
Defining Populations and Predicting Future Suitable Niche Space in the Geographically Disjunct, Narrowly Endemic Leafy Prairie-Clover (Dalea foliosa; Fabaceae)
 ,
,
Abstract
1. Introduction
2. Results
2.1. Summary Statistics
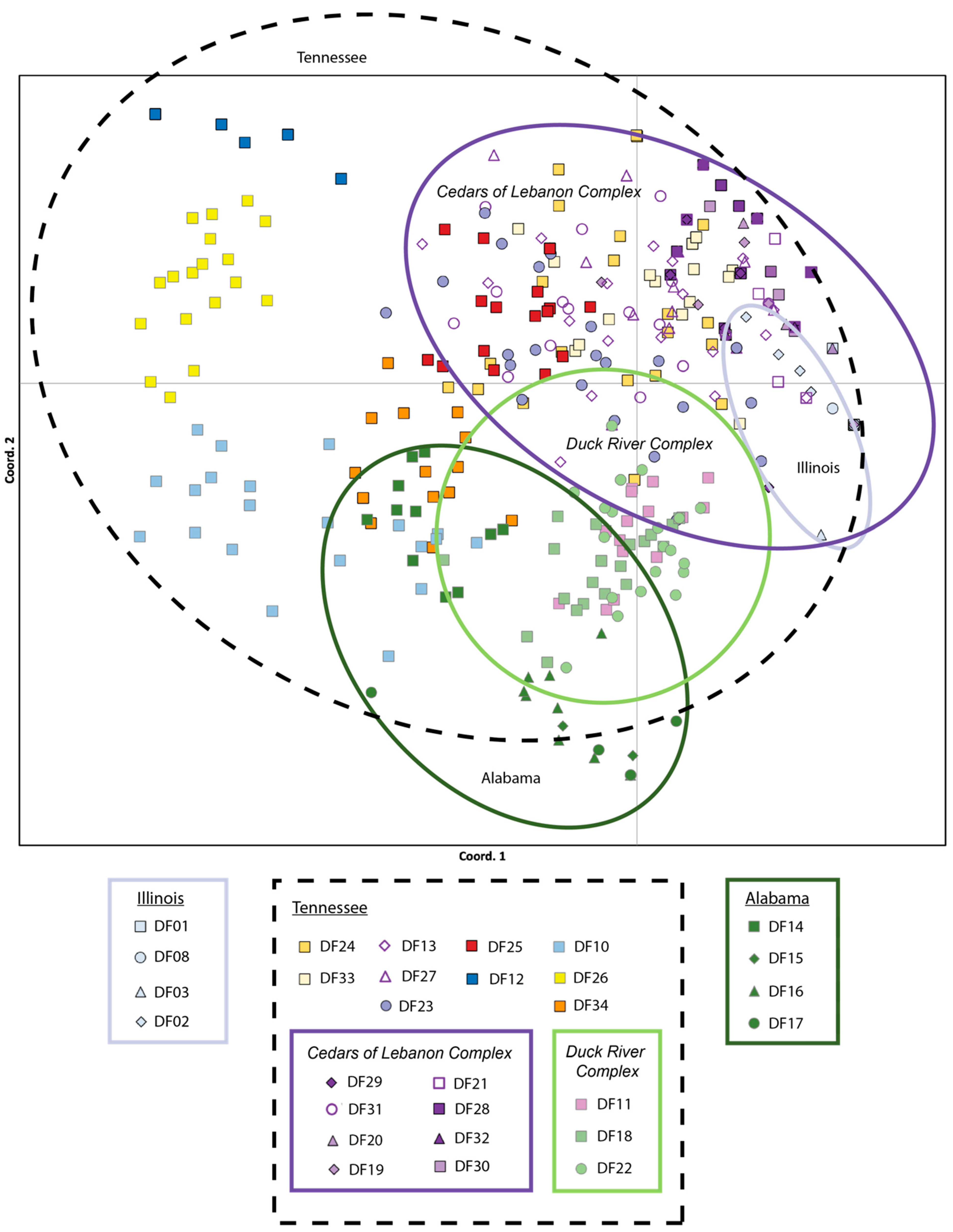
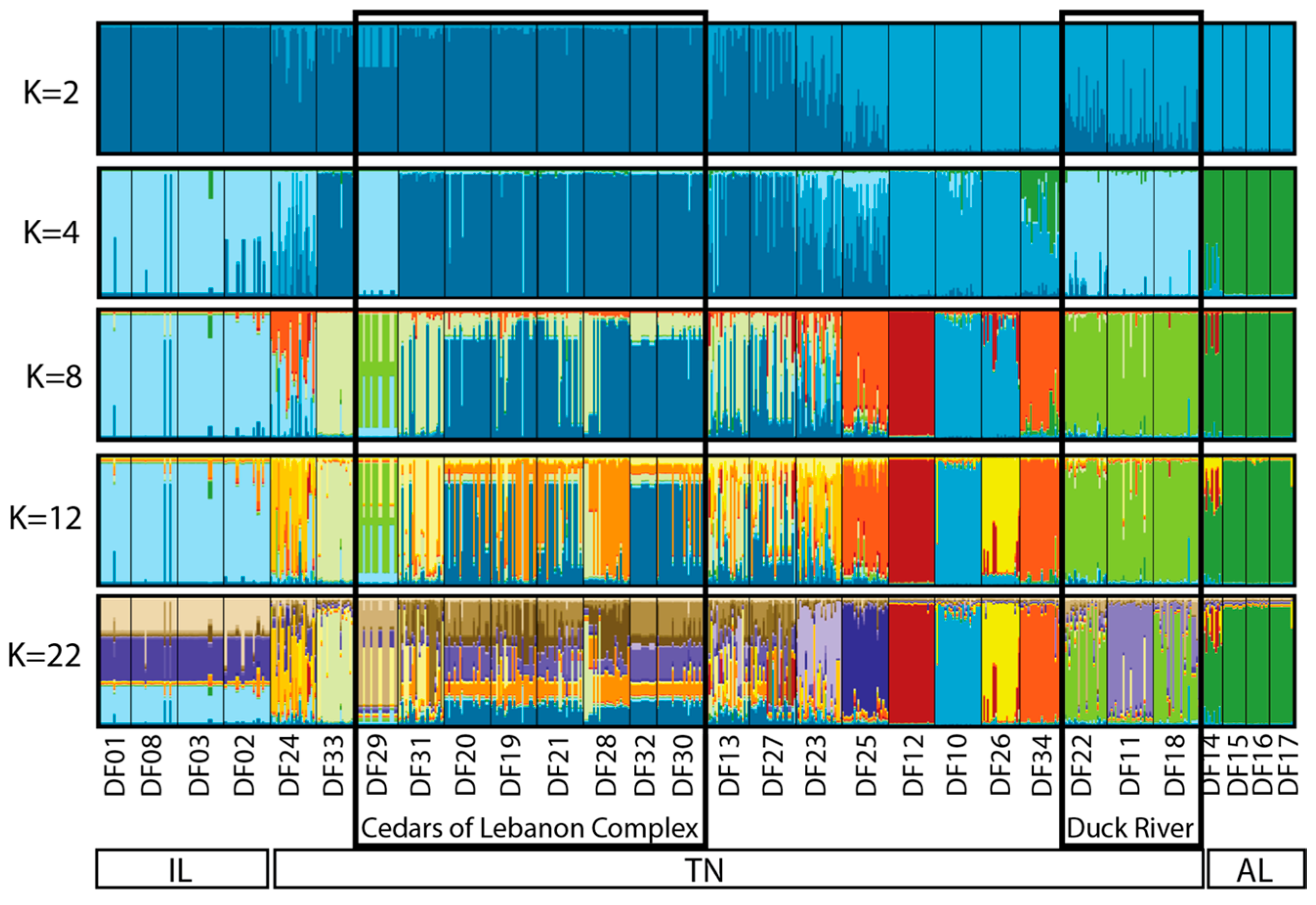
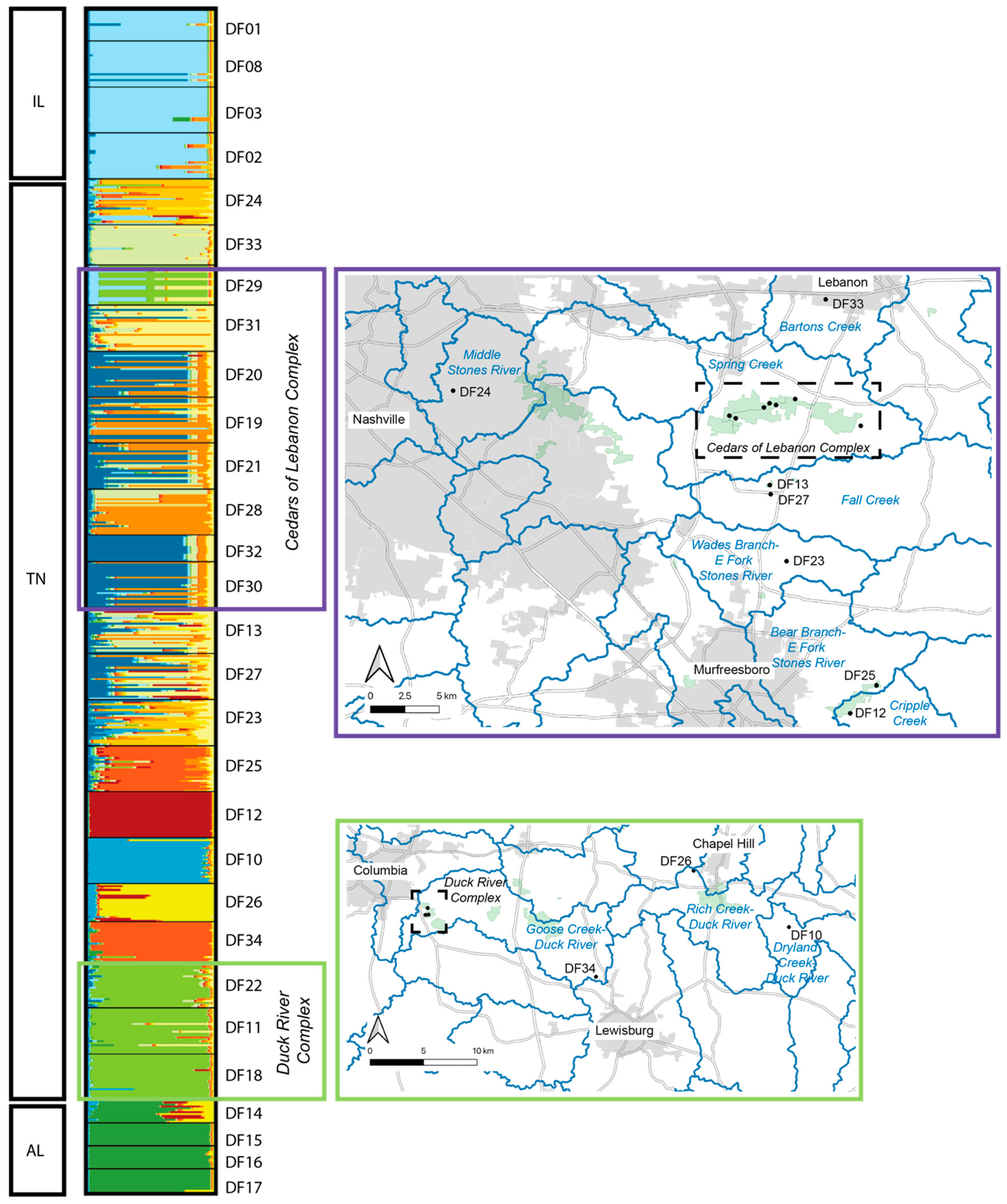
2.2. Individual and Population Assignment
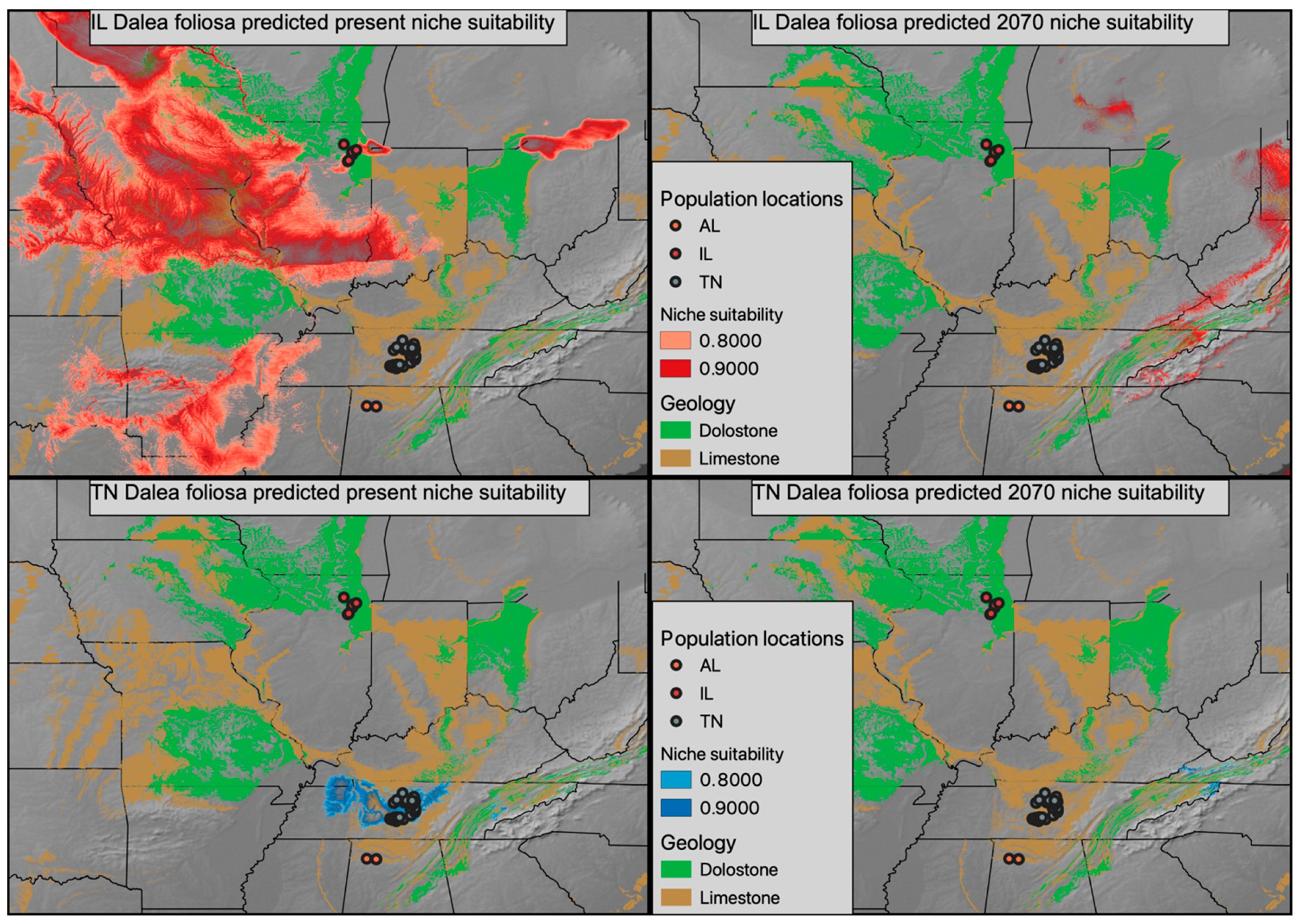
2.3. Current and Future Climatic Conditions
3. Discussion
3.1. Defining Populations in Dalea foliosa
3.2. Comparison with Other Calcareous Glade Endemics
3.3. Predicted Future Suitable Climatic Conditions for Dalea foliosa
4. Methods
4.1. Study Species
4.2. Field Sampling
4.3. Microsatellite Development and Genotyping
4.4. Summary Statistics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primer Sequences (5′-3′) | Repeat Motif | Allele Size (bp) |
|---|---|---|---|
| Dfol003 | F: GACATGGGTGGGTATGATTGAAG R: CGCGTGATGAGACCCTTATAAAG | (AG)8 | 260 |
| Dfol005 | F: ATGAAGGAAGATAATACCCGGCC R: CTTGCTGCTTTCGAATCATTCAC | (AG)13 | 164 |
| Dfol016 | F: CACACAAACAGGAAGAGAGATGG R: AACTAATGATTCCACCAGCCAAC | (AG)10 | 203 |
| Dfol020 1 | F: TCAGCGTCTTTGATCATCTGTTC R: TTTCAGGGTGTTTGACAAGGATC | (AT)15 | 255 |
| Dfol023 | F: ACCGATGATAGAAGAAAGCAAGG R: TGCTTTCATAGTCTTCAACGTCC | (AAC)7 | 194 |
| Dfol082 2 | F: CACACAAACAGGAAGAGAGATGG R: AACTAATGATTCCACCAGCCAAC | (AG)8 | 201 |
| Dfol092 | F: TTTCGCATCGTAACCTGAAGAAG R: GTCTCTGTGTCCTTCATTCCTTG | (AGG)7 | 213 |
| Dfol133 2 | F: CACACCGTGGAATCTTACTGTG R: ACCCTCCTTTCCACAAACAATAAG | (AC)8 | 190 |
| Dfol171 2 | F: TTCTTCACCTGCGTTGATTATGG R: ATCCAGCAAAGTCTATGAAGCTG | (AG)9 | 251 |
| Dfol241 | F: TGTGACACAAGTTGAACAAGATC R: AGAAATCGCTGTTTCCTTCCAAC | (AAG)6 | 173 |
4.5. Individual and Population Assignment
4.6. Current and Future Climatic Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caro, T.; Rowe, Z.; Berger, J.; Wholey, P.; Dobson, A. An Inconvenient Misconception: Climate Change Is Not the Principal Driver of Biodiversity Loss. Conserv. Lett. 2022, 15, e12868. [Google Scholar] [CrossRef]
- Isbell, F.; Balvanera, P.; Mori, A.S.; He, J.S. Expert Perspectives on Global Biodiversity Loss and Its Drivers and Impacts on People. Front. Ecol. Environ. 2023, 21, 94–103. [Google Scholar] [CrossRef]
