
Effects of Funneliformis mosseae on Growth and Photosynthetic Characteristics of Camellia oleifera under Different Nitrogen Forms


Abstract
1. Introduction
2. Results
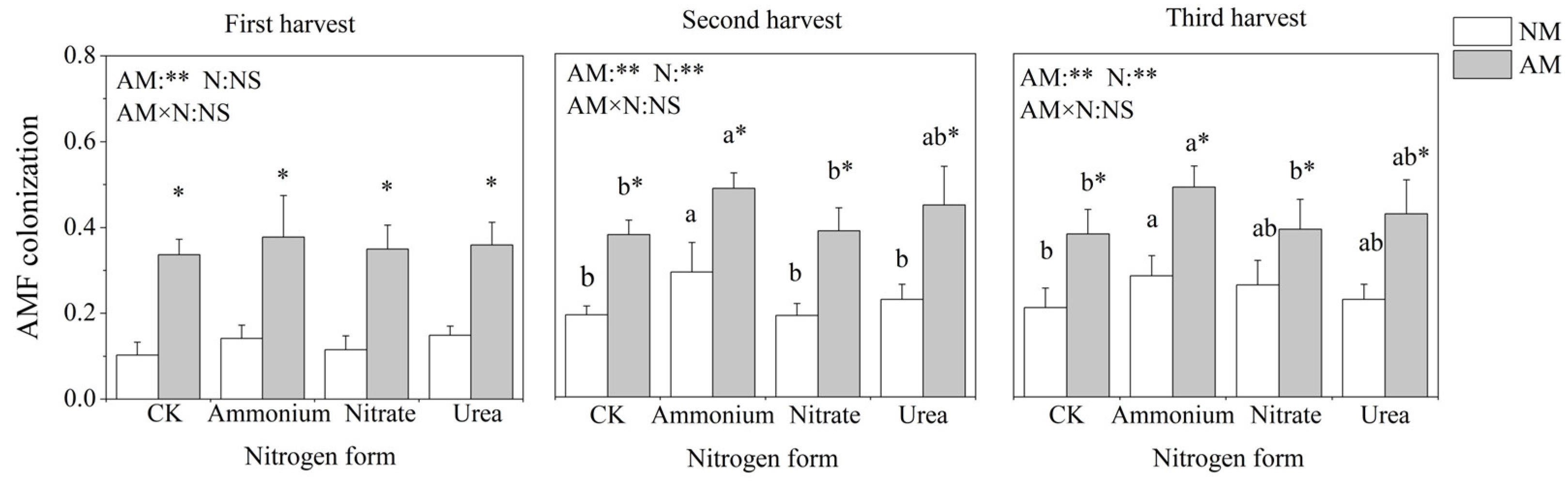
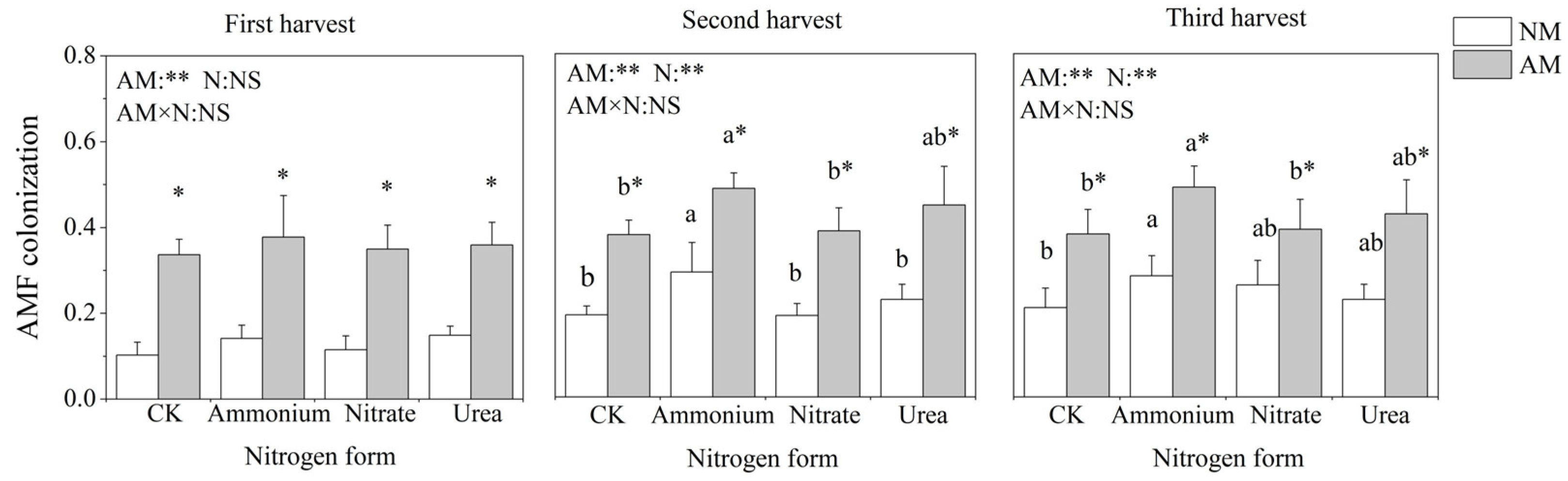
2.1. Mycorrhizal Colonization
2.2. Plant Growth
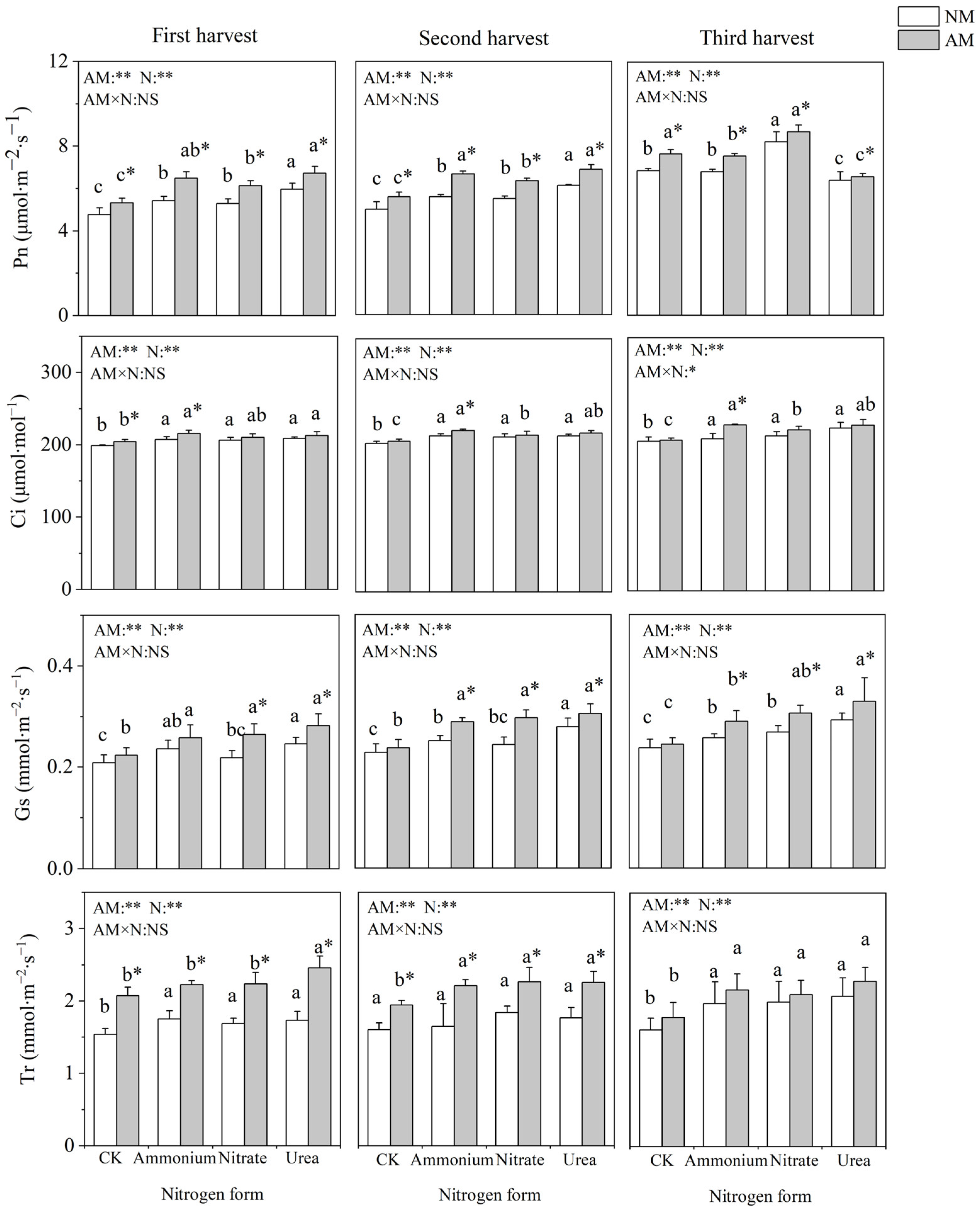
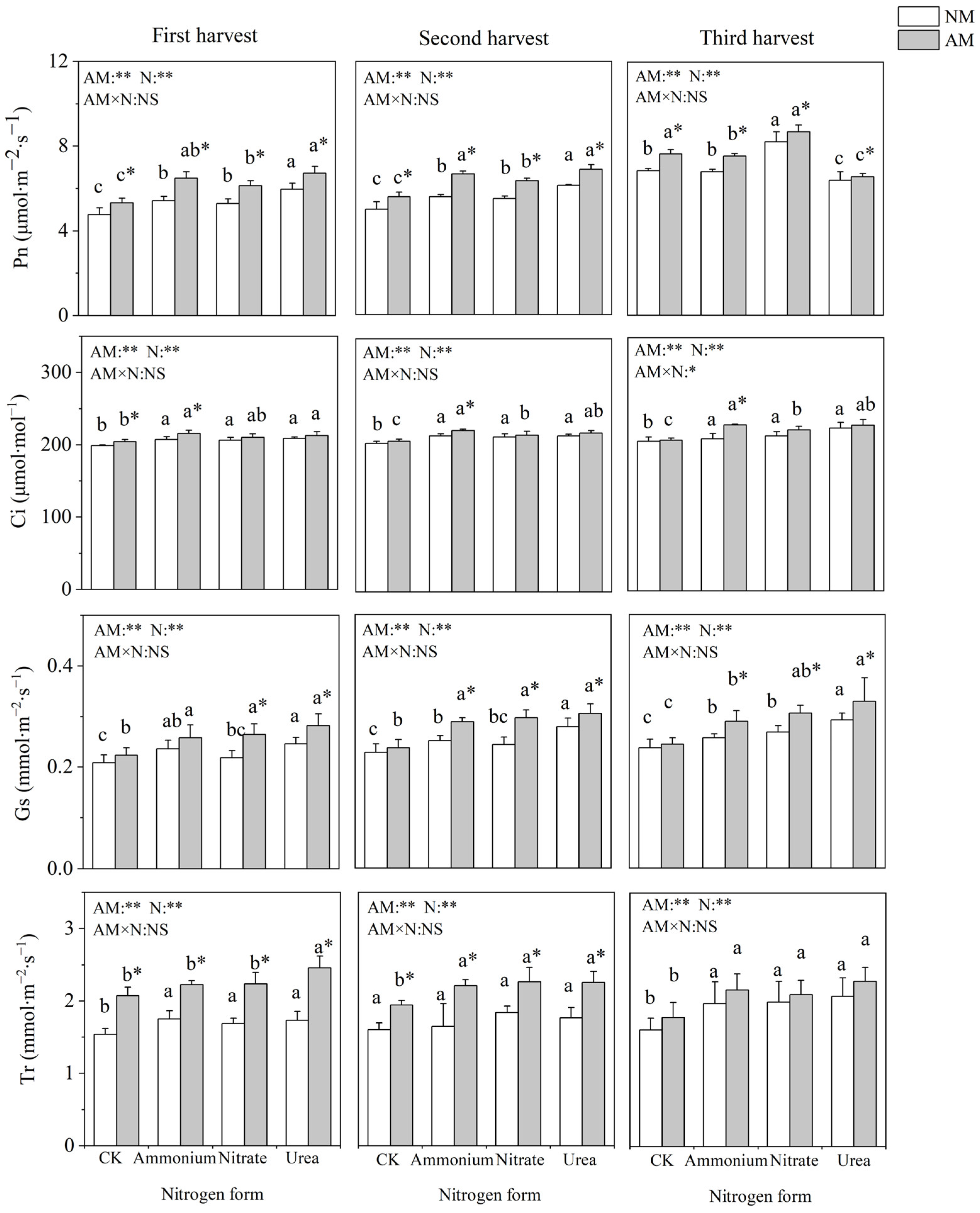
2.3. Gas Exchange Parameters
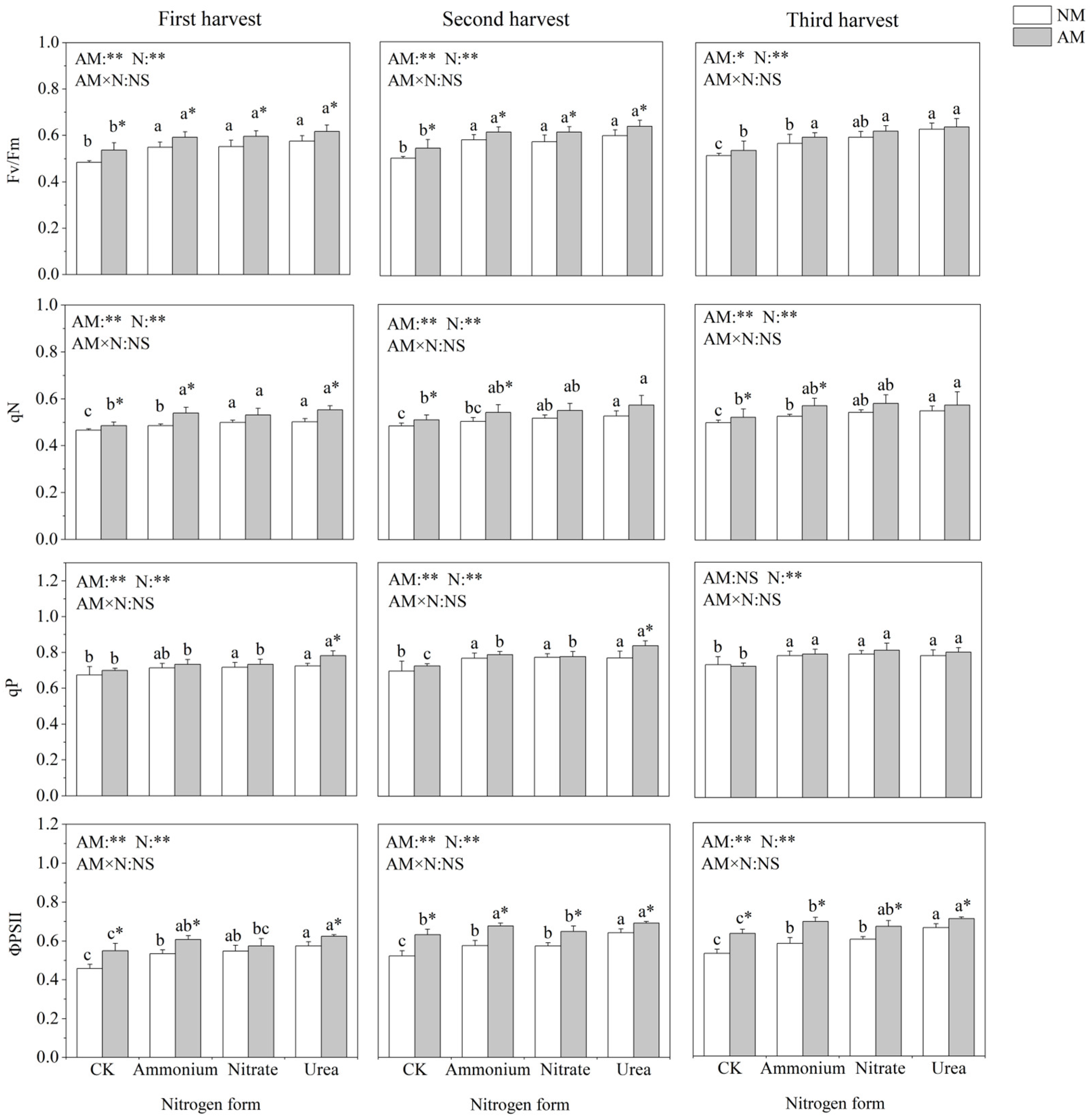
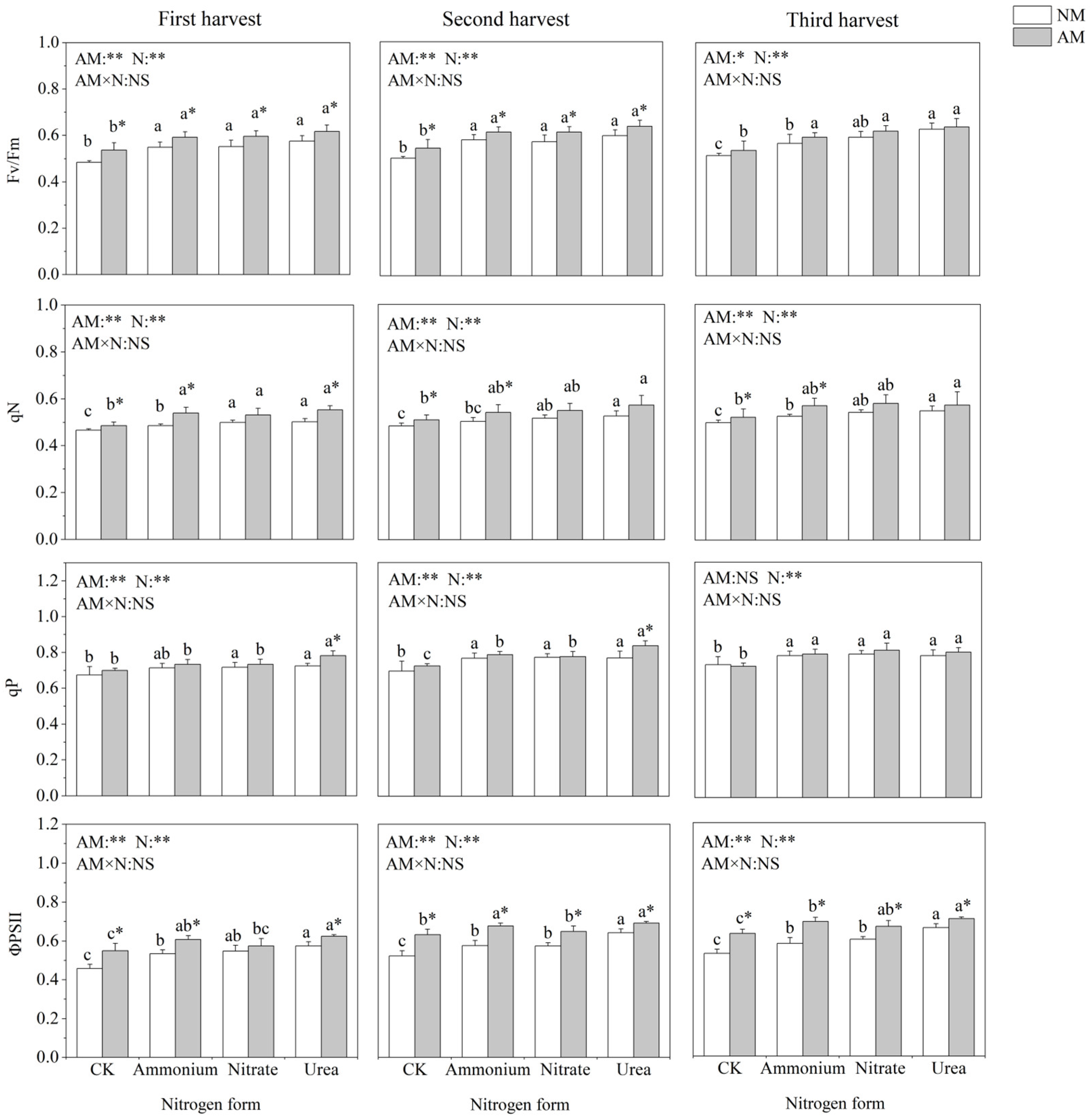
2.4. Chlorophyll Fluorescence Parameters
2.5. The Results of PCA
3. Discussion
3.1. AMF Colonization and Plant Growth
3.2. Funneliformis mosseae Improved Photosynthetic Characteristics of C. oleifera
3.3. Optimal Nitrogen Form of C. oleifera–AMF Symbiosis
4. Materials and Methods
4.1. Biological Materials and Soil
4.2. Experimental Design
4.3. Mycorrhizal Colonization
4.4. Plant Growth
4.5. Measurement of Gas Exchange Parameters and Chlorophyll Fluorescence Parameters
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Roosta, H.R.; Schjoerring, J.K. Effects of ammonium toxicity on nitrogen metabolism and elemental profile of cucumber plants. J. Plant Nutr. 2007, 30, 1933–1951. [Google Scholar] [CrossRef]
- Kronzucker, H.; Siddiqi, M.; Glass, A. Conifer root discrimination against soil nitrate and the ecology of forest succession. Nature 1997, 385, 59–61. [Google Scholar] [CrossRef]
- Mérigout, P.; Lelandais, M.; Bitton, F.; Renou, J.P.; Briand, X.; Meyer, C.; Daniel-Vedele, F. Physiological and transcriptomic aspects of urea uptake and assimilation in Arabidopsis plants. Plant Physiol. 2008, 147, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Houdusse, F.; Zamarreño, A.M.; Garnica, M.; García-Mina, J. The importance of nitrate in ameliorating the effects of ammonium and urea nutrition on plant development: The relationships with free polyamines and plant proline contents. Funct. Plant Biol. 2005, 32, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, G.; Erice, G.; Aroca, R.; Delgado-Huertas, A.; Ruiz-Lozano, J.M. Elucidating the possible involvement of maize aquaporins and arbuscular mycorrhizal symbiosis in the plant ammonium and urea transport under drought stress conditions. Plants 2020, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Fang, H.; Jiang, N.; Feng, W.; Luo, L.; Wang, J.; Wang, H.; Hu, D.; Guo, X.; Zhang, L. Biochar Is Comparable to dicyandiamide in the mitigation of nitrous oxide emissions from Camellia oleifera Abel. Fields. Forests 2019, 10, 1076. [Google Scholar] [CrossRef]
- Lee, W.T.; Tung, Y.T.; Wu, C.C.; Tu, P.S.; Yen, G.C. Camellia Oil (Camellia oleifera Abel.) Modifies the composition of gut microbiota and alleviates acetic acid-induced colitis in rats. J. Agric. Food Chem. 2018, 66, 7384–7392. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, Y.; Chen, M.; Tan, X.; Yuan, J. Transcriptomic and metabolomic profiling of Camellia oleifera seedling roots treated with different nitrogen forms. Plant Growth Regul. 2023, 99, 215–227. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal symbiosis. Q. Rev. Biol. 2008, 3, 273–281. [Google Scholar]
- Basu, S.; Rabara, R.C.; Negi, S. AMF: The future prospect for sustainable agriculture. Physiol. Mol. Plant Pathol. 2018, 102, 36–45. [Google Scholar] [CrossRef]
- Trouvelot, S.; Bonneau, L.; Redecker, D.; Van Tuinen, D.; Adrian, M.; Wipf, D. Arbuscular mycorrhiza symbiosis in viticulture: A review. Agron. Sustain. Dev. 2015, 35, 1449–1467. [Google Scholar] [CrossRef]
- Gui, H.; Purahong, W.; Wubet, T.; Peršoh, D.; Shi, L.; Khan, S.; Mortimer, P.E. Funneliformis mosseae alters soil fungal community dynamics and composition during litter decomposition. Fungal Ecol. 2020, 43, 100864. [Google Scholar] [CrossRef]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2015, 386, 1–19. [Google Scholar] [CrossRef]
