
Effects of Foliar Selenium Application on Oxidative Damage and Photosynthetic Properties of Greenhouse Tomato under Drought Stress


Abstract
1. Introduction
2. Results
2.1. Effect of Se on Oxidative Damage in Tomato
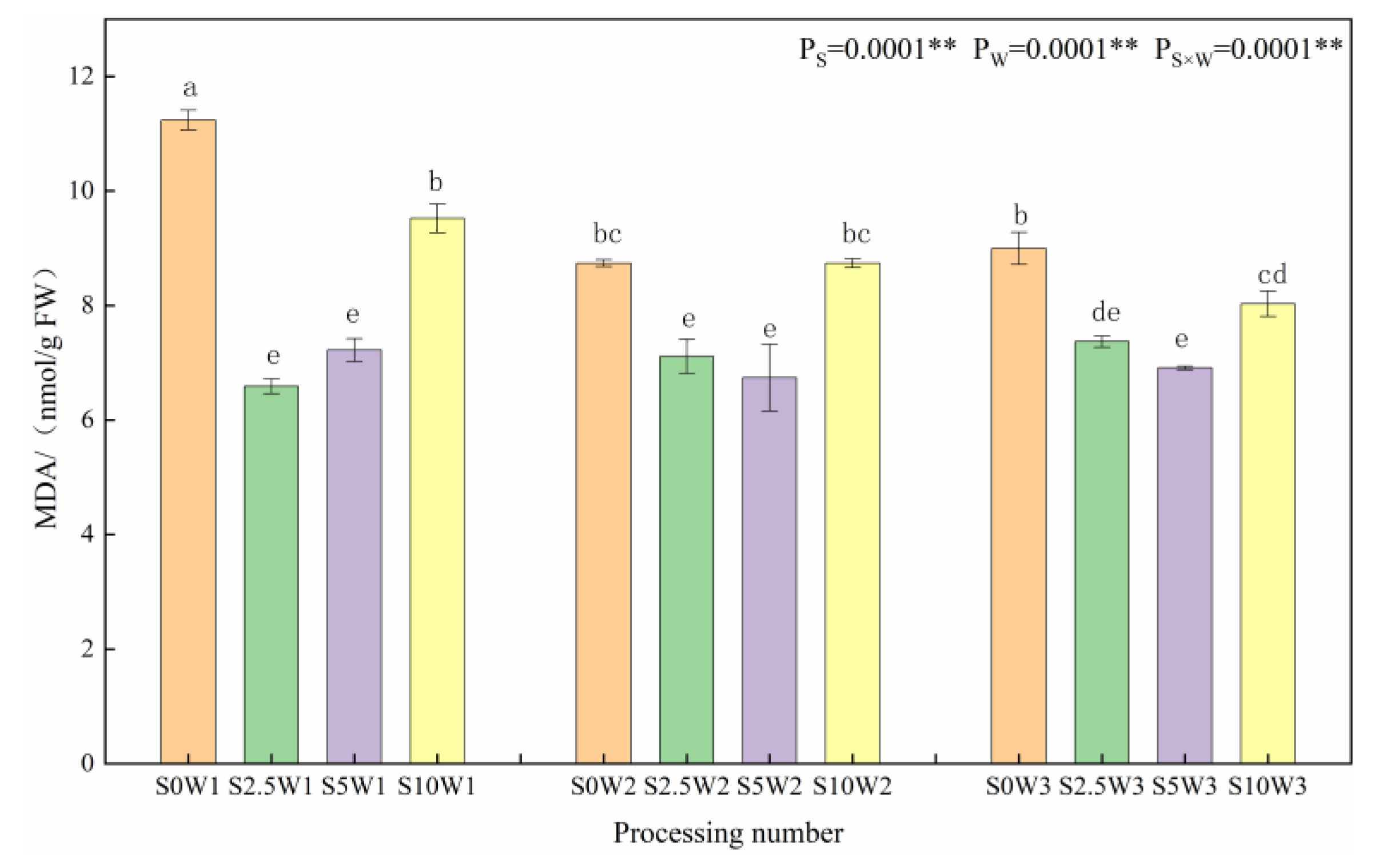
2.1.1. Leaf Malondialdehyde Content
2.1.2. Leaf Glutathione Peroxidase Activity
2.1.3. Leaf Superoxide Dismutase, Peroxidase, and Catalase Activities
2.2. Effect of Se on Osmoregulatory Substances in Leaves
2.2.1. Leaf Proline Content
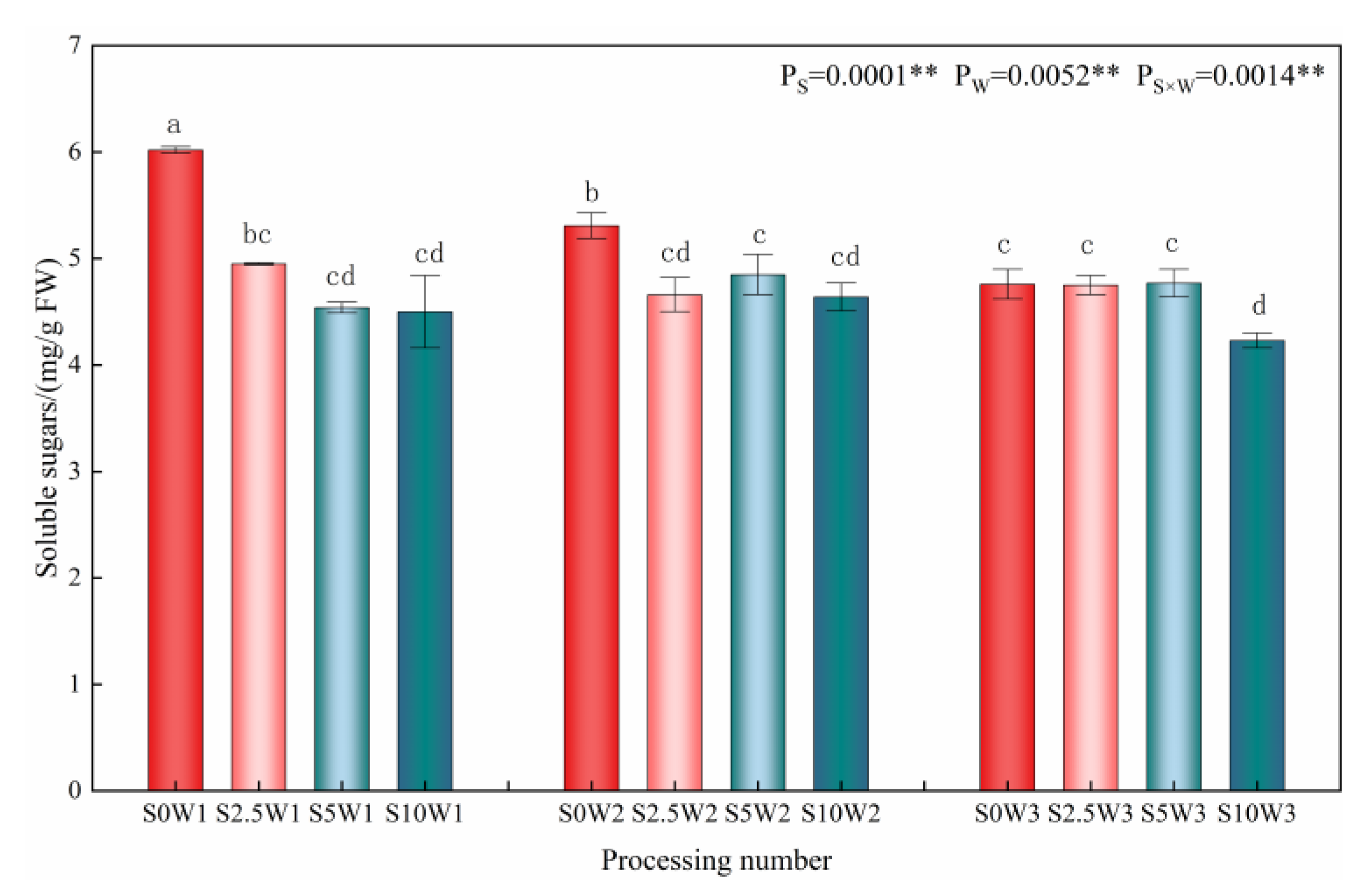
2.2.2. Leaf Soluble Sugar Content
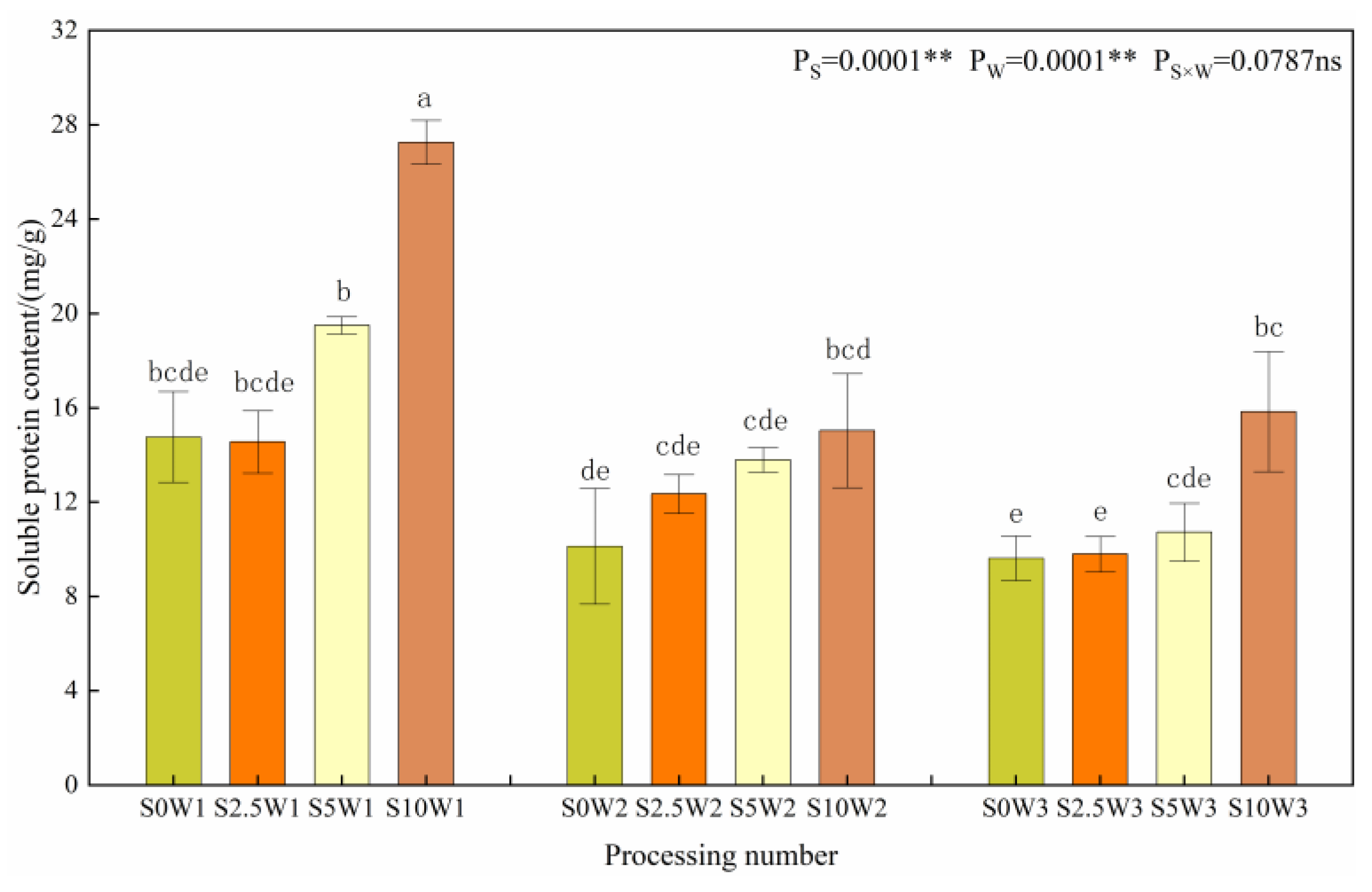
2.2.3. Leaf Soluble Protein Content
2.3. Effect of Se on Photosynthetic Characteristics of Tomato
2.3.1. Leaf Photosynthesis
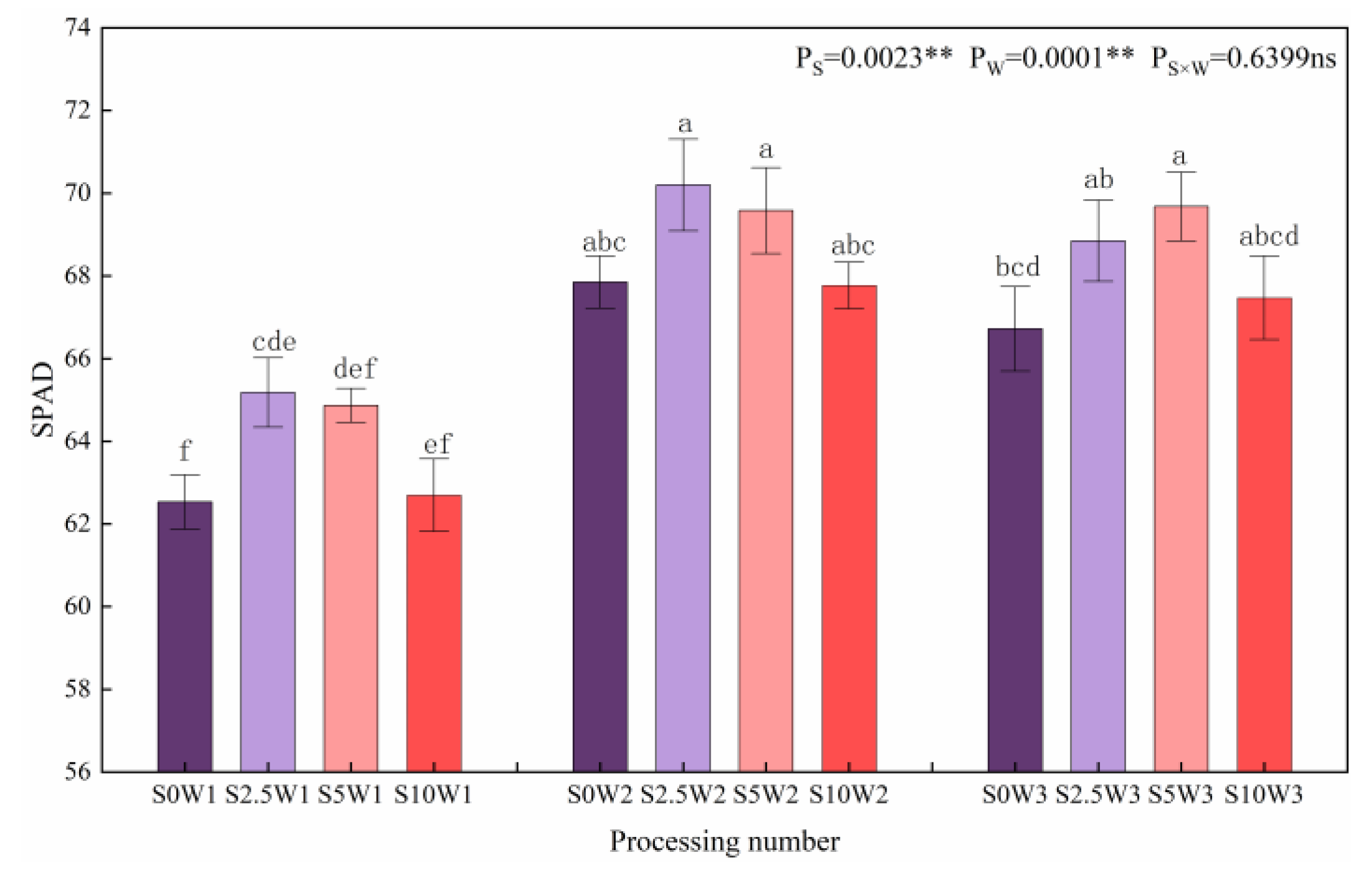
