
Advances in the Involvement of Metals and Metalloids in Plant Defense Response to External Stress

Abstract
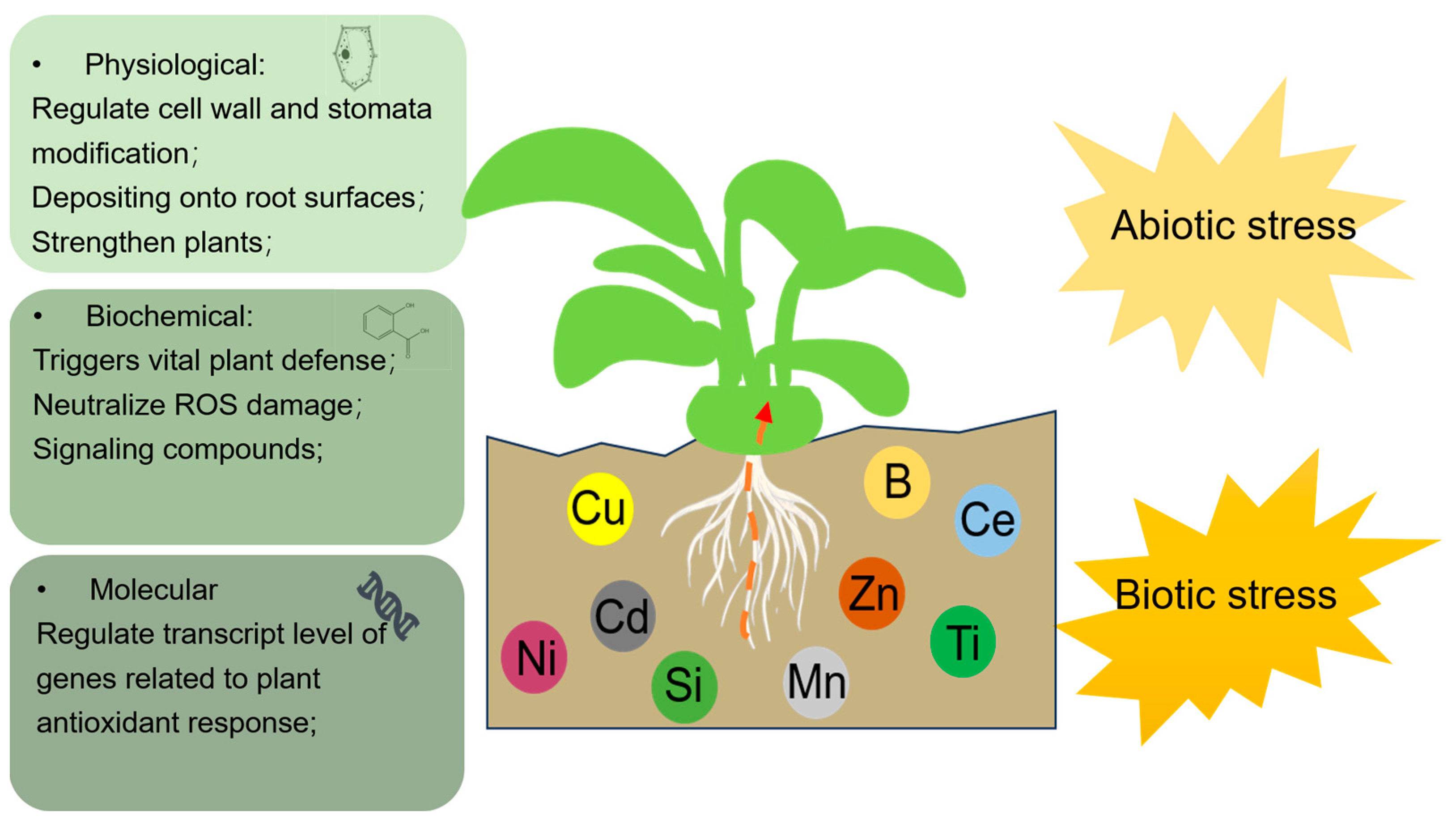
1. Introduction
2. Metal(Loid) Defense to Abiotic Stress
2.1. Zinc (Zn)
2.2. Silicon (Si)
2.3. Boron (B)
2.4. Cerium (Ce)
2.5. Titanium (Ti)

{kind=link}
| Metal(loid) | Species | Alleviation | Process | Reference |
|---|---|---|---|---|
| Zinc (Zn) | Cosmos bipinnatus | Cd toxicity | Inhibit Cd uptake, regulate Cd subcellular distributions, and induce the activities of plant antioxidative enzymes; | [28] |
| Triticum aestivum L. and Oryza sativa L. | Cd toxicity | Improve plant growth and enhance enzyme activity related to the plant’s antioxidant response; | [29] | |
| Lactuca sativa L. | Cd toxicity | Improve plant growth, maintain photosynthesis efficiency, induce plant antioxidant responses, and reduce Cd accumulation; | [31] | |
| Glycine max L. | As toxicity | Reverse growth inhibition, alleviate oxidative stress; | [34] | |
| Pleioblastus pygmaeus (Miq.) Nakai | As and Pb toxicity | Increase plant antioxidant activities, regulate stomatal closure, inhibit toxic metal uptake, and regulate their translocation; | [35] | |
| Zea mays L. | Salinity stress | Alleviate the adverse effect of salinity associated with photosynthesis and reverse the reduction of water and turgor potentials caused by salinity stress; | [36] | |
| Triticum aestivum L. | Salinity stress | Inhibit the negative effect of salinity, improve plant growth, and regulate physiological performance; | [37] | |
| Oryza sativa L. | Cr(VI) toxicity | Increase GSH and auxin synthesis and induce root hair formation; | [61] | |
| Oryza sativa L. | Cr(III) toxicity | Obstruct Cr(III) transport capacity via the apoplastic route; | [63] | |
| Solanum melongena | Pb toxicity | Induce plant antioxidant activity, promote plant growth, and reverse oxidant damage caused by Pb; | [64] | |
| Oryza sativa L. | As(III) toxicity | Compete binding sites and transporters; | [52] | |
| Zea mays L. | As(V) toxicity | Regulate the AsA-GSH cycle and counterbalance ROS content; | [54] | |
| Triticum aestivum L. | Drought | Reshape root architecture and induce plant defense; | [66] | |
| Brassica juncea | Drought, salinity | Induce plant defense and maintain chlorophyll biosynthesis; | [67,68] | |
| Boron (B) | Glycine max L. | Salinity stress | Enhance antioxidant defense and mitigate oxidative damage; | [80] |
| Gossypium spp. | Salinity stress | Reverse the damage effect and protect cell wall structure and antioxidant defense-related components; | [81] | |
| Poncirus trifoliate (L.) Raf. | Al toxicity | Reduce the biding domain and demethylation of pectin in Al; down-regulate genes for Al transport; | [71,75] | |
| Oryza sativa L., Pisum sativum, and Brassica napus | Cd toxicity | Increase Cd adsorption to cell walls and induce plant antioxidant responses; | [76,77] | |
| Citrullus lanatus | V toxicity | Sequester V in the root cell vacuole, increase the chelation of V in the cell wall, and induce the plant’s antioxidant defense; | [78] | |
| Cerium (Ce) | Arabidopsis thaliana | ROS damage | Act as a catalyzer used by chloroplast to increase the ROS scavenging; | [84] |
| Zea mays L. | Salinity stress | Modified root formation, altered Na+ fluxes, plant defense response, and enhanced rhizobacterial enrichment; | [85] | |
| Brassica napus | Salinity stress | Shorten root length to accelerate Na+ transport to aerial tissues to relieve root stress; | [86] | |
| Oryza sativa L. | Cd and salt | Trigger antioxidant defense systems and significantly relieve damage; | [87] | |
| Titanium (Ti) | Oryza sativa L. | Cd toxicity | Reduce Cd accumulation and alleviate negative effects caused by Cd stress; | [89,90] |
| Helianthus annuus L. | Cr(VI) toxicity | Reduce Cr accumulation, preserve the photosynthetic pigments, and bolster antioxidant enzyme activities; | [91] | |
| Vigna radiata L. | As toxicity | Reduce As accumulation and induce the expression of genes associated with antioxidant mechanisms; | [92] | |
| Oryza sativa L. | Pb toxicity | Decrease Pb bioavailability and regulate the transcript level of genes related to Pb transport; improve plant growth by promoting nutrient utilization; | [93] | |
| Vicia faba L. | Water deficit | Maintain the water content of leaves, decrease electrolyte leakage, and induce nitrate reductase activity; | [95] | |
| Calendula officinalis L. | Salinity stress | Reduce oxidant damage, improve photosynthetic efficiency, and increase antioxidant activity; | [96] | |
| Cicer arietinum L. | Cold stress | Enhance the expression level of genes related to plant antioxidative activity; | [97] | |
| Sesamum indicum L. | Heat stress | Improve plant growth, reduce hydrogen peroxide content | [98] |
3. Metal(Loid) Defense against Biotic Stress
3.1. Cadmium (Cd)
3.2. Zinc (Zn)
3.3. Copper (Cu)
3.4. Silicon (Si)
3.5. Nickel (Ni)
3.6. Titanium (Ti)
3.7. Other Metals
| Metal(loid) | Species | Alleviation | Process | Reference |
|---|---|---|---|---|
| Cadmium (Cd) | Thlaspi caerulescens | Pseudomonas syringae | Inhibit the growth of Pseudomonas syringae and enhance plant disease resistance; | [104] |
| Brassica juncea | Alternaria brassicicola | Enhance JA and SA accumulation in leaves and induce the expression of pathogen-related genes; | [107] | |
| Ageratina adenophora | Rhizoctonia solani | Inhibit the growth of pathogens and reduce lesion aeras; | [108] | |
| Populus yunnanensis | Pestalotiopsis microspora | Restructure the consistency of the phyllosphere microorganisms; improve plant resistance to pathogens; | [111] | |
| Arabidopsis halleri | Pieris rapae | Reduce the insect herbivory feeding rate; | [112] | |
| Populus yunnanensis | Botyodes diniasalis and Spodoptera exigua | Decrease the generalist herbivore survival rate; reduce the specialist herbivore feeding rate; | [113] | |
| Solanum lycopersicum | Tetranychus urticae | Toxic to herbivores and inhibit herbivore growth; | [114] | |
| Arabidopsis halleri | Myzus persicae | Present in phloem and can be toxic to aphids; | [115] | |
| Populus yunnanensis | Stauronematus compressicornis and Plagiodera versicolora | Induce volatile organic compounds; prevent the oviposition of females of both specialist and generalist herbivores; | [117] | |
| Larix olgensis | Lymantria dispar | Improve antioxidant enzyme activity and enhance resistance to gypsy moth larvae; reduce body weight of larvae; improve; | [120] | |
| Zinc (Zn) | Arabidopsis halleri | Alternaria brassicae | Induce antifungal defense and promote the synthesis of camalexin; | [122] |
