
Volatile Organic Compounds Emitted by Flowers: Ecological Roles, Production by Plants, Extraction, and Identification




Abstract
1. Introduction
2. Functions of Floral Volatile Organic Compounds
2.1. Role of VOCs in Attracting Pollinators
2.2. Role in the Defense of the Plant
2.3. Role in Communication with Other Plants
3. Formation and Accumulation Site
4. Spatial and Temporal Variation of VOCs
5. Extraction and Analysis of Floral VOCs
5.1. Extraction of VOCs
5.2. Analysis of VOCs
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and Distribution of Floral Scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Gershenzon, J. The Chemical Diversity of Floral Scent. Biol. Flor. Scent 2006, 27–52. [Google Scholar]
- Knudsen, J.T.; Gershenzon, J. The Chemical Diversity of Floral Scent. In Biology of Plant Volatiles; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 57–78. ISBN 978-0-429-45561-2. [Google Scholar]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of Plant Volatiles: Nature’s Diversity and Ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef]
- Raguso, R.A. Why Are Some Floral Nectars Scented? Ecology 2004, 85, 1486–1494. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral Volatiles: From Biosynthesis to Function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Ludwiczuk, A.; Skalicka-Woźniak, K.; Georgiev, M.I. Terpenoids. In Pharmacognosy: Fundamentals, Applications and Strategy; Badal, S., Delgoda, R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 233–266. ISBN 9780128020999. [Google Scholar]
- Loomis, W.D.; Croteau, R. Biochemistry of Terpenoids. In The Biochemistry of Plants: A Comprehensive Treatise; Stumpf, P.K., Ed.; Elsevier: Amsterdam, The Netherlands, 1980; pp. 363–418. ISBN 0126754047. [Google Scholar]
- Miziorko, H.M. Enzymes of the Mevalonate Pathway of Isoprenoid Biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and Biological Functions of Terpenoids in Plants. Adv. Biochem. Eng. Biotechnol. 2014, 123, 127–141. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid Pathway Engineering: An Emerging Approach towards Plant Defense. Pathogens 2020, 9, 1–25. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, Function and Metabolic Engineering of Plant Volatile Organic Compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Maffei, M.E. Sites of Synthesis, Biochemistry and Functional Role of Plant Volatiles. South Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef]
- Tholl, D.; Hossain, O.; Weinhold, A.; Röse, U.S.R.; Wei, Q. Trends and Applications in Plant Volatile Sampling and Analysis. Plant J. 2021, 106, 314–325. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Metabolic Engineering of Plant Volatiles. Curr. Opin. Biotechnol. 2008, 19, 181–189. [Google Scholar] [CrossRef]
- Levin, R.A.; McDade, L.A.; Raguso, R.A. The Systematic Utility of Floral and Vegetative Fragrance in Two Genera of Nyctaginaceae. Syst. Biol. 2003, 52, 334–351. [Google Scholar] [CrossRef]
- Raguso, R.A.; Pichersky, E. Floral Volatiles fromClarkia Breweri andC. Concinna (Onagraceae): Recent Evolution of Floral Scent and Moth Pollination. Pl. Syst. Evol. 1995, 194, 55–67. [Google Scholar] [CrossRef]
- Cseke, L.J.; Kaufman, P.B.; Kirakosyan, A. The Biology of Essential Oils in the Pollination of Flowers. Nat. Prod. Commun. 2007, 2, 1317–1336. [Google Scholar] [CrossRef]
