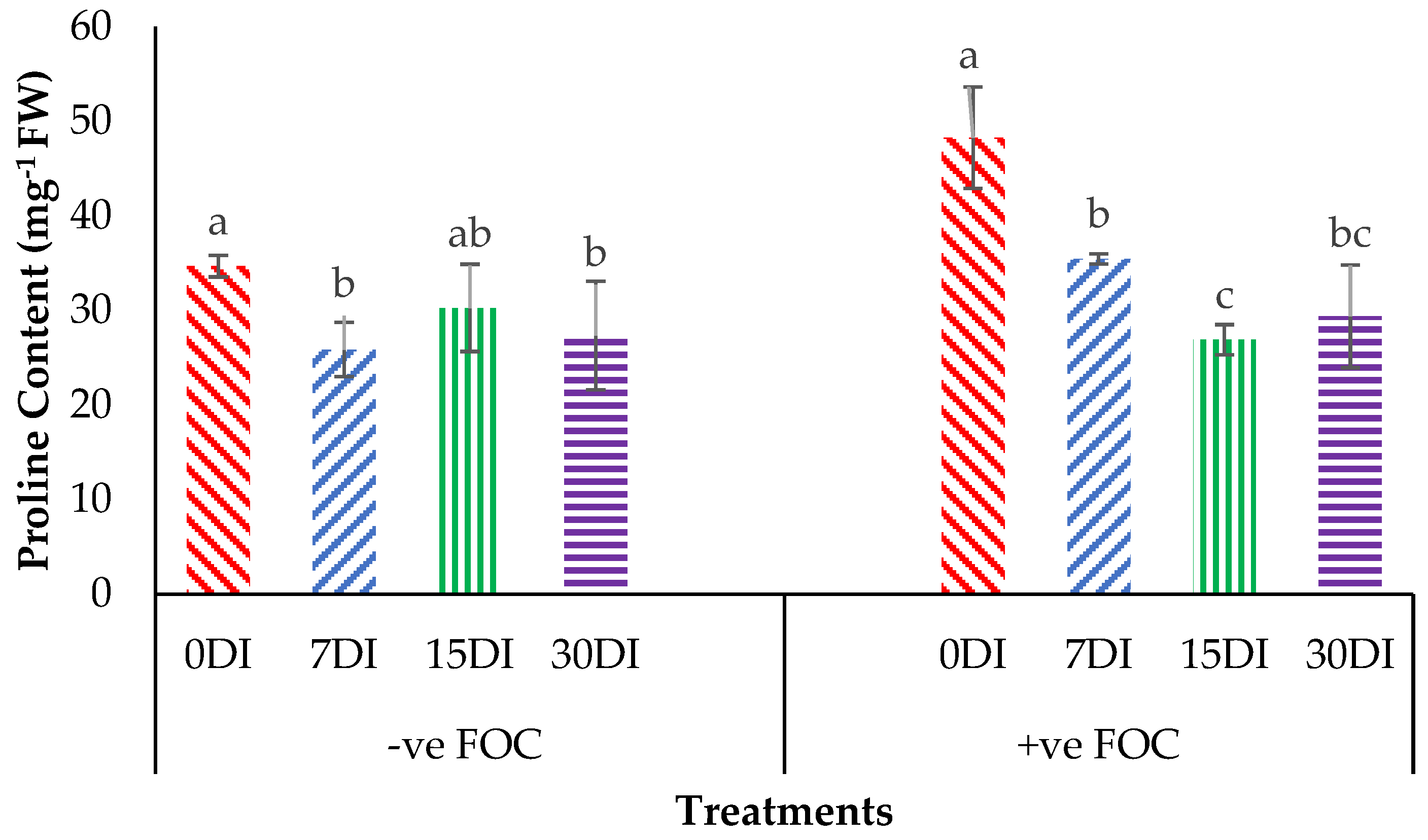
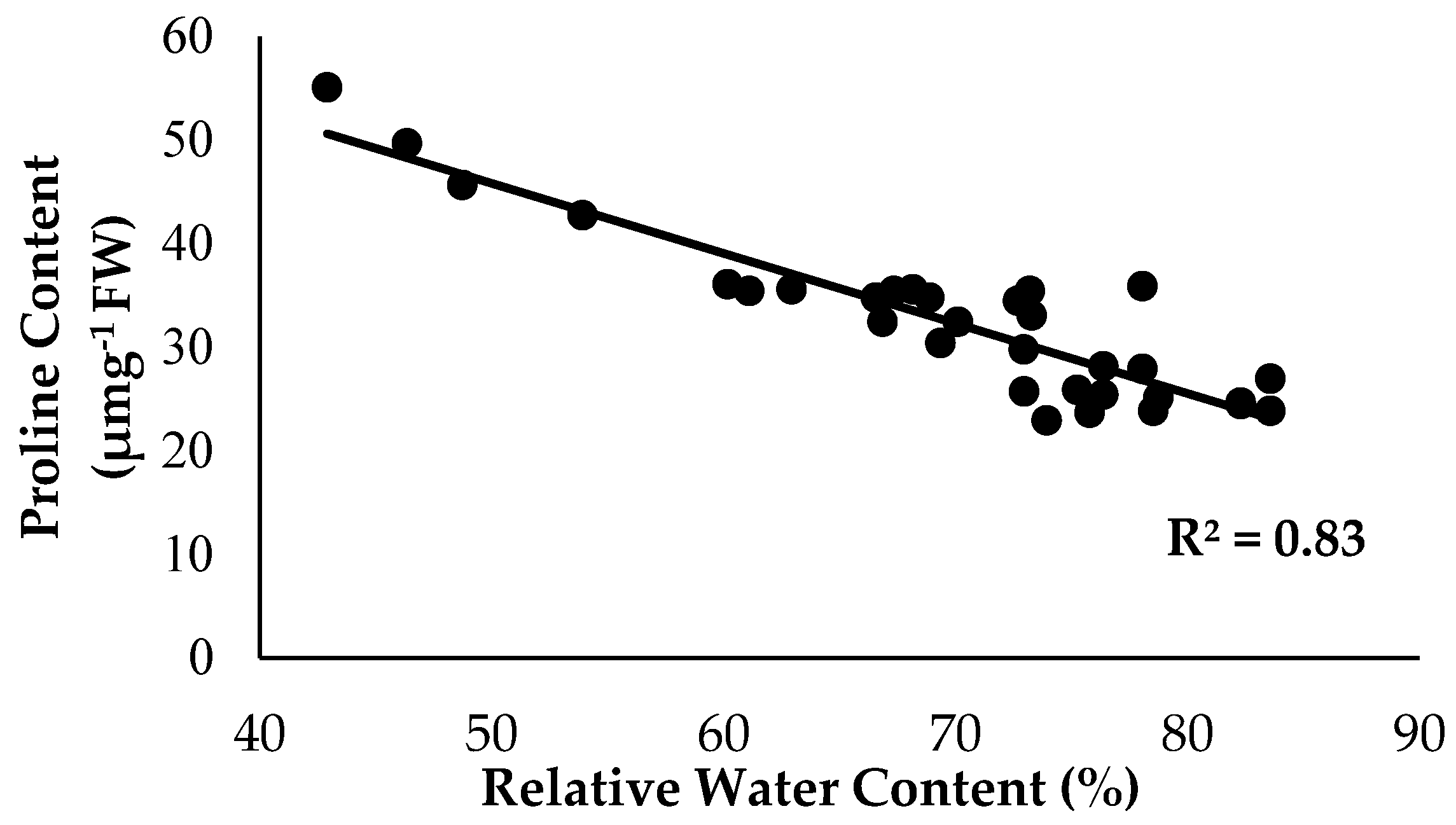
2.3. Physiological Attributes and Biochemical Content
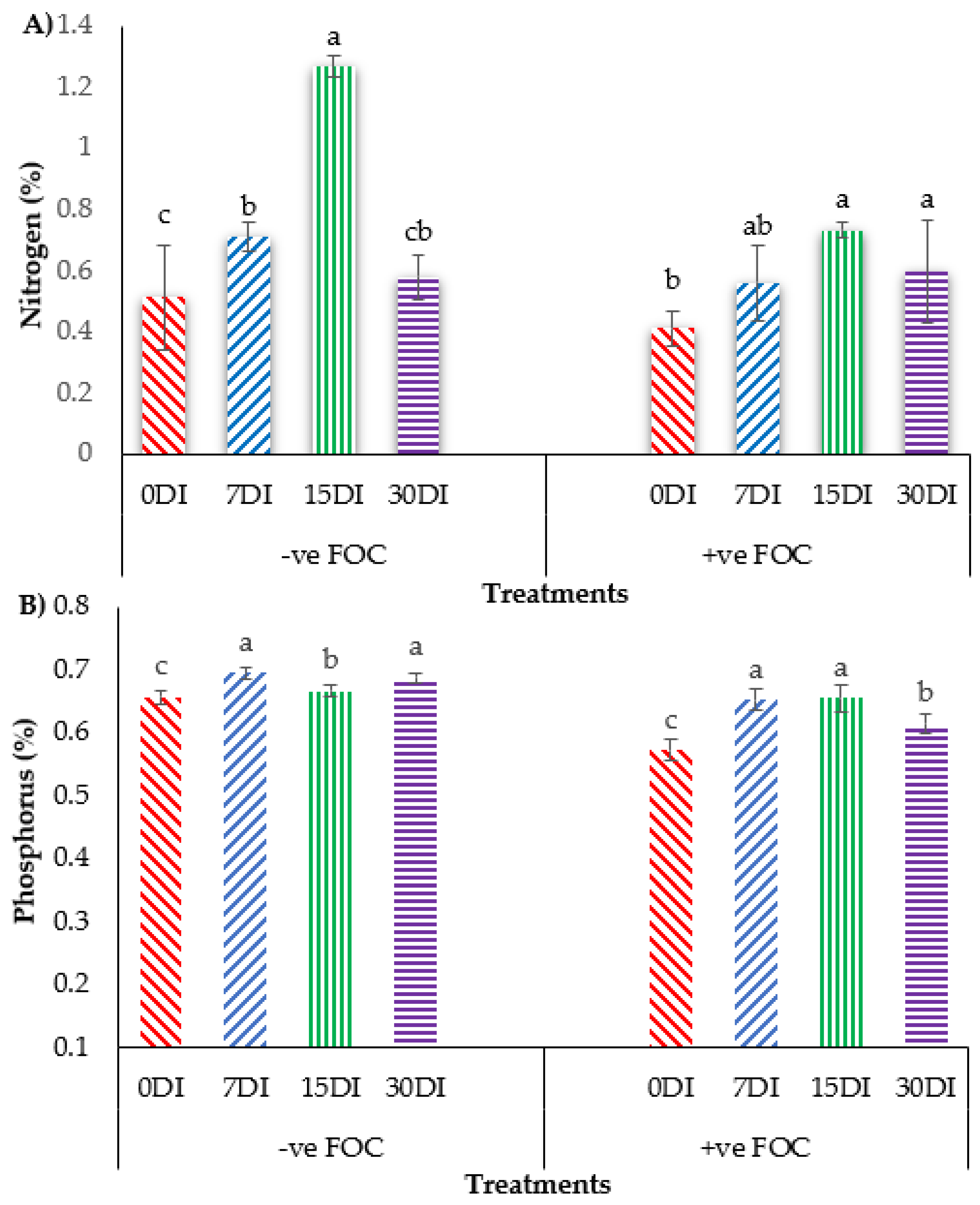
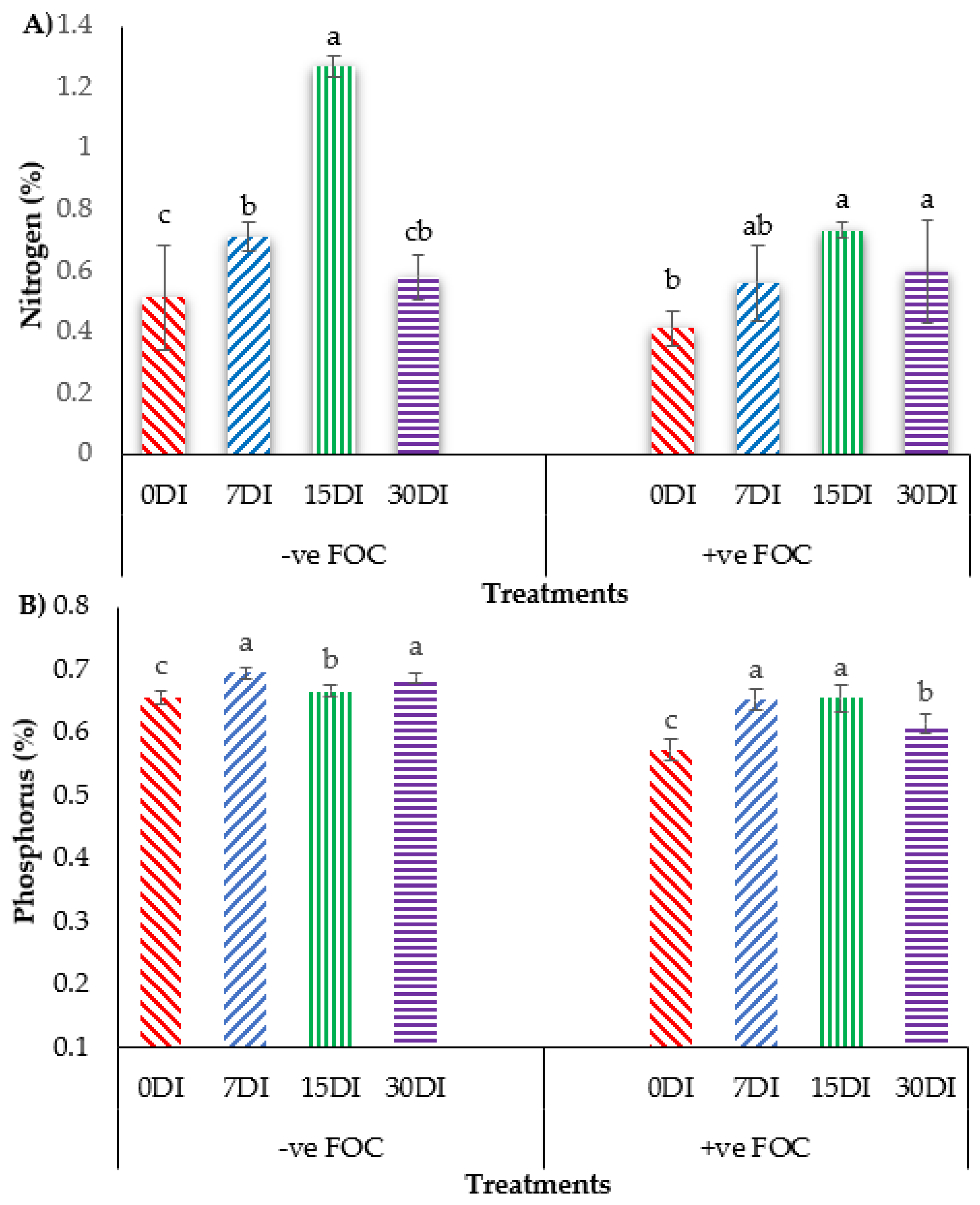
2.4. Plant Nutrient Uptake
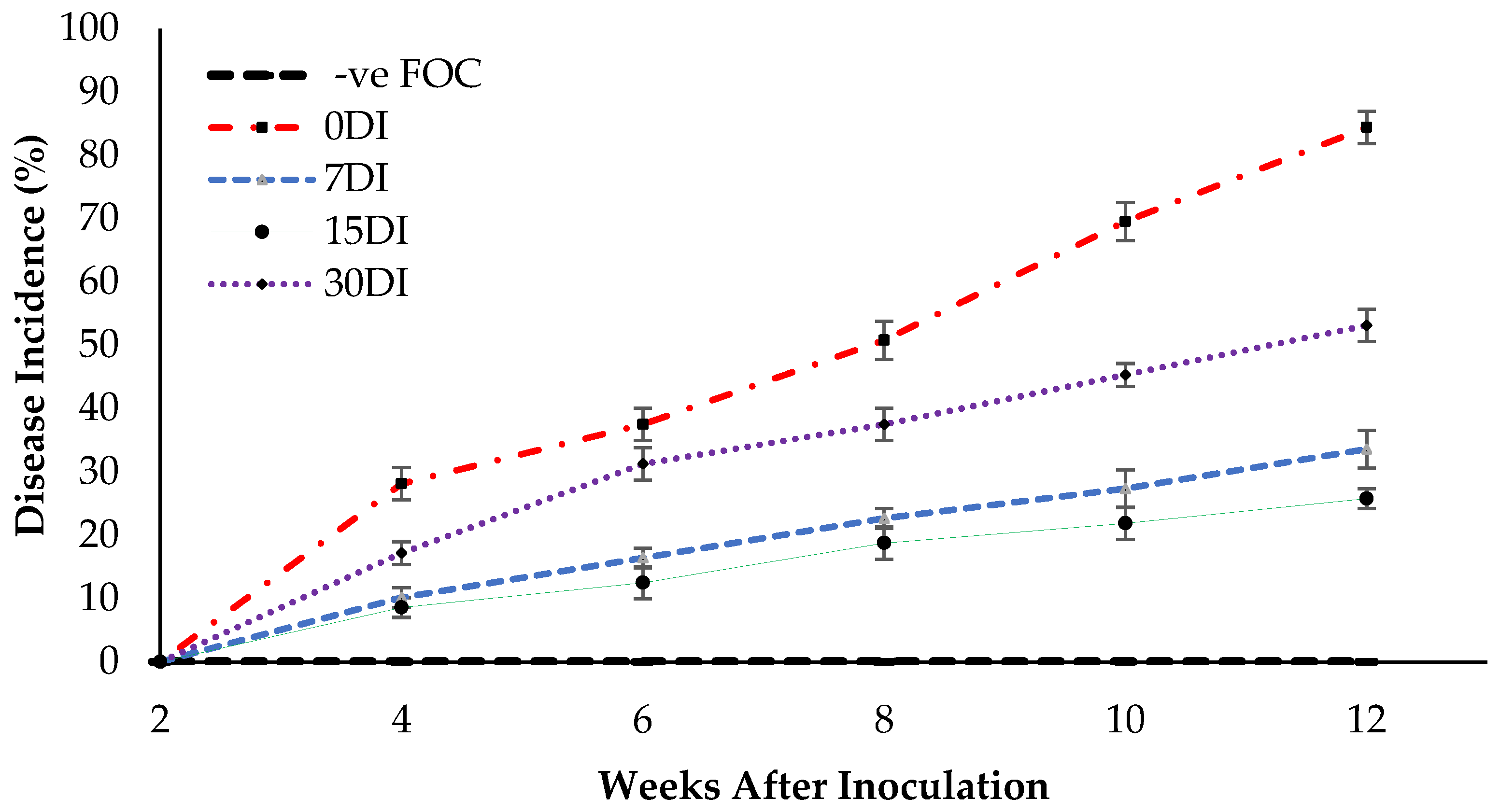
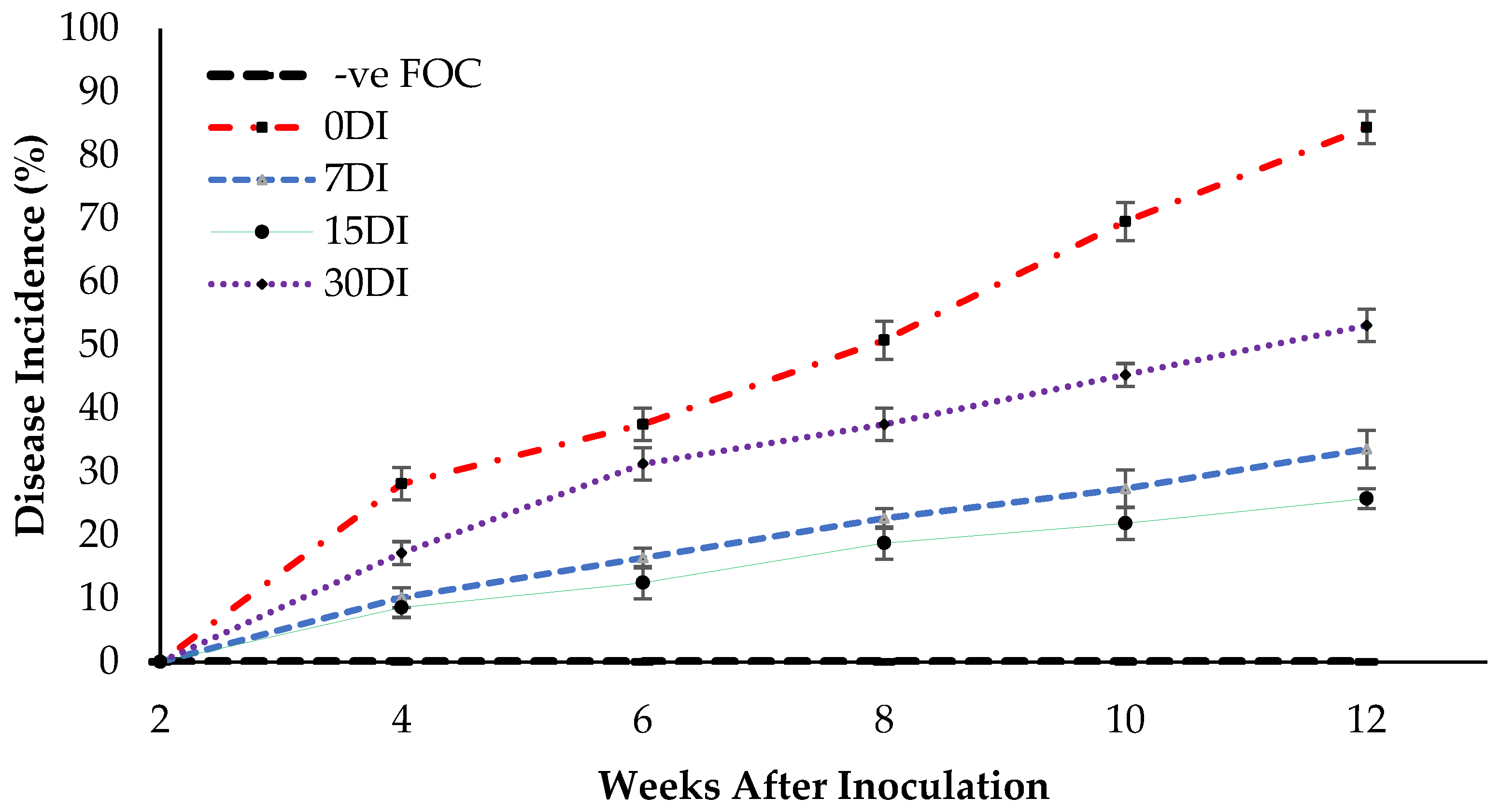
2.5. Disease Assessment
3. Discussion
4. Materials and Methods
4.1. Experimental Materials’ Preparation and Treatment
4.2. Data Collection
4.2.1. Determination of Crop Growth Traits
4.2.2. Determination of the Total Chlorophyll Content
4.2.3. Measurement of the Leaf Gas Exchange
4.2.4. Biochemical Assay of the Physiological Attributes
4.2.5. Determination of Plant Nutrient Uptake
4.2.6. Disease Assessment
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blomme, G.; Ocimati, W.; Zum Felde, A.; Amwonya, D.; Kantungeko, D. A literature review on yield gaps of various root, tuber and banana crops as a background for assessing banana yield reductions due to pests and diseases at a field site in western Burundi. Afr. J. Agric. Res. 2020, 8, 1169–1183. [Google Scholar]
- Maryani, N.; Lombard, L.; Poerba, Y.S.; Subandiyah, S.; Crous, P.W.; Kema, G.H.J. Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Stud. Mycol. 2019, 92, 155–194. [Google Scholar] [CrossRef] [PubMed]
- Sudantha, I.M.; Suwardji, S. Trichoderma biofungicides formulations on shallot growth, yield and Fusarium wilt disease resistance. IOP Conf. Ser. Earth Environ. Sci. 2021, 824, 012032. [Google Scholar] [CrossRef]
- Thangavelu, R.; Saraswathi, M.S.; Uma, S.; Loganathan, M.; Backiyarani, S.; Durai, P.; Swennen, R. Identification of sources resistant to a virulent Fusarium wilt strain (VCG 0124) infecting Cavendish bananas. Sci. Rep. 2021, 11, 3183. [Google Scholar] [CrossRef] [PubMed]
- Proboningrum, A.; Widono, S. Effectivity and compatibility of Azotobacter and Bacillus for biological control agents of Fusarium wilt on banana seedlings. IOP Conf. Ser. Earth Environ. Sci. 2019, 250, 012003. [Google Scholar] [CrossRef]
- Dadrasnia, A.; Usman, M.M.; Omar, R.; Ismail, S.; Abdullah, R. Potential use of Bacillus genus to control of bananas diseases: Approaches toward high yield production and sustainable management. J. King Saud Univ. Sci. 2020, 4, 2336–2342. [Google Scholar] [CrossRef]
