
Wood Anatomical and Demographic Similarities Between Self-Standing Liana and Tree Seedlings in Tropical Dry Forests of Colombia

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Results
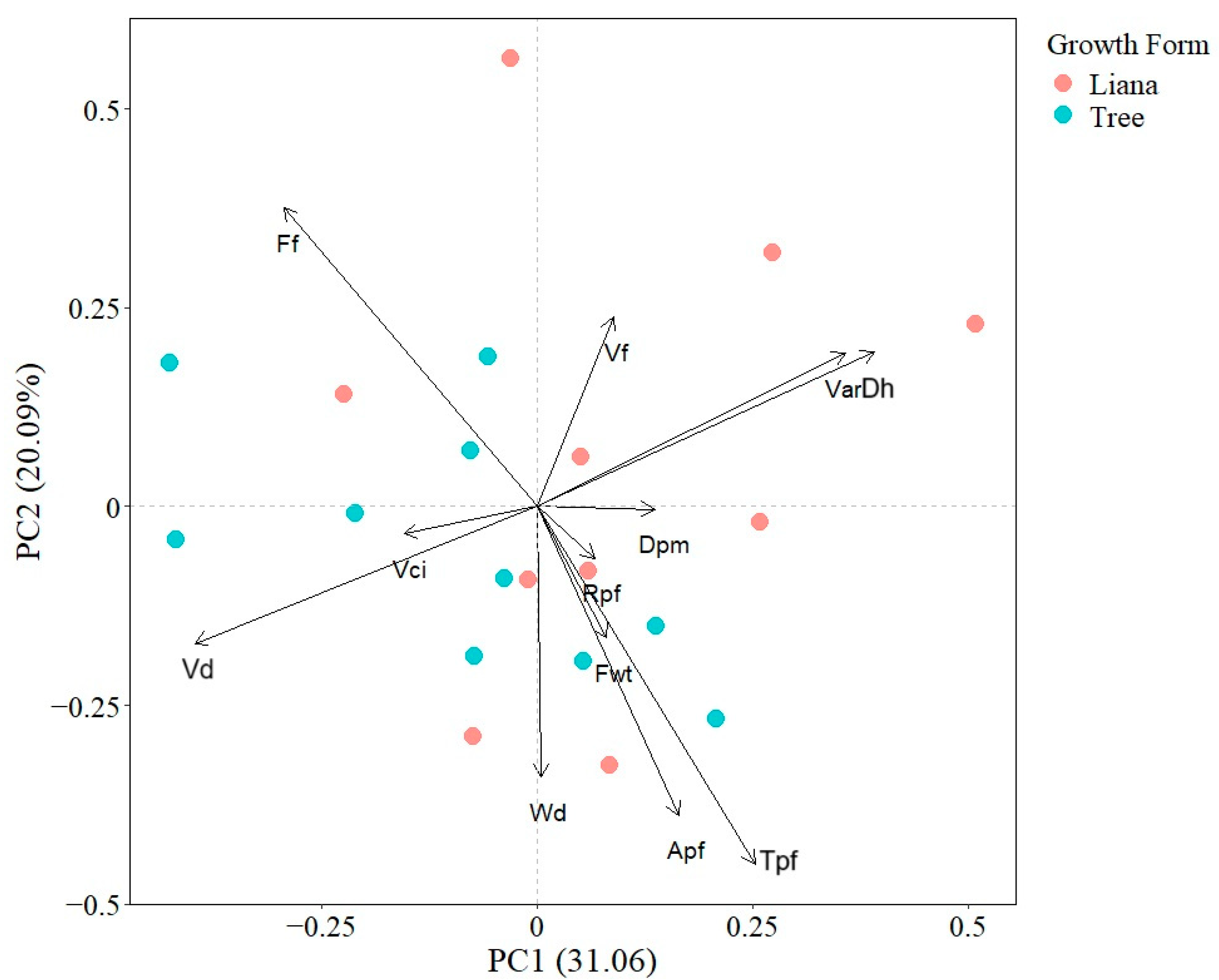
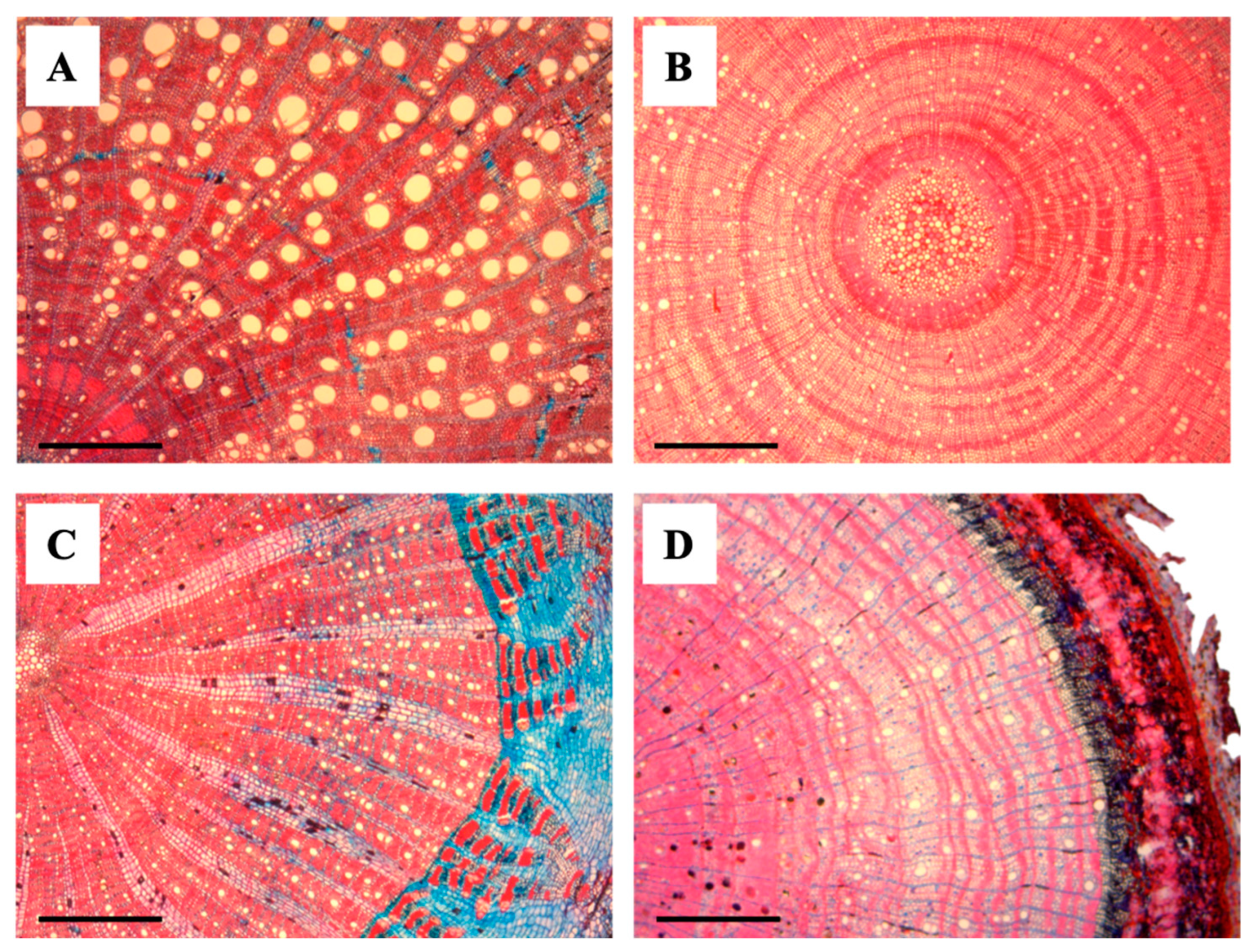
2.1. Wood Anatomical Differences Between Liana and Tree Seedlings
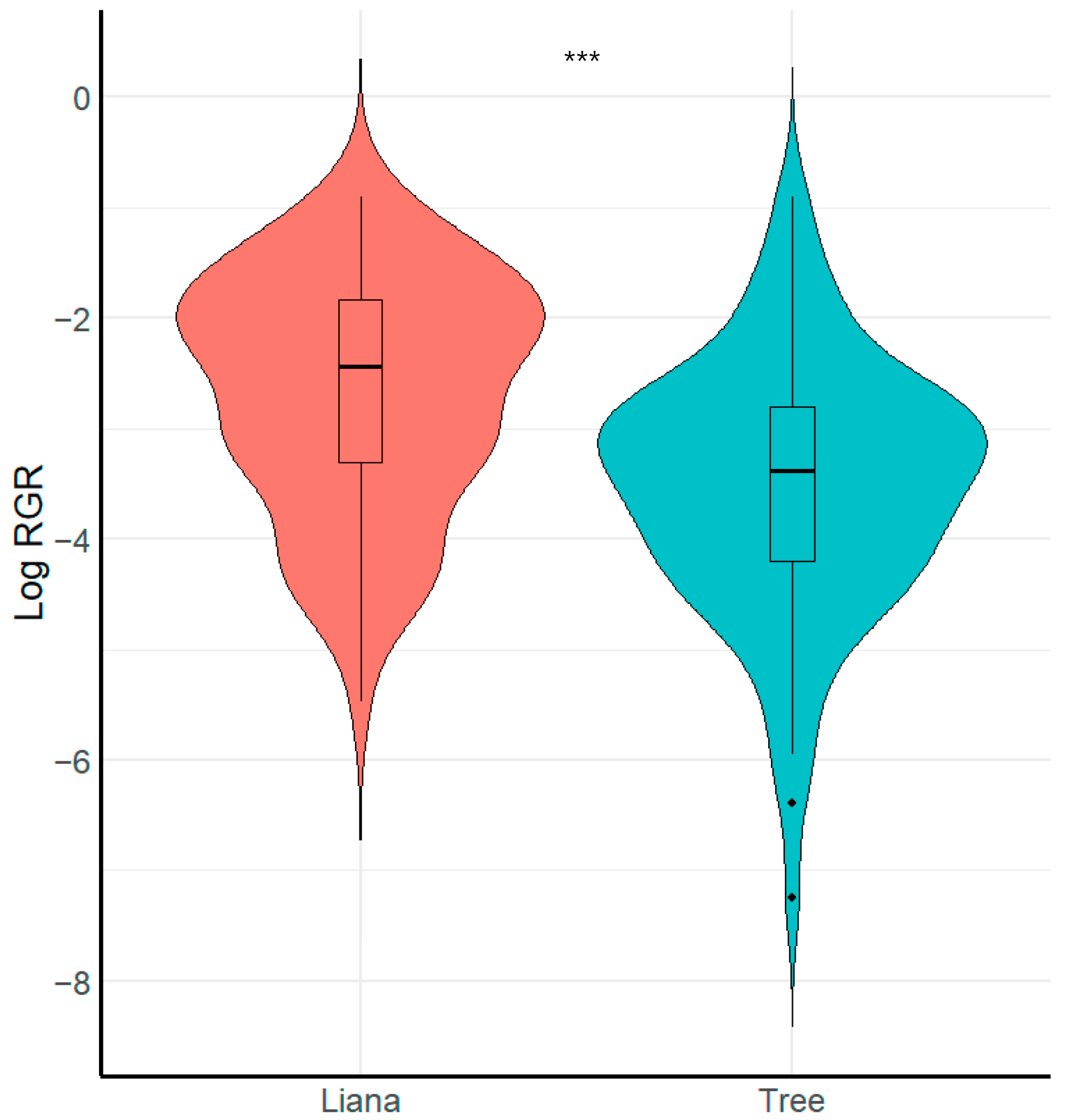
Differences in Relative Growth Rate (RGR) and Survival Between Liana and Tree Seedlings
3. Discussion
3.1. Wood Anatomical Differences and Similarities Between Liana and Tree Seedlings
3.2. Demographic Differences Between Liana and Tree Seedlings
4. Materials and Methods
4.1. Study Sites
4.2. Species Selection
4.3. Trait Measurement
4.4. Seedling Growth and Survival
4.5. Light Conditions
4.6. Data Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Putz, F. The Natural History of Lianas on Barro Colorado Island, Panama. Ecology 1984, 65, 1713–1724. [Google Scholar] [CrossRef]
- Ingwell, L.L.; Joseph Wright, S.; Becklund, K.K.; Hubbell, S.P.; Schnitzer, S.A. The impact of lianas on 10 years of tree growth and mortality on Barro Colorado Island, Panama. J. Ecol. 2010, 98, 879–887. [Google Scholar] [CrossRef]
- Álvarez-Cansino, L.; Schnitzer, S.A.; Reid, J.P.; Powers, J.S. Liana Competition with Tropical Trees Varies Seasonally but not with Tree Species Identity. Ecology 2015, 96, 39–45. [Google Scholar] [CrossRef] [PubMed]
- DeWalt, S.J.; Schnitzer, S.A.; Alves, L.F.; Bongers, F.; Burnham, R.J.; Cai, Z.; Carson, W.P.; Chave, J.; Chuyong, G.B.; Costa, F.R.C.; et al. Biogeographical patterns of liana abundance and diversity. In Ecology of Lianas; John Wiley & Sons: Chichester, UK, 2015; pp. 131–146. [Google Scholar]
- Schnitzer, S.A. Increasing liana abundance in neotropical forests: Causes and consequences. In Ecology of Lianas; John Wiley & Sons: Chichester, UK, 2015; pp. 451–464. [Google Scholar]
- Parolari, A.J.; Paul, K.; Griffing, A.; Condit, R.; Perez, R.; Aguilar, S.; Schnitzer, S.A. Liana abundance and diversity increase with rainfall seasonality along a precipitation gradient in Panama. Ecography 2020, 43, 25–33. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Carson, W.P. Lianas suppress tree regeneration and diversity in treefall gaps. Ecol. Lett. 2010, 13, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, S.C.; Wright, S.J.; Santiago, L.S. Lianas always outperform tree seedlings regardless of soil nutrients: Results from a long-term fertilization experiment. Ecology 2015, 96, 1866–1876. [Google Scholar] [CrossRef]
