
Metabolic and Molecular Rearrangements of Sauvignon Blanc (Vitis vinifera L.) Berries in Response to Foliar Applications of Specific Dry Yeast
 , ,
, ,  , , , ,
, , , ,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
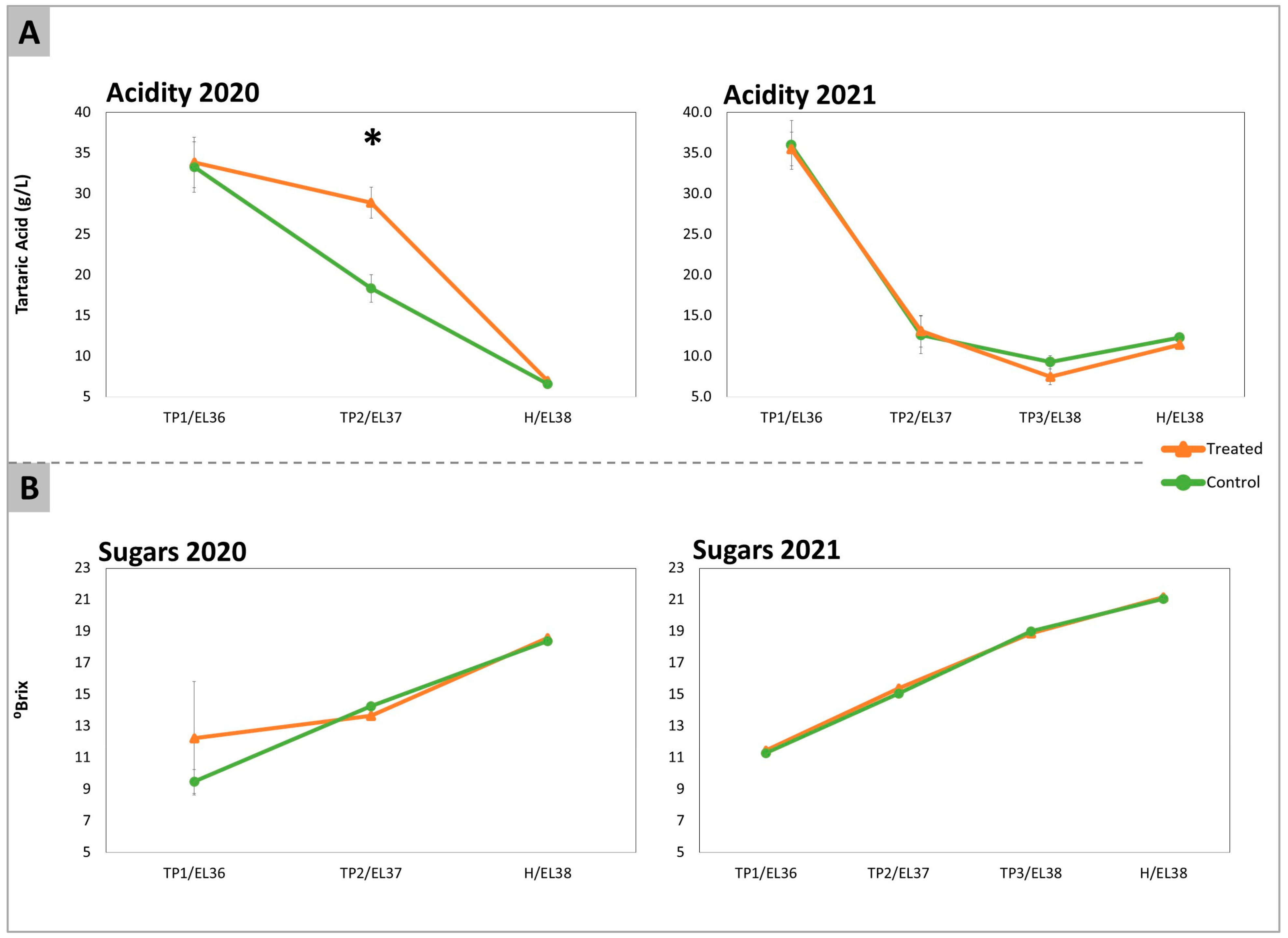
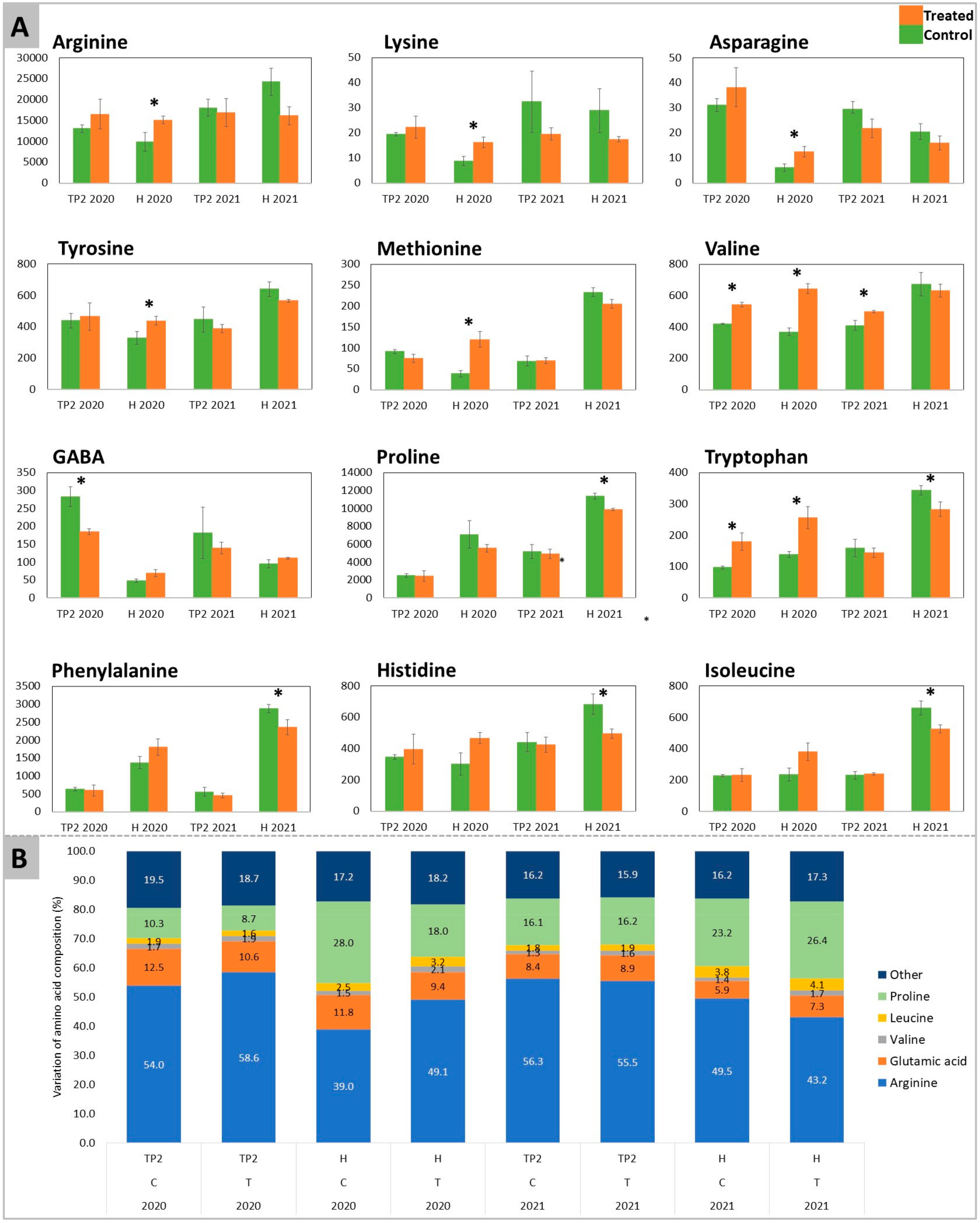
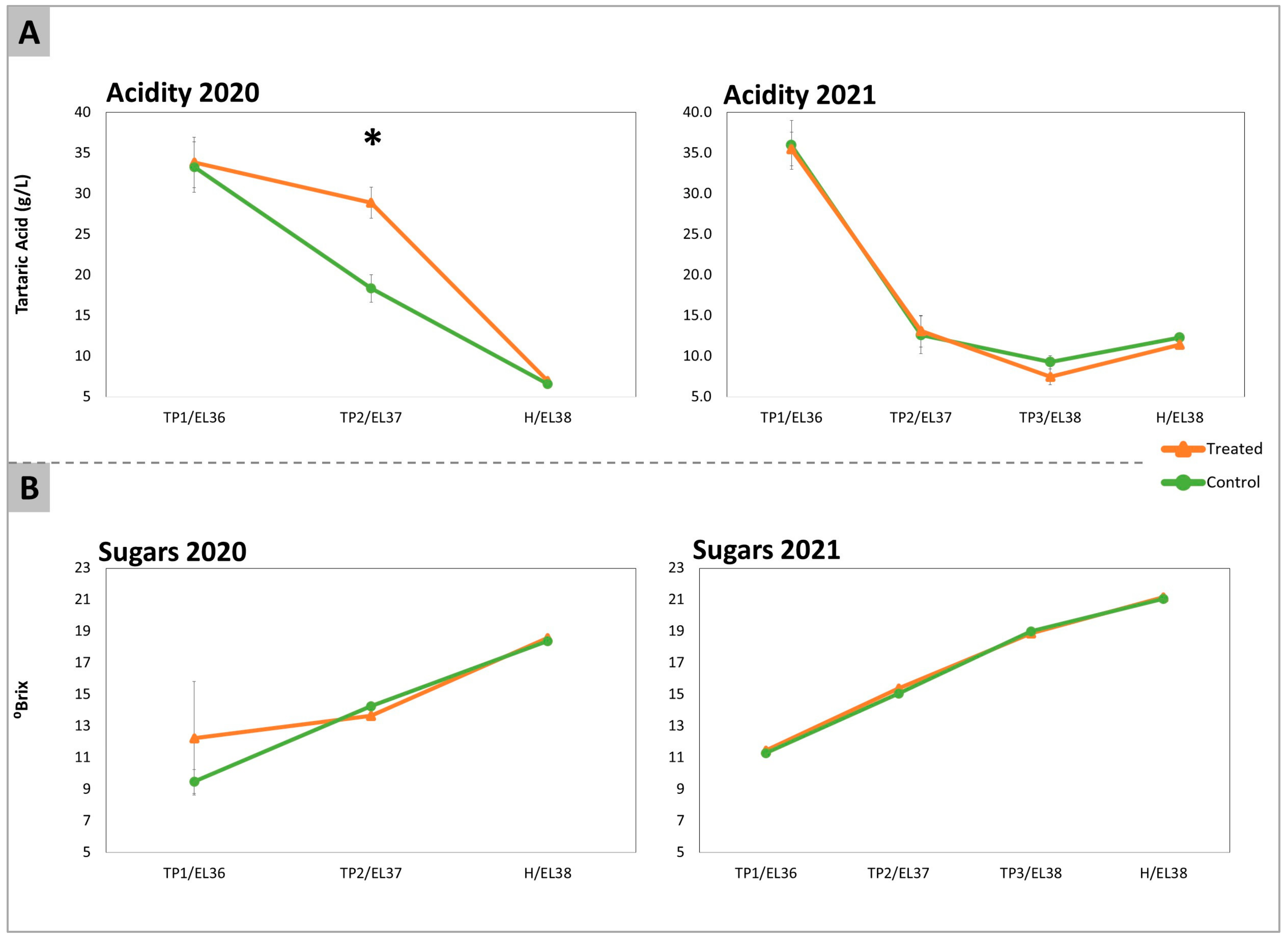
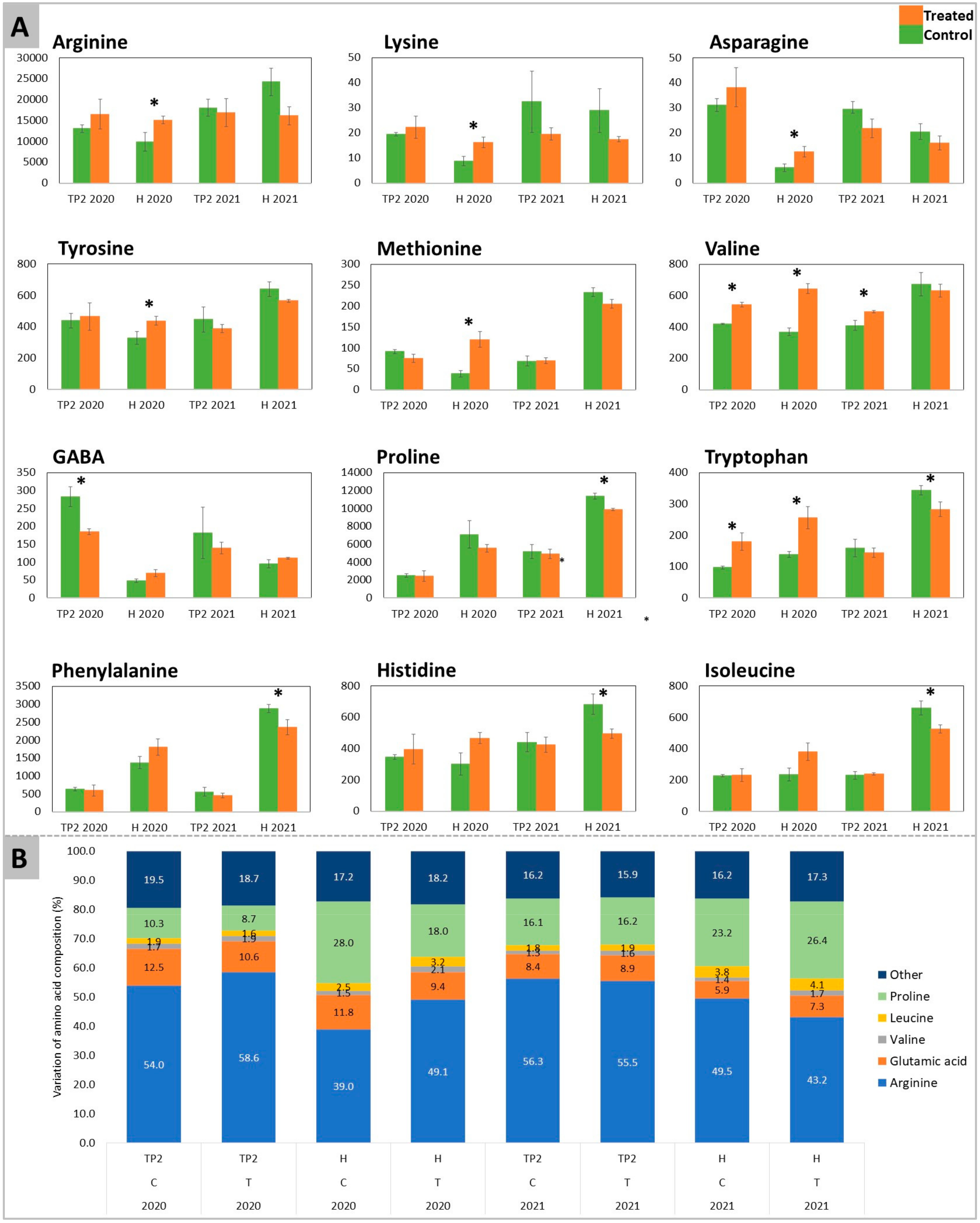
2.1. Acidity, Total Sugars, Amino Acids, and GABA
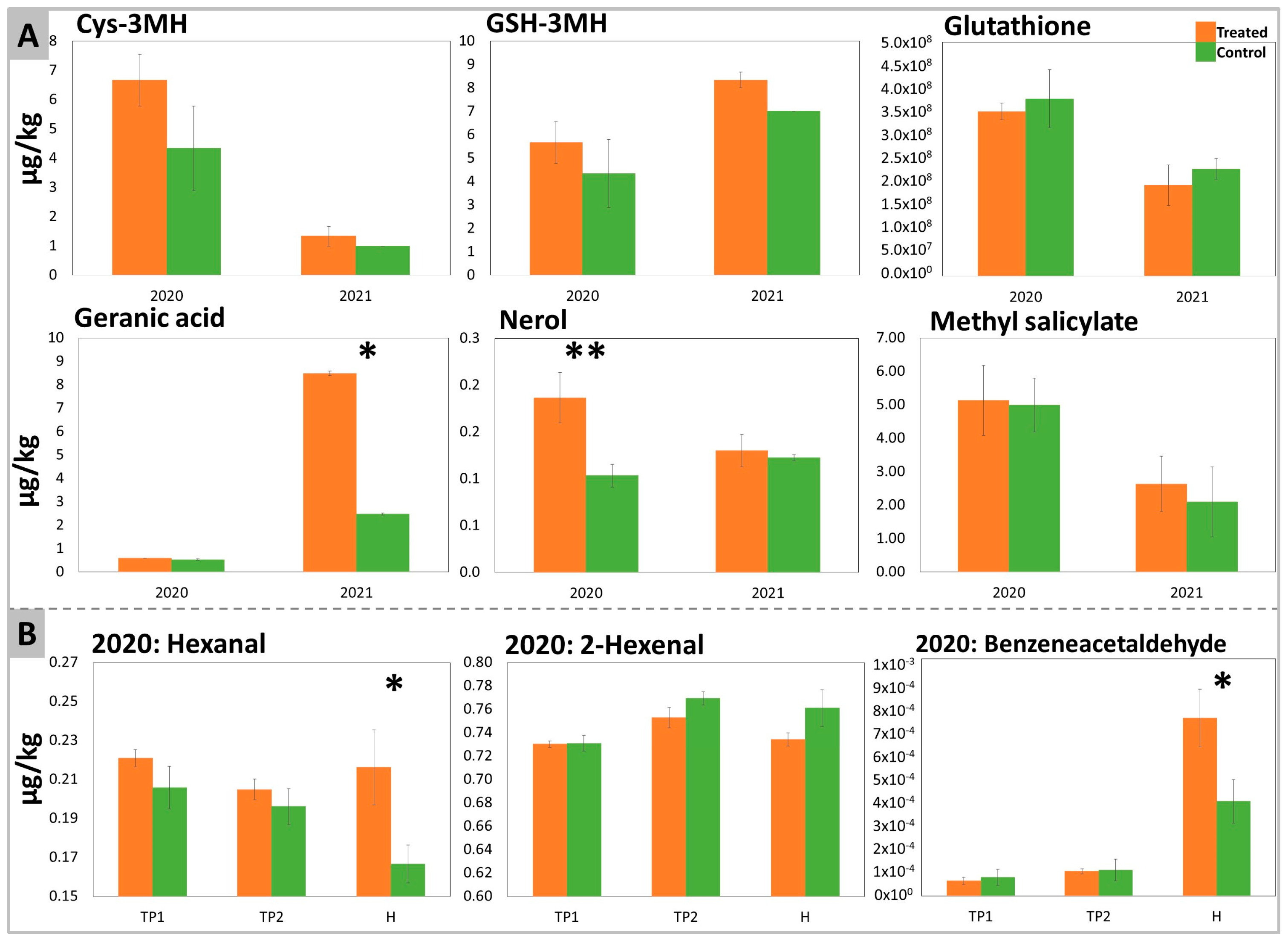
2.2. Aromatic Profile
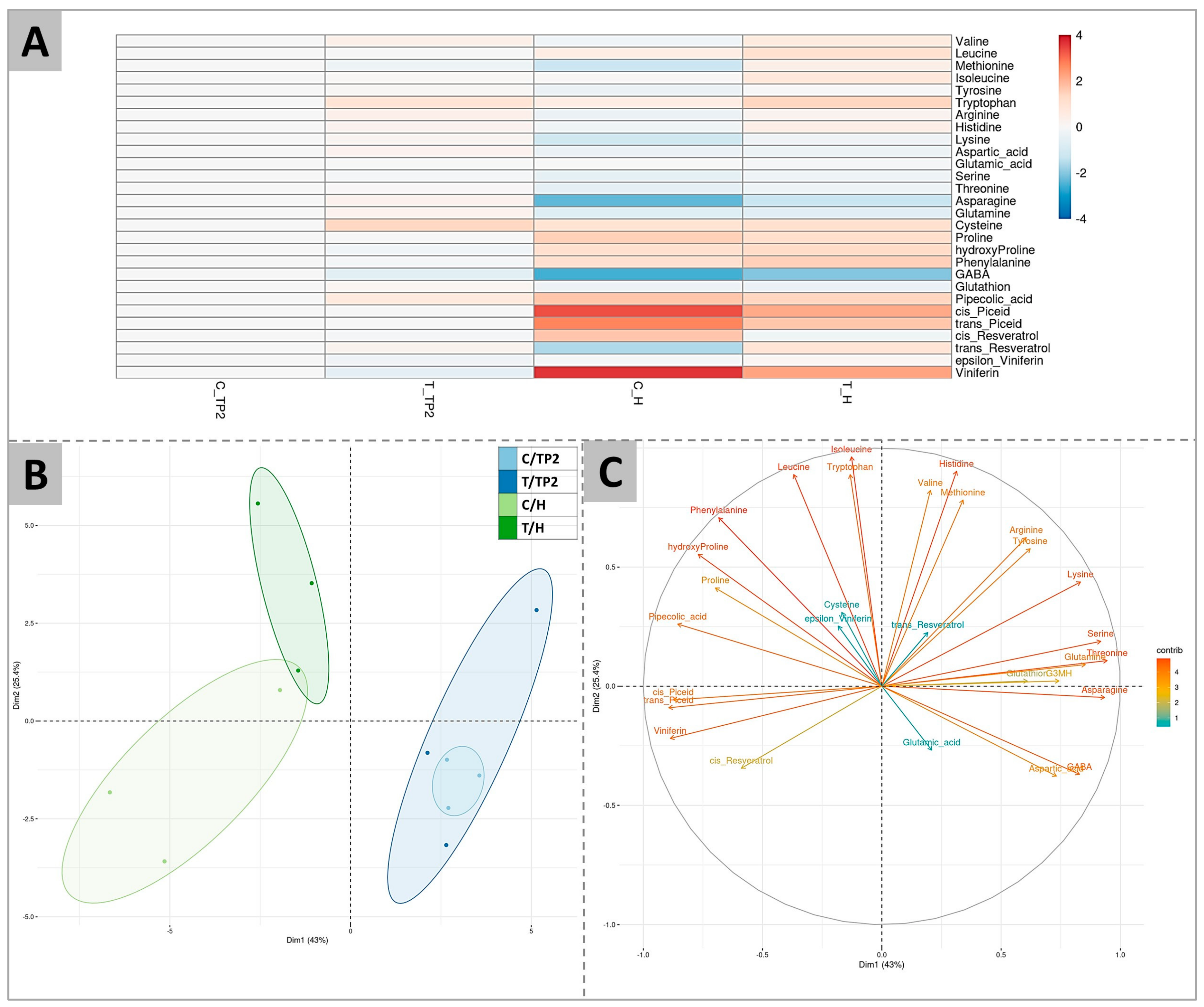
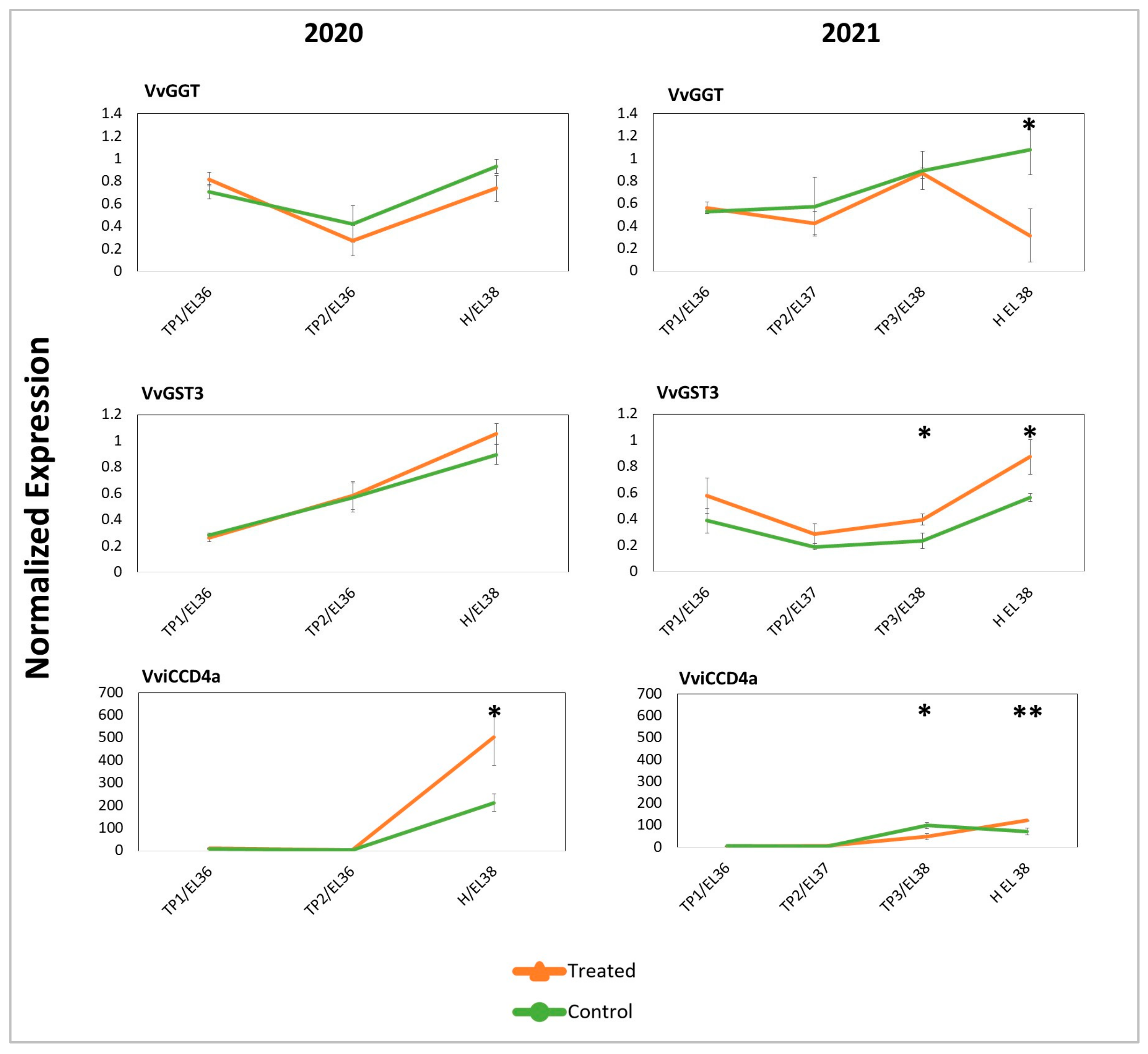
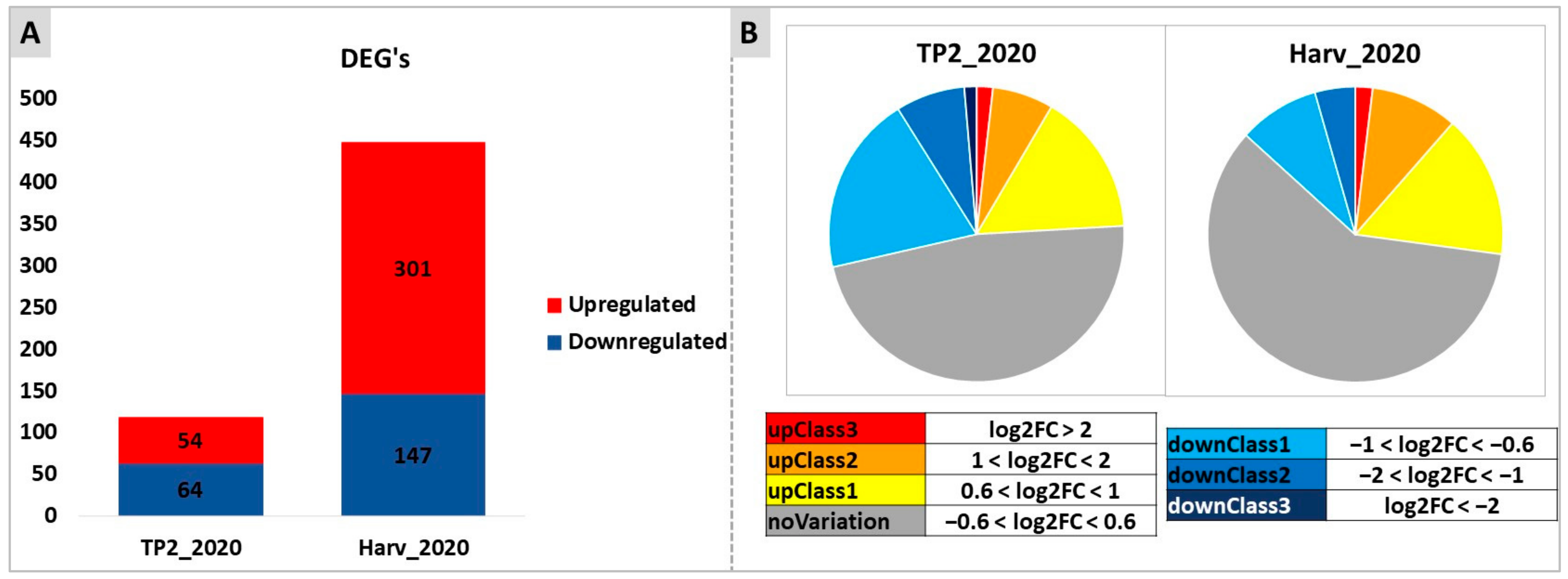
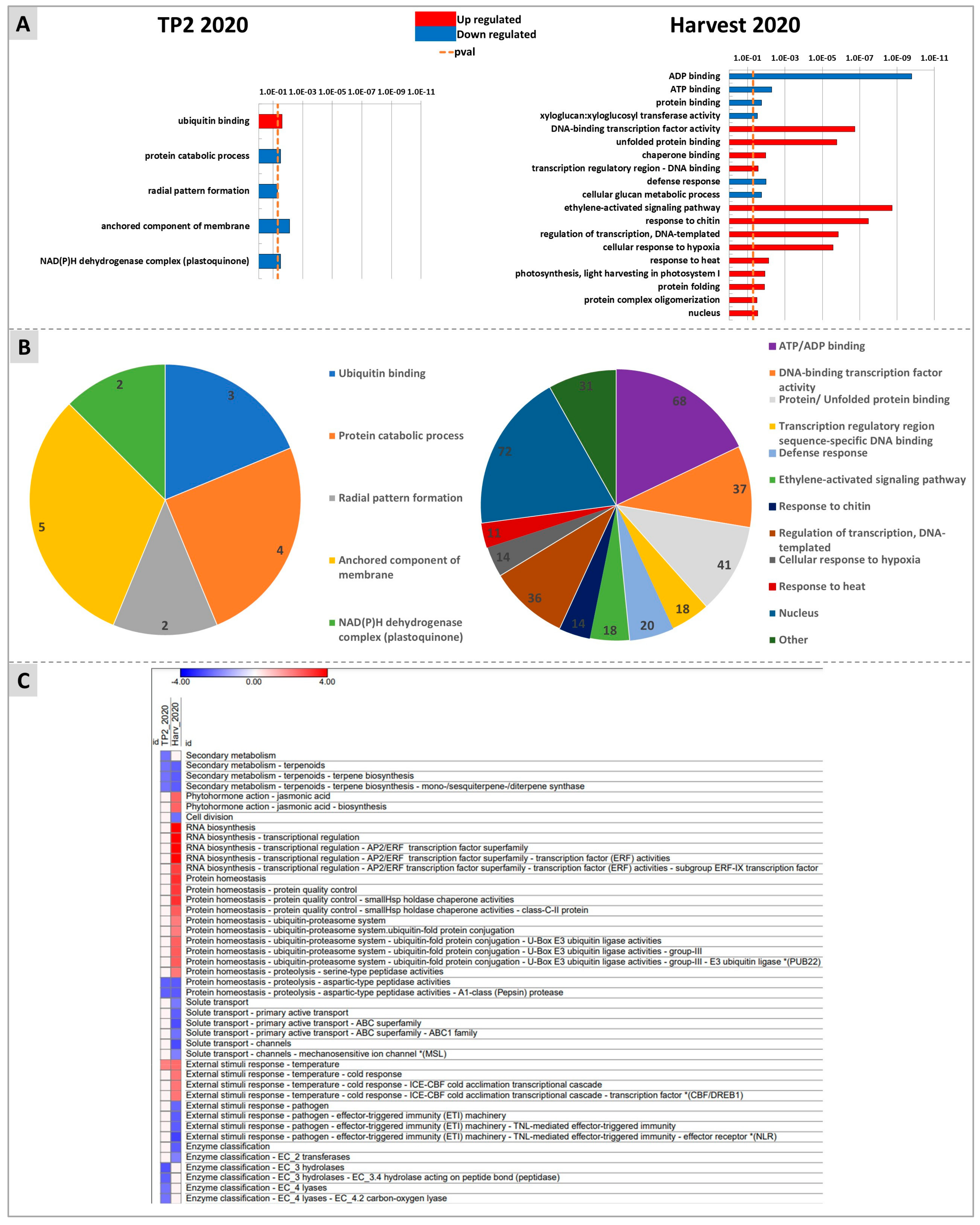
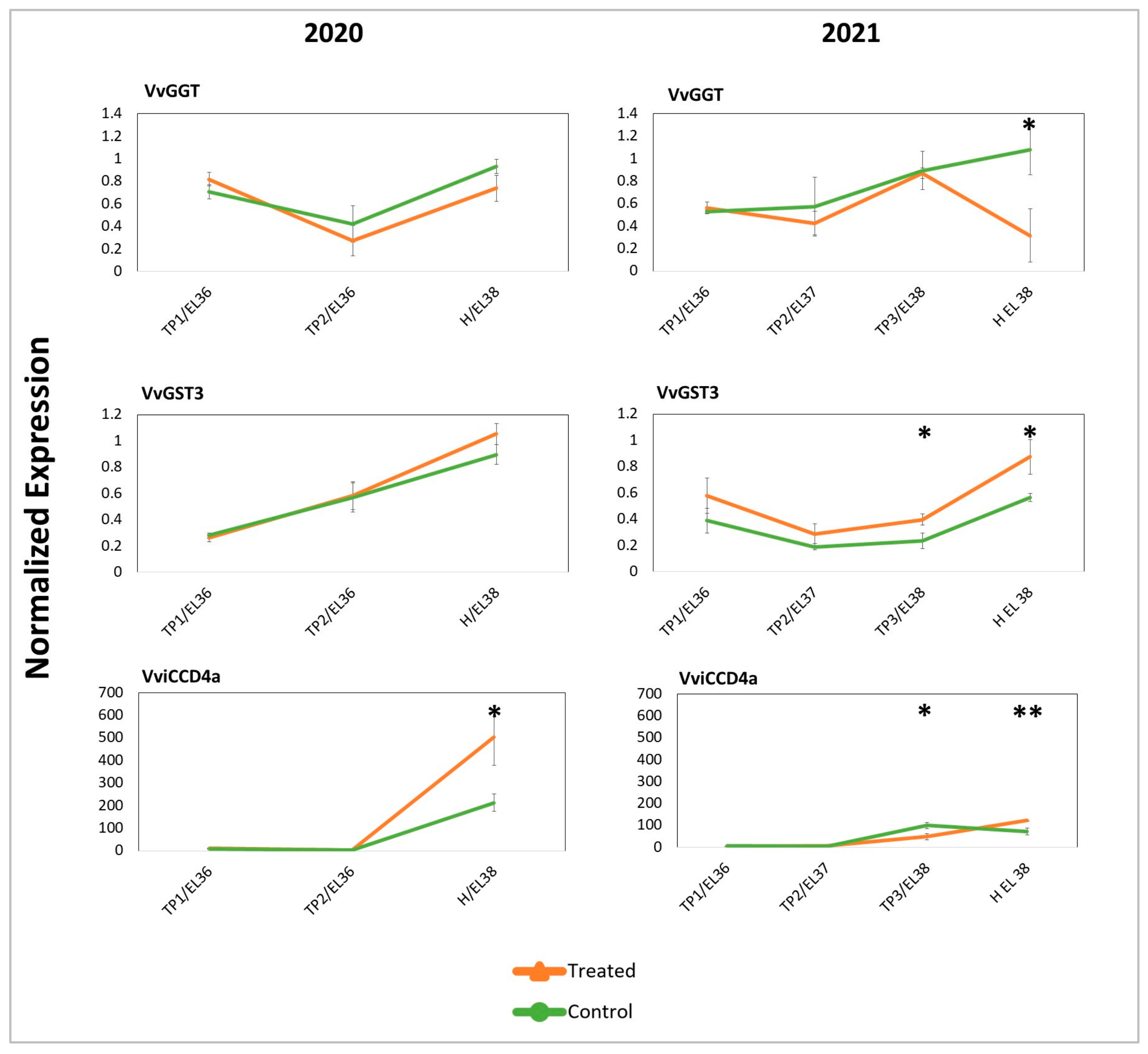
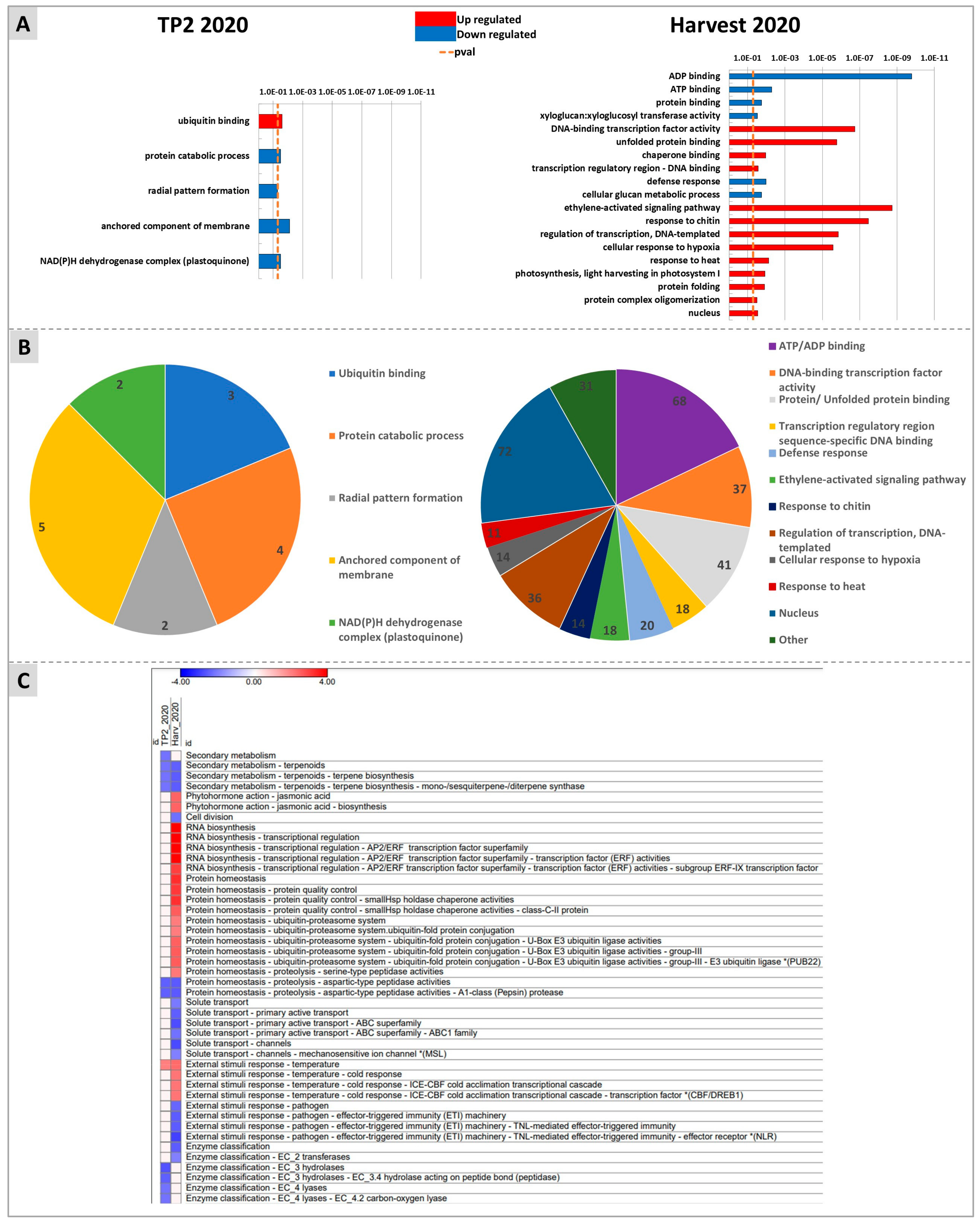
2.3. Targeted and Untargeted Molecular Responses: Comparison of DYE-Treated versus Untreated Berries
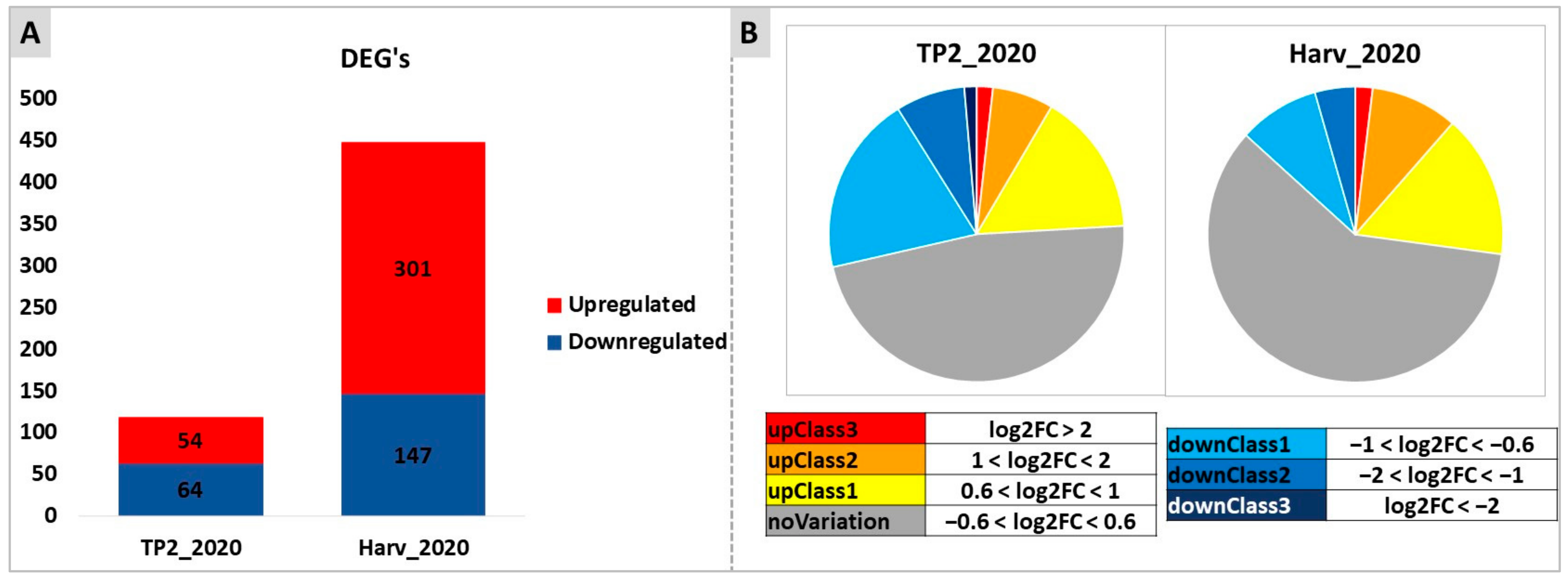
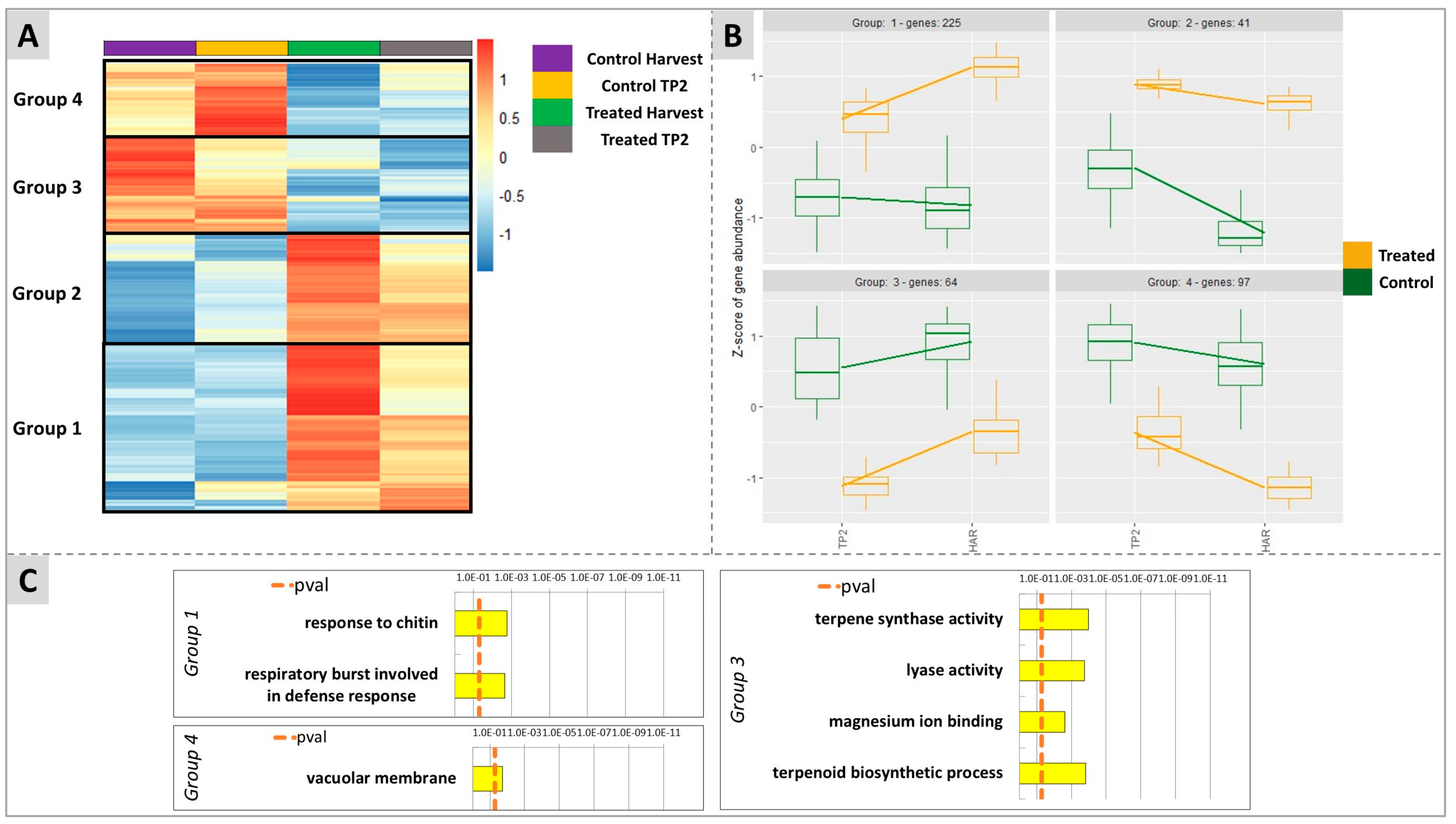
2.4. Untargeted Analysis of Molecular Responses to DYE Application: Combined Effects of Time and Treatment
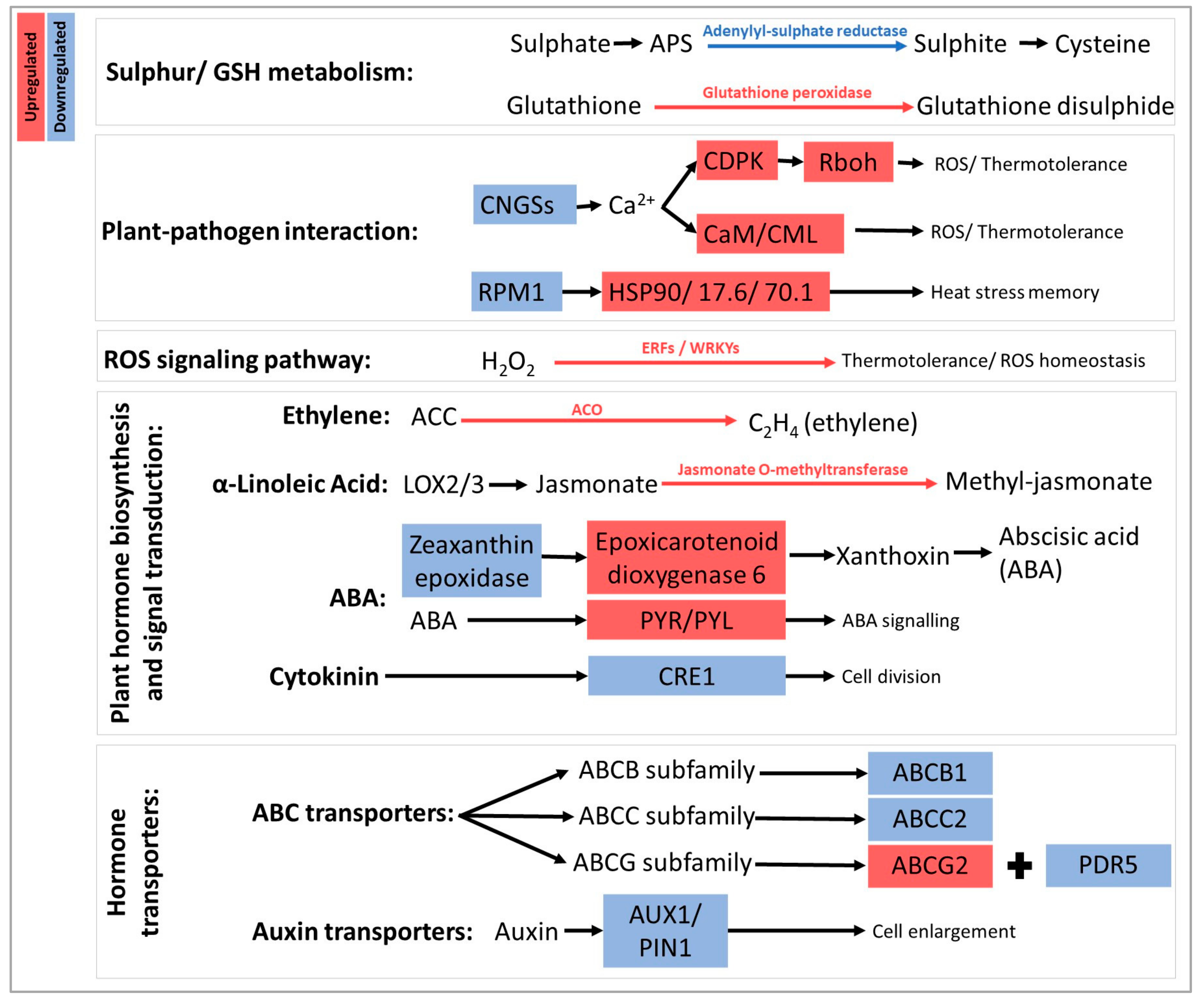
3. Discussion
4. Materials and Methods
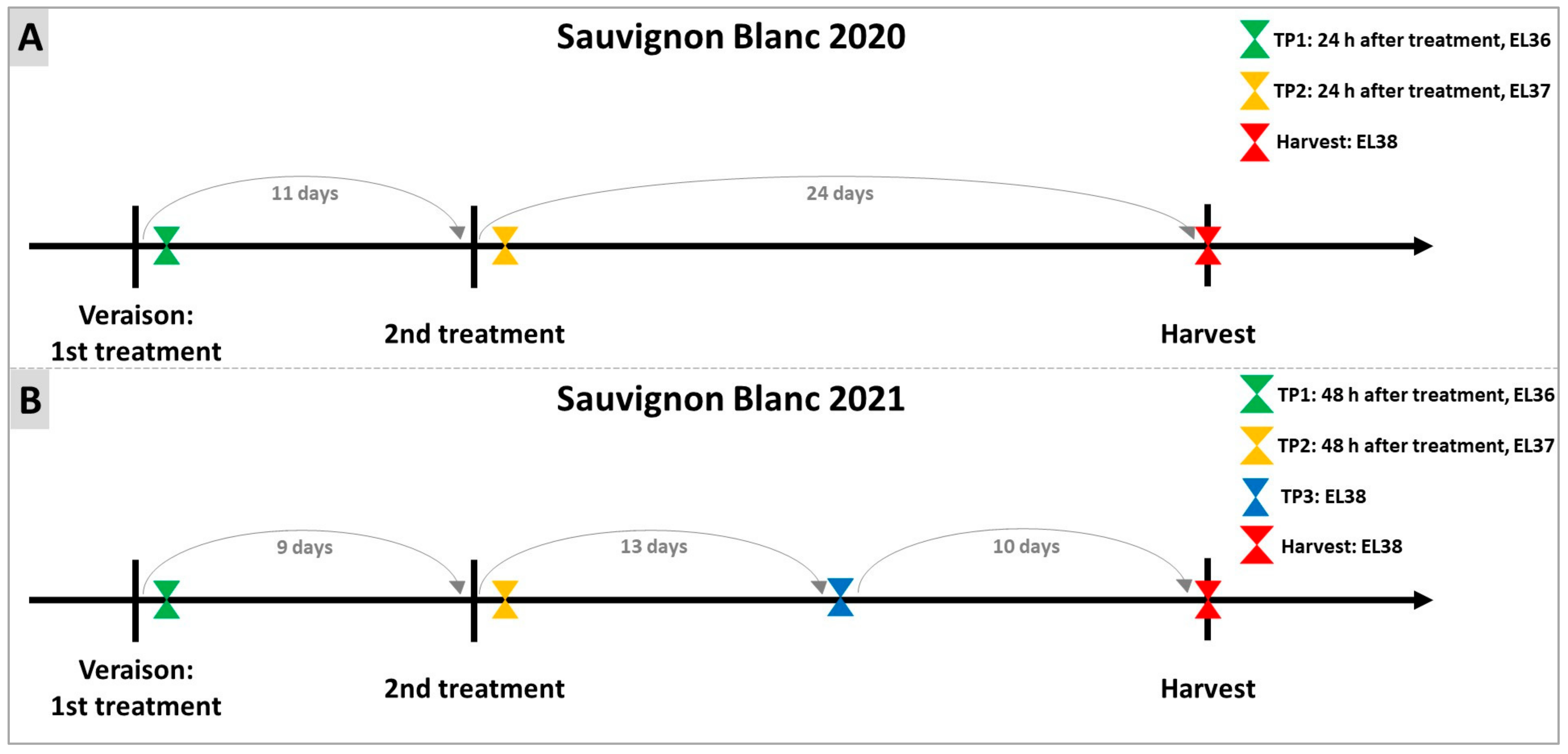
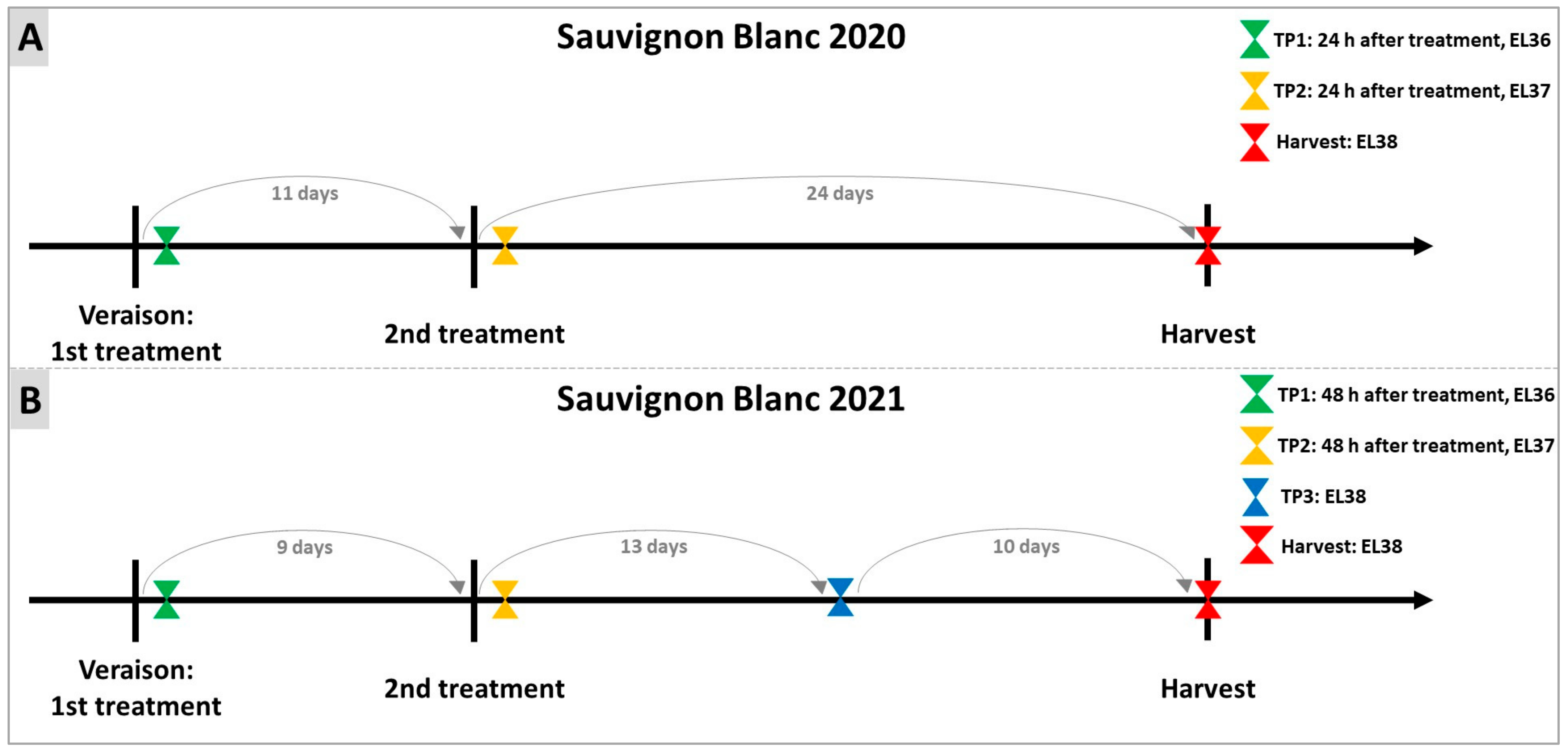
4.1. Experimental Design and Plant Material
4.2. Acidity and Total Sugars
4.3. Aroma Precursors
4.3.1. Free Precursors
4.3.2. Bound Precursors and Thiol Precursors
4.4. Relative Quantification of Amino Acids, GABA, and Secondary Metabolites
4.5. RNA Extraction, cDNA Synthesis and RT-qPCR Expression Analysis
4.6. RNA-Seq Analysis and Data Processing
4.7. Data Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, J.; Massonnet, M.; Cantu, D. The Genetic Basis of Grape and Wine Aroma. Hortic. Res. 2019, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Gattullo, C.E.; Mezzapesa, G.N.; Stellacci, A.M.; Ferrara, G.; Occhiogrosso, G.; Petrelli, G.; Castellini, M.; Spagnuolo, M. Cover Crop for a Sustainable Viticulture: Effects on Soil Properties and Table Grape Production. Agronomy 2020, 10, 1334. [Google Scholar] [CrossRef]
- Pons, A.; Allamy, L.; Schüttler, A.; Rauhut, D.; Thibon, C.; Darriet, P. What Is the Expected Impact of Climate Change on Wine Aroma Compounds and Their Precursors in Grape? Oeno One 2017, 51, 141–146. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Destrac-Irvine, A. Modified Grape Composition under Climate Change Conditions Requires Adaptations in the Vineyard. Oeno One 2016, 51, 147–154. [Google Scholar] [CrossRef]
- Giacosa, S.; Ossola, C.; Botto, R.; Río Segade, S.; Paissoni, M.A.; Pollon, M.; Gerbi, V.; Rolle, L. Impact of Specific Inactive Dry Yeast Application on Grape Skin Mechanical Properties, Phenolic Compounds Extractability, and Wine Composition. Food Res. Int. 2019, 116, 1084–1093. [Google Scholar] [CrossRef]
- Droulia, F.; Charalampopoulos, I. Future Climate Change Impacts on European Viticulture: A Review on Recent Scientific Advances. Atmosphere 2021, 12, 495. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A Review of the Potential Climate Change Impacts and Adaptation Options for European Viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Mira de Orduña, R. Climate Change Associated Effects on Grape and Wine Quality and Production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Crupi, P.; Santamaria, M.; Vallejo, F.; Tomás-Barberán, F.A.; Masi, G.; Caputo, A.R.; Battista, F.; Tarricone, L. How Pre-Harvest Inactivated Yeast Treatment May Influence the Norisoprenoid Aroma Potential in Wine Grapes. Appl. Sci. 2020, 10, 3369. [Google Scholar] [CrossRef]
- Vanzo, A.; Janeš, L.; Požgan, F.; Velikonja Bolta, Š.; Sivilotti, P.; Lisjak, K. UHPLC-MS/MS Determination of Varietal Thiol Precursors in Sauvignon Blanc Grapes. Sci. Rep. 2017, 7, 13122. [Google Scholar] [CrossRef]
- Ruiz, J.; Kiene, F.; Belda, I.; Fracassetti, D.; Marquina, D.; Navascués, E.; Calderón, F.; Benito, A.; Rauhut, D.; Santos, A.; et al. Effects on Varietal Aromas during Wine Making: A Review of the Impact of Varietal Aromas on the Flavor of Wine. Appl. Microbiol. Biotechnol. 2019, 103, 7425–7450. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Takase, H.; Suzuki, Y.; Tanzawa, F.; Takata, R.; Fujita, K.; Kohno, M.; Mochizuki, M.; Suzuki, S.; Konno, T. Environmental Stress Enhances Biosynthesis of Flavor Precursors, S-3-(Hexan-1-Ol)-Glutathione and S-3-(Hexan-1-Ol)-L-Cysteine, in Grapevine through Glutathione S-Transferase Activation. J. Exp. Bot. 2011, 62, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Paciello, P.; Mencarelli, F.; Palliotti, A.; Ceccantoni, B.; Thibon, C.; Darriet, P.; Pasquini, M.; Bellincontro, A. Nebulized Water Cooling of the Canopy Affects Leaf Temperature, Berry Composition and Wine Quality of Sauvignon Blanc. J. Sci. Food Agric. 2017, 97, 1267–1275. [Google Scholar] [CrossRef]
- Pastore, C.; Allegro, G.; Valentini, G.; Pizziolo, A.; Battista, F.; Spinelli, F.; Filippetti, I. Foliar Application of Specific Yeast Derivative Enhances Anthocyanins Accumulation and Gene Expression in Sangiovese Cv (Vitis vinifera L.). Sci. Rep. 2020, 10, 11627. [Google Scholar] [CrossRef] [PubMed]
- Šuklje, K.; Antalick, G.; Buica, A.; Coetzee, Z.A.; Brand, J.; Schmidtke, L.M.; Vivier, M.A. Inactive Dry Yeast Application on Grapes Modify Sauvignon Blanc Wine Aroma. Food Chem. 2016, 197, 1073–1084. [Google Scholar] [CrossRef]
- Coombe, B.G. Growth Stages of the Grapevine: Adoption of a System for Identifying Grapevine Growth Stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Spayd, S.; Andersen-Bagge, J. Free Amino Acid Composition of Grape Juice From 12 Vitis Vinifera Cultivars in Washington. Am. J. Enol. Vitic. 1996, 47, 389–402. [Google Scholar] [CrossRef]