- Martin, G.; Cardi, C.; Sarah, G.; Ricci, S.; Jenny, C.; Fondi, E.; Yahiaoui, N. Genome ancestry mosaics reveal multiple and cryptic contributors to cultivated banana. Plant J. 2020, 5, 1008–1025. [Google Scholar] [CrossRef]
- Dong, H.; Fan, H.; Lei, Z.; Wu, C.; Zhou, D.; Li, H. Histological and gene expression analyses in banana reveals the pathogenic differences between races 1 and 4 of banana Fusarium wilt pathogen. Phytopathology 2019, 6, 1029–1042. [Google Scholar] [CrossRef]
- Clement, W.K.F.; Vadamalai, G.; Saidi, N.B.; Zulperi, D. Research progress, challenges, future perspectives on the management of Fusarium wilt of banana in malaysia: A review. Malays. J. Sci. 2019, 2, 47–66. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef]
- Biryol, S.; Araz, N.; Eski, A.; Aktürk, R.; Aksu, Y.; Çelik Göktürk, B.; Demir, I. Biodiversity and pathogenicity of entomopathogenic fungi associated with the lesser spruce sawfly, Pristiphora abietina. Entomol. Exp. Appl. 2021, 5, 414–423. [Google Scholar] [CrossRef]
- Lishma, N.P.; Cherian, A. Effect of various bio control agents and plant extracts on the growth of Fusarium oxysporum f. sp. cubense (Foc) causing Fusarium wilt disease in banana. J. Pharm. Innov. 2021, 12, 2454–2457. [Google Scholar]
- He, Y.; Zhou, X.; Li, J.; Li, H.; Li, Y.; Nie, Y. In vitro secretome analysis suggests differential pathogenic mechanisms between Fusarium oxysporum f. sp. cubense Race 1 and Race 4. Biomolecules 2021, 9, 1353. [Google Scholar] [CrossRef]
- El-Mehrat, H.; Abdelaziz, A.M.R.A.; Ibrahim, W.; Hamed, L. Effectiveness of compost and potassium silicate under low rate of mineral fertilizers on production, quality, marketability and NPK contents of banana plants. J. Agric. Eng. 2017, 12, 787–795. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B. Combining biocontrol agents with chemical fungicides for integrated plant fungal disease control. Microorganisms 2020, 12, 1930. [Google Scholar] [CrossRef]