- Dobson, H.E. Relationship between Floral Fragrance Composition and Type of Pollinator. In Biology of Floral Scent; CRC Press: Boca Raton, FL, USA, 2006; pp. 147–198. ISBN 0-429-12666-2. [Google Scholar]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant Volatiles: Recent Advances and Future Perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Aharoni, A.; Jongsma, M.A.; Bouwmeester, H.J. Volatile Science? Metabolic Engineering of Terpenoids in Plants. Trends Plant Sci. 2005, 10, 594–602. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A. Identification of Geraniol as the Active Component in the Nassanoff Pheromone of the Honey Bee. Nature 1962, 194, 704–706. [Google Scholar] [CrossRef]
- Emura, M.; Nohara, I.; Toyoda, T.; Kanisawa, T. The Volatile Constituents of the Coffee Flower (Coffea Arabica L.). Flavour. Fragr. J. 1997, 12, 9–13. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R.; Cárdenas-Vargas, S.; Saavedra-Barrera, R.; Durán, D.C. GC-MS Study of Compounds Isolated from Coffea Arabica Flowers by Different Extraction Techniques: Sample Preparation. J. Sep. Sci. 2013, 36, 2901–2914. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Lewis, W.J.; Paré, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-Infested Plants Selectively Attract Parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Campbell, D.R.; Sosenski, P.; Raguso, R.A. Phenotypic Plasticity of Floral Volatiles in Response to Increasing Drought Stress. Ann. Bot. 2019, 123, 601–610. [Google Scholar] [CrossRef]
- Schnee, C.; Köllner, T.G.; Held, M.; Turlings, T.C.J.; Gershenzon, J.; Degenhardt, J. The Products of a Single Maize Sesquiterpene Synthase Form a Volatile Defense Signal That Attracts Natural Enemies of Maize Herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar] [CrossRef]
- Mayer, C.J.; Vilcinskas, A.; Gross, J. Phytopathogen Lures Its Insect Vector by Altering Host Plant Odor. J. Chem. Ecol. 2008, 34, 1045–1049. [Google Scholar] [CrossRef]
- Huang, M.; Sanchez-Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The Major Volatile Organic Compound Emitted from Arabidopsis Thaliana Flowers, the Sesquiterpene (E)-β-Caryophyllene, Is a Defense against a Bacterial Pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef]
- Horiuchi, J.I.; Badri, D.V.; Kimball, B.A.; Negre, F.; Dudareva, N.; Paschke, M.W.; Vivanco, J.M. The Floral Volatile, Methyl Benzoate, from Snapdragon (Antirrhinum majus) Triggers Phytotoxic Effects in Arabidopsis thaliana. Planta 2007, 226, 1–10. [Google Scholar] [CrossRef]
- Zhu, J.; Park, K.-C. Methyl Salicylate, a Soybean Aphid-Induced Plant Volatile Attractive to the Predator Coccinella Septempunctata. J. Chem. Ecol. 2005, 31, 1733–1746. [Google Scholar] [CrossRef]
- Hammerbacher, A.; Coutinho, T.A.; Gershenzon, J. Roles of Plant Volatiles in Defence against Microbial Pathogens and Microbial Exploitation of Volatiles. Plant Cell Env. 2019, 42, 2827–2843. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Ozawa, R.; Maffei, M.E. Recent Advances in Plant Early Signaling in Response to Herbivory. Int. J. Mol. Sci. 2011, 12, 3723–3739. [Google Scholar] [CrossRef]
- Farag, M.A.; Fokar, M.; Abd, H.; Zhang, H.; Allen, R.D.; Paré, P.W. (Z)-3-Hexenol Induces Defense Genes and Downstream Metabolites in Maize. Planta 2005, 220, 900–909. [Google Scholar] [CrossRef]
- Ruther, J.; Kleier, S. Plant–Plant Signaling: Ethylene Synergizes Volatile Emission In Zea Mays Induced by Exposure to (Z)-3-Hexen-1-Ol. J. Chem. Ecol. 2005, 31, 2217–2222. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne Signals Prime Plants against Insect Herbivore Attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R. Tropical Flower Scent Study by SPME. In Evolution of Solid Phase Microextraction Technology; Pawliszyn, J., Ed.; The Royal Society of Chemistry: London, UK, 2023; pp. 567–583. ISBN 978-1-83916-680-8. [Google Scholar]