- Cavagnaro, T.R.; Bender, S.F.; Asghari, H.R.; Van der Heijden, M.G. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015, 20, 283–290. [Google Scholar] [CrossRef]
- Tanaka, Y.; Yano, K. Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied. Plant Cell Environ. 2005, 28, 1247–1254. [Google Scholar] [CrossRef]
- Ngwene, B.; Gabriel, E.; George, E. Influence of different mineral nitrogen sources (NO3−-N vs. NH4+-N) on arbuscular mycorrhiza development and N transfer in a Glomus intraradices–cowpea symbiosis. Mycorrhiza 2013, 23, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Li, J.; Chen, Y.; Zhang, L.P.; Zhang, Y.; Wang, S.; Shi, X.; Li, L.; Liang, J.S. Effects of phosphate solubilizing bacteria on the growth, photosynthesis, and nutrient uptake of Camellia oleifera Abel. Forests 2019, 10, 348. [Google Scholar] [CrossRef]
- Balestrini, R.; Brunetti, C.; Chitarra, W.; Nerva, L. Photosynthetic traits and nitrogen uptake in crops: Which is the role of arbuscular mycorrhizal fungi? Plants 2020, 9, 1105. [Google Scholar] [CrossRef]
- Liu, T.; Wang, C.; Chen, H.; Fang, F.; Zhu, X.; Tang, M. Effects of arbuscular mycorrhizal colonization on the biomass and bioenergy production of Populus × canadensis ‘Neva’ in sterilized and unsterilized soil. Acta Physiol. Plant. 2014, 36, 871–880. [Google Scholar] [CrossRef]
- He, X.; Chen, Y.; Liu, S.; Gunina, A.; Wang, X.L.; Chen, W.L.; Shao, Y.H.; Shi, L.L.; Yao, Q.; Li, J.X.; et al. Cooperation of earthworm and arbuscular mycorrhizae enhanced plant N uptake by balancing absorption and supply of ammonia. Soil Biol. Biochem. 2018, 116, 351–359. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, C.; Li, J.; Wang, G.A. Effects of arbuscular mycorrhizal fungi on maize nitrogen uptake strategy under different soil water conditions. Plant Soil 2021, 464, 441–452. [Google Scholar] [CrossRef]
