
Effects of Extended Light/Dark Cycles on Solanaceae Plants

 ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Plant Growth and Development
2.2. Photosynthetic Pigments
2.3. Oxidative Stress and Antioxidants
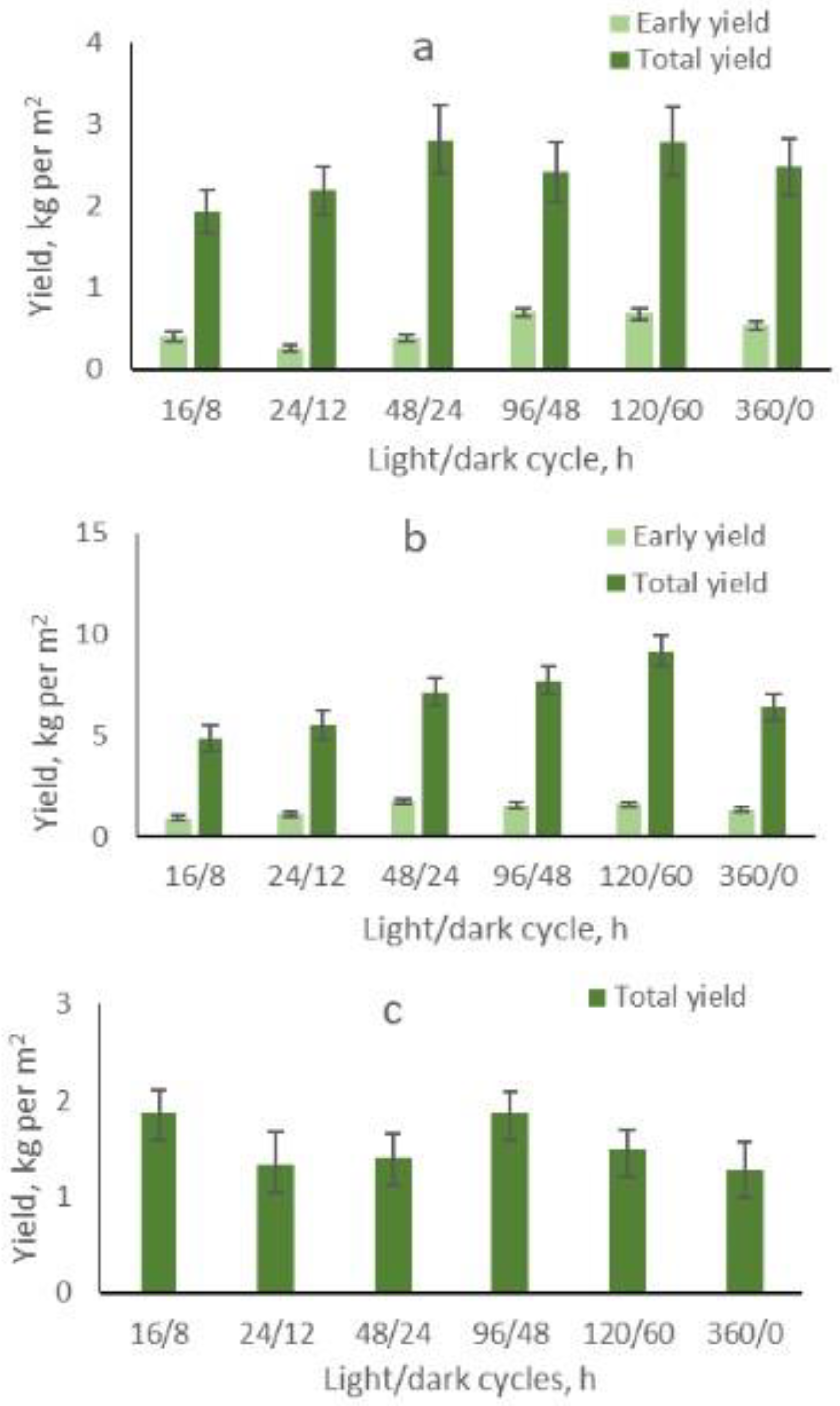
2.4. After-Effect of Extended Light/Dark Cycles on Fruit Yield
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Growth Conditions
5.2. Growth and Yield Measurements
5.3. Photosynthetic Pigment Content
5.4. Anthocyanins and Flavonoids Content
5.5. Malondialdehyde (MDA) Content
5.6. Hydrogen Peroxide Content
5.7. Data Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Despommier, D. The Vertical Farm: Feeding the World in the 21st Centure; Thomas Dunne Books: New York, NY, USA, 2010. [Google Scholar]
- Kozai, T.; Nui, G.; Takagaki, M. Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Academic Press: Cambridge, MA, USA, 2015; p. 516. [Google Scholar]
- Bartzas, G.; Zaharaki, D.; Komnitsas, K. Life cycle assessment of open field and greenhouse cultivation of lettuce and barley. Inf. Process. Agric. 2015, 2, 191–207. [Google Scholar] [CrossRef]
- Graamans, L.; Baeza, E.; van den Dobbelsteen, A.; Tsafaras, I.; Stanghellini, C. Plant factories versus greenhouses: Comparison of resource use efficiency. Agric. Syst. 2018, 160, 31–43. [Google Scholar] [CrossRef]
- SharathKumar, M.; Heuvelink, E.; Marcelis, L.F.M. Vertical farming: Moving from genetic to environmental modification. Trends Plant Sci. 2020, 25, 724–727. [Google Scholar] [CrossRef]
- van Delden, S.H.; Sharathkumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; et al. Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef]
- Inada, K.; Yabumoto, Y. Effect of light quality, day length and periodic temperature variation on the growth of lettuce and radish plants. Jpn. J. Crop Sci. 1989, 58, 689–694. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G. Role of the plant factory with artificial lighting (PFAL) in urban areas. In Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Kozai, T., Niu, G., Takagaki, M., Eds.; Academic Press: London, UK, 2020; pp. 7–33. [Google Scholar]
- Chen, X.; Li, Y.; Wang, L.; Yang, Q.; Guo, W. Responses of butter leaf lettuce to mixed red and blue light with extended light/dark cycle period. Sci. Rep. 2022, 12, 6924. [Google Scholar] [CrossRef] [PubMed]