- Sharma, M.; Chandra Dhakal, S.; Adhikari, R.K.; Tiwari, U. Profitability, productivity and resource use efficiency of banana production in Hetauda-Dumkibas road corridor, Nepal. Cogent Food Agric. 2021, 1, 1917134. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Importance of silicon in fruit nutrition: Agronomic and physiological implications. Fruit Crops Elsevier 2020, 9, 255–277. [Google Scholar]
- Gbongue, L.R.; Lalaymia, I.; Zeze, A.; Delvaux, B.; Declerck, S. Increased silicon acquisition in bananas colonized by Rhizophagus irregularis MUCL 41833 reduces the incidence of Pseudocercospora fijiensis. Front. Plant Sci. 2019, 9, 1977. [Google Scholar] [CrossRef]
- Vijayan, A.; Sriramachandrasekharan, M.V.; Manivannan, R.; Shakila, A. Effect of silicon through potassium silicate on yield, nutrient uptake and quality of Grand Naine Banana. Asian J. Agric. Sci. 2021, 3, 91–98. [Google Scholar] [CrossRef]
- Haroon, M.; Amruta, S.B.; Prakash, N.B.; Rangaswamy, K.T.; Lingaiah, H.B. Effect of silicon on incidence and severity of purple blotch disease (Alternaria porri (Ellis) Cif.) in onion (Allium cepa L.). Int J Curr. Microbiol App. Sci. 2020, 2, 429–439. [Google Scholar] [CrossRef]
- Christian, M.M.; Shimelis, H.; Laing, M.D.; Tsilo, T.J.; Mathew, I. Breeding for silicon-use efficiency, protein content and drought tolerance in bread wheat (Triticum aestivum L.): A review. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2021, 1, 17–29. [Google Scholar] [CrossRef]
- Yi, U.; Zaharah, S.S.; Ismail, S.I.; Musa, M.H. Effect of aqueous neem leaf extracts in controlling Fusarium wilt, soil physicochemical properties and growth performance of banana (Musa spp.). Sustainability 2021, 22, 12335. [Google Scholar] [CrossRef]
- Din, S.N.M.; Sakimin, S.Z.; Sijam, K.; Baghdadi, A.; Zakaria, M.A.T. Potential of Bacillus subtilis inoculated on biorichar amended soil for suppression of Fusarium wilt, biochemical changes and leaf gas exchange under water stress condition of banana (Musa acuminata) cv. Berangan. Fundam. Appl. Agric. 2018, 3, 515–524. [Google Scholar] [CrossRef]
- Damodaran, T.; Rajan, S.; Gopal, R.; Yadav, A.; Yadav, K.; Shukla, P.K.; Mishra, V.K. Successful community-based management of banana wilt caused by Fusarium oxysporum f. sp. cubense Tropical race-4 through ICAR-FUSICONT. J. Appl. Hortic. 2019, 21, 37–41. [Google Scholar] [CrossRef]