- Higo, M.; Takahashi, Y.; Gunji, K.; Isobe, K. How are arbuscular mycorrhizal associations related to maize growth performance during short-term cover crop rotation? J. Sci. Food Agric. 2018, 98, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.N.; Shu, B.; Wu, Q.S. Role of arbuscular mycorrhizas in citrus (Citrus spp.) crop production—A review. Curr. Hortic. 2020, 8, 3–6. [Google Scholar] [CrossRef]
- Bakonyi, G.; Posta, K.; Kiss, I.; Fábián, M.; Nagy, P.; Nosek, J.N. Density-dependent regulation of arbuscular mycorrhiza by collembola. Soil Biol. Biochem. 2002, 34, 661–664. [Google Scholar] [CrossRef]
- Ndoye, F.; Kane, A.; Bakhoum, N.; Sanon, A.; Fall, D.; Diouf, D.; Sylla, S.N.; Bâ, A.M.; Sy, M.O.; Noba, K. Response of Acacia senegal (L.) Willd. to inoculation with arbuscular mycorrhizal fungi isolates in sterilized and unsterilized soils in Senegal. Agrofor. Syst. 2013, 87, 941–952. [Google Scholar] [CrossRef]
- Berger, F.; Gutjahr, C. Factors affecting plant responsiveness to arbuscular mycorrhiza. Curr. Opin. Plant Biol. 2021, 59, 101994. [Google Scholar] [CrossRef]
- Magalhaes, J.R.; Wilcox, G.E. Ammonium toxicity development in tomato plants relative to nitrogen form and light intensity. J. Plant Nutr. 1984, 10, 1477–1496. [Google Scholar] [CrossRef]
- Song, D.L.; Zhao, Y.F.; Tang, F.P.; Zhang, Y.H.; Zhou, S.Q.; Dong, L.J. Effects of arbuscular mycorrhizal fungi on Solidago canadensis growth are independent of nitrogen form. J. Plant Ecol. 2021, 14, 648–661. [Google Scholar] [CrossRef]
- Wang, W.H.; Köhler, B.; Cao, F.Q.; Liu, G.W.; Gong, Y.Y.; Sheng, S.; Song, Q.C.; Cheng, X.Y.; Garnett, T.; Okamoto, M.; et al. Rice DUR3 mediates high-affinity urea transport and plays an effective role in improvement of urea acquisition and utilization when expressed in Arabidopsis. New Phytol. 2012, 193, 432–444. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef]