- Pérez-Díaz, R.; Madrid-Espinoza, J.; Salinas-Cornejo, J.; González-Villanueva, E.; Ruiz-Lara, S. Differential Roles for VviGST1, VviGST3, and VviGST4 in Proanthocyanidin and Anthocyanin Transport in Vitis Vinífera. Front. Plant Sci. 2016, 7, 1166. [Google Scholar] [CrossRef]
- Martin, D.M.; Aubourg, S.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional Annotation, Genome Organization and Phylogeny of the Grapevine (Vitis vinifera) Terpene Synthase Gene Family Based on Genome Assembly, FLcDNA Cloning, and Enzyme Assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef]
- Swarup, R.; Bhosale, R. Developmental Roles of AUX1/LAX Auxin Influx Carriers in Plants. Front. Plant Sci. 2019, 10, 1306. [Google Scholar] [CrossRef]
- Franco-Zorrilla, J.M.; Martín, A.C.; Leyva, A.; Paz-Ares, J. Interaction between Phosphate-Starvation, Sugar, and Cytokinin Signaling in Arabidopsis and the Roles of Cytokinin Receptors CRE1/AHK4 and AHK3. Plant Physiol. 2005, 138, 847–857. [Google Scholar] [CrossRef]
- Seok, K.C.; Moon, Y.R.; Song, C.; Kwak, J.M.; Woo, T.K. Arabidopsis PUB22 and PUB23 Are Homologous U-Box E3 Ubiquitin Ligases That Play Combinatory Roles in Response to Drought Stress. Plant Cell 2008, 20, 1899–1914. [Google Scholar] [CrossRef]
- Trujillo, M.; Ichimura, K.; Casais, C.; Shirasu, K. Negative Regulation of PAMP-Triggered Immunity by an E3 Ubiquitin Ligase Triplet in Arabidopsis. Curr. Biol. 2008, 18, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Pi, L.Y.; Chen, X.; Chakrabarty, P.K.; Jiang, J.; De Leon, A.L.; Liu, G.Z.; Li, A.; Benny, U.; Oard, J.; et al. Rice XA21 Binding Protein 3 Is a Ubiquitin Ligase Required for Full Xa21-Mediated Disease Resistance. Plant Cell 2006, 18, 3635–3646. [Google Scholar] [CrossRef] [PubMed]
- Scarpeci, T.E.; Zanor, M.I.; Mueller-Roeber, B.; Valle, E.M. Overexpression of AtWRKY30 Enhances Abiotic Stress Tolerance during Early Growth Stages in Arabidopsis thaliana. Plant Mol. Biol. 2013, 83, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kracher, B.; Ziegler, J.; Birkenbihl, R.P.; Somssich, I.E. Negative Regulation of ABA Signaling By WRKY33 Is Critical for Arabidopsis Immunity towards Botrytis Cinerea 2100. eLife 2015, 4, e07295. [Google Scholar] [CrossRef]
- Gaber, A.; Ogata, T.; Maruta, T.; Yoshimura, K.; Tamoi, M.; Shigeoka, S. The Involvement of Arabidopsis Glutathione Peroxidase 8 in the Suppression of Oxidative Damage in the Nucleus and Cytosol. Plant Cell Physiol. 2012, 53, 1596–1606. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, X.; Goatley, M.; Ervin, E. Heat Shock Proteins in Relation to Heat Stress Tolerance of Creeping Bentgrass at Different N Levels. PLoS ONE 2014, 9, e102914. [Google Scholar] [CrossRef]
- Müller, K.; Carstens, A.C.; Linkies, A.; Torres, M.A.; Leubner-Metzger, G. The NADPH-Oxidase AtrbohB Plays a Role in Arabidopsis Seed after-Ripening. New Phytol. 2009, 184, 885–897. [Google Scholar] [CrossRef]
- Friedrich, T.; Oberkofler, V.; Trindade, I.; Altmann, S.; Brzezinka, K.; Lämke, J.; Gorka, M.; Kappel, C.; Sokolowska, E.; Skirycz, A.; et al. Heteromeric HSFA2/HSFA3 Complexes Drive Transcriptional Memory after Heat Stress in Arabidopsis. Nat. Commun. 2021, 12, 3426. [Google Scholar] [CrossRef]
- Huang, Y.C.; Niu, C.Y.; Yang, C.R.; Jinn, T.L. The Heat Stress Factor HSFA6b Connects ABA Signaling and ABA-Mediated Heat Responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef]
- Lario, L.D.; Ramirez-Parra, E.; Gutierrez, C.; Spampinato, C.P.; Casati, P. ANTI-SILENCING FUNCTION1 Proteins Are Involved in Ultraviolet-Induced DNA Damage Repair and Are Cell Cycle Regulated by E2F Transcription Factors in Arabidopsis. Plant Physiol. 2013, 162, 1164–1177. [Google Scholar] [CrossRef] [PubMed]
- Reiner, T.; Hoefle, C.; Huesmann, C.; Ménesi, D.; Fehér, A.; Hückelhoven, R. The Arabidopsis ROP-Activated Receptor-like Cytoplasmic Kinase RLCK VI_A3 Is Involved in Control of Basal Resistance to Powdery Mildew and Trichome Branching. Plant Cell Rep. 2015, 34, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-Mediated Wound Signalling Promotes Plant Regeneration. Nat. Plants 2019, 5, 491–497. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The Versatile GABA in Plants. Plant Signal. Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef] [PubMed]
- Sewelam, N.; Kazan, K.; Hüdig, M.; Maurino, V.G.; Schenk, P.M. The ATHSP17.4c1 Gene Expression Is Mediated by Diverse Signals That Link Biotic and Abiotic Stress Factors with Ros and Can Be a Useful Molecular Marker for Oxidative Stress. Int. J. Mol. Sci. 2019, 20, 3201. [Google Scholar] [CrossRef]
- McLoughlin, F.; Basha, E.; Fowler, M.E.; Kim, M.; Bordowitz, J.; Katiyar-Agarwal, S.; Vierling, E. Class I and II Small Heat Shock Proteins Together with HSP101 Protect Protein Translation Factors during Heat Stress. Plant Physiol. 2016, 172, 1221–1236. [Google Scholar] [CrossRef]
- Tiwari, L.D.; Khungar, L.; Grover, A. AtHsc70-1 Negatively Regulates the Basal Heat Tolerance in Arabidopsis thaliana through Affecting the Activity of HsfAs and Hsp101. Plant J. 2020, 103, 2069–2083. [Google Scholar] [CrossRef]
- Lee, T.A.; Bailey-Serres, J. Integrative Analysis from the Epigenome to Translatome Uncovers Patterns of Dominant Nuclear Regulation during Transient Stress. Plant Cell 2019, 31, 2573–2595. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, Y.; Kieffer, M.; Yu, H.; Kepinski, S.; Estelle, M. HSP90 Regulates Temperature-Dependent Seedling Growth in Arabidopsis by Stabilizing the Auxin Co-Receptor F-Box Protein TIR1. Nat. Commun. 2016, 7, 10269. [Google Scholar] [CrossRef]