| Nicotiana attenuata | Pseudomonas syringae | Interrupt the growth of bacteria, regulate the transcript level of genes related to the hormone signal pathway, and stimulate the closeness of stomata; | [126] | |
| Cucumis melo | Acidovorax citrulli | Improve plant growth, activate the plant’s antioxidant response to bacterial infection, and restrict the incidence of bacteria; | [127] | |
| Oryza sativa L. | Magnaporthe grisea | Destruct the cell wall of Magnaporthe grisea; reverse the negative effect caused by fungi; | [128] | |
| Brassica juncea | Alternaria brassicae | Facilitate plant growth and suppress disease occurrence; | [129] | |
| Copper (Cu) | Lycopersicon esculentum | Fusarium oxysporum | Reduce Fusarium wilt fungus estimates and increase the growth and yield of tomato plants; | [133] |
| Eleusine coracana Gaertn. | Pyricularia grisea | Improve defense enzyme activities and inhibit the development of blast disease; | [134] | |
| Oryza sativa L. | Cnaphalocrocis medinalis | Inhibit Cnaphalocrocis medinalis growth; | [136] | |
| Silicon (Si) | Saccharum spp. hybrids | Xanthomonas albilineans | Reduce the disease index and simultaneously lower the bactericide concentration; | [142] |
| Solanum lycopersicum | Ralstonia solanacearum | Stimulate antioxidant activity, eliminate ROS, and inhibit gene expression levels related to the regulation of bacterial virulence to reduce bacterial pathogenicity; | [143] | |
| Arabidopsis thaliana | Pseudomonas syringae | Induce systemic, acquired resistance; | [138] | |
| Glycine max | Phytophthora sojae | Deposit under the cell wall, then form physical barriers and mechanically strengthen the plant, preventing interaction between P. sojae and the plant; | [145] | |
| Capsicum annuum L. | Colletotrichum sp. | Suppress the growth and germination of Colletotrichum sp. conidia; | [144] | |
| Avena sativa L. | Puccinia graminis f. sp. avenae | Ameliorate photosynthetic parameters affected by fungus and enhance plant resistance; | [146] | |
| Oryza sativa L. | Magnaporthe grisea | Stimulate plant immunity, establish plant resistance responses; | [147] | |
| Brassica napus | Plasmodiophora brassicae | Reverse the transcript-level changes caused by the pathogen and induce transcript-level changes related to plant antioxidant activity; | [148] | |
| Lycopersicon esculentum Mill. | Alternaria solani | Improve gene transcript levels related to JA and SA; increase antioxidative enzyme activity; | [149] | |
| Brachypodium distachyon | Helicoverpa armigera | Reduce herbivore damage, synchronized with the mandible wear of herbivores; | [151] | |
| Eleusine coracana Gaertn. | Sesamiainferens Walker. | Increase the mechanical strength of plants and shorten the pink stem borer feeding tunnel length; | [152] | |
| Oryza sativa L. | Pyricularia oryzae | Enhance plant growth and enhance plant defense to pathogens; | [159] | |
| Nickel (Ni) | Thlaspi caerulescens | Pseudomonas syringae | Inhibit the growth of Pseudomonas syringae; | [104] |
| Streptanthus polygaloides | Tetranychus urticae | Reduce the survival of herbivores and interrupt the growth of whiteflies; | [160] | |
| Vicia faba L. Fabaceae | Bean yellow mosaic virus | Reduce disease severity and increase plant growth indices; induce plant antioxidant responses; | [163] | |
| Titanium (Ti) | Vicia faba L. Fabaceae | broad bean stain virus | Reduce disease severity and induce the expression of genes involved in the SA signaling pathway; | [162] |
| Arabidopsis thaliana | Botrytis cinerea | Increase the transcript level of the phytoalexin-deficient gene and improve plant defense; | [164] | |
| Phytolacca americana | Spodoptera litura | Enhance plant defense against herbivore; inhibit herbivore growth; | [99] | |
| Manganese (Mn) | Betula pendula Roth | Epirrita autumnata | Improve the resistance of silver birch to autumnal moths and reduce pupal weight and immunity; | [165] |
| Boron (B) | Arabidopsis thaliana | Botrytis cinerea | Improve root elongation, induce plant defense, and reduce the proportion of lesions in leaves | [166] |
4. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Oldroyd, G.E.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, 6486. [Google Scholar] [CrossRef] [PubMed]
- Gondal, A.H.; Hussain, I.; Ijaz, A.B.; Zafar, A.; Ch, B.I.; Zafar, H.; Sohail, M.D.; Niazi, H.; Touseef, M.; Khan, A.A.; et al. Influence of Soil Ph and Microbes on Mineral Solubility and Plant Nutrition: A Review. J. Agric. Biol. Sci. 2021, 5, 71–81. [Google Scholar]
- Tang, Z.; Wang, H.Q.; Chen, J.; Chang, J.D.; Zhao, F.J. Molecular mechanisms underlying the toxicity and detoxification of trace metals and metalloids in plants. J. Integr. Plant Biol. 2023, 65, 570–593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic stress responses in plants. Nat. Rev. Genet. 2021, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Ma, Y.; Zhu, Y.G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Agrimonti, C.; Lauro, M.; Visioli, G. Smart agriculture for food quality: Facing climate change in the 21st century. Crit. Rev. Food Sci. Nutr. 2020, 61, 971–981. [Google Scholar] [CrossRef]
- Houmani, H.; Corpas, F.J. Can nutrients act as signals under abiotic stress? Plant Physiol. Biochem. 2024, 206, 108313. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant Immunity: From Signaling to Epigenetic Control of Defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- McNaughton, S.J.; Tarrants, J.L. Grass leaf silicification: Natural selection for an inducible defense against herbivores. Proc. Natl. Acad. Sci. USA 1983, 80, 790–791. [Google Scholar] [CrossRef]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Abiotic and Biotic Plant Stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Bhat, J.A.; Siddiqui, M.H.; Rinklebe, J.; Ahmad, P. Integration of silicon and secondary metabolites in plants: A significant association in stress tolerance. J. Exp. Bot. 2020, 71, 6758–6774. [Google Scholar] [CrossRef] [PubMed]
- Barco, B.; Kim, Y.; Clay, N.K. Expansion of a core regulon by transposable elements promotes Arabidopsis chemical diversity and pathogen defense. Nat. Commun. 2019, 10, 3444. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.J.M.; Brooks, R.R. Terrestrial higher plants which hyperaccumulate metallic elements—A review of their distribution, ecology and phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Reeves, R.D.; Baker, A.J.M.; Jaffre, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.W.A.D.; Lima, L.H.V.; Silva, Y.J.A.B.D.; Biondi, C.M. Ultramafic soils and nickel phytomining opportunities: A review. Rev. Bras. Ciência Solo 2022, 46, e0210099. [Google Scholar] [CrossRef]
- Fryzova, R.; Pohanka, M.; Martinkova, P.; Cihlarova, H.; Brtnicky, M.; Hladky, J.; Kynicky, J. Oxidative Stress and Heavy Metals in Plants. Rev. Environ. Contam. Toxicol. 2017, 245, 129–156. [Google Scholar]