- Raines, C.A. Increasing photosynthetic carbon assimilation in C3 plants to improve crop yield: Current and future strategies. Plant Physiol. 2011, 155, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The combined effects of arbuscular mycorrhizal fungi (AMF) and lead (Pb) stress on Pb accumulation, plant growth parameters, photosynthesis, and antioxidant enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef] [PubMed]
- Gavito, M.E.; Jakobsen, I.; Mikkelsen, T.N.; Mora, F. Direct evidence for modulation of photosynthesis by an arbuscular mycorrhiza-induced carbon sink strength. New Phytol. 2019, 223, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wang, Y.; Sun, S.; Mu, C.; Yan, X. Effects of arbuscular mycorrhizal fungi on the growth, photosynthesis and photosynthetic pigments of Leymus chinensis seedlings under salt-alkali stress and nitrogen deposition. Sci. Total Environ. 2017, 576, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcon, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef] [PubMed]
- Kaschuk, G.; Kuyper, T.W.; Leffelaar, P.A.; Hungria, M.; Giller, K.E. Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses? Soil Biol. Biochem. 2009, 41, 1233–1244. [Google Scholar] [CrossRef]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Hawkins, H.J.; Johansen, A.; George, E. Uptake and transport of organic and inorganic nitrogen by arbuscular mycorrhizal fungi. Plant Soil 2000, 226, 275–285. [Google Scholar] [CrossRef]
- Thirkell, T.; Cameron, D.; Hodge, A. Contrasting nitrogen fertilisation rates alter mycorrhizal contribution to barley nutrition in a field trial. Front. Plant Sci. 2019, 10, 1312. [Google Scholar] [CrossRef]
- Phillips, J.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| AM Treatment | N Treatment | Height (cm) | Stem Diameter (cm) | Root Length (cm) | Root Average Diameter (cm) | Root Volume (cm3) | Root Surface Area (cm2) |
|---|---|---|---|---|---|---|---|
| NM | CK | 11.06 ± 1.62 b | 2.6 ± 0.31 | 127.86 ± 11.72 b | 0.65 ± 0.11 b | 0.73 ± 0.11 b | 51.03 ± 2.34 c |