- Johnson, S.D.; Sivechurran, J.; Doarsamy, S.; Shuttleworth, A. Dung Mimicry: The Function of Volatile Emissions and Corolla Patterning in Fly-Pollinated Wurmbea Flowers. New Phytol. 2020, 228, 1662–1673. [Google Scholar] [CrossRef]
- Peakall, R.; Wong, D.C.J.; Bohman, B.; Flematti, G.R.; Pichersky, E. Floral Volatiles for Pollinator Attraction and Speciation in Sexually Deceptive Orchids. In Biology of Plant Volatiles; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 271–295. ISBN 978-0-429-45561-2. [Google Scholar]
- Slavković, F.; Bendahmane, A. Floral Phytochemistry: Impact of Volatile Organic Compounds and Nectar Secondary Metabolites on Pollinator Behavior and Health. Chem. Biodivers. 2023, 20, e202201139. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Biochemical and Molecular Genetic Aspects of Floral Scents. Plant Physiol. 2000, 122, 627–633. [Google Scholar] [CrossRef]
- Maoz, I.; Sun, P.; Haring, M.A.; Schuurink, R.C.; Dudareva, N. Emission and Perception of Plant Volatiles. In Biology of Plant Volatiles; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FK, USA, 2020; pp. 251–267. ISBN 978-0-429-45561-2. [Google Scholar]
- Knauer, A.C.; Bakhtiari, M.; Schiestl, F.P. Crab Spiders Impact Floral-Signal Evolution Indirectly through Removal of Florivores. Nat. Commun. 2018, 9, 1367. [Google Scholar] [CrossRef]
- Junker, R.R.; Gershenzon, J.; Unsicker, S.B. Floral Odor Bouquet Loses Its Ant Repellent Properties After Inhibition of Terpene Biosynthesis. J. Chem. Ecol. 2011, 37, 1323–1331. [Google Scholar] [CrossRef]
- Junker, R.R.; Tholl, D. Volatile Organic Compound Mediated Interactions at the Plant-Microbe Interface. J. Chem. Ecol. 2013, 39, 810–825. [Google Scholar] [CrossRef]
- Dötterl, S.; Wolfe, L.M.; Jürgens, A. Qualitative and Quantitative Analyses of Flower Scent in Silene Latifolia. Phytochemistry 2005, 66, 203–213. [Google Scholar] [CrossRef]
- Kessler, D.; Diezel, C.; Clark, D.G.; Colquhoun, T.A.; Baldwin, I.T. Petunia Flowers Solve the Defence/Apparency Dilemma of Pollinator Attraction by Deploying Complex Floral Blends. Ecol. Lett. 2013, 16, 299–306. [Google Scholar] [CrossRef]
- Junker, R.R.; Loewel, C.; Gross, R.; Dötterl, S.; Keller, A.; Blüthgen, N. Composition of Epiphytic Bacterial Communities Differs on Petals and Leaves. Plant Biol. 2011, 13, 918–924. [Google Scholar] [CrossRef]
- Paul, I.; Poddar Sarkar, M.; Bhadoria, P.B.S. Floral Secondary Metabolites in Context of Biotic and Abiotic Stress Factors. Chemoecology 2022, 32, 49–68. [Google Scholar] [CrossRef]
- Bouwmeester, H.; Schuurink, R.C.; Bleeker, P.M.; Schiestl, F. The Role of Volatiles in Plant Communication. Plant J. 2019, 100, 892–907. [Google Scholar] [CrossRef]
- Baldwin, I.T. Plant Volatiles. Curr. Biol. 2010, 20, 392–397. [Google Scholar] [CrossRef]
- Ton, J.; Ent, S.V.D.; Hulten, M.V.; Pozo, M.; van Oosten, V.; van Loon, L.C.; Mauch-Mani, B.; Turlings, T.C.J.; Pieterse, C.M.J. Priming as a Mechanism behind Induced Resistance against Pathogens, Insects and Abiotic Stress. IOBC/Wprs Bull. 2009, 44, 3–13. [Google Scholar]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Silva, R.F.; Rabeschini, G.B.P.; Peinado, G.L.R.; Cosmo, L.G.; Rezende, L.H.G.; Murayama, R.K.; Pareja, M. The Ecology of Plant Chemistry and Multi-Species Interactions in Diversified Agroecosystems. Front. Plant Sci. 2018, 9, 1713. [Google Scholar] [CrossRef]