2.3.2. Relative Amount of Chlorophyll in Leaves
2.4. Correlation Analysis of Indicators and TOPSIS Comprehensive Evaluation
2.4.1. Correlation Analysis of Indicators
2.4.2. TOPSIS-Based Analysis of Relationships between Treatments and Comprehensive Evaluation
3. Discussion
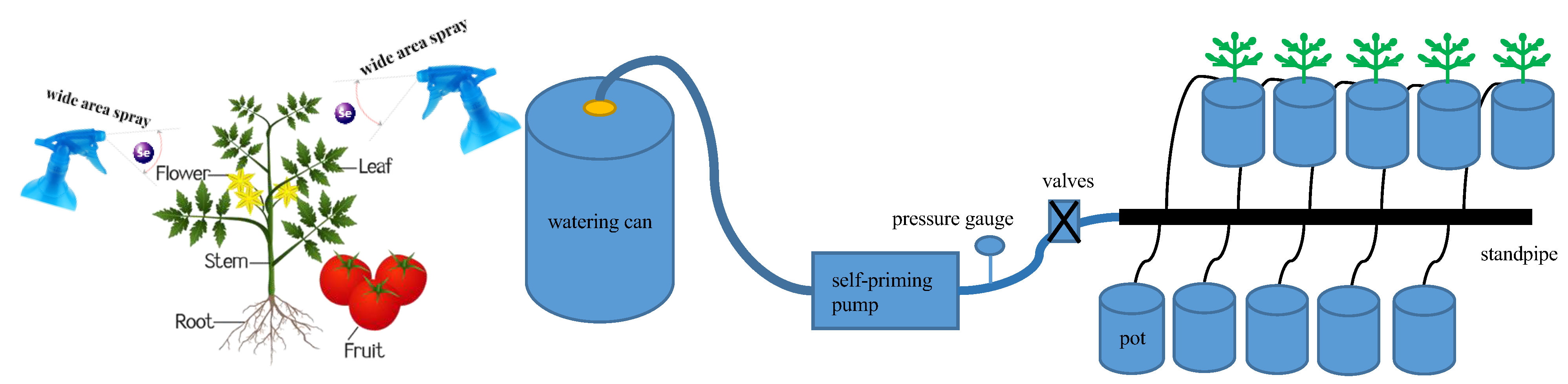
4. Materials and Methods
4.1. Overview of the Test Site
4.2. Experimental Design
4.3. Major Observation Programs and Methods
4.3.1. Soil Base Indicators
- (1)
- Soil capacity and field water-holding capacity: before the start of the experiment, the soil capacity and field water-holding capacity of the test field soil were determined by the ring knife method; since all the test soils were 20 cm topsoil, 0–20 cm ring knives were taken at different locations and repeated 6 times.