| Ammonium | 13.72 ± 2.03 ab | 2.81 ± 0.26 | 148.49 ± 14.25 a | 0.84 ± 0.13 a | 1.11 ± 0.1 a | 66 ± 4.35 b | |
| Nitrate | 13.44 ± 2.63 ab | 2.8 ± 0.36 | 153.37 ± 20.08 a | 0.98 ± 0.08 a | 1.17 ± 0.07 a | 67.45 ± 5.85 b | |
| Urea | 15.78 ± 2.43 a | 2.88 ± 0.49 | 162.07 ± 13.56 a | 0.85 ± 0.15 a | 1.18 ± 0.05 a | 78.35 ± 4.72 a | |
| AM | CK | 11.74 ± 1.96 b | 2.8 ± 0.31 | 151.7 ± 10.3b * | 0.73 ± 0.12 b | 0.85 ± 0.05 c * | 53.76 ± 2.34 c |
| Ammonium | 15.34 ± 2.05 a | 2.94 ± 0.23 | 165.75 ± 16.31 ab | 0.94 ± 0.19 a | 1.27 ± 0.17 b | 77.67 ± 6.19 b * | |
| Nitrate | 15.24 ± 1.93 a | 2.86 ± 0.22 | 178.94 ± 24.36 a | 1.02 ± 0.15 a | 1.37 ± 0.14 ab * | 76.25 ± 5.48 b * | |
| Urea | 16.92 ± 2.07 a | 2.87 ± 0.37 | 186.89 ± 11.86 a * | 0.98 ± 0.12 a | 1.47 ± 0.17 a * | 87.91 ± 5.12 a * | |
| Two-way ANOVA | |||||||
| N | 9.417 ** | 0.661 NS | 8.696 ** | 9.380 ** | 42.778 ** | 72.016 ** | |
| AM | 3.853 NS | 0.854 NS | 20.257 ** | 4.447 * | 27.163 ** | 29.631 ** | |
| N × AM | 0.143 NS | 0.189 NS | 0.143 NS | 0.198 NS | 0.999 NS | 1.626 NS | |
| AM Treatment | N Treatment | Height (cm) | Stem Diameter (mm) | Root Length (cm) | Root Average Diameter (cm) | Root Volume (cm3) | Root Surface Area (cm2) |
|---|---|---|---|---|---|---|---|
| NM | CK | 18.3 ± 0.58 c | 3.92 ± 0.15 b | 186.83 ± 9.01 d | 0.7 ± 0.11 c | 1.03 ± 0.07 c | 54.25 ± 2.09 c |
| Ammonium | 22.1 ± 1.6 b | 4.2 ± 0.19 b | 216.23 ± 9.84 c | 0.92 ± 0.12 ab | 1.23 ± 0.04 b | 72.11 ± 9.21 b | |
| Nitrate | 22.35 ± 0.47 b | 4.12 ± 0.13 b | 221.62 ± 10.62 b | 1.04 ± 0.07 a | 1.31 ± 0.11 ab | 74.37 ± 9.17 ab | |
| Urea | 24.3 ± 1.48 a | 4.66 ± 0.43 a | 262.16 ± 9.22 a | 0.82 ± 0.05 b | 1.39 ± 0.13 a | 83.28 ± 3.88 a | |
| AM | CK | 20.04 ± 0.84 c * | 3.98 ± 0.18 b | 197.79 ± 3.66 c * | 0.73 ± 0.1 c | 1.04 ± 0.11 b | 56.87 ± 1.96 c |
| Ammonium | 24.38 ± 1.84 ab | 4.14 ± 0.11 b | 220.19 ± 9.26 b | 0.9 ± 0.16 b | 1.33 ± 0.16 a | 81.48 ± 3.81 b | |