- Fu, Q.; Li, S.; Yu, D. Identification of an Arabidopsis Nodulin-Related Protein in Heat Stress. Mol. Cells 2010, 29, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Yang, Y.; Feng, H.; Pan, Z.; Shen, W.H.; Zhu, Y.; Dong, A. Histone Chaperone ASF1 Is Involved in Gene Transcription Activation in Response to Heat Stress in Arabidopsis thaliana. Plant Cell Environ. 2014, 37, 2128–2138. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Shang, Y.; Kang, H.K.; Kim, S.Y.; Kim, B.H.; Nam, K.H. Arabidopsis Galactinol Synthases 1 (AtGOLS1) Negatively Regulates Seed Germination. Plant Sci. 2018, 267, 94–101. [Google Scholar] [CrossRef] [PubMed]
- De Jesús Chávez Martínez, Á. Altered Levels of Glycosylated Sterols Affect Tomato Development and Stress Response. Ph.D. Thesis, University of Barcelona, Barcelona, Spain, 2020. [Google Scholar]
- Yang, X.; Chen, L.; Yang, Y.; Guo, X.; Chen, G.; Xiong, X.; Dong, D.; Li, G. Transcriptome Analysis Reveals That Exogenous Ethylene Activates Immune and Defense Responses in a High Late Blight Resistant Potato Genotype. Sci. Rep. 2020, 10, 21294. [Google Scholar] [CrossRef]
- Kim, E.H.; Kim, Y.S.; Park, S.H.; Koo, Y.J.; Choi, Y.D.; Chung, Y.Y.; Lee, I.J.; Kim, J.K. Methyl Jasmonate Reduces Grain Yield by Mediating Stress Signals to Alter Spikelet Development in Rice1. Plant Physiol. 2009, 149, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, L.; Baldwin, I.T. Methyl Jasmonate-Elicited Herbivore Resistance: Does MeJA Function as a Signal without Being Hydrolyzed to JA? Planta 2008, 227, 1161–1168. [Google Scholar] [CrossRef]
- Oh, I.S.; Park, A.R.; Bae, M.S.; Kwon, S.J.; Kim, Y.S.; Lee, J.E.; Kang, N.Y.; Lee, S.; Cheong, H.; Park, O.K. Secretome Analysis Reveals an Arabidopsis Lipase Involved in Defense against Altemaria brassicicola. Plant Cell 2005, 17, 2832–2847. [Google Scholar] [CrossRef]
- Dervisi, I.; Valassakis, C.; Agalou, A.; Papandreou, N.; Podia, V.; Haralampidis, K.; Iconomidou, V.A.; Kouvelis, V.N.; Spaink, H.P.; Roussis, A. Investigation of the Interaction of DAD1-LIKE LIPASE 3 (DALL3) with Selenium Binding Protein 1 (SBP1) in Arabidopsis thaliana. Plant Sci. 2020, 291, 110357. [Google Scholar] [CrossRef]
- Krčková, Z.; Brouzdová, J.; Daněk, M.; Kocourková, D.; Rainteau, D.; Ruelland, E.; Valentová, O.; Pejchar, P.; Martinec, J. Arabidopsis Non-Specific Phospholipase C1: Characterization and Its Involvement in Response to Heat Stress. Front. Plant Sci. 2015, 6, 928. [Google Scholar] [CrossRef]
- Gidda, S.K.; Miersch, O.; Levitin, A.; Schmidt, J.; Wasternack, C.; Varin, L. Biochemical and Molecular Characterization of a Hydroxyjasmonate Sulfotransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 17895–17900. [Google Scholar] [CrossRef]
- Funck, D.; Eckard, S.; Müller, G. Non-Redundant Functions of Two Proline Dehydrogenase Isoforms in Arabidopsis. BMC Plant Biol. 2010, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Wang, J.; Cheng, L.; Liu, S.; Wu, J.; Peng, Z.; Lu, G. Genome-Wide Analysis of the Mitogen-Activated Protein Kinase Gene Family in Solanum lycopersicum. Gene 2012, 499, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Gursanscky, N.R.; Jouannet, V.; Grünwald, K.; Sanchez, P.; Laaber-Schwarz, M.; Greb, T. MOL1 Is Required for Cambium Homeostasis in Arabidopsis. Plant J. 2016, 86, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Park, Y.J.; Son, S.H.; Roh, J.; Youn, J.H.; Kim, S.Y.; Kim, S.K. Brassinosteroids Signaling via BZR1 Down-Regulates Expression of ACC Oxidase 4 to Control Growth of Arabidopsis thaliana Seedlings. Plant Signal. Behav. 2020, 15, 1734333. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Wan, J.; Kim, H.J.; Nguyen, X.C.; Chung, W.S.; Hong, J.C.; Stacey, G. Ethylene-Responsive Element-Binding Factor 5, ERF5, Is Involved in Chitin-Induced Innate Immunity Response. Mol. Plant-Microbe Interact. 2012, 25, 48–60. [Google Scholar] [CrossRef]
- Moffat, C.S.; Ingle, R.A.; Wathugala, D.L.; Saunders, N.J.; Knight, H.; Knight, M.R. ERF5 and ERF6 Play Redundant Roles as Positive Regulators of JA/Et-Mediated Defense against Botrytis cinerea in Arabidopsis. PLoS ONE 2012, 7, e35995. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, Y.; Yamoto, N.; Suzuki, Y.; Chiba, Y.; Yamazaki, K.; Sato, T.; Yamaguchi, J. The Arabidopsis Transcriptional Repressor ERF9 Participates in Resistance against Necrotrophic Fungi. Plant Sci. 2013, 213, 79–87. [Google Scholar] [CrossRef]
- Yang, Z.; Tian, L.; Latoszek-Green, M.; Brown, D.; Wu, K. Arabidopsis ERF4 Is a Transcriptional Repressor Capable of Modulating Ethylene and Abscisic Acid Responses. Plant Mol. Biol. 2005, 58, 585–596. [Google Scholar] [CrossRef]
- Chen, W.H.; Li, P.F.; Chen, M.K.; Lee, Y.I.; Yang, C.H. FOREVER YOUNG FLOWER Negatively Regulates Ethylene Response DNA-Binding Factors by Activating an Ethylene-Responsive Factor to Control Arabidopsis Floral Organ Senescence and Abscission. Plant Physiol. 2015, 168, 1666–1683. [Google Scholar] [CrossRef]
- Schweighofer, A.; Kazanaviciute, V.; Scheikl, E.; Teige, M.; Doczi, R.; Hirt, H.; Schwanninger, M.; Kant, M.; Schuurink, R.; Mauch, F.; et al. The PP2C-Type Phosphatase AP2C1, Which Negatively Regulates MPK4 and MPK6, Modulates Innate Immunity, Jasmonic Acid, and Ethylene Levels in Arabidopsis. Plant Cell 2007, 19, 2213–2224. [Google Scholar] [CrossRef]