- Zhao, F.-J.; Tang, Z.; Song, J.-J.; Huang, X.-Y.; Wang, P. Toxic metals and metalloids: Uptake, transport, detoxification, phytoremediation, and crop improvement for safer food. Mol. Plant. 2022, 15, 27–44. [Google Scholar] [CrossRef]
- Okey-Onyesolu, C.F.; Hassanisaadi, M.; Bilal, M.; Barani, M.; Rahdar, A.; Iqbal, J.; Kyzas, G.Z. Nanomaterials as Nanofertilizers and Nanopesticides: An Overview. ChemistrySelect 2021, 6, 8645–8663. [Google Scholar] [CrossRef]
- Sun, D.; Hussain, H.I.; Yi, Z.; Rookes, J.E.; Kong, L.; Cahill, D.M. Mesoporous silica nanoparticles enhance seedling growth and photosynthesis in wheat and lupin. Chemosphere 2016, 152, 81–91. [Google Scholar] [CrossRef]
- Lowry, G.V.; Avellan, A.; Gilbertson, L.M. Opportunities and challenges for nanotechnology in the agri-tech revolution. Nat. Nanotechnol. 2019, 14, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Zain, M.; Ma, H.; Ur Rahman, S.; Nuruzzaman, M.; Chaudhary, S.; Azeem, I.; Mehmood, F.; Duan, A.; Sun, C. Nanotechnology in precision agriculture: Advancing towards sustainable crop production. Plant Physiol. Biochem. 2024, 206, 108244. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, R.; Kumar, L.; Mukerjee, N.; Anand, U.; Dhasmana, A.; Preetam, S.; Bhaumik, S.; Sihi, S.; Pal, S.; Khare, T.; et al. The emergence of metal oxide nanoparticles (NPs) as a phytomedicine: A two-facet role in plant growth, nano-toxicity and anti-phyto-microbial activity. Biomed. Pharmacother. 2022, 155, 113658. [Google Scholar] [CrossRef] [PubMed]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed priming with zinc oxide nanoparticles downplayed ultrastructural damage and improved photosynthetic apparatus in maize under cobalt stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef] [PubMed]
- Stanton, C.; Sanders, D.; Krämer, U.; Podar, D. Zinc in plants: Integrating homeostasis and biofortification. Mol. Plant. 2022, 15, 65–85. [Google Scholar] [CrossRef]
- Tan, L.; Qu, M.; Zhu, Y.; Peng, C.; Wang, J.; Gao, D.; Chen, C. ZINC TRANSPORTER5 and ZINC TRANSPORTER9 Function Synergistically in Zinc/Cadmium Uptake. Plant Physiol. 2020, 183, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Hussain, D.; Haydon, M.J.; Wang, Y.; Wong, E.; Sherson, S.M.; Young, J.; Camakaris, J.; Harper, J.F.; Cobbett, C.S. P-type ATPase heavy metal transporters with roles in essential zinc homeostasis in Arabidopsis. Plant Cell 2004, 16, 1327–1339. [Google Scholar] [CrossRef]
- Du, J.; Zeng, J.; Ming, X.; He, Q.; Tao, Q.; Jiang, M.; Gao, S.; Li, X.; Lei, T.; Pan, Y.; et al. The presence of zinc reduced cadmium uptake and translocation in Cosmos bipinnatus seedlings under cadmium/zinc combined stress. Plant Physiol. Biochem. 2020, 151, 223–232. [Google Scholar] [CrossRef]
- Ali, S.; Bani Mfarrej, M.F.; Hussain, A.; Akram, N.A.; Rizwan, M.; Wang, X.; Maqbool, A.; Nafees, M.; Ali, B. Zinc fortification and alleviation of cadmium stress by application of lysine chelated zinc on different varieties of wheat and rice in cadmium stressed soil. Chemosphere 2022, 295, 133829. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Noureen, S.; Anwar, S.; Ali, B.; Naveed, M.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Combined use of biochar and zinc oxide nanoparticle foliar spray improved the plant growth and decreased the cadmium accumulation in rice (Oryza sativa L.) plant. Environ. Sci. Pollut. Res. 2019, 26, 11288–11299. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, X.; Zhang, J.; Li, J.; Niu, T.; Tang, C.; Wang, C.; Xie, J. Zinc oxide nanoparticles improve lettuce (Lactuca sativa L.) plant tolerance to cadmium by stimulating antioxidant defense, enhancing lignin content and reducing the metal accumulation and translocation. Front. Plant Sci. 2022, 13, 1015745. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Bhat, J.A.; Hessini, K.; Yu, F.; Ahmad, P. Zinc oxide nanoparticles alleviates the adverse effects of cadmium stress on Oryza sativa via modulation of the photosynthesis and antioxidant defense system. Ecotoxicol. Environ. Saf. 2021, 220, 112401. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, C.; Du, B.; Cui, H.; Fan, X.; Zhou, D.; Zhou, J. Effects of zinc application on cadmium (Cd) accumulation and plant growth through modulation of the antioxidant system and translocation of Cd in low- and high-Cd wheat cultivars. Environ. Pollut. 2020, 265, 115045. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Faizan, M.; Bhat, M.A.; Huang, F.; Yu, D.; Ahmad, A.; Bajguz, A.; Ahmad, P. Defense interplay of the zinc-oxide nanoparticles and melatonin in alleviating the arsenic stress in soybean (Glycine max L.). Chemosphere 2022, 288, 132471. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Hasanuzzaman, M.; Ding, Y.; Barker, J.; Mokhberdoran, F.; Liu, G. Zinc Oxide Nanoparticles Improve Pleioblastus pygmaeus Plant Tolerance to Arsenic and Mercury by Stimulating Antioxidant Defense and Reducing the Metal Accumulation and Translocation. Front. Plant Sci. 2022, 13, 841501. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.N.; Rasheed, R.; Ashraf, M.Y.; Ashraf, M.A.; Hussain, I. Exogenously applied zinc and copper mitigate salinity effect in maize (Zea mays L.) by improving key physiological and biochemical attributes. Environ. Sci. Pollut. Res. Int. 2018, 25, 23883–23896. [Google Scholar] [CrossRef] [PubMed]
- Ul Aibdin, Z.; Nafees, M.; Rizwan, M.; Ahmad, S.; Ali, S.; Obaid, W.A.; Alsubeie, M.S.; Darwish, D.B.E.; Abeed, A.H.A. Combined effect of Zinc lysine and biochar on growth and physiology of wheat (Triticum aestivum L.) to alleviate salinity stress. Front. Plant Sci. 2023, 13, 1017282. [Google Scholar] [CrossRef]
- Faran, M.; Farooq, M.; Rehman, A.; Nawaz, A.; Saleem, M.K.; Ali, N.; Siddique, K.H.M. High intrinsic seed Zn concentration improves abiotic stress tolerance in wheat. Plant Soil. 2019, 437, 195–213. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Rizwan, M.; Ali, S.; Hassan, M.J.; Brestic, M.; Zhang, X.; Huang, L. Effects of silicon on heavy metal uptake at the soil-plant interphase: A review. Ecotoxicol. Environ. Saf. 2021, 222, 112510. [Google Scholar] [CrossRef]
- Wang, B.; Chu, C.; Wei, H.; Zhang, L.; Ahmad, Z.; Wu, S.; Xie, B. Ameliorative effects of silicon fertilizer on soil bacterial community and pakchoi (Brassica chinensis L.) grown on soil contaminated with multiple heavy metals. Environ. Poll. 2020, 267, 115411. [Google Scholar] [CrossRef]