- Gerber, L.R. Conservation Triage or Injurious Neglect in Endangered Species Recovery. Proc. Natl. Acad. Sci. USA 2016, 113, 3563–3566. [Google Scholar] [CrossRef] [PubMed]
- Buxton, R.T.; Avery-Gomm, S.; Lin, H.-Y.; Smith, P.A.; Cooke, S.J.; Bennett, J.R. Half of Resources in Threatened Species Conservation Plans Are Allocated to Research and Monitoring. Nat. Commun. 2020, 11, 4668. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.G.; Kehoe, L.; Mantyka-Pringle, C.; Chades, I.; Wilson, S.; Bloom, R.G.; Davis, S.K.; Fisher, R.; Keith, J.; Mehl, K.; et al. Prioritizing Recovery Funding to Maximize Conservation of Endangered Species. Conserv. Lett. 2018, 11, e12604. [Google Scholar] [CrossRef]
- Moritz, C. Defining “Evolutionarily Significant Units” for Conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Crandall, K.A.; Bininda-Emonds, O.R.; Mace, G.M.; Wayne, R.K. Considering Evolutionary Processes in Conservation Biology. Trends Ecol. Evol. 2000, 15, 290–295. [Google Scholar] [CrossRef]
- Kardos, M.; Armstrong, E.E.; Fitzpatrick, S.W.; Hauser, S.; Hedrick, P.W.; Miller, J.M.; Tallmon, D.A.; Funk, W.C. The Crucial Role of Genome-Wide Genetic Variation in Conservation. Proc. Natl. Acad. Sci. USA 2021, 118, e2104642118. [Google Scholar] [CrossRef]
- Reed, J.M.; Mills, L.S.; Dunning, J.B., Jr.; Menges, E.S.; McKelvey, K.S.; Frye, R.; Beissinger, S.R.; Anstett, M.-C.; Miller, P. Emerging Issues in Population Viability Analysis. Conserv. Biol. 2002, 16, 7–19. [Google Scholar] [CrossRef]
- Menges, E.S. The Application of Minimum Viable Population Theory to Plants. Genet. Conserv. Rare Plants 1991, 45, 158–164. [Google Scholar]
- Smith, D.R.; Allan, N.L.; McGowan, C.P.; Szymanski, J.A.; Oetker, S.R.; Bell, H.M. Development of a Species Status Assessment Process for Decisions under the U.S. Endangered Species Act. J. Fish Wildl. Manag. 2018, 9, 302–320. [Google Scholar] [CrossRef]
- Moritz, C. Conservation Units and Translocations: Strategies for Conserving Evolutionary Processes. Hereditas 2004, 130, 217–228. [Google Scholar] [CrossRef]
- McCarthy, M.A.; Possingham, H.P. Active Adaptive Management for Conservation. Conserv. Biol. 2007, 21, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Kadykalo, A.N.; Buxton, R.T.; Morrison, P.; Anderson, C.M.; Bickerton, H.; Francis, C.M.; Smith, A.C.; Fahrig, L. Bridging Research and Practice in Conservation. Conserv. Biol. 2021, 35, 1725–1737. [Google Scholar] [CrossRef]