- Licausi, F.; Van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, Two Hypoxia-Inducible Ethylene Response Factors, Affect Anaerobic Responses in Arabidopsis thaliana. Plant J. 2010, 62, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.P.; Diezel, C.; Somssich, I.E. Arabidopsis WRKY33 Is a Key Transcriptional Regulator of Hormonal and Metabolic Responses toward Botrytis Cinerea Infection. Plant Physiol. 2012, 159, 266–285. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Guo, J.; Zhang, T.; Bai, S.; He, K.; Wang, Z. Genome-Wide Analysis of WRKY Gene Family and the Dynamic Responses of Key WRKY Genes Involved in Ostrinia Furnacalis Attack in Zea Mays. Int. J. Mol. Sci. 2021, 22, 13045. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.C.; Chou, M.Y.; Chou, S.J.; Li, Y.R.; Peng, H.P.; Shih, M.C. Submergence Confers Immunity Mediated by the WRKY22 Transcription Factor in Arabidopsis. Plant Cell 2013, 25, 2699–2713. [Google Scholar] [CrossRef]
- Bolt, S.; Zuther, E.; Zintl, S.; Hincha, D.K.; Schmülling, T. ERF105 Is a Transcription Factor Gene of Arabidopsis thaliana Required for Freezing Tolerance and Cold Acclimation. Plant Cell Environ. 2017, 40, 108–120. [Google Scholar] [CrossRef]
- Lehti-Shiu, M.D.; Uygun, S.; Moghe, G.D.; Panchy, N.; Fang, L.; Hufnagel, D.E.; Jasicki, H.L.; Feig, M.; Shiu, S.H. Molecular Evidence for Functional Divergence and Decay of a Transcription Factor Derived from Whole-Genome Duplication in Arabidopsis thaliana. Plant Physiol. 2015, 168, 1717–1734. [Google Scholar] [CrossRef]
- Van Dingenen, J.; Vermeersch, M.; De Milde, L.; Hulsmans, S.; De Winne, N.; Van Leene, J.; Gonzalez, N.; Dhondt, S.; De Jaeger, G.; Rolland, F.; et al. The Role of HEXOKINASE1 in Arabidopsis Leaf Growth. Plant Mol. Biol. 2019, 99, 79–93. [Google Scholar] [CrossRef]
- Ding, Z.J.; Yan, J.Y.; Li, G.X.; Wu, Z.C.; Zhang, S.Q.; Zheng, S.J. WRKY41 Controls Arabidopsis Seed Dormancy via Direct Regulation of ABI3 Transcript Levels Not Downstream of ABA. Plant J. 2014, 79, 810–823. [Google Scholar] [CrossRef]
- AbuQamar, S.; Chen, X.; Dhawan, R.; Bluhm, B.; Salmeron, J.; Lam, S.; Dietrich, R.A.; Mengiste, T. Expression Profiling and Mutant Analysis Reveals Complex Regulatory Networks Involved in Arabidopsis Response to Botrytis Infection. Plant J. 2006, 48, 28–44. [Google Scholar] [CrossRef]
- Xie, Z.; Li, D.; Wang, L.; Sack, F.D.; Grotewold, E. Role of the Stomatal Development Regulators FLP/MYB88 in Abiotic Stress Responses. Plant J. 2010, 64, 731–739. [Google Scholar] [CrossRef]
- Yang, S.D.; Seo, P.J.; Yoon, H.K.; Park, C.M. The Arabidopsis NAC Transcription Factor VNI2 Integrates Abscisic Acid Signals into Leaf Senescence via the COR/RD Genes. Plant Cell 2011, 23, 2155–2168. [Google Scholar] [CrossRef]
- Chen, Y.F.; Li, L.Q.; Xu, Q.; Kong, Y.H.; Wang, H.; Wu, W.H. The WRKY6 Transcription Factor Modulates PHOSPHATE1 Expression in Response to Low Pi Stress in Arabidopsis. Plant Cell 2009, 21, 3554–3566. [Google Scholar] [CrossRef]
- Yu, X.C.; Zhu, S.Y.; Gao, G.F.; Wang, X.J.; Zhao, R.; Zou, K.Q.; Wang, X.F.; Zhang, X.Y.; Wu, F.Q.; Peng, C.C.; et al. Expression of a Grape Calcium-Dependent Protein Kinase ACPK1 in Arabidopsis thaliana Promotes Plant Growth and Confers Abscisic Acid-Hypersensitivity in Germination, Postgermination Growth, and Stomatal Movement. Plant Mol. Biol. 2007, 64, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Candat, A.; Paszkiewicz, G.; Neveu, M.; Gautier, R.; Logan, D.C.; Avelange-Macherel, M.H.; Macherel, D. The Ubiquitous Distribution of Late Embryogenesis Abundant Proteins across Cell Compartments in Arabidopsis Offers Tailored Protection against Abiotic Stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving Plant Drought, Salt and Freezing Tolerance by Gene Transfer of a Single Stress-Inducible Transcription Factor. Novartis Found. Symp. 2001, 236, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.; Koschmieder, J.; Wuest, F.; Schaub, P.; Fehling-Kaschek, M.; Timmer, J.; Beyer, P.; Al-Babili, S. Enzymatic Study on AtCCD4 and AtCCD7 and Their Potential to Form Acyclic Regulatory Metabolites. J. Exp. Bot. 2016, 67, 5993–6005. [Google Scholar] [CrossRef]
- Cao, M.J.; Wang, Z.; Zhao, Q.; Mao, J.L.; Speiser, A.; Wirtz, M.; Hell, R.; Zhu, J.K.; Xiang, C. Bin Sulfate Availability Affects ABA Levels and Germination Response to ABA and Salt Stress in Arabidopsis thaliana. Plant J. 2014, 77, 604–615. [Google Scholar] [CrossRef]
- Song, C.; Kim, T.; Chung, W.S.; Lim, C.O. The Arabidopsis Phytocystatin AtCYS5 Enhances Seed Germination and Seedling Growth under Heat Stress Conditions. Mol. Cells 2017, 40, 577–586. [Google Scholar] [CrossRef]
- Lim, C.W.; Yang, S.H.; Shin, K.H.; Lee, S.C.; Kim, S.H. The AtLRK10L1.2, Arabidopsis Ortholog of Wheat LRK10, Is Involved in ABA-Mediated Signaling and Drought Resistance. Plant Cell Rep. 2015, 34, 447–455. [Google Scholar] [CrossRef]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Brachypodium BdABCG25 is a homolog of Arabidopsis AtABCG25 involved in the transport of abscisic acid. FEBS Lett. 2021, 595, 954–959. [Google Scholar] [CrossRef]
- Kang, J.; Hwang, J.U.; Lee, M.; Kim, Y.Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-Type ABC Transporter Mediates Cellular Uptake of the Phytohormone Abscisic Acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Hanada, A.; Kuwahara, A.; Endo, A.; Okamoto, M.; Yamauchi, Y.; North, H.; Marion-Poll, A.; Sun, T.P.; Koshiba, T.; et al. Regulation of Hormone Metabolism in Arabidopsis Seeds: Phytochrome Regulation of Abscisic Acid Metabolism and Abscisic Acid Regulation of Gibberellin Metabolism. Plant J. 2006, 48, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Kang, S.G.; Hah, C.; Jang, J.C. Molecular and Cellular Characterization of GA-Stimulated Transcripts GASA4 and GASA6 in Arabidopsis thaliana. Plant Sci. 2016, 246, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wulfetange, K.; Lomin, S.N.; Romanov, G.A.; Stolz, A.; Heyl, A.; Schmülling, T. The Cytokinin Receptors of Arabidopsis Are Located Mainly to the Endoplasmic Reticulum. Plant Physiol. 2011, 156, 1808–1818. [Google Scholar] [CrossRef]