- Van der Heijden GM, F.; Powers, J.S.; Schnitzer, S.A. Lianas reduce carbon accumulation and storage in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 13267–13271. [Google Scholar] [CrossRef]
- Martínez-Izquierdo, L.; García, M.M.; Powers, J.S.; Schnitzer, S.A. Lianas suppress seedling growth and survival of 14 tree species in a Panamanian tropical forest. Ecology 2016, 97, 215–224. [Google Scholar] [CrossRef]
- Schnitzer, S.A. Testing ecological theory with lianas. New Phytol. 2018, 220, 366–380. [Google Scholar] [CrossRef]
- Angyalossy, V.; Pace, M.R.; Lima, A.C. Liana anatomy: A broad perspective on structural evolution of the vascular system. In Ecology of Lianas; John Wiley & Sons: Chichester, UK, 2015; pp. 251–287. [Google Scholar]
- Ewers, F.W.; Rosell, J.A.; Olson, M.E. Lianas as structural parasites. In Functional and Ecological Xylem Anatomy; Springer International Publishing: Cham, Switzerland, 2015; pp. 163–188. [Google Scholar]
- Schnitzer, S.A.; van der Heijden, G.M.F. Lianas have a seasonal growth advantage over co-occurring trees. Ecology 2019, 100, e02655. [Google Scholar] [CrossRef]
- Smith-Martin, C.M.; Bastos, C.L.; Lopez, O.R.; Powers, J.S.; Schnitzer, S.A. Effects of dry-season irrigation on leaf physiology and biomass allocation in tropical lianas and trees. Ecology 2019, 100, e02827. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A. A Mechanistic Explanation for Global Patterns of Liana Abundance and Distribution. Am. Nat. 2005, 166, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, B.; Wright, S.J.; Muller-Landau, H.C.; Kitajima, K.; Hernandé, A.S. Life history trade-offs in tropical trees and lianas. Ecology 2006, 87, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A.; Bongers, F. The ecology of lianas and their role in forests. Trends Ecol. Evol. 2002, 17, 223–230. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Mortality Rates of 205 Neotropical Tree and Shrub Species and the Impact of a Severe Drought. Ecol. Monogr. 1995, 65, 419–439. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragão, L.E.O.C.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; López-González, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the Amazon rainforest. Science 2002, 323, 1344–1347. [Google Scholar] [CrossRef]
- Rowe, N.; Speck, T. Plant growth forms: An ecological and evolutionary perspective. New Phytol. 2005, 166, 61–72. [Google Scholar] [CrossRef]