- Bowsher, C.G.; Long, D.M.; Oaks, A.; Rothstein, S.J. Effect of light/dark cycles on expression of nitrate assimilatory genes in maize shoots and roots. Plant Physiol. 1991, 95, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.C.; Yang, T.Y.; Riskowskic, G.L. Ascorbic acid, nitrate, and nitrite concentration relationship to the 24 hour light/dark cycle for spinach grown in different condition. Food Chem. 2013, 138, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Kurata, H.; Achioku, T.; Furusaki, S. The light/dark cycle operation with an hour-scale period enhances caffeine production by Coffea arabica, cells. Enzyme Microb. Technol. 1998, 23, 518–523. [Google Scholar] [CrossRef]
- Chen, X.L.; Yang, Q.C. Effects of intermittent light exposure with red and blue light emitting diodes on growth and carbohydrate accumulation of lettuce. Sci. Hortic. 2018, 234, 220–226. [Google Scholar] [CrossRef]
- Sivakumar, G.; Heo, J.W.; Kozai, T.; Paek, K.Y. Effect of continuous or intermittent radiation on sweet potato plantlets in vitro. J. Hortic. Sci. Biotechnol. 2006, 81, 546–548. [Google Scholar] [CrossRef]
- Vaštakaite, V.; Viršile, A.; Brazaityte, A.; Samuoliene, G.; Jankauskiene, J.; Novickovas, A.; Duchovskis, P. Pulsed light-emitting diodes for a higher phytochemical level in microgreens. J. Agric. Food Chem. 2017, 65, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.S.; He, D.X.; He, J.; Niu, G.H.; Ji, F. Effects of short light/dark cycles on photosynthetic pathway switching and growth of medicinal Dendrobium officinale in aeroponic cultivation. Int. J. Agric. Biol. Eng. 2019, 12, 38–43. [Google Scholar] [CrossRef]
- Avgoustaki, D.D.; Bartzanas, T.; Xydis, G. Minimizing the energy footprint of indoor food production while maintaining a high growth rate: Introducing disruptive cultivation protocols. Food Control 2021, 130, 108290. [Google Scholar] [CrossRef]
- Nitschke, S.; Cortleven, A.; Iven, T.; Feussner, I.; Havaux, M.; Riefler, M.; Schmülling, T. Circadian stress regimes affect the circadian clock and cause jasmonicacid-dependent cell death in cytokinin-deficient Arabidopsis plants. Plant Cell. 2016, 28, 1616–1639. [Google Scholar]
- Nitschke, S.; Cortleven, A.; Schmülling, T. Novel stress in plants by altering the photoperiod. Trends Plant Sci. 2017, 22, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Holtgrefe, S.; Jung, S.; Wunrau, C.; Kandlbinder, A.; Baier, M.; Dietz, K.-J.; Backhausen, J.E.; Scheibe, R. Influence of the photoperiod on redox regulation and stress responses in Arabidopsis thaliana L. (Heynh.) plants under long- and short-day conditions. Planta 2006, 224, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Roeber, V.M.; Schmulling, T.; Cortleven, A. The photoperiod: Handling and Causing Stress in Plants. Front. Plant Sci. 2021, 12, 781988. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; van Poppel, P.M.J.A.; Heuvelink, E.; Millenaar, F.F. A single locus confer tolerance to continuous light and allows substantial yield increase in tomato. Nat. Commun. 2014, 5, 4549. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Highkin, H.R.; Hanson, J.B. Possible interaction between light-dark cycles and endogenous daily rhythms on the growth of tomato plants. Plant Physiol. 1954, 29, 301–302. [Google Scholar] [CrossRef] [PubMed]
- Abuelsoud, W.; Cortleven, A.; Schmülling, T. Photoperiod stress induces an oxidative burst-like response and is associated with increased apoplastic peroxidase and decreased catalase activities. J. Plant Physiol. 2020, 253, 153252. [Google Scholar] [CrossRef] [PubMed]
- Krasensky-Wrzaczek, J.; Kangasjarvi, J. The role of reactive oxygen species in the integration of temperature and light signals. J. Exp. Bot. 2018, 69, 3347–3358. [Google Scholar] [CrossRef] [PubMed]
- Shibaeva, T.G.; Mamaev, A.V.; Titov, A.F. Possible physiological mechanisms of leaf photodamage in plants grown under continuous lighting. J. Plant Physiol. 2023, 70, 15. [Google Scholar] [CrossRef]
- Simon, N.M.L. The circad ian clock influences the long-term water use efficiency of Arabidopsis. Plant Physiol. 2020, 183, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Kopsell, D.E. Genetic and environmental factors affecting plant lutein/zeaxanthin. Agro Food Ind. Hi-Tech. 2008, 19, 44–46. [Google Scholar]
- Perez-Balibrea, S.; Moreno, D.A.; Garcia-Viguera, C. Influence of light on health promoting phytochemicals of broccoli sprouts. J. Sci. Food Agric. 2008, 88, 904–910. [Google Scholar] [CrossRef]
- Keller, M.; Hrazdina, G. Interaction of nitrogen availability during bloom and light intensity during veraison. II. Effects on anthocyanin and phenolic development during grape ripening. Am. J. Enol. Vitic. 1998, 49, 341–349. [Google Scholar] [CrossRef]
- Liu, W.; Zha, L.; Zhang, Y. Growth and nutrient element content of hydroponic lettuce are modified by LED continuous lighting of different intensities and spectral qualities. Agronomy 2020, 10, 1678. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A.; Auge, R.M.; Both, A.J. Biomass production and pigment accumulation in kale grown under increasing photoperiods. HortScience 2006, 41, 603–606. [Google Scholar] [CrossRef]
- Ali, M.B.; Khandaker, L.; Oba, S. Comparative study on functional components, antioxidant activity and color parameters of selected colored leafy vegetables as affected by photoperiods. J. Food Agric. Environ. 2009, 7, 392–398. [Google Scholar]