- Ninkovic, V.; Markovic, D.; Rensing, M. Plant Volatiles as Cues and Signals in Plant Communication. Plant Cell Environ. 2021, 44, 1030–1043. [Google Scholar] [CrossRef]
- Ninkovic, V. Volatile Interaction Between Undamaged Plants: A Short Cut to Coexistence. In Plant Communication from an Ecological Perspective; Baluska, F., Ninkovic, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 75–86. ISBN 9783642121623. [Google Scholar]
- Fahn, A. Secretory Tissues in Vascular Plants. New Phytol. 1988, 108, 229–257. [Google Scholar] [CrossRef]
- Duke, S.O.; Canel, C.; Rimando, A.M.; Telle, M.R.; Duke, M.V.; Paul, R.N. Current and Potential Exploitation of Plant Glandular Trichome Productivity; Elsevier: Amsterdam, The Netherlands, 2000; Volume 31, ISBN 0120059312. [Google Scholar]
- Jerković, I.; Mastelić, J. Composition of Free and Glycosidically Bound Volatiles of Mentha aquatica L. Croat. Chem. Acta 2001, 74, 431–439. [Google Scholar]
- Liao, P.; Ray, S.; Boachon, B.; Lynch, J.H.; Deshpande, A.; McAdam, S.; Morgan, J.A.; Dudareva, N. Cuticle Thickness Affects Dynamics of Volatile Emission from Petunia Flowers. Nat. Chem. Biol. 2021, 17, 138–145. [Google Scholar] [CrossRef]
- Liao, P.; Maoz, I.; Shih, M.-L.; Lee, J.H.; Huang, X.-Q.; Morgan, J.A.; Dudareva, N. Emission of Floral Volatiles Is Facilitated by Cell-Wall Non-Specific Lipid Transfer Proteins. Nat. Commun. 2023, 14, 330. [Google Scholar] [CrossRef]
- Effmert, U.; Buss, D.; Rohrbeck, D.; Piechulla, B. Localization of the Synthesis and Emission of Scent Compounds within the Flower. In Biology of Floral Scent; Taylor and Francis: Abingdon, UK, 2006; Volume 14, pp. 105–124. [Google Scholar] [CrossRef]
- Kolosova, N.; Sherman, D.; Karlson, D.; Dudareva, N. Cellular and Subcellular Localization of S-Adenosyl-L-Methionine: Benzoic Acid Carboxyl Methyltransferase, the Enzyme Responsible for Biosynthesis of the Volatile Ester Methylbenzoate in Snapdragon Flowers. Plant Physiol. 2001, 126, 956–964. [Google Scholar] [CrossRef]
- Bergougnoux, V.; Caissard, J.C.; Jullien, F.; Magnard, J.L.; Scalliet, G.; Cock, J.M.; Hugueney, P.; Baudino, S. Both the Adaxial and Abaxial Epidermal Layers of the Rose Petal Emit Volatile Scent Compounds. Planta 2007, 226, 853–866. [Google Scholar] [CrossRef]
- Burdon, R.C.F.; Raguso, R.A.; Kessler, A.; Parachnowitsch, A.L. Spatiotemporal Floral Scent Variation of Penstemon Digitalis. J. Chem. Ecol. 2015, 41, 641–650. [Google Scholar] [CrossRef]
- Gonzalez, A.; Benfodda, Z.; Bénimélis, D.; Fontaine, J.-X.; Molinié, R.; Meffre, P. Extraction and Identification of Volatile Organic Compounds in Scentless Flowers of 14 Tillandsia Species Using HS-SPME/GC-MS. Metabolites 2022, 12, 628. [Google Scholar] [CrossRef]
- Jakobsen, H.B.; Olsen, C.E. Influence of Climatic Factors on Emission of Flower Volatiles in Situ. Planta 1994, 192, 365–371. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of Plant Volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Ayasse, M. Post-Pollination Emission of a Repellent Compound in a Sexually Deceptive Orchid: A New Mechanism for Maximising Reproductive Success? Oecologia 2001, 126, 531–534. [Google Scholar] [CrossRef]
- Ryalls, J.M.W.; Langford, B.; Mullinger, N.J.; Bromfield, L.M.; Nemitz, E.; Pfrang, C.; Girling, R.D. Anthropogenic Air Pollutants Reduce Insect-Mediated Pollination Services. Environ. Pollut. 2022, 297, 118847. [Google Scholar] [CrossRef]
- Burkle, L.A.; Runyon, J.B. The Smell of Environmental Change: Using Floral Scent to Explain Shifts in Pollinator Attraction. Appl. Plant Sci. 2017, 5, 1600123. [Google Scholar] [CrossRef]