- USFWS. USFWS Species Status Assessment Framework: An Integrated Analytical Framework for Conservation; Version 3.4; U.S. USFWS: Atlanta, GA, USA, 2016. [Google Scholar]
- NatureServe. Habitat-Based Plant Element Occurrence Delimitation Guidance; Version 1.0; NatureServe: Arlington, VA, USA, 2020. [Google Scholar]
- Van Rossum, F.; Le Pajolec, S.; Raspé, O.; Godé, C. Assessing Population Genetic Status for Designing Plant Translocations. Front. Conserv. Sci. 2022, 3, 829332. [Google Scholar] [CrossRef]
- Novak, B.J.; Phelan, R.; Weber, M.U.S. Conservation Translocations: Over a Century of Intended Consequences. Conserv. Sci. Pract. 2021, 3, e394. [Google Scholar] [CrossRef]
- Draper, D.; Marques, I.; Iriondo, J.M. Species Distribution Models with Field Validation, a Key Approach for Successful Selection of Receptor Sites in Conservation Translocations. Glob. Ecol. Conserv. 2019, 19, e00653. [Google Scholar] [CrossRef]
- Maschinski, J.; Falk, D.A.; Wright, S.J.; Possley, J.; Roncal, J.; Wendelberger, K.S. Optimal Locations for Plant Reintroductions in a Changing World. In Plant Reintroduction in a Changing Climate: Promises and Perils; Maschinski, J., Haskins, K.E., Raven, P.H., Eds.; Island Press/Center for Resource Economics: Washington, DC, USA, 2012; pp. 109–129. ISBN 9781610911832. [Google Scholar]
- Parmesan, C.; Hanley, M.E. Plants and Climate Change: Complexities and Surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef]
- Anderson, J.; Song, B.-H. Plant Adaptation to Climate Change—Where Are We? J. Syst. Evol. 2020, 58, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Hällfors, M.H.; Liao, J.; Dzurisin, J.; Grundel, R.; Hyvärinen, M.; Towle, K.; Wu, G.C.; Hellmann, J.J. Addressing Potential Local Adaptation in Species Distribution Models: Implications for Conservation under Climate Change. Ecol. Appl. 2016, 26, 1154–1169. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W. Using Niche Models with Climate Projections to Inform Conservation Management Decisions. Biol. Conserv. 2012, 155, 149–156. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software STRUCTURE: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.B.; Visger, C.; Watkins, S.; Pollack, C. Extremely Low Levels of Genetic Variation and Predicted Shifts in Suitable Niche Space for a Geographically Disjunct, Federally Endangered Legume, Leafy Prairie-Clover (Dalea foliosa). Castanea 2023, 88, 91–110. [Google Scholar] [CrossRef]