| Nitrate | 22.42 ± 1.95 b | 4.25 ± 0.33 b | 233.58 ± 15.6 b | 1.04 ± 0.14 b | 1.3 ± 0.08 a | 79.17 ± 4.91 b | |
| Urea | 25.44 ± 2.04 a | 4.33 ± 0.33 a | 284.6 ± 10.7 a * | 1.01 ± 0.14 a * | 1.42 ± 0.1 6a | 87.4 ± 4.99 a | |
| Two-way ANOVA | |||||||
| N | 26.455 ** | 8.898 ** | 109.797 ** | 13.511 ** | 18.508 ** | 50.408 ** | |
| AM | 7.857 ** | 0.494 NS | 14.606 ** | 1.842 * | 0.841 ** | 8.538 ** | |
| N × AM | 1.029 NS | 1.863 NS | 1.395 NS | 1.600 NS | 0.361 NS | 0.663 NS | |
| AM Treatment | N Treatment | Height (cm) | Stem Diameter (cm) | Root Length (cm) | Root Average Diameter (cm) | Root Volume (cm3) | Root Surface Area (cm2) |
|---|---|---|---|---|---|---|---|
| NM | CK | 20.06 ± 2.65 c | 4.8 ± 0.46 b | 207.89 ± 11.06 c | 0.72 ± 0.11 c | 1.04 ± 0.07 c | 56.75±1.02 c |
| Ammonium | 24.42 ± 1.77 b | 4.52 ± 0.54 b | 238.34 ± 3.84 b | 0.93 ± 0.12 ab | 1.24 ± 0.04 b | 74.32±9.2 b | |
| Nitrate | 31 ± 3.8 a | 4.82 ± 0.39 b | 261.44 ± 10.6 b | 1.03 ± 0.04 a | 1.31 ± 0.11 ab | 76.57±9.11 b | |
| Urea | 32.1 ± 3.57 a | 5.62 ± 0.48 a | 280.67 ± 9.97 a | 0.89 ± 0.07 b | 1.39 ± 0.13 a | 85.21±3.57 a | |
| AM | CK | 19.92 ± 2.36 c | 4.84 ± 0.17 b | 209.89 ± 2.59 c | 0.74 ± 0.10 b | 1.04 ± 0.11 b | 57.36±3.31 c |
| Ammonium | 28.36 ± 2.36b * | 5.6 ± 0.50a * | 236.82 ± 8.01 b | 0.91 ± 0.16 ab | 1.33 ± 0.16 a | 82.86±3.77 b | |
| Nitrate | 32.9 ± 1.75 a | 5.56 ± 0.48 a * | 249.61 ± 14.57 b * | 1.05 ± 0.15 a | 1.3 ± 0.08 a | 80.5±5.18 b | |
| Urea | 33.84 ± 2.97 a | 5.96 ± 0.43 a | 302.47 ± 10.7 a * | 1.01 ± 0.14 a * | 1.42 ± 0.16 a | 89.63±4.99 a | |
| Two-way ANOVA | |||||||
| N | 47.244 ** | 8.630 ** | 128.236 ** | 12.816 ** | 18.338 ** | 51.706 ** | |
| AM | 4.585 * | 15.316 ** | 0.731 NS | 1.205 NS | 0.734 NS | 5.918 * | |
| N × AM | 0.921 NS | 2.621 NS | 5.308 ** | 0.740 NS | 0.343 NS | 0.819 NS | |
| Index | Principal Component 1 | Principal Component 2 |
|---|---|---|
| Height (X1) | 0.850 | 0.367 |
| Stem diameter (X2) | 0.732 | 0.558 |