- Lin, P.; Zhang, M.; Wang, M.; Li, Y.; Liu, J.; Chen, Y. Inoculation with arbuscular mycorrhizal fungus modulates defense-related genes expression in banana seedlings susceptible to wilt disease. Plant Signal. Behav. 2021, 16, 1884782. [Google Scholar] [CrossRef]
- Hu, W.; Yang, B.; He, Z.; Li, G. Magnesium may be a key nutrient mechanism related to Fusarium wilt resistance: A new banana cultivar. PeerJ 2021, 9, 11141. [Google Scholar] [CrossRef]
- Takrim, M.A.; Zaharah, S.S.; Ismail, M.R.; Kasim, S.; Ahmad, K. Effectiveness of K2SiO3 on growth and physio-biochemical changes of banana seedlings grown under tropical climate as influenced by application frequency. IOP Conf. Ser. Earth Environ. Sci. 2021, 948, 012011. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; Osman, S.A.; Abd-El-Kareem, F. Protective effects of silicon and silicate salts against white rot disease of onion and garlic, caused by Stromatinia cepivora. J. Plant Pathol. 2021, 103, 27–43. [Google Scholar] [CrossRef]
- Costa, B.N.S.; Neto, A.R.; Chagas, P.C.; Chagas, E.A.; Pasqual, M.; Vendrame, W.A. Silicon in the anatomy and physiology of banana plant leaves under temporary immersion bioreactors. Agric. Environ. Sci. 2021, 7, 10. [Google Scholar]
- Adisa, I.O.; Reddy Pullagurala, V.L.; Rawat, S.; Hernandez-Viezcas, J.A.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.L. Role of cerium compounds in Fusarium wilt suppression and growth enhancement in tomato (Solanum lycopersicum). J. Agric. Food Chem. 2018, 66, 5959–5970. [Google Scholar] [CrossRef]
- Panigrahi, N.; Thompson, A.J.; Zubelzu, S.; Knox, J.W. Identifying opportunities to improve management of water stress in banana production. Sci. Hortic. 2021, 276, 109735. [Google Scholar] [CrossRef]