- Umaña, M.N.; Manzané-Pinzón, E.; Comita, L.S. Long-term dynamics of liana seedlings suggest decelerating increases in liana relative abundance over time. J. Ecol. 2020, 108, 460–469. [Google Scholar] [CrossRef]
- Umaña, M.N.; Forero-Montaña, J.; Nytch, C.J.; Thompson, J.; Uriarte, M.; Zimmerman, J.; Swenson, N.G. Dry conditions and disturbance promote liana seedling survival and abundance. Ecology 2019, 100, e01502. [Google Scholar] [CrossRef]
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Hubbell, S.P.; Foster, R.B. Assessing recruitment limitation: Concepts, methods and case-studies from a tropical forest. In Seed Dispersal and Frugivory: Ecology, Evolution and Conservation; CABI Publishing: Wallingford, UK, 2003; pp. 35–53. [Google Scholar]
- Green, P.T.; Harms, K.E.; Connell, J.H. Nonrandom, diversifying processes are disproportionately strong in the smallest size classes of a tropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 18649–18654. [Google Scholar] [CrossRef]
- Carlquist, S. Observations on functional wood histology of vines and lianas: Vessel dimorphism, tracheids, vasicentric tracheids, narrow vessels and parenchyma. Aliso A J. Syst. Florist. Bot. 1985, 11, 139–157. [Google Scholar]
- Rosell, J.A.; Olson, M.E. Do lianas really have wide vessels? Vessel diameter-stem length scaling in non-self-supporting plants. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 288–295. [Google Scholar] [CrossRef]
- Olson, M.E.; Anfodillo, T.; Rosell, J.A.; Petit, G.; Crivellaro, A.; Isnard, S.; León-Gómez, C.; Alvarado-Cárdenas, L.O.; Castorena, M. Universal hydraulics of the flowering plants: Vessel diameter scales with stem length across angiosperm lineages, habits and climates. Ecol. Lett. 2014, 17, 988–997. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Yang, D.; Zhang, Y.B.; Liu, Q.; Wang, Y.S.D.; Ke, Y.; Xiao, Y.; Wang, Q.; Dossa, G.G.O.; Schnitzer, S.A.; et al. Vessel dimorphism and wood traits in lianas and trees among three contrasting environments. Am. J. Bot. 2023, 110, e16154. [Google Scholar] [CrossRef] [PubMed]
- Van der Sande, M.T.; Poorter, L.; Schnitzer, S.A.; Engelbrecht, B.M.J.; Markesteijn, L. The hydraulic efficiency–safety trade-off differs between lianas and trees. Ecology 2019, 100, e02666. [Google Scholar] [CrossRef]
- Morris, H.; Plavcová, L.; Cvecko, P.; Fichtler, E.; Gillingham, M.A.F.; Martínez-Cabrera, H.I.; Mcglinn, D.J.; Wheeler, E.; Zheng, J.; Ziemińska, K.; et al. A global analysis of parenchyma tissue fractions in secondary xylem of seed plants. New Phytol. 2016, 209, 1553–1565. [Google Scholar] [CrossRef]