- Cui, F.; Wu, S.; Sun, W.; Coaker, G.; Kunkel, B.; He, P.; Shan, L. The Pseudomonas Syringae Type III Effector AvrRpt2 Promotes Pathogen Virulence via Stimulating Arabidopsis Auxin/Indole Acetic Acid Protein Turnover. Plant Physiol. 2013, 162, 1018–1029. [Google Scholar] [CrossRef]
- Hazak, O.; Mamon, E.; Lavy, M.; Sternberg, H.; Behera, S.; Schmitz-Thom, I.; Bloch, D.; Dementiev, O.; Gutman, I.; Danziger, T.; et al. A Novel Ca2+-Binding Protein That Can Rapidly Transduce Auxin Responses during Root Growth. PLoS Biol. 2019, 17, e3000085. [Google Scholar] [CrossRef]
- Zwack, P.J.; Compton, M.A.; Adams, C.I.; Rashotte, A.M. Cytokinin Response Factor 4 (CRF4) Is Induced by Cold and Involved in Freezing Tolerance. Plant Cell Rep. 2016, 35, 573–584. [Google Scholar] [CrossRef]
- Banasiak, A.; Biedroń, M.; Dolzblasz, A.; Berezowski, M.A. Ontogenetic Changes in Auxin Biosynthesis and Distribution Determine the Organogenic Activity of the Shoot Apical Meristem in Pin1 Mutants. Int. J. Mol. Sci. 2019, 20, 180. [Google Scholar] [CrossRef]
- Bennett, M.J.; Marchant, A.; Green, H.G.; May, S.T.; Ward, S.P.; Millner, P.A.; Walker, A.R.; Schulz, B.F.K. Arabidopsis AUX1 Gene: A Permease-Like Regulator of Root Gravitropism. Science 1996, 273, 948–950. [Google Scholar] [CrossRef]
- Hanzawa, Y.; Takahashi, T.; Komeda, Y. ACL5: An Arabidopsis Gene Required for Internodal Elongation after Flowering. Plant J. 1997, 12, 863–874. [Google Scholar] [CrossRef]
- Morita, M.T.; Sakaguchi, K.; Kiyose, S.I.; Taira, K.; Kato, T.; Nakamura, M.; Tasaka, M. A C2H2-Type Zinc Finger Protein, SGR5, Is Involved in Early Events of Gravitropism in Arabidopsis Inflorescence Stems. Plant J. 2006, 47, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Hallmark, H.T.; Rashotte, A.M. Review—Cytokinin Response Factors: Responding to More than Cytokinin. Plant Sci. 2019, 289, 110251. [Google Scholar] [CrossRef] [PubMed]
- Gälweiler, L.; Guan, C.; Müller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of Polar Auxin Transport by AtPIN1 in Arabidopsis Vascular Tissue. Science 1998, 282, 2226–2230. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, D.; Lonardi, A.; Boscaro, D.; Nardi, T.; Marangon, C.M.; De Rosso, M.; Flamini, R.; Lovat, L.; Mian, G. Effects of Traditional and Modern Post-Harvest Withering Processes on the Composition of the Vitis v. Corvina Grape and the Sensory Profile of Amarone Wines. Molecules 2021, 26, 5198. [Google Scholar] [CrossRef]
- Tomasi, D.; Panighel, A.; Flamini, R.; Lovat, L.; Battista, F. Foliar Application of Specific Inactivated Yeast with Action on Phenolic and Aromatic Metabolism of Grapes. In Proceedings of the 40th World Vine and Wine Congress, OIV, Sofia, Bulgaria, 29 May 2017; p. 40. [Google Scholar]
- Mozzon, M.; Savini, S.; Boselli, E.; Thorngate, J.H. The Herbaceous Character of Wines. Ital. J. Food Sci. 2016, 28, 190–207. [Google Scholar]
- Stines, A.P.; Grubb, J.; Gockowiak, H.; Henschke, P.A.; Høj, P.B.; Van Heeswijck, R. Proline and Arginine Accumulation in Developing Berries of Vitis vinifera L. in Australian Vineyards: Influence of Vine Cultivar, Berry Maturity and Tissue Type. Aust. J. Grape Wine Res. 2000, 6, 150–158. [Google Scholar] [CrossRef]
- Stribny, J.; Gamero, A.; Pérez-Torrado, R.; Querol, A. Saccharomyces Kudriavzevii and Saccharomyces Uvarum Differ from Saccharomyces Cerevisiae during the Production of Aroma-Active Higher Alcohols and Acetate Esters Using Their Amino Acidic Precursors. Int. J. Food Microbiol. 2015, 205, 41–46. [Google Scholar] [CrossRef]
- Monforte, A.R.; Martins, S.I.F.S.; Silva Ferreira, A.C. Strecker Aldehyde Formation in Wine: New Insights into the Role of Gallic Acid, Glucose, and Metals in Phenylacetaldehyde Formation. J. Agric. Food Chem. 2018, 66, 2459–2466. [Google Scholar] [CrossRef]
- Tadmor, Y.; Fridman, E.; Gur, A.; Larkov, O.; Lastochkin, E.; Ravid, U.; Zamir, D.; Lewinsohn, E. Identification of Malodorous, a Wild Species Allele Affecting Tomato Aroma That Was Selected against during Domestication. J. Agric. Food Chem. 2002, 50, 2005–2009. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline Accumulation in Plants: A Review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Pereira, G.E.; Gaudillere, J.P.; Pieri, P.; Hilbert, G.; Maucourt, M.; Deborde, C.; Moing, A.; Rolin, D. Microclimate Influence on Mineral and Metabolic Profiles of Grape Berries. J. Agric. Food Chem. 2006, 54, 6765–6775. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.G. A Review of the Applications of Chitin and Its Derivatives in Agriculture to Modify Plant-Microbial Interactions and Improve Crop Yields. Agronomy 2013, 3, 757–793. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative Burst: An Early Plant Response to Pathogen Infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.B.; Butterfield, D.A. Measurement of Oxidized/Reduced Glutathione Ratio. In Protein Misfolding and Cellular Stress in Disease and Aging; Humana Press: Totowa, NJ, USA, 2010; pp. 269–277. [Google Scholar]
- Varala, K.; Marshall-Colón, A.; Cirrone, J.; Brooks, M.D.; Pasquino, A.V.; Léran, S.; Mittal, S.; Rock, T.M.; Edwards, M.B.; Kim, G.J.; et al. Temporal Transcriptional Logic of Dynamic Regulatory Networks Underlying Nitrogen Signaling and Use in Plants. Proc. Natl. Acad. Sci. USA 2018, 115, 6494–6499. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Wang, J.; Jiang, T.; Ma, T.; Wang, R. Effect of Nitrogen Regulation on Berry Quality and Flavonoids during Veraison Stage. Food Sci. Nutr. 2021, 9, 5448–5456. [Google Scholar] [CrossRef]