- (2)
- Soil base nutrients: before transplanting, air-dried and sieved test soils were taken for measuring total nitrogen, quick-acting nitrogen, phosphorus, potassium, pH value, conductivity, and organic matter; total nitrogen was measured by AA3 flow analyzer [48], fast-acting potassium was measured by flame photometry, fast-acting nitrogen was measured by alkaline dissolution and diffusion method, fast-acting phosphorus was measured by sodium bicarbonate leaching method, pH value was measured by potentiometric method, electrical conductivity was measured by DDS-307 conductivity meter method, and organic matter was measured by colorimetric method.
4.3.2. Physiological Indicators
- (1)
- Photosynthesis and other physiological indexes: a sunny and cloudless day was selected at the fruit ripening stage, and the leaf photosynthetic parameters were measured using the photosynthesis measurement system LI-6400 (LI-COR Inc., Lincoln, NE, USA) from 9:00–11:00 a.m.; the plants were selected to have the fourth well-grown functional leaf from top to bottom, and three representative plants were randomly selected for measurement in each treatment. The measurement items included photosynthetic physiological indexes such as net Pn, Tr, Gs, and Ci.
- (2)
- SPAD: at the same time as determining photosynthetic indexes, the SPAD value of functional leaves was determined by using a SPAD analyzer, and three plants with roughly the same growth were labeled for each treatment, and one leaf was labeled for each plant, and three points were measured and averaged.