- Edwards, A.L.; Wiltshire, B.; Nickrent, D.L. Genetic Diversity in Astragalus tennesseensis and the Federal Endangered Dalea foliosa (Fabaceae). J. Torrey Bot. Soc. 2004, 131, 279–291. [Google Scholar] [CrossRef]
- McMahon, M.; Hufford, L. Phylogeny of Amorpheae (Fabaceae: Papilionoideae). Am. J. Bot. 2004, 91, 1219–1230. [Google Scholar] [CrossRef]
- Dixon, A.L.; Herlihy, C.R.; Busch, J.W. Demographic and Population-Genetic Tests Provide Mixed Support for the Abundant Centre Hypothesis in the Endemic Plant Leavenworthia stylosa. Mol. Ecol. 2013, 22, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Abeli, T.; Gentili, R.; Mondoni, A.; Orsenigo, S.; Rossi, G. Effects of Marginality on Plant Population Performance. J. Biogeogr. 2014, 41, 239–249. [Google Scholar] [CrossRef]
- Kennedy, J.P.; Preziosi, R.F.; Rowntree, J.K.; Feller, I.C. Is the Central-Marginal Hypothesis a General Rule? Evidence from Three Distributions of an Expanding Mangrove Species, Avicennia germinans (L.) L. Mol. Ecol. 2020, 29, 704–719. [Google Scholar] [CrossRef]
- Sporbert, M.; Keil, P.; Seidler, G.; Bruelheide, H.; Jandt, U.; Aćić, S.; Biurrun, I.; Campos, J.A.; Čarni, A.; Chytrý, M.; et al. Testing Macroecological Abundance Patterns: The Relationship between Local Abundance and Range Size, Range Position and Climatic Suitability among European Vascular Plants. J. Biogeogr. 2020, 47, 2210–2222. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Germination Ecophysiology of Herbaceous Plant Species in a Temperate Region. Am. J. Bot. 1988, 75, 286–305. [Google Scholar] [CrossRef]
- Baskin, J.M. Greenhouse and Laboratory Studies on the Ecological Life Cycle of Dalea foliosa (Fabaceae), a Federal Endangered Species. Nat. Areas J. 1998, 18, 54–62. [Google Scholar]
- US Fish and Wildlife Service. Recovery Plan for the Leafy Prairie-Clover (Dalea foliosa); US Fish and Wildlife Service: Atlanta, GA, USA, 1996; pp. 1–74. [Google Scholar]
- Thompson, J.N.; Walck, J.L.; Hidayati, S.N. Microhabitat Requirements of the Federally Endangered Dalea foliosa, with Recommendations on Establishment of New Populations. Castanea 2006, 71, 94–104. [Google Scholar] [CrossRef]