- Bian, Z.-H.; Cheng, R.-F.; Yang, Q.-C.; Wang, J.; Lu, C. Continuous light from red, blue, and green light-emitting diodes reduces nitrate content and enhances phytochemical concentrations and antioxidant capacity in lettuce. J. Am. Soc. Hort. Sci. 2016, 141, 186. [Google Scholar] [CrossRef]
- Ohtake, N.; Ishikura, M.; Suzuki, H. Continuous irradiation with alternating red and blue light enhances plant growth while keeping nutritional quality in lettuce. Hort. Sci. 2018, 53, 1804. [Google Scholar] [CrossRef]
- Shibaeva, T.G.; Sherudilo, E.G.; Rubaeva, A.A.; Titov, A.F. Continuous LED lighting enhances yield and nutritional value of four genotypes of Brassicaceae microgreens. Planta 2022, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Alrajhi, A.A.; Alsahli, A.S.; Alhelal, I.M.; Rihan, H.Z.; Fuller, M.P.; Alsadon, A.A.; Ibrahim, A.A. The Effect of LED light spectra on the growth, yield and nutritional value of red and green lettuce (Lactuca sativa). Plants 2023, 12, 463. [Google Scholar] [CrossRef]
- Podsedek, A.; Fraszczak, B.; Sosnowska, D.; Kajszczak, D.; Szymczak, K.; Bonikowski, R. LED light quality affected bioactive compounds, antioxidant potential, and nutritional value of red and white cabbage microgreens. Appl. Sci. 2023, 13, 5435. [Google Scholar] [CrossRef]
- Fraszczak, B.; Kula-Maximenko, M.; Podsedek, A.; Sosnowska, D.; Unegbu, K.C.; Spizewski, T. Morphological and Photosynthetic Parameters of Green and Red Kale Microgreens Cultivated under Different Light Spectra. Plants 2023, 12, 3800. [Google Scholar] [CrossRef]
- Kang, J.H.; Sugumaran, K.; Atulba, S.L.S.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hort. Environ. Biotechnol. 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Steyn, W.J.; Wand, S.J.E. Anthocyanins in vegetative tissues: A proposed unified function in photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Timmins, G.S.; Holbrook, N.M.; Field, T.S. Le rouge et le noir:Are anthocyanins plant melanins? Adv. Bot. Res. 2002, 37, 17–35. [Google Scholar]
- Nielsen, S.L.; Simonsen, A.M. Photosynthesis and photoinhibition in two differently coloured varieties of Oxalis triangularis—The effect of anthocyanin content. Photosynthetica 2011, 49, 346–352. [Google Scholar] [CrossRef]
- Trojak, M.; Skowron, E. Role of anthocyanins in high-light stress response. World Sci. News 2017, 81, 150–168. [Google Scholar]