- Hodson, M.J.; Evans, D.E.; Dietz, K.-J. Aluminium–silicon interactions in higher plants: An update. J. Exp. Bot. 2020, 71, 6719–6729. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Song, Z.; Li, Q.; Ellam, R.M.; Tang, J.; Wang, Y.; Sarkar, B.; Wang, H. Accumulation and partitioning of toxic trace metal(loid)s in phytoliths of wheat grown in a multi-element contaminated soil. Environ. Pollut. 2022, 294, 118645. [Google Scholar] [CrossRef]
- Guerriero, G.; Hausman, J.-F.; Legay, S. Silicon and the Plant Extracellular Matrix. Front. Plant Sci. 2016, 7, 463. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Liang, Y. Silicon prevents aluminum from entering root tip by promoting formation of root border cells in rice. Plant Physiol. Biochem. 2022, 175, 12–22. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, A.; Saleh, A.M.; Habeeb, T.H.; Hassan, Y.M.; Zrieq, R.; Wadaan, M.A.M.; Hozzein, W.N.; Selim, S.; Matos, M.; AbdElgawad, H. Silicon dioxide nanoparticles ameliorate the phytotoxic hazards of aluminum in maize grown on acidic soil. Sci. Total Environ. 2019, 693, 133636. [Google Scholar] [CrossRef]
- Zhang, Z.; Fu, D.; Xie, D.; Wang, Z.; Zhao, Y.; Ma, X.; Huang, P.; Ju, C.; Wang, C. CBL1/9–CIPK23–NRAMP1 axis regulates manganese toxicity. New Phytologist. 2023, 239, 660–672. [Google Scholar] [CrossRef]
- O’Neal, S.L.; Zheng, W. Manganese Toxicity Upon Overexposure: A Decade in Review. Curr. Environ. Health Rep. 2015, 2, 315–328. [Google Scholar] [CrossRef]
- Che, J.; Yamaji, N.; Shao, J.F.; Ma, J.F.; Shen, R.F. Silicon decreases both uptake and root-to-shoot translocation of manganese in rice. J. Exp. Bot. 2016, 67, 1535–1544. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Alam, M.M.; Nahar, K.; Mohsin, S.M.; Bhuyan, M.; Parvin, K.; Hawrylak-Nowak, B.; Fujita, M. Silicon-induced antioxidant defense and methylglyoxal detoxification works coordinately in alleviating nickel toxicity in Oryza sativa L. Ecotoxicology 2019, 28, 261–276. [Google Scholar] [CrossRef]
- Gaur, S.; Kumar, J.; Prasad, S.M.; Sharma, S.; Bhat, J.A.; Sahi, S.; Singh, V.P.; Tripathi, D.K.; Chauhan, D.K. Silicon and nitric oxide interplay alleviates copper induced toxicity in mung bean seedlings. Plant Physiol. Biochem. 2021, 167, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Kumarathilaka, P.; Seneweera, S.; Meharg, A.; Bundschuh, J. Arsenic speciation dynamics in paddy rice soil-water environment: Sources, physico-chemical, and biological factors-A review. Water Res. 2018, 140, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Tai, F.; Wang, S.; Liang, B.; Li, Y.; Wu, J.; Fan, C.; Hu, X.; Wang, H.; He, R.; Wang, W. Quaternary ammonium iminofullerenes improve root growth of oxidative-stress maize through ASA-GSH cycle modulating redox homeostasis of roots and ROS-mediated root-hair elongation. J. Nanobiotechnol. 2022, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon Nanoparticles More Efficiently Alleviate Arsenate Toxicity than Silicon in Maize Cultiver and Hybrid Differing in Arsenate Tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef]
- Bist, V.; Anand, V.; Srivastava, S.; Kaur, J.; Naseem, M.; Mishra, S.; Srivastava, P.K.; Tripathi, R.D.; Srivastava, S. Alleviative mechanisms of silicon solubilizing Bacillus amyloliquefaciens mediated diminution of arsenic toxicity in rice. J. Hazard. Mater. 2022, 428, 128170. [Google Scholar] [CrossRef]
- Ur Rahman, S.; Xuebin, Q.; Kamran, M.; Yasin, G.; Cheng, H.; Rehim, A.; Riaz, L.; Rizwan, M.; Ali, S.; Alsahli, A.A.; et al. Silicon elevated cadmium tolerance in wheat (Triticum aestivum L.) by endorsing nutrients uptake and antioxidative defense mechanisms in the leaves. Plant Physiol. Biochem. 2021, 166, 148–159. [Google Scholar] [CrossRef]
- Vaculík, M.; Landberg, T.; Greger, M.; Luxová, M.; Stoláriková, M.; Lux, A. Silicon modifies root anatomy, and uptake and subcellular distribution of cadmium in young maize plants. Ann. Bot. 2012, 110, 433–443. [Google Scholar] [CrossRef]
- Rizwan, M.; Meunier, J.D.; Davidian, J.C.; Pokrovsky, O.S.; Bovet, N.C.K. Silicon alleviates Cd stress of wheat seedlings (Triticum turgidum L. cv. Claudio) grown in hydroponics. Environ. Sci. Pollut. Res. Int. 2016, 23, 1414–1427. [Google Scholar] [CrossRef]
- Ahmed, T.; Masood, H.A.; Noman, M.; Al-Huqail, A.A.; Alghanem, S.M.S.; Khan, M.M.; Muhammad, S.; Manzoor, N.; Rizwan, M.; Qi, X.; et al. Biogenic silicon nanoparticles mitigate cadmium (Cd) toxicity in rapeseed (Brassica napus L.) by modulating the cellular oxidative stress metabolism and reducing Cd translocation. J. Hazard. Mater. 2023, 459, 132070. [Google Scholar] [CrossRef]
- Ulhassan, Z.; Yang, S.; He, D.; Khan, A.R.; Salam, A.; Azhar, W.; Muhammad, S.; Ali, S.; Hamid, Y.; Khan, I.; et al. Seed priming with nano-silica effectively ameliorates chromium toxicity in Brassica napus. J. Hazard. Mater. 2023, 458, 131906. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Rai, P.; Kandhol, N.; Kumar, A.; Sahi, S.; Corpas, F.J.; Sharma, S.; Singh, V.P. Silicon palliates chromium toxicity through the formation of root hairs in rice (Oryza sativa) mediated by GSH and IAA. Plant Cell Physiol. 2023, 63, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Rai, P.; Prakash, V.; Tripathi, S.; Tiwari, K.; Gahlawat, N.; Tripathi, D.K.; Sharma, S. Ameliorative effects of Si-SNP synergy to mitigate chromium induced stress in Brassica juncea. Environ. Pollut. 2023, 335, 122031. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Mei, Y.; Nikolic, N.; Nikolic, M.; Li, T.; Peng, H.; Liang, Y. From promoting aggregation to enhancing obstruction: A negative feedback regulatory mechanism of alleviation of trivalent chromium toxicity by silicon in rice. J. Hazard. Mater. 2023, 457, 131720. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Yasin, N.A.; Akram, K.; Ahmad, A.; Khan, W.U.; Akram, W.; Akbar, M. Ameliorative role of FBL-10 and silicon against lead induced stress in Solanum melongena. Plant Physiol. Biochem. 2021, 158, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, R.; Mur, L.A.J.; Ruan, J.; Shen, Q.; Guo, S. Functions of silicon in plant drought stress responses. Hortic. Res. 2021, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Wang, X.; Abbas, T.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Irfan, M.; Butt, M.; Wahid, M.A.; Cheema, M.; et al. Combined application of zinc and silicon alleviates terminal drought stress in wheat by triggering morpho-physiological and antioxidants defense mechanisms. PLoS ONE 2021, 16, e0256984. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.; Hu, Y.; Mukherjee, S.; Aftab, T.; Fahad, S.; Raza, A.; Ahmad, M.; Siddiqui, M.H. Silicon-induced postponement of leaf senescence is accompanied by modulation of antioxidative defense and ion homeostasis in mustard (Brassica juncea) seedlings exposed to salinity and drought stress. Plant Physiol. Biochem. 2020, 157, 47–59. [Google Scholar] [CrossRef]