- Fisher, J.B.; Ewerst, F.W. Wound healing in stems of lianas after twisting and girdling injuries. Bot. Gaz. 1989, 150, 251–265. [Google Scholar] [CrossRef]
- Muscarella, R.; Uriarte, M.; Forero-Montaña, J.; Comita, L.S.; Swenson, N.G.; Thompson, J.; Nytch, C.J.; Jonckheere, I.; Zimmerman, J.K. Life-history trade-offs during the seed-to-seedling transition in a subtropical wet forest community. J. Ecol. 2013, 101, 171–182. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Santiago, L.S.; Goldstein, G.; Meinzer, F.C.; Fisher, J.B.; Machado, K.; Woodruff, D.; Jones, T. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia 2004, 140, 543–550. [Google Scholar] [CrossRef]
- De Guzman, M.E.; Santiago, L.S.; Schnitzer, S.A.; Álvarez-Cansino, L. Trade-offs between water transport capacity and drought resistance in neotropical canopy liana and tree species. Tree Physiol. 2017, 37, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Carlquist, S. Cambial Variants (Anomalous Secondary Growth). In Comparative Wood Anatomy: Systematic, Ecological, and Evolutionary Aspects of Dicotyledon Wood; Springer: Berlin/Heidelberg, Germany, 2001; pp. 271–295. [Google Scholar]
- Isnard, S.; Feild, T.S. The evolution of angiosperm lianescence: A perspective from xylem structure-function. In Ecology of Lianas; John Wiley & Sons: Chichester, UK, 2015; pp. 221–238. [Google Scholar]
- Pace, M.R.; Lohmann, L.G.; Angyalossy, V. The rise and evolution of the cambial variant in Bignonieae (Bignoniaceae). Evol. Dev. 2009, 11, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Luizon Dias Leme, C.; da Cunha Neto, I.L.; Angyalossy, V. How the neotropical liana Machaerium multifoliolatum (Fabaceae) develop their distinctive flattened stems? Flora 2020, 269, 151629. [Google Scholar] [CrossRef]
- Hacke, U.G.; Spicer, R.; Schreiber, S.G.; Plavcová, L. An ecophysiological and developmental perspective on variation in vessel diameter. Plant Cell Environ. 2017, 40, 831–845. [Google Scholar] [CrossRef] [PubMed]
- Zanne, A.E.; Westoby, M.; Falster, D.S.; Ackerly, D.D.; Loarie, S.R.; Arnold, S.E.J.; Coomes, D.A. Angiosperm wood structure: Global patterns in vessel anatomy and their relation to wood density and potential conductivity. Am. J. Bot. 2010, 97, 207–215. [Google Scholar] [CrossRef]
- Poorter, L.; McDonald, I.; Alarcón, A.; Fichtler, E.; Licona, J.C.; Peña-Claros, M.; Sterck, F.; Villegas, Z.; Sass-Klaassen, U. The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytol. 2010, 185, 481–492. [Google Scholar] [CrossRef]