- Lallemand LalVigne AROMA. Available online: https://lalvigne.com/it/italy/our-products/product-details/aroma-it/ (accessed on 28 May 2021).
- Thermo Fisher Scientific. Thermo Fisher Scientific Titratable Acidity of Red Wine by Manual Titration (Potentiometric) Application Note AN010; Thermo Fisher Scientific: Waltham, MA, USA, 2022. [Google Scholar]
- Modesti, M.; Brizzolara, S.; Forniti, R.; Ceccantoni, B.; Bellincontro, A.; Catelli, C.; Mencarelli, F.; Tonutti, P. Postharvest Ozone Fumigation of Grapes (Cv Sangiovese) Differently Affects Volatile Organic Compounds and Polyphenol Profiles of Berries and Wine. Aust. J. Grape Wine Res. 2023, 2023, 8244309. [Google Scholar] [CrossRef]
- Paolini, M.; Tonidandel, L.; Moser, S.; Larcher, R. Development of a Fast Gas Chromatography–Tandem Mass Spectrometry Method for Volatile Aromatic Compound Analysis in Oenological Products. J. Mass Spectrom. 2018, 53, 801–810. [Google Scholar] [CrossRef]
- Tonidandel, L.; Larcher, R.; Barbero, A.; Jelley, R.E.; Fedrizzi, B. A Single Run Liquid Chromatography-Tandem Mass Spectrometry Method for the Analysis of Varietal Thiols and Their Precursors in Wine. J. Chromatogr. A 2021, 1658, 462603. [Google Scholar] [CrossRef]
- Schilling, M.; Maia-Grondard, A.; Baltenweck, R.; Robert, E.; Hugueney, P.; Bertsch, C.; Farine, S.; Gelhaye, E. Wood Degradation by Fomitiporia Mediterranea M. Fischer: Physiologic, Metabolomic and Proteomic Approaches. Front. Plant Sci. 2022, 13, 988709. [Google Scholar] [CrossRef]
- Nonis, A.; Vezzaro, A.; Ruperti, B. Evaluation of RNA Extraction Methods and Identification of Putative Reference Genes for Real-Time Quantitative Polymerase Chain Reaction Expression Studies on Olive (Olea europaea L.) Fruits. J. Agric. Food Chem. 2012, 60, 6855–6865. [Google Scholar] [CrossRef]
- González-Agüero, M.; García-Rojas, M.; Di Genova, A.; Correa, J.; Maass, A.; Orellana, A.; Hinrichsen, P. Identification of Two Putative Reference Genes from Grapevine Suitable for Gene Expression Analysis in Berry and Related Tissues Derived from RNA-Seq Data. BMC Genom. 2013, 14, 878. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]
- Del Fabbro, C.; Scalabrin, S.; Morgante, M.; Giorgi, F.M. An Extensive Evaluation of Read Trimming Effects on Illumina NGS Data Analysis. PLoS ONE 2013, 8, e85024. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grapevine Gene | Arabidopsis Homologue | Function in Arabidopsis | Group | Reference | Putative Role |
|---|---|---|---|---|---|
| Response to chitin and regulatory burst involved in defence response | |||||
| Vitvi08g02053 | AT2G35930 | Plant U-box 23 | 1 | Cho et al., 2008 [22] Trujillo et al., 2008 [23] | Ubiquitin ligase homologous to U-Box E3 playing a role in response to drought stress. |
| Vitvi08g02051 | AT2G35930 | Plant U-box 23 | 1 | Cho et al., 2008 [22] Trujillo et al., 2009 [23] | Ubiquitin ligase homologous to U-Box E3 playing a role in response to drought stress. |
| Vitvi12g00139 | AT1G29340 | Plant U-box 17 | 4 | Wang et al., 2006 [24] | In tobacco: ubiquitin-proteasome system in defences against pathogens. |
| Vitvi15g01003 | AT5G24110 | WRKY Transcription Factor Group III | 1 | Scarpeci et al., 2013 [25] | AtWRKY30, putative role in the activation of defence responses. |
| Vitvi07g03110 | AT5G65530 | Protein kinase superfamily protein | 1 | Reiner et al., 2015 [33] | RLCK VI, regulating basal resistance to powdery mildew. |
| ROS Homeostasis | |||||
| Vitvi02g00332 | AT1G63460 | Glutathione peroxidase 8 | 1 | Gaber et al., 2012 [27] | Involvement in the suppression of oxidative damage. |
| Vitvi03g00500 | AT4G34410 | ERF109 | 1 | Zhang et al., 2019 [34] Li et al., 2021 [35] | Wounding, JA and salt stress inducible; regulates ROS production and stress adaptation. |
| Vitvi13g00097 | AT5G58530 | Glutathione oxidoreductase | 2 | No reference | Reduction of thiol groups in proteins. |
| Vitvi14g00183 | AT1G09090 | RBohB | 2 | Wang et al., 2014 [28] Müller et al., 2009 [29] | ABA inducible in seed after ripening and involved in conferring thermotolerance. |
| Heat Shock Proteins | |||||
| Vitvi13g00491 | AT3G46230 | Heat shock protein 17.4 | 1 | Sewelam et al., 2019 [36] McLoughlin et al., 2016 [37] | Response to ROS and several abiotic stresses, including cold and heat. |
| Vitvi06g00443 | AT5G02500 | Heat shock cognate protein 70.1 | 2 | Tiwari et al., 2020 [38] | Negative regulator of basal heat tolerance. |
| Vitvi08g02189 | AT5G02500 | Heat shock cognate protein 70.1 | 1 | Tiwari et al., 2020 [38] | Negative regulator of basal heat tolerance. |
| Vitvi09g00045 | AT1G54050 | HSP20-like chaperones superfamily | 1 | Lee and Bailey-Serres, 2019 [39] | Hypoxia related stress induction. |
| Vitvi02g00025 | AT5G52640 | Heat shock-like protein 90.1 | 2 | Wang et al., 2016 [40] | Interacts with disease resistance signaling components and is required for RPS2-mediated resistance. |
| Vitvi16g01103 | AT5G52640 | Heat shock-like protein 90.1 | 2 | Wang et al., 2016 [40] | Interacts with disease resistance signaling components and is required for RPS2-mediated resistance. |
| Vitvi01g01846 | AT2G03440 | Nodulin-related protein 1 | 1 | Fu et al., 2010 [41] | Negative regulation of ABA response and abiotic stress (cold) inducible. |
| Vitvi04g00092 | AT2G26150 | Heat shock transcription factor A2 | 1 | Friedrich et al., 2021 [30] | Regulating heat stress memory genes and thermotolerance. |
| Vitvi07g00078 | AT3G22830 | Heat shock transcription factor A6B | 1 | Huang et al., 2016 [31] | Positive mediator of ABA dependent thermotolerance and drought resistance. |