- Calero Hurtado, A.; Chiconato, D.A.; Prado, R.M.; Sousa Junior, G.D.S.; Gratao, P.L.; Felisberto, G.; Olivera Viciedo, D.; Mathias Dos Santos, D.M. Different methods of silicon application attenuate salt stress in sorghum and sunflower by modifying the antioxidative defense mechanism. Ecotoxicol. Environ. Saf. 2020, 203, 110964. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Keya, S.S.; Abdelrahman, M.; Miah, M.G.; Phan Tran, L.S. Silicon in mitigation of abiotic stress-induced oxidative damage in plants. Crit. Rev. Biotechnol. 2021, 41, 918–934. [Google Scholar] [CrossRef]
- Wu, X.; Riaz, M.; Yan, L.; Du, C.; Liu, Y.; Jiang, C. Boron Deficiency in Trifoliate Orange Induces Changes in Pectin Composition and Architecture of Components in Root Cell Walls. Front Plant Sci. 2017, 8, 1882. [Google Scholar] [CrossRef]
- Yan, L.; Riaz, M.; Liu, J.; Liu, Y.; Zeng, Y.; Jiang, C. Boron reduces aluminum deposition in alkali-soluble pectin and cytoplasm to release aluminum toxicity. J. Hazard Mater. 2021, 401, 123388. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Yan, L.; Wu, X.; Hussain, S.; Aziz, O.; Jiang, C. Boron deprivation induced inhibition of root elongation is provoked by oxidative damage, root injuries and changes in cell wall structure. Environ. Exp. Bot. 2018, 156, 74–85. [Google Scholar] [CrossRef]
- Wu, X.; Lu, X.; Riaz, M.; Yan, L.; Jiang, C. Boron toxicity induced specific changes of cell ultrastructure and architecture of components in leaf center and tip of trifoliate orange [Poncirus trifoliata (L.) Raf.]. J. Environ. Manag. 2019, 246, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sanchez, F.; Simon-Grao, S.; Martinez-Nicolas, J.J.; Alfosea-Simon, M.; Liu, C.; Chatzissavvidis, C.; Perez-Perez, J.G.; Camara-Zapata, J.M. Multiple stresses occurring with boron toxicity and deficiency in plants. J. Hazard Mater. 2020, 397, 122713. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Riaz, M.; Wu, X.; Du, C.; Liu, Y.; Jiang, C. Ameliorative effects of boron on aluminum induced variations of cell wall cellulose and pectin components in trifoliate orange (Poncirus trifoliate (L.) Raf.) Rootstock. Environ. Pollut. 2018, 240, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Kamran, M.; Fang, Y.; Yang, G.; Rizwan, M.; Ali, S.; Zhou, Y.; Wang, Q.; Deng, L.; Wang, Y.; et al. Boron supply alleviates cadmium toxicity in rice (Oryza sativa L.) by enhancing cadmium adsorption on cell wall and triggering antioxidant defense system in roots. Chemosphere 2021, 266, 128938. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Song, H.; Guan, C.; Zhang, Z. Boron mitigates cadmium toxicity to rapeseed (Brassica napus) shoots by relieving oxidative stress and enhancing cadmium chelation onto cell walls. Environ. Pollut. 2020, 263, 114546. [Google Scholar] [CrossRef]
- Shireen, F.; Nawaz, M.A.; Lu, J.; Xiong, M.; Kaleem, M.; Huang, Y.; Bie, Z. Application of boron reduces vanadium toxicity by altering the subcellular distribution of vanadium, enhancing boron uptake and enhancing the antioxidant defense system of watermelon. Ecotoxicol. Environ. Saf. 2021, 226, 112828. [Google Scholar] [CrossRef]
- Shireen, F.; Nawaz, M.A.; Xiong, M.; Ahmad, A.; Sohail, H.; Chen, Z.; Abouseif, Y.; Huang, Y.; Bie, Z. Pumpkin rootstock improves the growth and development of watermelon by enhancing uptake and transport of boron and regulating the gene expression. Plant Physiol. Biochem. 2020, 154, 204–218. [Google Scholar] [CrossRef]
- Alharby, H.F.; Nahar, K.; Al-Zahrani, H.S.; Hakeem, K.R.; Hasanuzzaman, M. Enhancing Salt Tolerance in Soybean by Exogenous Boron: Intrinsic Study of the Ascorbate-Glutathione and Glyoxalase Pathways. Plants 2021, 10, 2085. [Google Scholar] [CrossRef]
- Lu, K.; Yan, L.; Riaz, M.; Babar, S.; Hou, J.; Zhang, Y.; Jiang, C. Exogenous boron alleviates salt stress in cotton by maintaining cell wall structure and ion homeostasis. Plant Physiol. Biochem. 2023, 201, 107858. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant nanobionics approach to augment photosynthesis and biochemical sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Tito, N.; Giraldo, J.P. Anionic Cerium Oxide Nanoparticles Protect Plant Photosynthesis from Abiotic Stress by Scavenging Reactive Oxygen Species. ACS Nano 2017, 11, 11283–11297. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, X.; Yue, L.; Wang, C.; Tao, M.; Wang, Z.; Xing, B. Foliar-applied cerium oxide nanomaterials improve maize yield under salinity stress: Reactive oxygen species homeostasis and rhizobacteria regulation. Environ. Pollut. 2022, 299, 118900. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Zhang, W.; Ma, X. Cerium oxide nanoparticles alter the salt stress tolerance of Brassica napus L. by modifying the formation of root apoplastic barriers. Environ. Pollut. 2017, 229, 132–138. [Google Scholar]
- Wang, Y.; Wang, L.; Ma, C.; Wang, K.; Hao, Y.; Chen, Q.; Mo, Y.; Rui, Y. Effects of cerium oxide on rice seedlings as affected by co-exposure of cadmium and salt. Environ. Pollut. 2019, 252, 1087–1096. [Google Scholar] [CrossRef]
- Trela-Makowej, A.; Orzechowska, A.; Szymańska, R. Less is more: The hormetic effect of titanium dioxide nanoparticles on plants. Sci. Total Environ. 2024, 910, 168669. [Google Scholar] [CrossRef]
- Lai, M.; Ghouri, F.; Sarwar, S.; Alomrani, S.O.; Riaz, M.; Haider, F.U.; Liu, J.; Imran, M.; Ali, S.; Liu, X.; et al. Modulation of metal transporters, oxidative stress and cell abnormalities by synergistic application of silicon and titanium oxide nanoparticles: A strategy for cadmium tolerance in rice. Chemosphere 2023, 345, 140439. [Google Scholar] [CrossRef]
- Iqbal, A.; Mo, Z.; Pan, S.G.; Qi, J.Y.; Hua, T.; Imran, M.; Duan, M.; Gu, Q.; Yao, X.B.; Tang, X. Exogenous TiO(2) Nanoparticles Alleviate Cd Toxicity by Reducing Cd Uptake and Regulating Plant Physiological Activity and Antioxidant Defense Systems in Rice (Oryza sativa L.). Metabolites 2023, 13, 765. [Google Scholar] [CrossRef]
- Kumar, D.; Dhankher, O.P.; Tripathi, R.D.; Seth, C.S. Titanium dioxide nanoparticles potentially regulate the mechanism(s) for photosynthetic attributes, genotoxicity, antioxidants defense machinery, and phytochelatins synthesis in relation to hexavalent chromium toxicity in Helianthus annuus L. J. Hazard Mater. 2023, 454, 131418. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, P.; Yadu, B.; Korram, J.; Satnami, M.L.; Kumar, M.; Keshavkant, S. Titanium nanoparticles attenuates arsenic toxicity by up-regulating expressions of defensive genes in Vigna radiata L. J. Environ. Sci. 2020, 92, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Ghouri, F.; Shahid, M.J.; Zhong, M.; Zia, M.A.; Alomrani, S.O.; Liu, J.; Sun, L.; Ali, S.; Liu, X.; Shahid, M.Q. Alleviated lead toxicity in rice plant by co-augmented action of genome doubling and TiO2 nanoparticles on gene expression, cytological and physiological changes. Sci. Total Environ. 2024, 911, 168709. [Google Scholar] [CrossRef] [PubMed]