- Coley, P.D. Hervibory and defensive characteristics of tree species in a lowland tropical forest. Ecol. Monogr. 1983, 53, 209–234. [Google Scholar] [CrossRef]
- Poorter, L.; Kitajima, K. Carbohydrate storage and light requirements of tropical moist and dry forest tree species. Ecology 2007, 88, 1000–1011. [Google Scholar] [CrossRef]
- Morris, H.; Brodersen, C.; Schwarze, F.W.M.R.; Jansen, S. The Parenchyma of Secondary Xylem and Its Critical Role in Tree Defense against Fungal Decay in Relation to the CODIT Model. Front. Plant Sci. 2016, 7, 1665. [Google Scholar] [CrossRef]
- Hubbell, S.P. The maintenance of diversity in a neotropical tree community: Conceptual issues, current evidence, and challenges ahead. In Forest Biodiversity Research, Monitoring and Modeling: Conceptual Background and Old World Case Studies; Dallmeier, F., Comiskey, J.A., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 17–44. [Google Scholar]
- Chang-Yang, C.H.; Needham, J.; Lu, C.L.; Hsieh, C.F.; Sun, I.F.; McMahon, S.M. Closing the life cycle of forest trees: The difficult dynamics of seedling-to-sapling transitions in a subtropical rainforest. J. Ecol. 2021, 109, 2705–2716. [Google Scholar] [CrossRef]
- González-M., R.; Posada, J.M.; Carmona, C.P.; Garzón, F.; Salinas, V.; Idárraga-Piedrahita, Á.; Pizano, C.; Avella, A.; López-Camacho, R.; Norden, N.; et al. Diverging functional strategies but high sensitivity to an extreme drought in tropical dry forests. Ecol. Lett. 2021, 24, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.B.; Wiemann, M.C. Measuring wood specific gravity Correctly. Am. J. Bot. 2010, 97, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Carlquist, S. Fibre dimorphism: Cell type diversification as an evolutionary strategy in angiosperm woods. Bot. J. Linn. Soc. 2014, 174, 44–67. [Google Scholar] [CrossRef]
- Scholz, A.; Klepsch, M.; Karimi, Z.; Jansen, S. How to quantify conduits in wood? Front. Plant Sci. 2013, 4, 56. [Google Scholar] [CrossRef]
- Tyree, M.T.; Zimmermann, M.H. The Vessel Network in the Stem. In Xylem Structure and the Ascent of Sap; Springer Series in Wood Science; Springer: Berlin/Heidelberg, Germany, 2002; pp. 27–48. [Google Scholar]
- Hietz, P.; Rosner, S.; Hietz-Seifert, U.; Wright, S.J. Wood traits related to size and life history of trees in a Panamanian rainforest. New Phytol. 2017, 213, 170–180. [Google Scholar] [CrossRef]