4.3.3. Leaf Enzyme Activities and Osmoregulatory Substances
4.4. Data Processing
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Processing Number | Number of Fruits per Plant/N | Average Fruit Mass per Fruit/g | Yield per Plant/kg |
|---|---|---|---|
| S0W1 | 7.11 ± 0.42 c | 22.85 ± 0.47 ef | 0.162 ± 0.010 e |
| S2.5W1 | 7.33 ± 0.33 c | 26.82 ± 0.48 e | 0.197 ± 0.010 e |
| S5W1 | 7.89 ± 0.51 c | 23.61 ± 0.72 ef | 0.185 ± 0.010 e |
| S10W1 | 8.11 ± 0.42 c | 22.18 ± 0.57 f | 0.179 ± 0.008 e |
| S0W2 | 10.33 ± 0.41 b | 81.67 ± 1.60 d | 0.844 ± 0.036 d |
| S2.5W2 | 10.67 ± 0.37 ab | 89.41 ± 1.36 c | 0.955 ± 0.041 c |
| S5W2 | 11.44 ± 0.47 ab | 82.44 ± 1.31 d | 0.940 ± 0.034 c |
| S10W2 | 11.11 ± 0.35 ab | 81.04 ± 1.13 d | 0.899 ± 0.027 cd |
| S0W3 | 11.00 ± 0.37 ab | 129.92 ± 2.07 ab | 1.427 ± 0.046 b |
| S2.5W3 | 11.67 ± 0.17 a | 133.95 ± 2.20 a | 1.562 ± 0.028 a |
| S5W3 | 11.56 ± 0.18 a | 131.50 ± 2.09 ab | 1.519 ± 0.028 a |
| S10W3 | 11.00 ± 0.29 ab | 128.49 ± 2.36 b | 1.413 ± 0.042 b |
| W | 0.0001 *** | 0.0001 *** | 0.0001 *** |
| S | 0.0584 ns | 0.0001 *** | 0.0005 *** |
| S × W | 0.5629 ns | 0.7580 ns | 0.2946 ns |
References
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef] [PubMed]
- Ghorai, M.; Kumar, V.; Kumar, V.; Al-Tawaha, A.R.; Shekhawat, M.S.; Pandey, D.K.; Batiha, G.E.; Bursal, E.; Jha, N.K.; Gadekar, V.S.; et al. Beneficial Role of Selenium (Se) Biofortification in Developing Resilience Against Potentially Toxic Metal and Metalloid Stress in Crops: Recent Trends in Genetic Engineering and Omics Approaches. J. Soil Sci. Plant Nutr. 2022, 22, 2347–2377. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Y.; Huang, Y.; Lin, Z.; Bañuelos, G.S.; Lam, M.H.; Yin, X. Daily selenium intake in a moderate selenium deficiency area of Suzhou, China. Food Chem. 2011, 126, 1088–1093. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Pezzarossa, B. Selenium Enrichment of Horticultural Crops. Molecules 2017, 22, 933. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Conesa, M.À.; Douthe, C.; El Aou-Ouad, H.; Ribas-Carbó, M.; Galmés, J. Tomato landraces as a source to minimize yield losses and improve fruit quality under water deficit conditions. Agric. Water Manag. 2019, 223, 105722. [Google Scholar] [CrossRef]
- Lu, J.; Shao, G.; Cui, J.; Wang, X.; Keabetswe, L. Yield, fruit quality and water use efficiency of tomato for processing under regulated deficit irrigation: A meta-analysis. Agric. Water Manag. 2019, 222, 301–312. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Ning, H.; Zhang, X.; Li, S.; Pang, J.; Wang, G.; Sun, J. Optimizing irrigation frequency and amount to balance yield, fruit quality and water use efficiency of greenhouse tomato. Agric. Water Manag. 2019, 226, 105787. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, Y.; Zhang, X.; Li, M. Effect of foliar treatment of sodium selenate on postharvest decay and quality of tomato fruits. Sci. Hortic. 2016, 198, 304–310. [Google Scholar] [CrossRef]
- Rady, M.M.; Belal, H.E.E.; Gadallah, F.M.; Semida, W.M. Selenium application in two methods promotes drought tolerance in Solanum lycopersicum plant by inducing the antioxidant defense system. Sci. Hortic. 2020, 266, 109290. [Google Scholar] [CrossRef]
- Garza-García, J.J.O.; Hernández-Díaz, J.A.; Zamudio-Ojeda, A.; León-Morales, J.M.; Guerrero-Guzmán, A.; Sánchez-Chiprés, D.R.; López-Velázquez, J.C.; García-Morales, S. The Role of Selenium Nanoparticles in Agriculture and Food Technology. Biol. Trace Elem. Res. 2022, 200, 2528–2548. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Lu, C.D.; Shen, H.Y.; Liang, Y.; Chen, Z.H.; Zhong, J.H.; Tan, J.J.; Wang, X.Y.; Liang, H.G. Adaptive capacity and physiological response to high-temperature stress in the downy parasitic armor. Chin. J. Biol. Control 2021, 37, 1179–1188. [Google Scholar]
- Shuang, L.; Mounkaila, H.A.K.; Yingying, Z.; Yueping, L.; Yang, G.; Aiwang, D. Coordination of leaf hydraulic, anatomical, and economical traits in tomato seedlings acclimation to long-term drought. BMC Plant Biol. 2021, 21, 536. [Google Scholar]
- Pezzarossa, B.; Rosellini, I.; Borghesi, E.; Tonutti, P.; Malorgio, F. Effects of Se-enrichment on yield, fruit composition and ripening of tomato (Solanum lycopersicum) plants grown in hydroponics. Sci. Hortic. 2014, 165, 106–110. [Google Scholar] [CrossRef]