- Song, X.P.; Verma, K.K.; Tian, D.D.; Zhang, X.Q.; Liang, Y.J.; Huang, X.; Li, Y.R. Exploration of silicon functions to integrate with biotic stress tolerance and crop improvement. Biol. Res. 2021, 54, 19. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Zia-ur-Rehman, M.; Akhtar, T.; Zia, M.H.; Aslam, M. Silicon nutrition lowers cadmium content of wheat cultivars by regulating transpiration rate and activity of antioxidant enzymes. Environ. Pollut. 2018, 242, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.; Correia, B.; António, C.; Rodrigues, A.M.; Gómez-Cadenas, A.; Valledor, L.; Pinto, G. Pinus susceptibility to pitch canker triggers specific physiological responses in symptomatic plants: An integrated approach. Front. Plant Sci. 2019, 10, 509. [Google Scholar] [CrossRef]
- Liu, S.; Li, J.; Zhang, Y.; Liu, N.; Viljoen, A.; Mostert, D.; Li, C. Fusaric acid instigates the invasion of banana by Fusarium oxysporum f. sp. cubense TR 4. New Phytol. 2020, 225, 913–929. [Google Scholar] [CrossRef]
- Li, Y.; Lv, J.; Zhao, Q.; Chen, L.; Dong, Y.; Dong, K. Wheat/faba bean intercropping improves physiological and structural resistance of faba bean to Fusaric acid stress. Plant Pathol. 2021, 70, 827–840. [Google Scholar] [CrossRef]
- Xia, Y.; Farooq, M.A.; Javed, M.T.; Kamran, M.A.; Mukhtar, T.; Ali, J.; Chaudhary, H.J. Multi-stress tolerant PGPR Bacillus xiamenensis PM14 activating sugarcane (Saccharum officinarum L.) red rot disease resistance. Plant Physiol. Biochem. 2020, 151, 640–649. [Google Scholar]
- Bhardwaj, S.; Kapoor, D. Fascinating regulatory mechanism of silicon for alleviating drought stress in plants. Plant Physiol. Biochem. 2021, 166, 1044–1053. [Google Scholar] [CrossRef]
- Wong, C.K.F.; Zulperi, D.; Saidi, N.B.; Vadamalai, G. A Consortium of Pseudomonas aeruginosa and Trichoderma harzianum for improving growth and induced biochemical changes in Fusarium wilt infected bananas. Trop. Life Sci. Res. 2021, 32, 23. [Google Scholar] [CrossRef]
- Thakral, V.; Bhat, J.A.; Kumar, N.; Myaka, B.; Sudhakaran, S.; Patil, G.; Deshmukh, R. Role of silicon under contrasting biotic and abiotic stress conditions provides benefits for climate smart cropping. Environ. Exp. Bot. 2021, 189, 104545. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Choudhary, R. Effects of silicon and silicon-based nanoparticles on rhizosphere microbiome, plant stress and growth. Biology 2021, 10, 791. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Bélanger, R.R. The controversies of silicon’s role in plant biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Fahadi Hoveizeh, N.; Gholami, R.; Abdelrahman, M.; Tran, L.S.P. Exogenous melatonin mitigates salinity-induced damage in olive seedlings by modulating ion homeostasis, antioxidant defense, and phytohormone balance. Physiol. Plant. 2021, 173, 1682–1694. [Google Scholar] [CrossRef]
- Thangavelu, R.; Loganathan, M.; Arthee, R.; Prabakaran, M.; Uma, S. Fusarium wilt: A threat to banana cultivation and its management. Cab. Rev. 2020, 15, 004. [Google Scholar] [CrossRef]
- Campos, O.; Araya-Alman, M.; Acevedo-Opazo, C.; Olivares, B.O.; Rey, J.C.; Cañete-Salinas, P.; Kurina, F.G.; Balzarini, M.; Lobo, D.; Navas-Cortés, J.A. Relationship Between Soil Properties and Banana Productivity in the Two Main Cultivation Areas in Venezuela. J. Soil Sci. Plant Nutr. 2020, 20, 2512–2524. [Google Scholar]
- Olivares, B. Determination of the Potential Influence of Soil in the Differentiation of Productivity and in the Classification of Susceptible Areas to Banana Wilt in Venezuela. UCOPress: Córdoba, Spain, 2022; pp. 89–111. [Google Scholar]
- Campos, O.; Calero, J.; Rey, J.C.; Lobo, D.; Landa, B.B.; Gómez, J.A. Correlation of banana productivity levels and soil morphological properties using regularized optimal scaling regression. Catena 2022, 208, 105718. [Google Scholar]
- Olivares, B.; Rey, J.C.; Lobo, D.; Navas-Cortés, J.A.; Gómez, J.A.; Landa, B.B. Fusarium Wilt of Bananas: A Review of Agro-Environmental Factors in the Venezuelan Production System Affecting Its Development. Agronomy 2021, 11, 986. [Google Scholar] [CrossRef]
- Rey, J.C.; Martínez-Solórzano, G.; Ramírez, H.; Pargas-Pichardo, R. Cavendish Banana Wilt, and its relationship with agroecological conditions in a lacustrine plain of Venezuela. Agron. Trop. 2020, 70, 1–12. [Google Scholar]
- Paredes, F.; Rey, J.; Lobo, D.; Galvis-Causil, S.; Olivares, B. The relationship between the normalized difference vegetation index, rainfall, and potential evapotranspiration in a banana plantation of Venezuela. SAINS TANAH-J. Soil Sci. Agroclimatol. 2021, 18, 58–64. [Google Scholar]