- Molano-Flores, B.; Zaya, D.N.; Baty, J.; Spyreas, G. An Assessment of the Vulnerability of Illinois’ Rarest Plant Species to Climate Change. Castanea 2019, 84, 115–127. [Google Scholar] [CrossRef]
- Molano-Flores, B.; Bell, T.J. Projected Population Dynamics for a Federally Endangered Plant under Different Climate Change Emission Scenarios. Biol. Conserv. 2012, 145, 130–138. [Google Scholar] [CrossRef]
- USFWS. Recovery Plan for Dalea foliosa (Leafy Prairie-Clover) (Gray) Barneby; U.S. Fish and Wildlife Service: Atlanta, GA, USA, 1996. [Google Scholar]
- USFWS. Leafy Prairie-Clover (Dalea foliosa) 5-Year Review: Summary and Evaluation; U.S. Fish and Wildlife Service, Tennessee Ecological Services Field Office: Cookeville, TN, USA, 2022. [Google Scholar]
- Morris, A.B.; Scalf, C.; Burleyson, A.; Johnson, L.T.; Trostel, K. Development and Characterization of Microsatellite Primers in the Federally Endangered Astragalus bibullatus (Fabaceae). Appl. Plant Sci. 2016, 4, 1500126. [Google Scholar] [CrossRef]
- Schuelke, M. An Economic Method for the Fluorescent Labeling of PCR Fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. Genalex 6: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E.; Huff, D.R. Evolutionary Implications of Allozyme and RAPD Variation in Diploid Populations of Dioecious Buffalograss Buchloë Dactyloides. Mol. Ecol. 1995, 4, 135–148. [Google Scholar] [CrossRef]
- Smouse, P.E.; Peakall, R. Spatial Autocorrelation Analysis of Individual Multiallele and Multilocus Genetic Structure. Heredity 1999, 82 Pt 5, 561–573. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring Weak Population Structure with the Assistance of Sample Group Information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
- Besnier, F.; Glover, K.A. ParallelStructure: A R Package to Distribute Parallel Runs of the Population Genetics Program STRUCTURE on Multi-Core Computers. PLoS ONE 2013, 8, e70651. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A Program for Identifying Clustering Modes and Packaging Population Structure Inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- Funk, S.M.; Guedaoura, S.; Juras, R.; Raziq, A.; Landolsi, F.; Luís, C.; Martínez, A.M.; Musa Mayaki, A.; Mujica, F.; do Mar Oom, M.; et al. Major Inconsistencies of Inferred Population Genetic Structure Estimated in a Large Set of Domestic Horse Breeds Using Microsatellites. Ecol. Evol. 2020, 10, 4261–4279. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A Cluster Matching and Permutation Program for Dealing with Label Switching and Multimodality in Analysis of Population Structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.A. Distruct: A Program for the Graphical Display of Population Structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Paulsen, J.; Körner, C. A Climate-Based Model to Predict Potential Treeline Position around the Globe. Alp. Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at High Resolution for the Earth’s Land Surface Areas. Sci Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.org (accessed on 19 December 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 19 December 2023).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A Maximum Entropy Approach to Species Distribution Modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; Association for Computing Machinery: New York, NY, USA, 2004; p. 83. [Google Scholar]
- Horton, J.D.; San Juan, C.A.; Stoeser, D.B. The State Geologic Map Compilation (SGMC) Geodatabase of the Conterminous United States; Data Series; Version 1.0: Originally Posted on 30 June 2017; Version 1.1: August 2017; U.S. Geological Survey: Reston, VA, USA, 2017. [Google Scholar]
- Brook, B.W.; O’Grady, J.J.; Chapman, A.P.; Burgman, M.A.; Akcakaya, H.R.; Frankham, R. Predictive Accuracy of Population Viability Analysis in Conservation Biology. Nature 2000, 404, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Coulson, T.; Mace, G.M.; Hudson, E.; Possingham, H. The Use and Abuse of Population Viability Analysis. Trends Ecol. Evol. 2001, 16, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Akçakaya, H.R.; Sjögren-Gulve, P. Population Viability Analyses in Conservation Planning: An Overview. Ecol. Bull. 2000, 48, 9–21. [Google Scholar]

| Site Name | Site Code | Month Sampled | Collector(s) | |
|---|---|---|---|---|
| Dalea foliosa sites sampled in Illinois | ||||
| Keepataw | DF01 | July 2014 | Pollack | |
| Material Services | DF08 | Aug 2014 | Pollack | |
| Dellwood | DF03 | July 2014 | Pollack | |
| Midewin | DF02 | Aug 2014 | Pollack | |
| Dalea foliosa sites sampled in Tennessee | ||||
| Hamilton Creek | DF24 | Aug 2015 | Fleming | |
| Lebanon | DF33 | 2022 | Elam | |
| Cedar Forest Rd W |  | DF29 | June 2021 | Elam, Fox, Morris |