- Hemmati, M.; Delkhosh, B.; Shirani Rad, A.H.; Noor Mohammadi, G.; Söylemezoğlu, G. Effect of the Application of Foliar Selenium on Canola Cultivars as Influenced by Different Irrigation Regimes. Ank. Üniversitesi Ziraat Fakültesi Tarım Bilim. Derg. 2019, 25, 309–318. [Google Scholar] [CrossRef]
- Sattar, A.; Cheema, M.A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Nawaz, A.; Abbas, T.; Ali, Q. Physiological and biochemical attributes of bread wheat (Triticum aestivum L.) seedlings are influenced by foliar application of silicon and selenium under water deficit. Acta Physiol. Plant 2019, 41, 146. [Google Scholar] [CrossRef]
- Saleem, M.; Fariduddin, Q. Novel mechanistic insights of selenium induced microscopic, histochemical and physio-biochemical changes in tomato (Solanum lycopersicum L.) plant. An account of beneficiality or toxicity. J. Hazard. Mater. 2022, 434, 128830. [Google Scholar] [CrossRef]
- Hajlaoui, F.; Hajlaoui, H.; Krouma, A. Physio-Biochemical Response to Exogenous Selenium Application of Tomatoes (Solanum lycopersicum L.) Cultivated in the Field under Saline Irrigation. Russ. J. Plant Physiol. 2023, 70, 142. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Adamakis, I.D.S.; Şaş, B.; İşgören, S.; Moustaka, J.; Morales, F. Mechanistic Approach on Melatonin-Induced Hormesis of Photosystem II Function in the Medicinal PlantMentha spicata. Plants 2023, 12, 4025. [Google Scholar] [CrossRef]
- Rezaei, M.A.; Jokar, I.; Ghorbanli, M.; Kaviani, B.; Kharabian-Masouleh, A. Morpho-physiological improving effects of exogenous glycine betaine on tomato (Lycopersicum esculentum Mill.) cv. PS under drought stress conditions. Plant Omics 2012, 5, 79–86. [Google Scholar]
- Mohamad, A.H.; Marina, M.F.; José, R.S.F.; Oscar, V.; Monica, B. Effects of Salt and Water Stress on Plant Growth and on Accumulation of Osmolytes and Antioxidant Compounds in Cherry Tomato. Not. Bot. Horti Agrobot. Cluj-Na 2015, 43, 1–11. [Google Scholar]
- Song, J.W.; Li, H.H.; Xin, L.; Shen, J.; Liu, H.; Wang, X.P.; Zhong, Y. Effects of foliar selenium application and soil moisture interaction on tomato growth and water use efficiency in greenhouse. J. Irrig. Drain. 2023, 42, 52–59. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Khan, R.; Ma, X.; Zhang, J.; Wu, X.; Iqbal, A.; Wu, Y.; Zhou, L.; Wang, S. Circular drought-hardening confers drought tolerance via modulation of the antioxidant defense system, osmoregulation, and gene expression in tobacco. Physiol. Plant. 2021, 172, 1073–1088. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide Dismutase and Stress Tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Kozi, A. The Water-Water Cycle in Chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.R.; Pereira, A. Molecular and Physiological Analysis of Drought Stress in Arabidopsis Reveals Early Responses Leading to Acclimation in Plant Growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef]
- Haodong, L.; Chunmei, X.; Tianci, Q.; Jie, D.; Hua, C.; Xin, C.; Shuiyuan, C.; Shen, R.; Yue, Z. Selenium Regulates Antioxidant, Photosynthesis, and Cell Permeability in Plants under Various Abiotic Stresses: A Review. Plants 2022, 12, 44. [Google Scholar]
- Qi, J.E.A. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef]
- Giti Verma, D.S.P.T. ROS Modulation in Crop Plants Under Drought Stress. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Wiley: Hoboken, NJ, USA, 2019; pp. 311–336. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Induction of Drought Tolerance in Maize (Zea mays L.) due to Exogenous Application of Trehalose: Growth, Photosynthesis, Water Relations and Oxidative Defence Mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Rady, M.M.; Hemida, K.A. Sequenced application of ascorbate-proline-glutathione improves salt tolerance in maize seedlings. Ecotoxicol. Environ. Saf. 2016, 133, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen Peroxide in Plants: A Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross Talk between H2O2 and Interacting Signal Molecules under Plant Stress Response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef]