- Poorter, H.; Garnier, E. Ecological Significance of Inherent Variation in Relative Growth Rate and Its Components. In Functional Plant Ecology; Pugnaire, F., Valladares, F., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 67–100. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Trait | Unit | t-Value | p-Value | Mean | Range |
|---|---|---|---|---|---|
| Mean hydraulically weighted vessel diameter | µm | 1.815 | 0.089 | 47.124 | 24.365–121.283 |
| Vessel fraction | % | −0.091 | 0.928 | 7.423 | 2.897–20.522 |
| Vessel density | Vessels per mm2 | −2.395 | 0.028 | 53.794 | 5.671–198.8 |
| Horizontal pit membrane diameter aperture | µm | −0.293 | 0.774 | 7.533 | 5.008–11.712 |
| Variance in vessel diameter | unitless | 2.122 | 0.048 | 121.22 | 3.15–885.353 |
| Vessel clustering index | unitless | −0.389 | 0.702 | 1.384 | 1–4.04 |
| Fiber wall thickness | µm | 0.397 | 0.696 | 3.798 | 2.573–8.56 |
| Fiber fraction | % | −0.41703 | 0.6817 | 54.952 | 33.469–83.461 |
| Wood density | g/cm−3 | −0.274 | 0.788 | 0.563 | 0.281–0.665 |
| Axial parenchyma fraction | % | −0.934 | 0.363 | 23.503 | 0.0652–43.238 |
| Radial parenchyma fraction | % | 1.551 | 0.139 | 15.651 | 4.049–45.58 |
| Total parenchyma fraction | % | 0.184 | 0.855 | 37.625 | 10.643–56.75 |
| Growth Form | Family | Species | Mean | Sd | Range |
|---|---|---|---|---|---|
| Liana | Apocynaceae | Forsteronia spicata | 0.125 | 0.103 | 0.004–0.408 |
| Bignoniaceae | Bignonia pterocalyx | 0.088 | 0.051 | 0.009–0.152 | |
| Celastraceae | Hippocratea volubilis | 0.039 | 0 | 0.039–0.039 | |
| Fabaceae | Abrus precatorius | - | - | - | |
| Fabaceae | Machaerium isadelphum | 0.038 | 0.013 | 0.024–0.051 | |
| Nyctaginaceae | Pisonia aculeata | 0.045 | 0.043 | 0.009–0.108 | |
| Sapindaceae | Paullinia aff. cururu | 0.040 | 0 | 0.040–0.040 | |
| Sapindaceae | Paullinia cururu | 0.108 | 0.076 | 0.014–0.200 | |
| Sapindaceae | Paullinia sp1 | 0.010 | 0.069 | 2.928–3.066 | |
| Sapindaceae | Paullinia sp2 | 0.068 | 1.362 | 0.103–3.060 | |
| Tree | Annonaceae | Oxandra venezuelana | 0.026 | 1.441 | 0.012–2.993 |
| Fabaceae | Coursetia ferruginea | 0.052 | 0.011 | 0.042–0.063 | |
| Fabaceae | Inga edulis | 0.008 | 0 | 2.575–2.575 | |
| Fabaceae | Neltuma juliflora | 0.067 | 0 | 0.067–0.067 | |
| Fabaceae | Pterocarpus rohrii | 0.068 | 0.062 | 0.003–0.151 | |
| Malpighiaceae | Malpighia glabra | 0.074 | 0.178 | 0.001–0.804 | |
| Meliaceae | Trichilia acuminata | 0.051 | 1.464 | 0.006–3.095 | |
| Phyllanthaceae | Phyllanthus botryanthus | 0.012 | 0.003 | 0.009–0.015 | |