| Vesta | DF31 | June 2021 | Elam, Fox, Morris | |
| Rowland Barrens E | DF20 | Sept 2015 | Bishop, Fleming, Williams | |
| Rowland Barrens W | DF19 | Sept 2015 | Bishop, Fleming, Williams | |
| Richmond Shop Rd | DF21 | Sept 2015 | Fleming | |
| Cedar Forest Rd S | DF28 | June 2021 | Elam, Fox, Morris | |
| Cedars Powerline | DF32 | June 2021 | Elam, Fox, Morris | |
| Hidden Springs | DF30 | June 2021 | Elam, Fox, Morris | |
| Lane Farm | DF13 | Aug 2015 | Bishop, Williams | |
| HWY 452 (Lane Farm) | DF27 | Sept 2015 | ||
| Holly Grove Rd | DF23 | Sept 2015 | Bishop, Fleming | |
| Hall Farm | DF25 | Aug 2015 | Bishop, Crabtree | |
| Flat Rock | DF12 | 2015 | ||
| Burnt Hill Rd | DF10 | Sept 2015 | Bishop, Williams, Call | |
| TVA Powerline (Chapel Hill) | DF26 | Sept 2015 | Bishop, Fleming | |
| Berlin Glade | DF34 | Aug 2022 | Elam, Call, Cogburn | |
| Blue Springs |  | DF22 | Sept 2015 | Bishop, Fleming |
| Columbia Glade W | DF11 | Sept 2015 | Fleming | |
| Columbia Glade E | DF18 | Sept 2015 | Bishop, Williams | |
| Dalea foliosa sites sampled in Alabama | ||||
| Franklin Co., AL | DF14 | July 2015 | Barger | |
| Lawrence Co., AL TVA ROW | DF15 | July 2015 | Barger | |
| Lawrence Co., AL Paved roadside | DF16 | July 2015 | Barger | |
| Lawrence Co., AL Dirt roadside | DF17 | July 2015 | Barger | |
| Site Name | N | %P | Na | Ne | Ho | He | F | ||
|---|---|---|---|---|---|---|---|---|---|
| Illinois sampling locations | |||||||||
| DF01 | 16 | 11.11 | 1.111 | 1.007 | 0.007 | 0.007 | −0.032 | ||
| DF08 | 24 | 11.11 | 1.222 | 1.026 | 0.005 | 0.021 | 0.781 | ||
| DF03 | 24 | 11.11 | 1.111 | 1.020 | 0.000 | 0.017 | 1.000 | ||
| DF02 | 24 | 22.22 | 1.222 | 1.091 | 0.023 | 0.065 | 0.632 | ||
| Tennessee sampling locations | |||||||||
| DF24 | 23 | 66.67 | 2.444 | 1.708 | 0.219 | 0.330 | 0.400 | ||
| DF33 | 20 | 44.44 | 1.556 | 1.314 | 0.106 | 0.144 | 0.252 | ||
| DF29 | 21 | 11.11 | 1.111 | 1.077 | 0.000 | 0.045 | 1.000 |  | |
| DF31 | 21 | 44.44 | 1.444 | 1.283 | 0.101 | 0.158 | 0.285 | ||
| DF20 | 23 | 44.44 | 1.444 | 1.057 | 0.033 | 0.047 | 0.127 | ||
| DF19 | 24 | 55.56 | 1.556 | 1.234 | 0.046 | 0.138 | 0.744 | ||
| DF21 | 24 | 33.33 | 1.444 | 1.109 | 0.056 | 0.077 | 0.455 | ||
| DF28 | 23 | 22.22 | 1.333 | 1.269 | 0.108 | 0.119 | 0.080 | ||
| DF32 | 12 | 0.00 | 1.000 | 1.000 | 0.000 | 0.000 | |||