| Vitvi01g00372 | AT5G38110 | Anti- silencing function 1b | 1 | Lario et al., 2013 [32] Weng et al., 2014 [42] | A positive regulator of basal and acquired thermotolerance. |
| Vitvi14g02461 | AT2G47180 | Galactinol synthase 131 | 3 | Jang et at., 2018 [43] | Role in improving oxidative stress tolerance by increasing galactinol biosynthesis in Arabidopsis. |
| Hormone biosynthesis and signalling—Lipid metabolism, JA biosynthesis and Response | |||||
| Vitvi13g01780 | AT3G45140 | Lipoxygenase 2 | 1 | No reference | Lipoxygenase involved in JA biosynthesis. |
| Vitvi09g00085 | AT1G17420 | Lipoxygenase 3 | 1 | Chávez-Martínez et al., 2020 [44] Yang et al., 2020 [45] | Lipoxygenase involved in JA biosynthesis. |
| Vitvi04g02169 | AT1G19640 | Jasmonic acid carboxyl methyltransferase | 1 | Kim et al., 2009 [46] Wu et al., 2008 [47] | Catalyses the formation of methyl jasmonate from JA. |
| Vitvi09g00038 | AT3G14225 | GDSL-motif lipase 4 hydrolase | 1 | Oh et al., 2005 [48] | Lipase involved in resistance to necrotrophic pathogens. |
| Vitvi10g00669 | AT5G45670 | GDSL-like Lipase/Acylhydrolase | 1 | No reference | Lipase. Unknown role. |
| Vitvi07g00039 | AT2G30550 | Alpha/beta-Hydrolases superfamily protein | 1 | Dervisi et al., 2020 [49] | Role in JA biosynthesis. Lipase hydrolysing phosphatidylcholine, glycolipids, triacylglycerol. |
| Vitvi15g00298 | AT3G03520 | Phospholipase C3 | 1 | Krčková et al., 2015 [50] | Positive regulator of thermotolerance, induced by phosphate starvation and abiotic stresses. |
| Vitvi13g00864 | AT5G07010 | Sulfotransferase 2A | 1 | Gidda et al., 2003 [51] | Acts on 11- and 12-hydroxyjasmonic acid. Involved in reducing excess JA levels. |
| Vitvi13g01379 | AT5G07010 | Sulfotransferase 2A | 3 | Gidda et al., 2003 [51] | Acts on 11- and 12-hydroxyjasmonic acid. Involved in reducing excess JA levels. |
| Vitvi14g01283 | AT5G38710 | Proline Dehydrogenase | 4 | Funck et al., 2010 [52] | Upregulated during salt stress. |
| Vitvi12g02545 | AT4G08500 | MAPK/ERK kinase 1 | 4 | Kong et al., 2012 [53] | Negative regulator of wound and immune responses. |
| Vitvi11g01479 | AT5G51350 | Leucine-rich repeat transmembrane protein kinase | 4 | Gursanscky et al., 2016 [54] | Represses genes associated with ethylene and JA. |
| Hormone biosynthesis and signalling—Ethylene Biosynthesis and signaling and hypoxic responses | |||||
| Vitvi10g02409 | AT1G05010 | Ethylene-forming enzyme ACO4 | 1 | Moon et al., 2020 [55] | BR repressible ethylene biosynthetic gene involved in inhibition of growth. |
| Vitvi16g01438 | AT5G47230 | ERF5 | 1 | Son et al., 2012 [56] Moffat et al., 2012 [57] | Involved in chitin-induced innate immunity and a positive regulator of JA/ethylene-responsive genes. |
| Vitvi12g00274 | AT5G44210 | ERF9 | 1 | Maruyama et al., 2013 [58] | Regulator of plant defence against necrotrophic fungi mediated by the DEAR1-dependent ethylene/JA signaling pathway. |
| Vitvi19g01784 | AT3G15210 | ERF4 | 1 | Yang et al., 2005 [59] | Modulates ethylene and ABA responses. |
| Vitvi18g02240 | AT1G71450 | Integrase-type DNA-binding superfamily | 1 | Chen et al., 2015 [60] | Negatively regulates ethylene response. |
| Vitvi06g00667 | AT2G30020 | AP2C1 | 1 | Schweighofer et al., 2007 [61] | Negative regulator of ethylene and JA synthesis and responses of resistance to Botrytis c. |
| Vitvi07g00357 | AT2G47520 | ERF B-2 of ERF/AP2 transcription factor family | 1 | Licausi et al., 2010 [62] | Hypoxia responsive ERF. |
| Vitvi08g00793 | AT2G38470 | WRKY33 | 1 | Birkenbihl et al., 2012 [63] Tang et al., 2021 [64] | WRKY transcription factor regulator of Botrytis resistance and hypoxia responses by direct regulation of RAP2.2. |
| Vitvi15g01090 | AT4G01250 | WRKY22 | 1 | Hsu et al., 2013 [65] | WRKY22 family transcription factor, hypoxia inducible and regulating innate immunity in hypoxia. |
| Vitvi16g01444 | AT5G51190 | ERF105 | 1 | Bolt et al., 2017 [66] | Transcription factor required for freezing tolerance and cold acclimation. |
| Vitvi17g00787 | AT1G60190 | ERF105 | 1 | Bolt et al., 2017 [66] | Transcription factor required for freezing tolerance and cold acclimation. |
| Vitvi16g01434 | AT1G60190 | ERF105 | 1 | Bolt et al., 2017 [66] | Transcription factor required for freezing tolerance and cold acclimation. |
| Vitvi16g01432 | AT1G60190 | ERF105 | 1 | Bolt et al., 2017 [66] | Transcription factor required for freezing tolerance and cold acclimation. |
| Vitvi18g00295 | AT1G60190 | ERF105 | 1 | Bolt et al., 2017 [66] | Transcription factor required for freezing tolerance and cold acclimation. |
| Vitvi16g01423 | AT1G60190 | ERF105 | 1 | Bolt et al., 2017 [66] | Transcription factor required for freezing tolerance and cold acclimation. |
| Vitvi02g00407 | AT1G63030 | Integrase-type DNA-binding superfamily | 1 | Lehti-Shiu et al., 2015 [67] | Negative regulator of gibberellic acid biosynthesis. |
| Hormone biosynthesis and signalling—ABA biosynthesis and signaling | |||||
| Vitvi17g00116 | AT3G48530 | SNF1-related protein kinase regulatory subunit gamma 1 | 1 | Van Dingenen et al., 2019 [68] | Regulates responses to sugar availability, negative regulator of HXK1. |
| Vitvi02g00114 | AT4G11070 | WRKY family transcription factor | 1 | Ding et al., 2014 [69] | Regulation of ABI3 independent from ABA, positive regulating ABA sensitivity. |
| Vitvi13g00058 | AT2G40140 | Zinc finger (CCCH-type) family protein | 1 | AbuQamar et al., 2006 [70] | Positive regulator of Botrytis resistance and negative regulator of ABA. |
| Vitvi01g00956 | AT2G02820 | MYB domain protein 88 | 1 | Xie et al., 2010 [71] | Sensing and/or transducing drought and saline stress. |