- Ordonez, N.; García-Bastidas, F.; Laghari, H.B.; Akkary, M.Y.; Harfouche, E.N.; Al Awar, B.N.; Kema, G.H.J. First report of Fusarium oxysporum f. sp. cubense tropical race 4 causing Panama disease in Cavendish bananas in Pakistan and Lebanon. Plant Dis. 2016, 100, 209. [Google Scholar] [CrossRef]
- Huang, Y.H.; Wang, R.C.; Li, C.H.; Zuo, C.W.; Wei, Y.R.; Zhang, L.; Yi, G.J. Control of Fusarium wilt in banana with Chinese leek. Eur. J. Plant Pathol. 2012, 1, 87–95. [Google Scholar] [CrossRef]
- Zulkarnaini, Z.M.; Sakimin, S.Z.; Mohamed, M.T.M.; Jaafar, H.Z.E. Relationship between chlorophyll content and soil plant analytical development values in two cultivars of fig (Ficus carica L.) as brassinolide effect at an open field. IOP Conf. Ser. Earth Environ. Sci. 2019, 250, 012025. [Google Scholar] [CrossRef]
- De Camargo, M.S.; Bezerra, B.K.L.; Holanda, L.A.; Oliveira, A.L.; Vitti, A.C.; Silva, M.A. Silicon fertilization improves physiological responses in sugarcane cultivars grown under water deficit. J. Soil Sci. Plant Nutr. 2019, 19, 81–91. [Google Scholar] [CrossRef]
- Hoang, H.L.; de Guzman, C.C.; Cadiz, N.M.; Hoang, T.T.H.; Tran, D.H.; Rehman, H. Salicylic acid and calcium signaling induce physiological and phytochemical changes to improve salinity tolerance in Red Amaranth (Amaranthus tricolor L.). J. Soil Sci. Plant Nutr. 2020, 20, 1759–1769. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Root Dry Weight | Shoot Dry Weight | Root to Shoot Ratio |
|---|---|---|---|
| (g) | (g) | ||
| Main plot means: Treatments | |||
| −ve FOC | 3.29 ± 0.45 a | 19.831 ± 4.55 a | 0.172 ± 0.03 a |
| +ve FOC | 3.21 ± 0.58 a | 20.444 ± 6.25 a | 0.173 ± 0.05 a |
| LSD (p < 0.05) | NS | NS | NS |
| Sub-plot means: Si compounds | |||
| T0 | 2.53 ± 0.48 b | 12.08 ± 3.66 b | 0.226 ± 0.06 a |
| T1 | 3.37 ± 0.2 a | 23.13 ± 2.21 a | 0.146 ± 0.01 b |
| T2 | 3.6 ± 0.24 a | 23.2 ± 2.46 a | 0.156 ± 0.01 b |
| T3 | 3.5 ± 0.21 a | 22.12 ± 2.18 a | 0.159 ± 0.02 b |
| LSD (p < 0.05) | 0.27 *** | 2.84 *** | 0.04 ** |
| Significance interaction | NS | NS | NS |
| Factors | Stomata Conductance | Transpiration Rate |
|---|---|---|
| (mmol m−2s−1) | (mmol H2O m−2s−1) | |
| Main plot means: Treatments | ||