| DF30 | 23 | 33.33 | 1.333 | 1.052 | 0.037 | 0.040 | 0.019 | ||
| DF13 | 22 | 88.89 | 2.333 | 1.389 | 0.147 | 0.250 | 0.417 | ||
| DF27 | 23 | 66.67 | 2.000 | 1.316 | 0.044 | 0.201 | 0.764 | ||
| DF23 | 23 | 100.00 | 2.333 | 1.640 | 0.235 | 0.363 | 0.412 | ||
| DF25 | 23 | 44.44 | 1.667 | 1.229 | 0.130 | 0.132 | −0.021 | ||
| DF12 | 23 | 11.11 | 1.111 | 1.107 | 0.010 | 0.055 | 0.823 | ||
| DF10 | 21 | 55.56 | 1.667 | 1.396 | 0.119 | 0.214 | 0.359 | ||
| DF26 | 19 | 66.67 | 2.000 | 1.412 | 0.163 | 0.243 | 0.373 | ||
| DF34 | 21 | 44.44 | 1.444 | 1.310 | 0.069 | 0.175 | 0.580 | ||
| DF22 | 23 | 44.44 | 1.556 | 1.386 | 0.169 | 0.206 | 0.185 |  | |
| DF11 | 22 | 22.22 | 1.222 | 1.205 | 0.087 | 0.107 | 0.188 | ||
| DF18 | 23 | 55.56 | 1.778 | 1.344 | 0.110 | 0.182 | 0.424 | ||
| Alabama sampling locations | |||||||||
| DF14 | 11 | 33.33 | 1.667 | 1.546 | 0.148 | 0.206 | 0.286 | ||
| DF15 | 11 | 11.11 | 1.111 | 1.010 | 0.009 | 0.009 | −0.043 | ||
| DF16 | 11 | 44.44 | 1.556 | 1.184 | 0.058 | 0.114 | 0.267 | ||
| DF17 | 12 | 44.44 | 1.556 | 1.092 | 0.028 | 0.076 | 0.635 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morris, A.B.; Visger, C.J.; Fox, S.J.; Scalf, C.; Fleming, S.; Call, G. Defining Populations and Predicting Future Suitable Niche Space in the Geographically Disjunct, Narrowly Endemic Leafy Prairie-Clover (Dalea foliosa; Fabaceae). Plants 2024, 13, 495. https://doi.org/10.3390/plants13040495
Morris AB, Visger CJ, Fox SJ, Scalf C, Fleming S, Call G. Defining Populations and Predicting Future Suitable Niche Space in the Geographically Disjunct, Narrowly Endemic Leafy Prairie-Clover (Dalea foliosa; Fabaceae). Plants. 2024; 13(4):495. https://doi.org/10.3390/plants13040495
Chicago/Turabian StyleMorris, Ashley B., Clayton J. Visger, Skyler J. Fox, Cassandra Scalf, Sunny Fleming, and Geoff Call. 2024. "Defining Populations and Predicting Future Suitable Niche Space in the Geographically Disjunct, Narrowly Endemic Leafy Prairie-Clover (Dalea foliosa; Fabaceae)" Plants 13, no. 4: 495. https://doi.org/10.3390/plants13040495
APA StyleMorris, A. B., Visger, C. J., Fox, S. J., Scalf, C., Fleming, S., & Call, G. (2024). Defining Populations and Predicting Future Suitable Niche Space in the Geographically Disjunct, Narrowly Endemic Leafy Prairie-Clover (Dalea foliosa; Fabaceae). Plants, 13(4), 495. https://doi.org/10.3390/plants13040495