- Havaux, M.; Kloppstech, K. The protective functions of carotenoid and flavonoid pigments against excess visible radiation at chilling temperature investigated in Arabidopsisnpq and tt mutants. Planta 2001, 213, 953. [Google Scholar] [CrossRef]
- Neill, S.O.; Gould, K.S. Anthocyanins in leaves: Light attenuators or antioxidants? Funct. Plant Biol. 2003, 30, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Zang, K.-M.; Yu, H.-J.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q.; Xia, X.-J. Photoprotective roles of anthocyanins in Begonia semperflorens. Plant Sci. 2010, 179, 202–208. [Google Scholar] [CrossRef]
- Zhang, T.-J.; Chow, W.S.; Liu, X.-T.; Zhang, P.; Liu, N.; Peng, C.-L. A magic red coat on the surface of young leaves: Anthocyanins distributed in trichome layer protect Castanopsis fissa leaves from photoinhibition. Tree Physiol. 2016, 36, 1296–1306. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, T.J.; Zheng, J. Anthocyanins function as a light attenuator to compensate for insufficient photoprotection mediated by nonphotochemical quenching in young leaves of Acmena acuminatissima in winter. Photosynthetica 2018, 56, 445–454. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The case for anthocyanin consumption to promote human health: A review. Comp. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Llorente, B.; Martinez-Garcia, J.; Stange, C.; Rodriguez-Concepcion, M. Illuminating colors: Regulation of carotenoid biosynthesis and accumulation by light. Curr. Opin. Plant Biol. 2017, 37, 49–55. [Google Scholar] [CrossRef]
- Maslova, T.G.; Markovskaya, E.F.; Slemnev, N.N. Functions of carotenoids in the leaves of higher plants (review). Zh. Obsch. Biol. 2020, 81, 297–310. [Google Scholar] [CrossRef]
- Shibaeva, T.G.; Mamaev, A.V.; Sherudilo, E.G.; Titov, A.F. The role of photosyntheticdaily light integral in plant response to extended photoperiods. Russ. J. Plant Physiol. 2022, 69, 1–8. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigment of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Meng, X.; Xing, T.; Wang, X. The role of light in the regulation of anthocyanin accumulation in Gerbera hybrid. Plant Growth Regul. 2004, 44, 243–250. [Google Scholar] [CrossRef]
- Nogues, S.; Backer, N.R. Effect of drought on photosynthesis in Mediterranean plants under UV-B radiation. J. Exp. Bot. 2000, 51, 1309–1317. [Google Scholar]
- Baroowa, B.; Gogoi, N. Biochemical changes in two Vigna spp. during drought and subsequent recovery. Ind. J. Plant Physiol. 2013, 18, 319–325. [Google Scholar] [CrossRef]