- Christine, H.F.; Maud, L.; Karl, J.K. Photooxidative stress in plants. Physiol. Plant. 1994, 92, 696–717. [Google Scholar]
- Morales-Espinoza, M.C.; Cadenas-Pliego, G.; Pérez-Alvarez, M.; Hernández-Fuentes, A.D.; Cabrera De La Fuente, M.; Benavides-Mendoza, A.; Valdés-Reyna, J.; Juárez-Maldonado, A. Se Nanoparticles Induce Changes in the Growth, Antioxidant Responses, and Fruit Quality of Tomato Developed under NaCl Stress. Molecules 2019, 24, 3030. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Hajiboland, R.; Sadeghzadeh, N.; Sadeghzadeh, B. Effect of Se application on photosynthesis, osmolytes and water relations in two durum wheat (Triticum durum L.) genotypes under drought stress. Acta Agric. Slov. 2015, 103, 167–179. [Google Scholar] [CrossRef]
- Ma, C.C.; Gao, Y.B.; Li, Q.F.; Guo, H.Y.; Chen, D.; Dou, S.; Wang, J.L. Morphological and physiological ecological adap-tations of the genus Cinchona on the Inner Mongolian Plateau. J. Ecol. 2011, 31, 6451–6459. [Google Scholar]
- Erika Mangiapane, A.P.E.P. Selenium and selenoproteins: An overview on different biological systems. Curr. Protein Pept. Sci. 2017, 15, 598–607. [Google Scholar] [CrossRef]
- Hu Ying, M.G.H.; Le Leu, R.K.; Woodman, R.; Young, G.P. Suppression of colorectal oncogenesis by selenium-enriched milk proteins: Apoptosis and K-ras mutations. Cancer Res. 2008, 68, 4936–4944. [Google Scholar]
- Dong, Z.; Lin, Y.; Wu, H.; Zhang, M. Selenium accumulation in protein fractions of Tenebrio molitor larvae and the antioxidant and immunoregulatory activity of protein hydrolysates. Food Chem. 2021, 334, 127475. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Yan, H.; Mengzhuo, Z.; Junyou, L.; Xiangbin, X.; Xuequn, S.; Lanhuan, M. Slltpg3, a non-specific lipid transfer protein, acts on the cuticle synthetic pathway to delay water loss and softening of tomato fruit. Postharvest Biol. Technol. 2022, 188, 111899. [Google Scholar]
- Chen, M. Chlorophyll Modifications and Their Spectral Extension in Oxygenic Photosynthesis. Annu. Rev. Biochem. 2014, 83, 313–340. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.; Vasconcelos, M.W.; Soares, C.; Fidalgo, F.; Heuvelink, E.; Carvalho, S.M. Young Tomato Plants Respond Differently under Single or Combined Mild Nitrogen and Water Deficit: An Insight into Morphophysiological Responses and Primary Metabolism. Plants 2023, 12, 1181. [Google Scholar] [CrossRef]
- Li, H.H. Water and Nitrogen Interactions in Greenhouse Tomato and Optimization of Irrigation Nitrogen Application Mode. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2021. [Google Scholar]
- Liu, H.; Pang, J.; Li, H.H.; Qiang, X.M.; Zhang, Y.Y.; Song, J.W. Effects of foliar selenium spray coupling with soil moisture on tomato yield and quality. Chin. Agric. Sci. 2022, 55, 4433–4444. [Google Scholar]
- Li, S.; Hamani, A.K.M.; Si, Z.; Liang, Y.; Gao, Y.; Duan, A. Leaf Gas Exchange of Tomato Depends on Abscisic Acid and Jasmonic Acid in Response to Neighboring Plants under Different Soil Nitrogen Regimes. Plants 2020, 9, 1674. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; Sun, X.; Xing, S.; Cong, W.; Liu, X. Study on Dormancy Mechanism and Breaking Dormancy Method of Seeds. Am. J. Plant Sci. 2019, 10, 65–78. [Google Scholar] [CrossRef]




| Processing Number | Pn (μmol CO2 m−2 s−1) | Gs (mol H2O m−2 s−1) | Ci (μmol mol−1) | Tr (mmol H2O m−2 s−1) |
|---|---|---|---|---|
| S0W1 | 15.88 ± 1.08 d | 0.08 ± 0.01 e | 35.67 ± 16.86 c | 3.83 ± 0.39 e |
| S2.5W1 | 15.90 ± 0.31 d | 0.07 ± 0.00 e | 34.32 ± 3.55 c | 3.53 ± 0.10 e |
| S5W1 | 16.04 ± 0.29 d | 0.08 ± 0.00 e | 50.13 ± 11.98 c | 3.79 ± 0.04 e |
| S10W1 | 14.67 ± 1.16 d | 0.07 ± 0.01 e | 41.24 ± 10.76 c | 3.67 ± 0.28 e |
| S0W2 | 22.83 ± 0.52 c | 0.33 ± 0.03 cd | 264.4 ± 7.27 ab | 7.52 ± 0.28 d |
| S2.5W2 | 26.02 ± 0.18 b | 0.39 ± 0.02 bcd | 266.29 ± 4.04 ab | 9.58 ± 0.28 c |
| S5W2 | 28.50 ± 0.42 a | 0.39 ± 0.01 bcd | 252.96 ± 3.18 b | 10.28 ± 0.34 c |
| S10W2 | 23.71 ± 0.25 c | 0.30 ± 0.02 d | 240.65 ± 11.87 b | 9.27 ± 0.30 c |
| S0W3 | 26.69 ± 0.38 b | 0.42 ± 0.08 bc | 263.85 ± 16.77 ab | 11.45 ± 0.59 b |
| S2.5W3 | 29.58 ± 0.41 a | 0.62 ± 0.07 a | 293.87 ± 8.08 a | 13.38 ± 0.60 a |