- Kiany, T.; Pishkar, L.; Sartipnia, N.; Iranbakhsh, A.; Barzin, G. Effects of silicon and titanium dioxide nanoparticles on arsenic accumulation, phytochelatin metabolism, and antioxidant system by rice under arsenic toxicity. Environ. Sci. Pollut. Res. Int. 2022, 29, 34725–34737. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; AlSolami, M.A.; Basahi, R.A.; Siddiqui, M.H.; Al-Huqail, A.A.; Abbas, Z.K.; Siddiqui, Z.H.; Ali, H.M.; Khan, F. Nitric oxide is involved in nano-titanium dioxide-induced activation of antioxidant defense system and accumulation of osmolytes under water-deficit stress in Vicia faba L. Ecotoxicol. Environ. Saf. 2020, 190, 110152. [Google Scholar] [CrossRef]
- Lashkary, M.; Moghaddam, M.; Asgharzade, A.; Tatari, M. Titanium dioxide nanoparticle is involved in mitigating NaCl-induced Calendula officinalis L. by activation of antioxidant defense system and accumulation of osmolytes. Plant Physiol. Biochem. 2021, 166, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Amini, S.; Maali-Amiri, R.; Mohammadi, R.; Kazemi-Shahandashti, S.S. cDNA-AFLP analysis of transcripts induced in chickpea plants by TiO(2) nanoparticles during cold stress. Plant Physiol. Biochem. 2017, 111, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, N.E.; Abdelhameed, R.M. Use of titanium dioxide doped multi-wall carbon nanotubes as promoter for the growth, biochemical indices of Sesamum indicum L. under heat stress conditions. Plant Physiol. Biochem. 2023, 201, 107844. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, C.; Xiong, Y.; Xiao, F.; Wang, Y. Heavy metal induced resistance to herbivore of invasive plant: Implications from inter- and intraspecific comparisons. Front. Plant Sci. 2023, 14, 1222867. [Google Scholar] [CrossRef]
- Khan, M.M.; Fan, Z.-Y.; Wang, X.-M.; Qiu, B.-L. Distribution and accumulation of Cadmium in different trophic levels affecting Serangium japonicum, the predatory beetle of whitefly Bemisia tabaci, biologically, physiologically and genetically: An experimental study. J. Hazard. Mater. 2024, 465, 133244. [Google Scholar] [CrossRef]
- Kramer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Cappa, J.J.; Pilon-Smits, E.A.H. Evolutionary aspects of elemental hyperaccumulation. Planta 2013, 239, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Asad, S.A.; Ahmad, R.M.S. Heavy metals and glucosinolates based defence mechanisms in metal accumulating plants and counter adaptations by insect herbivores: A review. Int. J. Agric. Biol. 2018, 20, 811–820. [Google Scholar]
- Dangl, J.L.; Fones, H.; Davis, C.A.R.; Rico, A.; Fang, F.; Smith, J.A.C.; Preston, G.M. Metal Hyperaccumulation Armors Plants against Disease. PLoS Pathog. 2010, 6, e1001093. [Google Scholar]
- Gurajala, H.K.; Cao, X.; Tang, L.; Ramesh, T.M.; Lu, M.; Yang, X. Comparative assessment of Indian mustard (Brassica juncea L.) genotypes for phytoremediation of Cd Pb Contam. Soils. Environ. Pollut. 2019, 254, 113085. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.P.; Kumari, P.; Rai, P.K. Current Status of the Disease-Resistant Gene(s)/QTLs, and Strategies for Improvement in Brassica juncea. Front. Plant Sci. 2021, 12, 617405. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Z.; Zeng, C.; Dong, X.; Li, M.; Liu, Z.; Yan, M. The elemental defense effect of cadmium on Alternaria brassicicola in Brassica juncea. BMC Plant Biol. 2022, 22, 17. [Google Scholar] [CrossRef]
- Dai, Z.C.; Cai, H.H.; Qi, S.S.; Li, J.; Zhai, D.L.; Wan, J.S.H.; Du, D.L. Cadmium hyperaccumulation as an inexpensive metal armor against disease in Crofton weed. Environ. Poll. 2020, 267, 115649. [Google Scholar] [CrossRef]
- Lin, T.; He, W.; Yang, M.; Wang, X.; Vrieling, K.; Chen, G. Soil cadmium pollution facilitated the invasion of alligator weed through enhanced herbivore resistance and competitive ability over a congeneric species. Plant Cell Environ. 2023, 47, 585–599. [Google Scholar] [CrossRef]
- Pottier, M.; Garcia de la Torre, V.S.; Victor, C.; David, L.C.; Chalot, M.; Thomine, S. Genotypic variations in the dynamics of metal concentrations in poplar leaves: A field study with a perspective on phytoremediation. Environ. Pollut. 2015, 199, 73–82. [Google Scholar] [CrossRef]
- Lin, T.; Lu, Q.; Zheng, Z.; Li, S.; Li, S.; Liu, Y.; Zhu, T.; Chen, L.; Yang, C.; Han, S. Soil cadmium stress affects the phyllosphere microbiome and associated pathogen resistance differently in male and female poplars. J. Exp. Bot. 2023, 74, 2188–2202. [Google Scholar] [CrossRef] [PubMed]
- Plaza, S.; Weber, J.; Pajonk, S.; Thomas, J.; Talke, I.N.; Schellenberg, M.; Pradervand, S.; Burla, B.; Geisler, M.; Martinoia, E.; et al. Wounding of Arabidopsis halleri leaves enhances cadmium accumulation that acts as a defense against herbivory. BioMetals 2015, 28, 521–528. [Google Scholar] [CrossRef]
- Kazemi-Dinan, A.; Thomaschky, S.; Stein, R.J.; Kramer, U.; Muller, C. Zinc and cadmium hyperaccumulation act as deterrents towards specialist herbivores and impede the performance of a generalist herbivore. New Phytol. 2014, 202, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Godinho, D.P.; Serrano, H.C.; Magalhães, S.; Branquinho, C. Concurrent herbivory and metal accumulation: The outcome for plants and herbivores. Plant-Environ. Interact. 2022, 3, 170–178. [Google Scholar] [CrossRef]
- Stolpe, C.; Giehren, F.; Krämer, U.; Müller, C. Both heavy metal-amendment of soil and aphid-infestation increase Cd and Zn concentrations in phloem exudates of a metal-hyperaccumulating plant. Phytochemistry 2017, 139, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Chen, J.; Zhou, S.; Yu, W.; Chen, G.; Chen, L.; Wang, X.; Shi, H.; Han, S.; Zhang, F. Testing the elemental defense hypothesis with a woody plant species: Cadmium accumulation protects Populus yunnanensis from leaf herbivory and pathogen infection. Chemosphere 2020, 247, 125851. [Google Scholar] [CrossRef]
- Lin, T.; Zhu, G.; He, W.; Xie, J.; Li, S.; Han, S.; Li, S.; Yang, C.; Liu, Y.; Zhu, T. Soil cadmium stress reduced host plant odor selection and oviposition preference of leaf herbivores through the changes in leaf volatile emissions. Sci. Total Environ. 2022, 814, 152728. [Google Scholar] [CrossRef]
- Qin, S.; Wu, Z.; Tang, J.; Zhu, G.; Chen, G.; Chen, L.; Lei, H.; Wang, X.; Zhu, T.; Lin, T. Effects of exogenous spermidine on poplar resistance to leaf and root herbivory as affected by soil cadmium stress. J. Environ. Manag. 2021, 288, 112467. [Google Scholar] [CrossRef]