- Kolupaev, Y.E.; Fisova, E.N.; Yastreb, T.O.; Ryabchun, N.I.; Kirichenko, V.V. Effect of hydrogen sulfide donor on antioxidant state of wheat plants and their resistance to soil drought. Russ. J. Plant Physiol. 2019, 66, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperioxidation in isolated chloroplasts I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Velikova, V.; Edreva, A. Oxidative stress and some antioxidant system in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]

{kind=link}
| Light/Dark Cycle, h | Leaf Area, dm2 | LMA, mg cm−2 | Leaf Number | Chl a + b, mg gDW−1 | Car, mg gDW−1 |
|---|---|---|---|---|---|
| Eggplant | |||||
| 16/8 | 8.2 ± 0.8 | 1.71 ± 0.26 | 8.3 ± 0.2 | 17.1 ± 1.8 | 2.1 ± 0.3 |
| 24/12 | 7.5 ± 0.7 | 2.22 ± 0.17 * | 8.0 ± 0.2 | 14.7 ± 0.3 * | 1.8 ± 0.3 |
| 48/24 | 7.2 ± 0.6 * | 2.56 ± 0.43 * | 6.9 ± 0.2 * | 14.0 ± 2.8 * | 1.8 ± 0.1 |
| 96/48 | 6.0 ± 0.6 * | 2.30 ± 0.11 * | 7.9 ± 0.3 | 12.3 ± 2.1 * | 1.8 ± 0.4 |
| 120/60 | 5.0 ± 0.6 * | 2.44 ± 0.17 * | 7.1 ± 0.2 * | 12.0 ± 0.9 * | 1.6 ± 0.3 * |
| 360/0 | 7.5 ± 0.8 | 3.22 ± 0.42 * | 8.6 ± 0.2 * | 8.9 ± 1.0 * | 1.1 ± 0.1 * |
| Pepper | |||||
| 16/8 | 6.1 ± 0.6 | 1.78 ± 0.06 | 15.3 ± 0.6 | 19.9 ± 1.0 | 2.5 ± 0.2 |
| 24/12 | 5.5 ± 0.6 | 2.09 ± 0.07 * | 16.8 ± 0.5 | 14.5 ± 1.2 * | 1.9 ± 0.1 * |
| 48/24 | 6.3 ± 0.5 | 2.88 ± 0.25 * | 16.0 ± 0.8 | 12.9 ± 0.8 * | 1.6 ± 0.1 * |
| 96/48 | 6.0 ± 0.6 | 2.40 ± 0.08 * | 13.4 ± 0.8 * | 10.9 ± 3.3 * | 1.5 ± 0.5 * |
| 120/60 | 3.5 ± 0.4 * | 2.93 ± 0.11 * | 14.3 ± 0.4 * | 12.7 ± 1.7 * | 1.6 ± 0.1 * |
| 360/0 | 5.5 ± 0.6 | 2.76 ± 0.12 * | 16.0 ± 0.6 | 9.5 ± 0.6 * | 1.4 ± 0.1 * |
| Tobacco | |||||
| 16/8 | 4.5 ± 0.5 | 3.13 ± 0.30 | 7.1 ± 0.2 | 9.9 ± 0.3 | 1.2 ± 0.1 |
| 24/12 | 6.8 ± 0.6 * | 2.90 ± 0.39 | 7.9 ± 0.3 * | 7.5 ± 0.6 * | 0.9 ± 0.1 * |
| 48/24 | 3.4 ± 0.4 * | 2.59 ± 0.18 | 6.7 ± 0.3 | 4.5 ± 0.6 * | 0.5 ± 0.3 * |
| 96/48 | 2.6 ± 0.3 * | 1.94 ± 0.18 * | 5.8 ± 0.3 * | 6.0 ± 1.1 * | 0.8 ± 0.1 |
| 120/60 | 5.4 ± 0.5 * | 1.63 ± 0.17 * | 7.8 ± 0.3 * | 6.5 ± 0.3 * | 0.8 ± 0.1 * |
| 360/0 | 4.4 ± 0.4 | 5.02 ± 0.28 * | 6.6 ± 0.3 | 5.5 ± 0.3 * | 0.6 ± 0.1 * |
| Tomato | |||||
| 16/8 | 8.8 ± 0.7 | 2.92 ± 0.44 | 12.0 ± 0.3 | 19.6 ± 0.3 | 1.4 ± 0.1 |
| 24/12 | 10.4 ± 0.9 * | 2.42 ± 0.45 | 11.5 ± 0.5 | 16.2 ± 1.2 * | 1.3 ± 0.1 |
| 48/24 | 9.1 ± 0.8 | 2.75 ± 0.20 | 12.0 ± 0.2 | 10.3 ± 1.9 * | 1.5 ± 0.1 |
| 96/48 | 9.2 ± 0.9 | 2.57 ± 0.43 | 12.1 ± 0.3 | 13.9 ± 2.0 * | 1.4 ± 0.1 |
| 120/60 | 7.5 ± 0.7 | 2.85 ± 0.38 | 11.2 ± 0.5 | 18.2 ± 5.7 | 1.4 ± 0.1 |
| 360/0 | 12.1 ± 0.9 * | 3.88 ± 0.36 * | 13.2 ± 0.3 * | 7.8 ± 0.5 * | 0.9 ± 0.1 * |
| Light/Dark Cycle, h | H2O2, mg gDW−1 | MDA, µmol gDW−1 | Anthocyanins, (A530 − 0.25A657) gFW−1 | Flavonoids, A300gFW−1 |