- Tholl, D.; Lee, S. Terpene Specialized Metabolism in Arabidopsis Thaliana. In The Arabidopsis Book; The American Society of Plant Biologists: Rockville, MD, USA, 2011; Volume 9, p. e0143. [Google Scholar] [CrossRef]
- Shanker Dubey, V.; Bhalla, R.; Luthra, R. An Overview of the Non-Mevalonate Pathway for Terpenoid Biosynthesis in Plants. J. Biosci. 2003, 28, 637–646. [Google Scholar] [CrossRef]
- Mak, W.C.K.; Walsh, S. The Characterization of Steam Distillation as an Extraction Method to Extract Volatile Compounds from Prunella vulgaris and the Investigation of Their Anti-Tumorous Effect. J. Biosci. Med. 2021, 09, 120–142. [Google Scholar] [CrossRef]
- Ayalew, Y.; Berhe, D.T.; Retta, N.; Desse, G.; Mohammed, A.; Kim, K.S. Volatile Organic Compounds of Anchote Tuber and Leaf Extracted Using Simultaneous Steam Distillation and Solvent Extraction. Int. J. Food Sci. 2022, 2022, 3265488. [Google Scholar] [CrossRef] [PubMed]
- Almeida, V.; Gonçalves, V.; Galego, L.; Miguel, G.; Costa, M. Volatile Constituents of Leaves and Flowers of Thymus Mastichina by Headspace Solid-Phase Microextraction. Acta Hortic. 2006, 723, 239–242. [Google Scholar] [CrossRef]
- Báez, D.; Pino, J.A.; Morales, D. Floral Scent Composition in Hedychium Coronarium J. Koenig Analyzed by SPME. J. Essent. Oil Res. 2011, 23, 64–67. [Google Scholar] [CrossRef]
- Song, G.; Xiao, J.; Deng, C.; Zhang, X.; Hu, Y. Use of Solid-Phase Microextraction as a Sampling Technique for the Characterization of Volatile Compounds Emitted from Chinese Daffodil Flowers. J. Anal. Chem. 2007, 62, 674–679. [Google Scholar] [CrossRef]
- Mohammadhosseini, M. Chemical Composition of the Volatile Fractions from Flowers, Leaves and Stems of Salvia Mirzayanii by HS-SPME-GC-MS. J. Essent. Oil Bear. Plants 2015, 18, 464–476. [Google Scholar] [CrossRef]
- Adam, M.; Juklová, M.; Bajer, T.; Eisner, A.; Ventura, K. Comparison of Three Different Solid-Phase Microextraction Fibres for Analysis of Essential Oils in Yacon (Smallanthus Sonchifolius) Leaves. J. Chromatogr. A 2005, 1084, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Stashenko, E.E.; Martínez, J.R. Sampling Flower Scent for Chromatographic Analysis. J. Sep. Sci. 2008, 31, 2022–2031. [Google Scholar] [CrossRef]
- Bicchi, C.; Drigo, S.; Rubiolo, P. Influence of Fibre Coating in Headspace Solid-Phase Microextraction-Gas Chromatographic Analysis of Aromatic and Medicinal Plants. J. Chromatogr. A 2000, 892, 469–485. [Google Scholar] [CrossRef]
- Bogusz, S.; de Marchi Tavares de Melo, A.; Zini, C.A.; Godoy, H.T. Optimization of the Extraction Conditions of the Volatile Compounds from Chili Peppers by Headspace Solid Phase Micro-Extraction. J. Chromatogr. A 2011, 1218, 3345–3350. [Google Scholar] [CrossRef]
- Deng, C.; Song, G.; Hu, Y. Application of HS-SPME and GC-MS to Characterization of Volatile Compounds Emitted from Osmanthus Flowers. Ann. Chim. 2004, 94, 921–927. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Umashankar, S.; Liang, X.; Lee, H.; Swarup, S.; Ong, C. Characterization of Plant Volatiles Reveals Distinct Metabolic Profiles and Pathways among 12 Brassicaceae Vegetables. Metabolites 2018, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.-M.; Benfodda, Z.; Bénimélis, D.; Fontaine, J.-X.; Molinié, R.; Meffre, P. Development of a HS-SPME/GC-MS Method for the Extraction and Identification of the Volatile Compounds Emitted by Flowers of Tillandsia Xiphioides. ACS Omega 2021, 6, 12691–12698. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.; Benfodda, Z.; Bénimélis, D.; Fontaine, J.; Molinié, R.; Meffre, P. Extraction and Identification of Volatile Organic Compounds Emitted by Fragrant Flowers of Three Tillandsia Species by HS-SPME/GC-MS. Metabolites 2021, 11, 594. [Google Scholar] [CrossRef] [PubMed]