- Chen, J.; Qin, S.; Tang, J.; Chen, G.; Xie, J.; Chen, L.; Han, S.; Wang, X.; Zhu, T.; Liu, Y.; et al. Exogenous nitrogen enhances poplar resistance to leaf herbivory and pathogen infection after exposure to soil cadmium stress. Ecotoxicol. Environ. Saf. 2021, 208, 111688. [Google Scholar] [CrossRef]
- Jiang, D.; Wang, G.R.; Yan, S.C. The improved resistance against gypsy moth in Larix olgensis seedlings exposed to Cd stress association with elemental and chemical defenses. Pest. Manag. Sci. 2020, 76, 1713–1721. [Google Scholar] [CrossRef]
- Glawischnig, E. Camalexin. Phytochem 2007, 68, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Martos, S.; Gallego, B.; Cabot, C.; Llugany, M.; Barcelo, J.; Poschenrieder, C. Zinc triggers signaling mechanisms and defense responses promoting resistance to Alternaria brassicicola in Arabidopsis thaliana. Plant Sci. 2016, 249, 13–24. [Google Scholar] [CrossRef]
- Escudero, V.; Ferreira Sánchez, D.; Abreu, I.; Sopeña-Torres, S.; Makarovsky-Saavedra, N.; Bernal, M.; Krämer, U.; Grolimund, D.; González-Guerrero, M.; Jordá, L.; et al. Arabidopsis thaliana Zn2+—Efflux ATPases HMA2 and HMA4 are required for resistance to the necrotrophic fungus Plectosphaerella cucumerina BMM. J. Exp. Bot. 2022, 73, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Awan, Z.A.; Shoaib, A.; Khan, K.A. Crosstalk of Zn in Combination with Other Fertilizers Underpins Interactive Effects and Induces Resistance in Tomato Plant against Early Blight Disease. Plant Pathol. J. 2019, 35, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Huq, M.A.; Apu, M.A.I.; Ashrafudoulla, M.; Rahman, M.M.; Parvez, M.A.K.; Balusamy, S.R.; Akter, S.; Rahman, M.S. Bioactive ZnO Nanoparticles: Biosynthesis, Characterization and Potential Antimicrobial Applications. Pharmaceutics 2023, 15, 2634. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Xiao, Q.; Li, Q.; Xia, Y.; Feng, H.; Ma, X.; Cai, L.; Sun, X. Antimicrobial mechanisms of ZnO nanoparticles to phytopathogen Pseudomonas syringae: Damage of cell envelope, suppression of metabolism, biofilm and motility, and stimulation of stomatal immunity on host plant. Pestic. Biochem. Physiol. 2023, 194, 105455. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qian, C.; Jiang, H.; Liu, S.; Yang, D.; Cui, J. Visible-Light-Driven Zinc Oxide Quantum Dots for the Management of Bacterial Fruit Blotch Disease and the Improvement of Melon Seedlings Growth. J. Agric. Food Chem. 2023, 71, 2773–2783. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, T.; Li, B.; Wu, Y.; Li, Z. Bio-Fabrication of ZnONPs from Alkalescent Nucleoside Antibiotic to Control Rice Blast: Impact on Pathogen (Magnaporthe grisea) and Host (Rice). Int. J. Mol. Sci. 2023, 24, 2778. [Google Scholar] [CrossRef]
- Dhiman, S.; Varma, A.; Rao, M.; Prasad, R.; Goel, A. Deciphering the fertilizing and disease suppression potential of phytofabricated zinc oxide nanoparticles on Brassica juncea. Environ. Res. 2023, 231, 116276. [Google Scholar] [CrossRef]
- Kah, M.; Tufenkji, N.; White, J.C. Nano-enabled strategies to enhance crop nutrition and protection. Nat. Nanotechnol. 2019, 14, 532–540. [Google Scholar] [CrossRef]
- Valdes, C.; Cota-Ruiz, K.; Flores, K.; Ye, Y.; Hernandez-Viezcas, J.A.; Gardea-Torresdey, J.L. Antioxidant and defense genetic expressions in corn at early-developmental stage are differentially modulated by copper form exposure (nano, bulk, ionic): Nutrient and physiological effects. Ecotoxicol. Environ. Saf. 2020, 206, 111197. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hu, Q.; Huang, Y.; Fulton, A.N.; Hannah-Bick, C.; Adeleye, A.S.; Keller, A.A. Activation of antioxidant and detoxification gene expression in cucumber plants exposed to a Cu(OH)2 nanopesticide. Environ. Sci. Nano 2017, 4, 1750–1760. [Google Scholar] [CrossRef]
- Elmer, W.H.; White, J.C. The use of metallic oxide nanoparticles to enhance growth of tomatoes and eggplants in disease infested soil or soilless medium. Environ. Sci. Nano 2016, 3, 1072–1079. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Manikandan, A. Application of Copper-Chitosan Nanoparticles Stimulate Growth and Induce Resistance in Finger Millet (Eleusine coracana Gaertn.) Plants Against Blast Disease. J. Agric. Food Chem. 2018, 66, 1784–1790. [Google Scholar] [CrossRef]
- Mondal, K.K.; Mani, C. Investigation of the antibacterial properties of nanocopper against Xanthomonas axonopodis pv. punicae, the incitant of pomegranate bacterial blight. Ann. Microbiol. 2011, 62, 889–893. [Google Scholar] [CrossRef]
- Cheah, B.H.; Chuang, W.-P.; Lo, J.-C.; Li, Y.; Cheng, C.-Y.; Yang, Z.-W.; Liao, C.-T.; Lin, Y.-F. Exogenous Copper Application for the Elemental Defense of Rice Plants against Rice Leaffolder (Cnaphalocrocis medinalis). Plants 2022, 11, 1104. [Google Scholar] [CrossRef] [PubMed]
- Naidu, S.; Pandey, J.; Mishra, L.C.; Chakraborty, A.; Roy, A.; Singh, I.K.; Singh, A. Silicon nanoparticles: Synthesis, uptake and their role in mitigation of biotic stress. Ecotoxicol. Environ. Saf. 2023, 255, 114783. [Google Scholar] [CrossRef] [PubMed]
- El-Shetehy, M.; Moradi, A.; Maceroni, M.; Reinhardt, D.; Petri-Fink, A.; Rothen-Rutishauser, B.; Mauch, F.; Schwab, F. Silica nanoparticles enhance disease resistance in Arabidopsis plants. Nat. Nanotechnol. 2020, 16, 344–353. [Google Scholar] [CrossRef]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of Silicon on Plant-Pathogen Interactions. Front. Plant Sci. 2017, 8, 701. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Yang, Y. Mechanisms of silicon-induced fungal disease resistance in plants. Plant Physiol. Biochem. 2021, 165, 200–206. [Google Scholar] [CrossRef]
- Peng, H.; Deng, K.; Shi, Y.; Liu, S.; Jian, Z.; Li, C.; Ji, X.; Li, S. Alleviation of Cd-polluted paddy soils through Si fertilizer application and its effects on the soil microbial community. Sci. Total Environ. 2023, 855, 158735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-Y.; Hong, D.-K.; Chen, Y.-H.; Gao, S.-J.; Fu, H.-Y.; Zheng, H.-K.; Fang, Y.; Wang, J.-D. Synergistic Effects of Organosilicon and Cu(OH)2 in Controlling Sugarcane Leaf Scald Disease. Int. J. Mol. Sci. 2022, 23, 13532. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pan, T.; Gao, X.; An, J.; Ning, C.; Li, S.; Cai, K. Silica nanoparticles activate defense responses by reducing reactive oxygen species under Ralstonia solanacearum infection in tomato plants. NanoImpact 2022, 28, 100418. [Google Scholar] [CrossRef] [PubMed]
- Hersanti Siti Choir, W.; Rizkie, L.; Subakti Pu, S.N. Effects of Chitosan and Silica Nanoparticles Against the Development and Growth of Red Chilli Anthracnose Disease Colletotrichum sp. Pak. J. Biol. Sci. 2022, 25, 748–754. [Google Scholar]