| Rutaceae | Amyris pinnata | 0.058 | 0.046 | 0.018–0.134 | |
| Sapindaceae | Melicoccus bijugatus | 0.048 | 0.036 | 0.005–0.171 |
| Plot | Altitude (AMSL) | Coordinates | Total Annual Precipitation (TAP; mm) | Mean Annual Temperature (MAT; °C) |
|---|---|---|---|---|
| Tayrona | 15 | 11.31° N, −74.13° W | 899.4 | 27.38 |
| Colorados | 301 | 9.94° N, −75.11° W | 1528.4 | 26.1 |
| Cotové | 385 | 6.53° N, −75.83° W | 1193.8 | 26.92 |
| Growth Form | Family | Species | Plot |
|---|---|---|---|
| Liana | Apocynaceae | Forsteronia spicata (Jacq.) G. Mey. | Cotové |
| Bignoniaceae | Bignonia pterocalyx (Sprague ex Urb.) L.G. Lohmann | Tayrona | |
| Celastraceae | Hippocratea volubilis L. | Tayrona | |
| Fabaceae | Abrus precatorius L. | Cotové | |
| Fabaceae | Machaerium microphyllum (E. Mey.) Standl. | Cotové | |
| Nyctaginaceae | Pisonia aculeata L. | Cotové | |
| Sapindaceae | Paullinia aff. cururu L. | Tayrona | |
| Sapindaceae | Paullinia cururu L. | Cotové | |
| Sapindaceae | Paullinia sp1 | Colorados | |
| Sapindaceae | Paullinia sp2 | Colorados | |
| Tree | Annonaceae | Oxandra venezuelana R.E. Fr. | Colorados |
| Fabaceae | Coursetia ferruginea (Kunth) Lavin | Tayrona | |
| Fabaceae | Inga vera Willd | Colorados | |
| Fabaceae | Prosopis juliflora (Sw.) DC. | Tayrona | |
| Fabaceae | Pterocarpus rohrii Vahl | Tayrona | |
| Malpighiaceae | Malpighia glabra L. | Cotové | |
| Meliaceae | Trichilia acuminata (Humb. & Bonpl. ex Roem. & Schult.) C. DC. | Colorados | |
| Phyllanthaceae | Phyllanthus botryanthus Müll. Arg. | Cotové | |
| Rutaceae | Amyris pinnata Kunth | Cotové | |
| Sapindaceae | Melicoccus bijugatus Jacq. | Cotové |
| Trait (Abbreviation) | Unit | Description | Function |
|---|---|---|---|
| Axial parenchyma fraction (Apf) | % | Percentage of stem cross-sectional area allocated to axial parenchyma | Storage and structural flexibility |
| Mean hydraulically weighted vessel diameter (Dh) | µm | Mean diameter that all of the vessels in a stem would have in order to correspond to the overall conductivity for the same numbers of conduits | Water transport efficiency and safety |
| Horizontal pit membrane diameter aperture (Dpm) | µm | Horizontal pit membrane diameter | Water transport safety |
| Fiber fraction (Ff) | % | Percentage of stem cross-sectional area allocated to fibers | Mechanical stability |
| Fiber wall thickness (Fwt) | µm | Double wall between adjacent fibers | Mechanical stability |
| Radial parenchyma fraction (Rpf) | % | Percentage of stem cross-sectional area allocated to radial parenchyma | Storage and structural flexibility |