- Ciotlaus, I.; Pojar-Fenesan, M.; Balea, A. Analysis of Volatile Organic Compounds from the Aerial Parts of Medicinal Plant, Galium verum. Rev. Chim. 2020, 71, 136–144. [Google Scholar] [CrossRef]
- Custódio, L.; Serra, H.; Nogueira, J.M.F.; Gonçalves, S.; Romano, A. Analysis of the Volatiles Emitted by Whole Flowers and Isolated Flower Organs of the Carob Tree Using HS-SPME-GC/MS. J. Chem. Ecol. 2006, 32, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, T.; Fan, Q.; Qi, X.; Zhang, F.; Fang, W.; Jiang, J.; Chen, F.; Chen, S. Identification of Floral Scent in Chrysanthemum Cultivars and Wild Relatives by Gas Chromatography-Mass Spectrometry. Molecules 2015, 20, 5346–5359. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yu, C.; Cheng, B.; Wan, H.; Luo, L.; Pan, H.; Zhang, Q. Volatile Compound Analysis and Aroma Evaluation of Tea-Scented Roses in China. Ind. Crops Prod. 2020, 155, 112735. [Google Scholar] [CrossRef]
- Shang, C.; Hu, Y.; Deng, C.; Hu, K. Rapid Determination of Volatile Constituents of Michelia Alba Flowers by Gas Chromatography-Mass Spectrometry with Solid-Phase Microextraction. J. Chromatogr. A 2002, 942, 283–288. [Google Scholar] [CrossRef]
- Bertrand, C.; Comte, G.; Piola, F. Solid-Phase Microextraction of Volatile Compounds from Flowers of Two Brunfelsia Species. Biochem. Syst. Ecol. 2006, 34, 371–375. [Google Scholar] [CrossRef]
- Sarti, G.C.; Gostoli, C.; Bandini, S. Extraction of Organic Components from Aqueous Streams by Vacuum Membrane Distillation. J. Membr. Sci. 1993, 80, 21–33. [Google Scholar] [CrossRef]
- Ormeño, E.; Goldstein, A.; Niinemets, Ü. Extracting and Trapping Biogenic Volatile Organic Compounds Stored in Plant Species. TrAC Trends Anal. Chem. 2011, 30, 978–989. [Google Scholar] [CrossRef]
- Epping, R.; Koch, M. On-Site Detection of Volatile Organic Compounds (VOCs). Molecules 2023, 28, 1598. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral Scents - a Checklist of Volatile Compounds Isolated by Headspace Techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Bicchi, C.; Joulain, D. Headspace-gas Chromatographic Analysis of Medicinal and Aromatic Plants and Flowers. Flavour. Fragr. J. 1990, 5, 131–145. [Google Scholar] [CrossRef]
- Arthur, C.L.; Pawliszyn, J. Solid Phase Microextraction with Thermal Desorption Using Fused Silica Optical Fibers. Anal. Chem. 1990, 62, 2145–2148. [Google Scholar] [CrossRef]
- Martendal, E.; de Souza Silveira, C.D.; Nardini, G.S.; Carasek, E. Use of Different Sample Temperatures in a Single Extraction Procedure for the Screening of the Aroma Profile of Plant Matrices by Headspace Solid-Phase Microextraction. J. Chromatogr. A 2011, 1218, 3731–3736. [Google Scholar] [CrossRef]
- Tholl, D.; Boland, W.; Hansel, A.; Loreto, F.; Röse, U.S.R.; Schnitzler, J.-P. Practical Approaches to Plant Volatile Analysis. Plant J. 2006, 45, 540–560. [Google Scholar] [CrossRef]
- Mejıas, R.C.; Marin, R.N.; Moreno, d.V.G.; Barroso, C.G. Optimisation of Headspace Solid-Phase Microextraction for Analysis of Aromatic Compounds in Vinegar. J. Chromatogr. A 2002, 953, 7–15. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Panigrahi, S. Solid-Phase Microextraction (SPME) Techniques for Quality Characterization of Food Products: A Review. Food Bioprocess. Technol. 2011, 4, 1–26. [Google Scholar] [CrossRef]