- Rasoolizadeh, A.; Labbe, C.; Sonah, H.; Deshmukh, R.K.; Belzile, F.; Menzies, J.G.; Belanger, R.R. Silicon protects soybean plants against Phytophthora sojae by interfering with effector-receptor expression. BMC Plant Biol. 2018, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J.; Lv, P.; Mi, J.; Zhao, B. Silicon improves the photosynthetic performance of oat leaves infected with Puccinia graminis f. sp. avenae. Front. Plant Sci. 2022, 13, 1037136. [Google Scholar] [CrossRef]
- Du, J.; Liu, B.; Zhao, T.; Xu, X.; Lin, H.; Ji, Y.; Li, Y.; Li, Z.; Lu, C.; Li, P.; et al. Silica nanoparticles protect rice against biotic and abiotic stresses. J. Nanobiotechnol. 2022, 20, 197. [Google Scholar] [CrossRef]
- Sarkar, A.; Kisiala, A.; Adhikary, D.; Basu, U.; Emery, R.J.N.; Rahman, H.; Kav, N.N.V. Silicon ameliorates clubroot responses in canola (Brassica napus): A “multi-omics”-based investigation into possible mechanisms. Physiol. Plantarum 2023, 175, e13900. [Google Scholar] [CrossRef]
- Gulzar, N.; Ali, S.; Shah, M.A.; Kamili, A.N. Silicon supplementation improves early blight resistance in Lycopersicon esculentum Mill. by modulating the expression of defense-related genes and antioxidant enzymes. 3 Biotech 2021, 11, 232. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, A.; Hartley, S.; Singh, I.K. Silicon: Its ameliorative effect on plant defense against herbivory. J. Exp. Bot. 2020, 71, 6730–6743. [Google Scholar] [CrossRef]
- Waterman, J.M.; Cibils-Stewart, X.; Cazzonelli, C.I.; Hartley, S.E.; Johnson, S.N. Short-term exposure to silicon rapidly enhances plant resistance to herbivory. Ecology 2021, 102, e03438. [Google Scholar] [CrossRef] [PubMed]
- Jadhao, K.R.; Bansal, A.; Rout, G.R. Silicon amendment induces synergistic plant defense mechanism against pink stem borer (Sesamia inferens Walker.) in finger millet (Eleusine coracana Gaertn.). Sci. Rep. 2020, 10, 4229. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Song, J.; Jeong, B.R. Drenched Silicon Suppresses Disease and Insect Pests in Coffee Plant Grown in Controlled Environment by Improving Physiology and Upregulating Defense Genes. Int. J. Mol. Sci. 2022, 23, 3543. [Google Scholar] [CrossRef]
- Douma, J.C.; Ganzeveld, L.N.; Unsicker, S.B.; Boeckler, G.A.; Dicke, M. What makes a volatile organic compound a reliable indicator of insect herbivory? Plant Cell Environ. 2019, 42, 3308–3325. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, O.L.; Padula, M.P.; Zeng, R.; Gurr, G.M. Silicon: Potential to Promote Direct and Indirect Effects on Plant Defense Against Arthropod Pests in Agriculture. Front. Plant Sci. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed]
- Gurr, G.M.; Kvedaras, O.L. Synergizing biological control: Scope for sterile insect technique, induced plant defences and cultural techniques to enhance natural enemy impact. Biol. Control. 2010, 52, 198–207. [Google Scholar] [CrossRef]
- Allmann, S.; Baldwin, I.T. Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles. Science 2010, 329, 1075–1078. [Google Scholar] [CrossRef]
- Ye, M.; Song, Y.; Long, J.; Wang, R.; Baerson, S.R.; Pan, Z.; Zhu-Salzman, K.; Xie, J.; Cai, K.; Luo, S.; et al. Priming of jasmonate-mediated antiherbivore defense responses in rice by silicon. Proc. Natl. Acad. Sci. USA 2013, 110, E3631–E3639. [Google Scholar] [CrossRef]
- Parthasarathy, R.; Jayabaskaran, C.; Manikandan, A.; Anusuya, S. Synthesis of Nickel-Chitosan Nanoparticles for Controlling Blast Diseases in Asian Rice. Appl. Biochem. Biotechnol. 2022, 195, 2134–2148. [Google Scholar] [CrossRef]
- Jhee, E.M.; Boyd, R.S.; Eubanks, M.D. Nickel hyperaccumulation as an elemental defense of Streptanthus polygaloides (Brassicaceae): Influence of herbivore feeding mode. New Phytologist. 2005, 168, 331–344. [Google Scholar] [CrossRef]
- Cheruiyot, D.J.; Boyd, R.S.; Moar, W. Testing the joint effects hypothesis of elemental defense using Spodoptera exigua. J. Chem. Ecol. 2015, 41, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Elsharkawy, M.M.; Derbalah, A. Antiviral activity of titanium dioxide nanostructures as a control strategy for broad bean strain virus in faba bean. Pest Manag. Sci. 2019, 75, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Sofy, A.R.; Hmed, A.A.; Alnaggar, A.E.M.; Dawoud, R.A.; Elshaarawy, R.F.M.; Sofy, M.R. Mitigating effects of Bean yellow mosaic virus infection in faba bean using new carboxymethyl chitosan-titania nanobiocomposites. Int. J. Biol. Macromol. 2020, 163, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Zavala, F.G.; Atriztán-Hernández, K.; Martínez-Irastorza, P.; Oropeza-Aburto, A.; López-Arredondo, D.; Herrera-Estrella, L. Titanium nanoparticles activate a transcriptional response in Arabidopsis that enhances tolerance to low phosphate, osmotic stress and pathogen infection. Front. Plant Sci. 2022, 13, 994523. [Google Scholar] [CrossRef]
- Ruuhola, T.; Leppanen, T.; Julkunen-Tiitto, R.; Rantala, M.J.; Lehto, T. Boron fertilization enhances the induced defense of silver birch. J. Chem. Ecol. 2011, 37, 460–471. [Google Scholar] [CrossRef]
- Batista-Oliveira, J.S.; Formey, D.; Torres, M.; Aragon, W.; Romero-Contreras, Y.J.; Maruri-Lopez, I.; Tromas, A.; Schwan-Estrada, K.R.F.; Serrano, M. Gadolinium Protects Arabidopsis thaliana against Botrytis cinerea through the Activation of JA/ET-Induced Defense Responses. Int. J. Mol. Sci. 2021, 22, 4938. [Google Scholar] [CrossRef]
- Beig, B.; Niazi, M.B.K.; Jahan, Z.; Haider, G.; Zia, M.; Shah, G.A.; Iqbal, Z.; Hayat, A. Development and testing of zinc sulfate and zinc oxide nanoparticle-coated urea fertilizer to improve N and Zn use efficiency. Front. Plant Sci. 2023, 13, 1058219. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Liu, Z.; Song, Y.; Sui, J.; Hua, X. Advances in the Involvement of Metals and Metalloids in Plant Defense Response to External Stress. Plants 2024, 13, 313. https://doi.org/10.3390/plants13020313
Zhang L, Liu Z, Song Y, Sui J, Hua X. Advances in the Involvement of Metals and Metalloids in Plant Defense Response to External Stress. Plants. 2024; 13(2):313. https://doi.org/10.3390/plants13020313
Chicago/Turabian StyleZhang, Lingxiao, Zhengyan Liu, Yun Song, Junkang Sui, and Xuewen Hua. 2024. "Advances in the Involvement of Metals and Metalloids in Plant Defense Response to External Stress" Plants 13, no. 2: 313. https://doi.org/10.3390/plants13020313
APA StyleZhang, L., Liu, Z., Song, Y., Sui, J., & Hua, X. (2024). Advances in the Involvement of Metals and Metalloids in Plant Defense Response to External Stress. Plants, 13(2), 313. https://doi.org/10.3390/plants13020313