| Total parenchyma fraction (Tpf) | % | Percentage of stem cross-sectional area allocated to total parenchyma | Storage and structural flexibility |
| Variance in vessel diameter (VarDh) | Unitless | Vessel diameter variance | Water transport efficiency and safety |
| Vessel clustering index (Vci) | Unitless | Total number of vessels divided by the number of vessel groups. | Water transport efficiency and safety |
| Vessel density (Vd) | Vessels per mm2 | Number of conduits per cross-sectional area | Water transport safety |
| Vessel fraction (Vf) | % | Percentage of stem cross-sectional area allocated to vessels | Water transport capacity |
| Wood (density (Wd)) | g/cm−3 | Oven-dry mass divided by saturated volume of the wood section. | Mechanical stability Water transport safety |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puentes-Marín, J.; González-Melo, A.; Salgado-Negret, B.; González-M, R.; Abad Ferrer, J.; Benavides, J.P.; Cely, J.M.; Idárraga-Piedrahita, Á.; Moreno, E.; Pizano, C.; et al. Wood Anatomical and Demographic Similarities Between Self-Standing Liana and Tree Seedlings in Tropical Dry Forests of Colombia. Plants 2024, 13, 3023. https://doi.org/10.3390/plants13213023
Puentes-Marín J, González-Melo A, Salgado-Negret B, González-M R, Abad Ferrer J, Benavides JP, Cely JM, Idárraga-Piedrahita Á, Moreno E, Pizano C, et al. Wood Anatomical and Demographic Similarities Between Self-Standing Liana and Tree Seedlings in Tropical Dry Forests of Colombia. Plants. 2024; 13(21):3023. https://doi.org/10.3390/plants13213023
Chicago/Turabian StylePuentes-Marín, Juliana, Andrés González-Melo, Beatriz Salgado-Negret, Roy González-M, Julio Abad Ferrer, Juan Pablo Benavides, Juan Manuel Cely, Álvaro Idárraga-Piedrahita, Esteban Moreno, Camila Pizano, and et al. 2024. "Wood Anatomical and Demographic Similarities Between Self-Standing Liana and Tree Seedlings in Tropical Dry Forests of Colombia" Plants 13, no. 21: 3023. https://doi.org/10.3390/plants13213023
APA StylePuentes-Marín, J., González-Melo, A., Salgado-Negret, B., González-M, R., Abad Ferrer, J., Benavides, J. P., Cely, J. M., Idárraga-Piedrahita, Á., Moreno, E., Pizano, C., Pulido, N., Rivera, K., Rojas-Bautista, F., Solorzano, J. F., & Umaña, M. N. (2024). Wood Anatomical and Demographic Similarities Between Self-Standing Liana and Tree Seedlings in Tropical Dry Forests of Colombia. Plants, 13(21), 3023. https://doi.org/10.3390/plants13213023