- Górecki, T.; Yu, X.; Pawliszyn, J. Theory of Analyte Extraction by Selected Porous Polymer SPME Fibres. Analyst 1999, 124, 643–649. [Google Scholar] [CrossRef]
- Shirey, R.E.; Mindrup, R.F. SPME-Adsorption versus Absorption: Which Fiber Is Best for Your Application? Supelco Sigma Aldrich Corp. 1999, 1–31. [Google Scholar]
- Gherghel, S.; Morgan, R.M.; Arrebola-liébanas, J.; Romero-gonzález, R.; Blackman, C.S.; Garrido-frenich, A.; Parkin, I.P. Development of a HS-SPME/GC- MS Method for the Analysis of Volatile Organic Compounds from Fabrics for Forensic Reconstruction Applications. Forensic Sci. Int. 2018, 290, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Wardencki, W.; Michulec, M.; Curyło, J. A Review of Theoretical and Practical Aspects of Solid-Phase Microextraction in Food Analysis. Int. J. Food Sci. Technol. 2004, 39, 703–717. [Google Scholar] [CrossRef]
- Bouchonnet, S. Introduction to GC-MS Coupling, 1st ed.; CRC Press: New York, NY, USA, 2013. [Google Scholar]
- Bouchonnet, S.; Libong, D. Le Couplage Chromatographie En Phase Gazeuse-Spectrométrie de Masse. Actual. Chim. 2004, 7–14. [Google Scholar]
- Hoffmann, E.d.; Stroobant, V. Mass Spectrometry: Principles and Applications, 3rd ed.; John Wiley & Sons: West Sussex, UK, 2007. [Google Scholar]
- El-Aneed, A.; Cohen, A.; Banoub, J. Mass Spectrometry, Review of the Basics: Electrospray, MALDI, and Commonly Used Mass Analyzers. Appl. Spectrosc. Rev. 2009, 44, 210–230. [Google Scholar] [CrossRef]
- Kováts, E. Gas-chromatographische Charakterisierung Organischer Verbindungen. Teil 1: Retentionsindices Aliphatischer Halogenide, Alkohole, Aldehyde Und Ketone. Helv. Chim. Acta 1958, 41, 1915–1932. [Google Scholar] [CrossRef]

| Compounds | Chemical Structure | Fam. 1 | Examples of Bioactivity: Pollination/Defense/Communication | Ref. |
|---|---|---|---|---|
| linalool | 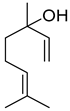 | M | Moth-pollinated Clarkia breweri flowers emission | [18] |
| Pollination by butterflies or nocturnal moths | [19,20,21] | |||
| Aphids repulsion | [14,22] | |||
| geraniol |  | M | Dominent messenger in chemical communication among bees, abundant emission from Coffea arabica flowers | [23,24,25] |
| (E)-β-ocimene |  | M | Systematic release by tobacco plants damaged by herbivore Heliothis virescens | [26] |
| α-pinene | 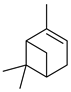 | M | Response to increasing drought stress: Ipomopsis aggregata flower emission highest percentage of the scent mixture when intermediate soil moisture | [27] |
| (E,E)-α-farnesene | 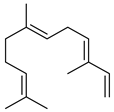 | S | Response to increasing drought stress: Ipomopsis aggregata flower emission highest percentage of the scent mixture when intense soil drought | [27] |
| (E)-α-bergamotene (α-trans-bergamotene) | 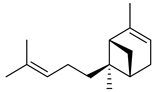 | S | Herbivore induced sesquiterpene | [28] |
| β-caryophyllene |  | S | Systematic release by tobacco plants damaged by herbivore Heliothis virescens | [26] |
| Emission by apple trees infected by Candidatus Phytoplasma mali | [29] | |||
| Arabidopsis thaliana floral defense against microbial pathogen | [30] | |||
| methyl benzoate |  | P/B | Snapdragon Antirrhinum majus floral VOC with negative allelopathic effects on Arabidopsis thaliana. | [31] |
| methyl salicylate and other salicylates | 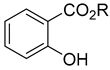 R=CH3, other | P/B | Released at a significantly higher rate from aphid-infested plants | [32,33,34] |