| S5W3 | 30.00 ± 0.5 a | 0.46 ± 0.02 b | 265.33 ± 6.84 ab | 12.85 ± 0.36 a |
| S10W3 | 26.74 ± 0.31 b | 0.37 ± 0.03 bcd | 250.40 ± 10.31 b | 11.67 ± 0.32 b |
| W | 0.0001 ** | 0.0001 ** | 0.0001 ** | 0.0001 ** |
| S | 0.0001 ** | 0.0023 ** | 0.1353 ns | 0.0003 ** |
| S × W | 0.0039 ** | 0.031 * | 0.2357 ns | 0.0159 * |
| Correlation Coefficient | GSH-PX | CAT | SOD | POD | MDA | Proline | Soluble Sugar | Soluble Proteins | Pn | Gs | Ci | Tr | SPAD | Yield per Plant |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GSH-PX | 1 | 0.72 ** | −0.26 | −0.53 | −0.3 | −0.84 ** | −0.49 | −0.46 | 0.89 ** | 0.83 ** | 0.86 ** | 0.93 ** | 0.74 ** | 0.93 ** |
| CAT | 0.72 ** | 1 | −0.39 | −0.61 * | −0.62 * | −0.79 ** | −0.54 | −0.3 | 0.77 ** | 0.71 ** | 0.74 ** | 0.71 ** | 0.80 ** | 0.66 * |
| SOD | −0.26 | −0.39 | 1 | 0.21 | 0.55 | 0.17 | 0.78 ** | −0.29 | −0.24 | −0.28 | −0.16 | −0.28 | −0.25 | −0.27 |
| POD | −0.53 | −0.61 * | 0.21 | 1 | 0.60 * | 0.73 ** | 0.48 | 0.41 | −0.61 * | −0.48 | −0.63 * | −0.53 | −0.76 ** | −0.48 |
| MDA | −0.3 | −0.62 * | 0.55 | 0.60 * | 1 | 0.57 * | 0.54 | 0.24 | −0.42 | −0.35 | −0.31 | −0.34 | −0.65 * | −0.3 |
| Proline | −0.84 ** | −0.79 ** | 0.17 | 0.73 ** | 0.57 * | 1 | 0.37 | 0.74 ** | −0.92 ** | −0.88 ** | −0.92 ** | −0.89 ** | −0.94 ** | −0.88 ** |
| Soluble Sugar | −0.49 | −0.54 | 0.78 ** | 0.48 | 0.54 | 0.37 | 1 | −0.28 | −0.29 | −0.26 | −0.28 | −0.35 | −0.33 | −0.35 |
| Soluble proteins | −0.46 | −0.3 | −0.29 | 0.41 | 0.24 | 0.74 ** | −0.28 | 1 | −0.69 ** | −0.71 ** | −0.70 ** | −0.65 * | −0.67 * | −0.66 * |
| Pn | 0.89 ** | 0.77 ** | −0.24 | −0.61 * | −0.42 | −0.92 ** | −0.29 | −0.69 ** | 1 | 0.96 ** | 0.95 ** | 0.98 ** | 0.90 ** | 0.95 ** |
| Gs | 0.83 ** | 0.71 ** | −0.28 | −0.48 | −0.35 | −0.88 ** | −0.26 | −0.71 ** | 0.96 ** | 1 | 0.94 ** | 0.96 ** | 0.84 ** | 0.95 ** |
| Ci | 0.86 ** | 0.74 ** | −0.16 | −0.63 * | −0.31 | −0.92 ** | −0.28 | −0.70 ** | 0.95 ** | 0.94 ** | 1 | 0.93 ** | 0.89 ** | 0.92 ** |
| Tr | 0.93 ** | 0.71 ** | −0.28 | −0.53 | −0.34 | −0.89 ** | −0.35 | −0.65 * | 0.98 ** | 0.96 ** | 0.93 ** | 1 | 0.82 ** | 0.99 ** |
| SPAD | 0.74 ** | 0.80 ** | −0.25 | −0.76 ** | −0.65 * | −0.94 ** | −0.33 | −0.67 * | 0.90 ** | 0.84 ** | 0.89 ** | 0.82 ** | 1 | 0.77 ** |
| Yield per plant | 0.93 ** | 0.66 * | −0.27 | −0.48 | −0.3 | −0.88 ** | −0.35 | −0.66 * | 0.95 ** | 0.95 ** | 0.92 ** | 0.99 ** | 0.77 ** | 1 |
| Assessment Criteria | GSH-PX | CAT | SOD | MDA | Proline | Soluble Sugar | Pn | Gs | Ci | Tr | SPAD | Yield per Plant |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Weight | 0.0807 | 0.0628 | 0.0717 | 0.0734 | 0.0940 | 0.0625 | 0.0938 | 0.0814 | 0.1039 | 0.0947 | 0.0836 | 0.0976 |
| Processing Number | D+ | D− | C | Sorting Results |
|---|---|---|---|---|
| S0W1 | 0.329 | 0.065 | 0.165 | 12 |
| S2.5W1 | 0.255 | 0.159 | 0.384 | 10 |
| S5W1 | 0.249 | 0.181 | 0.421 | 9 |
| S10W1 | 0.299 | 0.135 | 0.311 | 11 |
| S0W2 | 0.162 | 0.205 | 0.559 | 8 |
| S2.5W2 | 0.096 | 0.272 | 0.738 | 4 |
| S5W2 | 0.098 | 0.265 | 0.729 | 6 |
| S10W2 | 0.139 | 0.218 | 0.611 | 7 |
| S0W3 | 0.098 | 0.265 | 0.73 | 5 |
| S2.5W3 | 0.088 | 0.307 | 0.778 | 2 |
| S5W3 | 0.075 | 0.292 | 0.795 | 1 |
| S10W3 | 0.091 | 0.271 | 0.749 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Xin, L.; Gao, F.; Liu, H.; Wang, X. Effects of Foliar Selenium Application on Oxidative Damage and Photosynthetic Properties of Greenhouse Tomato under Drought Stress. Plants 2024, 13, 302. https://doi.org/10.3390/plants13020302
Song J, Xin L, Gao F, Liu H, Wang X. Effects of Foliar Selenium Application on Oxidative Damage and Photosynthetic Properties of Greenhouse Tomato under Drought Stress. Plants. 2024; 13(2):302. https://doi.org/10.3390/plants13020302
Chicago/Turabian StyleSong, Jiawen, Lang Xin, Fukui Gao, Hao Liu, and Xingpeng Wang. 2024. "Effects of Foliar Selenium Application on Oxidative Damage and Photosynthetic Properties of Greenhouse Tomato under Drought Stress" Plants 13, no. 2: 302. https://doi.org/10.3390/plants13020302
APA StyleSong, J., Xin, L., Gao, F., Liu, H., & Wang, X. (2024). Effects of Foliar Selenium Application on Oxidative Damage and Photosynthetic Properties of Greenhouse Tomato under Drought Stress. Plants, 13(2), 302. https://doi.org/10.3390/plants13020302