| Vitvi14g01499 | AT5G13180 | NAC domain containing protein 83 | 1 | Yang et al., 2011 [72] | Molecular link integrating plant responses to environmental stresses and leaf longevity. |
| Vitvi10g00063 | AT1G62300 | WRKY family transcription factor | 1 | Chen et al., 2009 [73] | Regulates the ABA responsive gene Phosphate1 (Pho1) expression in response to low phosphate. |
| Vitvi08g00793 | AT2G38470 | WRKY transcription factor family. | 1 | Liu et al., 2015 [26] | Negative regulator of ABA signalling and positive regulator of resistance to Botrytis. |
| Vitvi08g00298 | AT5G04870 | CDPK1 | 1 | Yu et al., 2007 [74] | Positive regulator of ABA responses in grape berry. |
| Vitvi15g01084 | AT2G46140 | Late embryosis abundant protein | 1 | Candat et al., 2014 [75] | Involved in protection from abiotic stress: desiccation and cold. |
| Vitvi16g00941 | AT4G25480 | DREB subfamily A-1 of ERF/AP2 transcription factor family (CBF3) | 1 | Kasuga et al., 1999 [76] | Involved in conferring resistance to drought and freezing stress. |
| Vitvi02g01288 | AT4G19170 | 9-cis-epoxycarotenoid dioxygenase 4 | 2 | Bruno et al., 2016 [77] | Production of acyclic regulatory metabolites. |
| Vitvi05g00548 | AT3G51895 | Sulphate transporter 3;1 | 1 | Cao et al., 2014 [78] | Co-regulation of S-metabolism and ABA biosynthesis. |
| Vitvi18g03100 | AT5G47550 | Cystatin/monellin superfamily protein | 1 | Song et al., 2017 [79] | Positive role in the heat shock-responsive expression of AtCYS5. |
| Vitvi01g00353 | AT1G18390 | Serine/Threonine kinase | 2 | Lim et al., 2015 [80] | Involved in ABA-mediated signaling and drought resistance. |
| Vitvi18g01703 | AT1G71960 | ATP-binding cassette family G25 ABCG25 | 3 | Kuromori et al. 2021 [81] | Involved in intercellular ABA import inside cells. |
| Vitvi04g00423 | AT1G15520 | ATP-binding cassette family G40 ABCG40 | 3 | Kang et al., 2015 [82] | Necessary for ABA export from cells. |
| Vitvi05g00963 | AT3G24220 | 9-cis-epoxycarotenoid dioxygenase 6 | 3 | Seo et al., 2006 [83] | ABA biosynthetic gene. |
| Vitvi17g00601 | AT1G74670 | GASA6 | 3 | Qu et al., 2016 [84] | ABA repressible and GA inducible; cell wall regulator. |
| Vitvi12g00685 | AT5G35750 | Histidine kinase 2 | 4 | Wulfetange et al., 2011 [85] | Negative regulator of ABA sensitivity in stomata. |
| Hormone biosynthesis and signalling—Auxin and CK Response | |||||
| Vitvi11g00394 | AT3G23050 | Indole-3-acetic acid 7 | 2 | Cui et al., 2013 [86] | AUX/IAA family factor. Negative regulator of pathogenicity, through regulation of auxin response. |
| Vitvi12g00594 | AT4G27280 | Calcium-binding EF-hand family protein | 1 | Hazak et al., 2019 [87] | Ca2+ dependent transducer of auxin-regulated gene expression. |
| Vitvi16g01201 | AT4G27950 | Cytokinin response factor 4 | 1 | Zwack et al., 2016 [88] | Cytokinin response factor 4 (CRF4) is a positive regulator of freezing tolerance |
| Vitvi17g00210 | AT1G73590 | PIN1 auxin efflux carrier | 3 | Banasiak et al., 2019 [89] | Auxin efflux transporter. |
| Vitvi13g00019 | AT2G38120 | AUX1 | 4 | Bennet et al., 1996 [90] | Auxin influx transporter. |
| Vitvi05g00476 | AT5G19530 | Spermine synthase | 4 | Hanzawa et al., 2003 [91] | Biosynthesis of polyamine spermine, promoting growth of organs. |
| Vitvi17g00157 | AT2G01940 | C2H2-like zinc finger protein | 4 | Morita et al., 2006 [92] | Transcription factor involved in shoot gravitropism and gravity perception. |
| Gene | Primer | Primer F | Primer R | Reference |
|---|---|---|---|---|
| VvTCPB (Vitvi18g00138) | TCPB | CAGACAGTGATTGACAGCCGAGTT | ATCCCTGCGTGGCTTTCTTCC | [116] |
| VvAIG1 (Vitvi18g02131) | AIG1 | GGAAGATTATTTGGGCCGTGAG | ACTTCTTGGCTTCATCCTTGGTC | [116] |
| VviLinNer1 (Vitvi10g02128) | Ner1 | GTGGGCGAGTTTATGCAACG | CCCTGAACTAACGGCCCCAT | [19] |
| VviCCD4a (Vitvi02g01286) | CCD4a | CGAGGCATCCGCTATCCACA | CACGTCCAGCTTCACCACTCC | [1] |
| VvGGT (Vitvi11g00234) | GGT | TGGCAACAGCTTAGAGGCAGTA | CCCACCTGCCTTTCTCACAT | [12] |
| VvGST3 (Vitvi17g01467) | GST3 | TGCAAAGGTGTTGGACATCTATG | TGTGAATGGAAGGTGGCTAAGA | [12] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, M.; Forestan, C.; Ravazzolo, L.; Hugueney, P.; Baltenweck, R.; Rasori, A.; Cardillo, V.; Carraro, P.; Malagoli, M.; Brizzolara, S.; et al. Metabolic and Molecular Rearrangements of Sauvignon Blanc (Vitis vinifera L.) Berries in Response to Foliar Applications of Specific Dry Yeast. Plants 2023, 12, 3423. https://doi.org/10.3390/plants12193423
Rodrigues M, Forestan C, Ravazzolo L, Hugueney P, Baltenweck R, Rasori A, Cardillo V, Carraro P, Malagoli M, Brizzolara S, et al. Metabolic and Molecular Rearrangements of Sauvignon Blanc (Vitis vinifera L.) Berries in Response to Foliar Applications of Specific Dry Yeast. Plants. 2023; 12(19):3423. https://doi.org/10.3390/plants12193423
Chicago/Turabian StyleRodrigues, Marta, Cristian Forestan, Laura Ravazzolo, Philippe Hugueney, Raymonde Baltenweck, Angela Rasori, Valerio Cardillo, Pietro Carraro, Mario Malagoli, Stefano Brizzolara, and et al. 2023. "Metabolic and Molecular Rearrangements of Sauvignon Blanc (Vitis vinifera L.) Berries in Response to Foliar Applications of Specific Dry Yeast" Plants 12, no. 19: 3423. https://doi.org/10.3390/plants12193423
APA StyleRodrigues, M., Forestan, C., Ravazzolo, L., Hugueney, P., Baltenweck, R., Rasori, A., Cardillo, V., Carraro, P., Malagoli, M., Brizzolara, S., Quaggiotti, S., Porro, D., Meggio, F., Bonghi, C., Battista, F., & Ruperti, B. (2023). Metabolic and Molecular Rearrangements of Sauvignon Blanc (Vitis vinifera L.) Berries in Response to Foliar Applications of Specific Dry Yeast. Plants, 12(19), 3423. https://doi.org/10.3390/plants12193423