| −ve FOC | 0.74 ± 0.03 a | 3.86 ± 0.42 a |
| +ve FOC | 0.54 ± 0.26 b | 2.81 ±0.67 b |
| LSD (p < 0.05) | 0.11 * | 0.39 ** |
| Sub-plot means: Si compounds | ||
| T0 | 0.53 ±0.21 b | 3.03 ±0.85 b |
| T1 | 0.56 ±0.22 b | 3.05 ± 3.05 b |
| T2 | 0.74 ± 0.17 a | 3.64 ± 0.56 a |
| T3 | 0.74 ± 0.16 a | 3.61 ± 0.64 a |
| LSD (p < 0.05) | 0.17 * | 0.52 * |
| Significance interaction | NS | NS |
| Factors | K | Ca | Mg |
|---|---|---|---|
| (%) | (%) | (%) | |
| Main plot means: Treatments | |||
| -FOC | 1.74 ± 0.15 a | 0.39 ± 0.05 a | 0.53 ± 0.09 a |
| FOC | 1.65 ± 0.16 a | 0.38 ± 0.04 a | 0.52 ± 0.10 a |
| LSD (p < 0.05) | NS | NS | NS |
| Sub-plot means: Frequencies: day interval (DI) | |||
| 0DI | 1.60 ± 0.22 b | 0.38 ± 0.02 a | 0.46 ± 0.04 b |
| 7DI | 1.83 ± 0.16 a | 0.41 ± 0.03 a | 0.54 ± 0.13 ab |
| 15DI | 1.78 ± 0.15 a | 0.33 ± 0.01 b | 0.56 ± 0.11 a |
| 30DI | 1.56 ± 0.07 b | 0.40 ± 0.03 a | 0.52 + 0.05 ab |
| LSD (p < 0.05) | 0.10 *** | 0.03 *** | 0.10 * |
| Significance interaction | NS | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakaria, M.A.T.; Sakimin, S.Z.; Ismail, M.R.; Ahmad, K.; Kasim, S. Growth Enhancement and Resistance of Banana Plants to Fusarium Wilt Disease as Affected by Silicate Compounds and Application Frequency. Plants 2024, 13, 542. https://doi.org/10.3390/plants13040542
Zakaria MAT, Sakimin SZ, Ismail MR, Ahmad K, Kasim S. Growth Enhancement and Resistance of Banana Plants to Fusarium Wilt Disease as Affected by Silicate Compounds and Application Frequency. Plants. 2024; 13(4):542. https://doi.org/10.3390/plants13040542
Chicago/Turabian StyleZakaria, Md Aiman Takrim, Siti Zaharah Sakimin, Mohd Razi Ismail, Khairulmazmi Ahmad, and Susilawati Kasim. 2024. "Growth Enhancement and Resistance of Banana Plants to Fusarium Wilt Disease as Affected by Silicate Compounds and Application Frequency" Plants 13, no. 4: 542. https://doi.org/10.3390/plants13040542
APA StyleZakaria, M. A. T., Sakimin, S. Z., Ismail, M. R., Ahmad, K., & Kasim, S. (2024). Growth Enhancement and Resistance of Banana Plants to Fusarium Wilt Disease as Affected by Silicate Compounds and Application Frequency. Plants, 13(4), 542. https://doi.org/10.3390/plants13040542