| (Z)-3-hexen-1-ol | 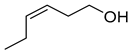 | Al | Can initiate a subset of regulatory steps and downstream metabolites to initiate defense responses in maize. Function as indirect plant defense mediating plant-plant signaling and intraplant information transfer. | [35,36] |
| (E) and (Z) 3-hexenal |  | A | Implicated in plants protection against insect herbivore attack. Numerous activities in plants pathogen resistance | [33,37] |
| indoles |  R=CH3: skatole | O | Stapelia gigantea or Wurmbea elatior flower emission for coprophagous insects attraction | [38,39] |
| Methods of Extraction of VOCs | Analytical Methods of VOCs | Plants | References (Year) |
|---|---|---|---|
| HS-SPME | GC-MS | Tropical flowers | [38] (2023) (review) |
| Steam distillation | GC-MS | Prunella vulgaris whole plant | [75] (2021) |
| Simultaneous steam distillation and solvent extraction | GC-MS | Coccinia abyssinica (Anchote) leaf and tuber | [76] (2022) |
| Hydrodistillation and HS-SPME | GC-MS | Thymus mastichina flowers and leaves | [77] (2006) |
| HS-SPME | GC-MS | Hedychium coronarium (ginger lily) flowers | [78] (2011) |
| HS-SPME | GC-MS | Narcissus tazetta var. chinensis (chinese daffodil) flowers | [79] (2007) |
| HS-SPME | GC-MS | Salvia mirzayanii flowers, leaves and stems | [80] (2015) |
| HS-SPME | GC-FID | Smallanthus sonchifolius leaves | [81] (2005) |
| HS-SPME | GC-MS, GC-FID and GC-ECD | Brugmansia suaveolens flowers | [82] (2008) (review) |
| HS-SPME and S-HS | GC-MS | Rosmarinus officinalis, Salvia officinalis, Thymus vulgaris, Valeriana officinalis | [83] (2000) |
| HS-SPME | GI-FID and GC-MS | Capsicum frutescens and other Capsicum chili peppers | [84] (2011) |
| HS-SPME | GC-MS | Osmanthus fragrans var. latifolius and Osmanthus fragrans var. thunbergii flowers | [85] (2004) |
| HS-SPME | GC-HRMS | Brassicaceae vegetables | [86] (2018) |
| HS-SPME | GC-MS | Tillandsia species | [67,87,88] (2021, 2022) |
| Hydrodistillation and HS-SPME | GC-MS | Galium verum aerial parts (leaves and flowers) | [89] (2020) |
| HS-SPME | GC-MS | Ceratonia siliqua (carob trees) flowers | [90] (2006) |
| HS-SPME | GC-MS | Silene latifolia flowers | [47] (2005) |
| HS-SPME | GC-MS | Chrysanthemum cultivar flowers | [91] (2015) |
| HS-SPME and HS-DTD | GC-MS | Rosa odorata and Rosa chinensis | [92] (2020) |
| HS-SPME | GC-MS | Michelia alba flowers | [93] (2002) |
| HS-SPME | GC-MS | Brunfelsia pauciflora and Brunfelsia australis | [94] (2006) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, M.-M.; Benfodda, Z.; Molinié, R.; Meffre, P. Volatile Organic Compounds Emitted by Flowers: Ecological Roles, Production by Plants, Extraction, and Identification. Plants 2024, 13, 417. https://doi.org/10.3390/plants13030417
Lo M-M, Benfodda Z, Molinié R, Meffre P. Volatile Organic Compounds Emitted by Flowers: Ecological Roles, Production by Plants, Extraction, and Identification. Plants. 2024; 13(3):417. https://doi.org/10.3390/plants13030417
Chicago/Turabian StyleLo, Mame-Marietou, Zohra Benfodda, Roland Molinié, and Patrick Meffre. 2024. "Volatile Organic Compounds Emitted by Flowers: Ecological Roles, Production by Plants, Extraction, and Identification" Plants 13, no. 3: 417. https://doi.org/10.3390/plants13030417
APA StyleLo, M.-M., Benfodda, Z., Molinié, R., & Meffre, P. (2024). Volatile Organic Compounds Emitted by Flowers: Ecological Roles, Production by Plants, Extraction, and Identification. Plants, 13(3), 417. https://doi.org/10.3390/plants13030417