| Root length (X3) | 0.869 | 0.321 |
| Root average diameter (X4) | 0.514 | −0.546 |
| Root volume (X5) | 0.753 | −0.405 |
| Root surface area (X6) | 0.811 | −0.396 |
| Pn (X7) | 0.89 | 0.273 |
| Gs (X8) | 0.852 | −0.043 |
| Ci (X9) | 0.844 | 0.017 |
| Tr (X10) | −0.101 | 0.318 |
| Fv/Fm (X11) | 0.802 | −0.327 |
| qP (X12) | 0.866 | 0.052 |
| qP (X13) | 0.776 | −0.092 |
| qN (X13) | 0.776 | 0.008 |
| ΦPSⅡ (X14) | 0.798 | 0.367 |
| AM Treatment | N Treatment | Y1 | Y2 | Comprehensive Score | Rank |
|---|---|---|---|---|---|
| NM | CK | 128.486 | 92.156 | 8168.572 | 7 |
| Ammonium | 151.076 | 48.249 | 8176.879 | 6 | |
| Nitrate | 155.811 | 51.355 | 8471.011 | 5 | |
| Urea | 167.637 | 56.171 | 9135.742 | 2 | |
| AM | CK | 140.568 | 47.017 | 7658.589 | 8 |
| Ammonium | 160.421 | 48.937 | 8628.098 | 4 | |
| Nitrate | 162.188 | 52.896 | 8804.381 | 3 | |
| Urea | 178.048 | 61.283 | 9741.72 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Wang, C.; Ma, Z.; Zhang, L.; Wu, F. Effects of Funneliformis mosseae on Growth and Photosynthetic Characteristics of Camellia oleifera under Different Nitrogen Forms. Plants 2024, 13, 370. https://doi.org/10.3390/plants13030370
Huang Y, Wang C, Ma Z, Zhang L, Wu F. Effects of Funneliformis mosseae on Growth and Photosynthetic Characteristics of Camellia oleifera under Different Nitrogen Forms. Plants. 2024; 13(3):370. https://doi.org/10.3390/plants13030370
Chicago/Turabian StyleHuang, Yuxuan, Chuangxin Wang, Ziran Ma, Linping Zhang, and Fei Wu. 2024. "Effects of Funneliformis mosseae on Growth and Photosynthetic Characteristics of Camellia oleifera under Different Nitrogen Forms" Plants 13, no. 3: 370. https://doi.org/10.3390/plants13030370
APA StyleHuang, Y., Wang, C., Ma, Z., Zhang, L., & Wu, F. (2024). Effects of Funneliformis mosseae on Growth and Photosynthetic Characteristics of Camellia oleifera under Different Nitrogen Forms. Plants, 13(3), 370. https://doi.org/10.3390/plants13030370