|---|---|---|---|---|
| Eggplant | ||||
| 16/8 | 0.33 ± 0.04 | 144.5 ± 10.5 | 0.54 ± 0.04 | 22.2 ± 2.1 |
| 24/12 | 0.38 ± 0.03 | 173.0 ± 13.3 * | 0.58 ± 0.11 | 22.4 ± 0.3 |
| 48/24 | 0.41 ± 0.06 * | 163.9 ± 15.0 * | 0.53 ± 0.05 | 37.1 ± 3.5 * |
| 96/48 | 0.47 ± 0.02 * | 189.3 ± 17.4 * | 0.55 ± 0.04 | 40.6 ± 2.7 * |
| 120/60 | 0.53 ± 0.06 * | 163.1 ± 4.7 * | 0.51 ± 0.03 | 33.5 ± 2.9 * |
| 360/0 | 0.49 ± 0.01 * | 172.3 ± 15.5 * | 0.50 ± 0.03 | 26.2 ± 1.1 * |
| Pepper | ||||
| 16/8 | 0.40 ± 0.02 | 126.9 ± 13.1 | 0.69 ± 0.04 | 24.6 ± 1.6 |
| 24/12 | 0.42 ± 0.02 | 142.0 ± 8.5 | 0.60 ± 0.08 | 29.0 ± 3.8 * |
| 48/24 | 0.44 ± 0.02 | 127.5 ± 14.0 | 0.66 ± 0.05 | 36.6 ± 2.9 * |
| 96/48 | 0.61 ± 0.02 * | 151.0 ± 6.1 | 0.52 ± 0.06 * | 34.0 ± 2.9 * |
| 120/60 | 0.58 ± 0.06 * | 152.5 ± 12.8 | 0.61 ± 0.06 | 37.7 ± 3.6 * |
| 360/0 | 0.47 ± 0.02 * | 175.5 ± 13.5 * | 0.94 ± 0.02 * | 30.8 ± 1.4 * |
| Tobacco | ||||
| 16/8 | 0.23 ± 0.02 | 138.3 ± 13.0 | 1.08 ± 0.14 | 106.5 ±2.8 |
| 24/12 | 0.31 ± 0.05 * | 123.6 ± 14.6 | 1.00 ± 0.08 * | 93.5 ± 7.4 |
| 48/24 | 0.32 ± 0.07 * | 215.9 ± 22.7 * | 0.95 ± 0.21 * | 97.3 ± 9.5 |
| 96/48 | 0.26 ± 0.04 | 213.6 ± 7.0 * | 1.07 ± 0.18 | 87.7 ± 4.0 * |
| 120/60 | 0.36 ± 0.04 * | 200.0 ± 9.0 * | 0.90 ± 0.12 * | 78.4 ± 9.9 * |
| 360/0 | 0.31 ± 0.05 * | 169.8 ± 25.0 * | 1.23 ± 0.09 | 142.0 ± 3.9 * |
| Tomato | ||||
| 16/8 | 0.47 ± 0.02 | 164.9 ± 12.1 | 0.86 ± 0.13 | 33.9 ± 3.9 |
| 24/12 | 0.62 ± 0.12 * | 148.7 ± 18.4 | 0.87 ± 0.02 | 34.9 ± 1.3 |
| 48/24 | 0.78 ± 0.11 * | 194.6 ± 27.6 * | 1.18 ± 0.07 * | 34.9 ± 2.0 |
| 96/48 | 0.69 ± 0.06 * | 195.1 ± 40.4 * | 2.12 ± 0.30 * | 41.8 ± 3.8 * |
| 120/60 | 0.75 ± 0.07 * | 250.7 ± 27.5 * | 2.15 ± 0.30 * | 36.7 ± 3.4 * |
| 360/0 | 0.85 ± 0.05 * | 253.9 ± 24.5 * | 1.74 ± 0.15 * | 41.6 ± 2.2 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibaeva, T.G.; Sherudilo, E.G.; Ikkonen, E.; Rubaeva, A.A.; Levkin, I.A.; Titov, A.F. Effects of Extended Light/Dark Cycles on Solanaceae Plants. Plants 2024, 13, 244. https://doi.org/10.3390/plants13020244
Shibaeva TG, Sherudilo EG, Ikkonen E, Rubaeva AA, Levkin IA, Titov AF. Effects of Extended Light/Dark Cycles on Solanaceae Plants. Plants. 2024; 13(2):244. https://doi.org/10.3390/plants13020244
Chicago/Turabian StyleShibaeva, Tatjana G., Elena G. Sherudilo, Elena Ikkonen, Alexandra A. Rubaeva, Ilya A. Levkin, and Alexander F. Titov. 2024. "Effects of Extended Light/Dark Cycles on Solanaceae Plants" Plants 13, no. 2: 244. https://doi.org/10.3390/plants13020244
APA StyleShibaeva, T. G., Sherudilo, E. G., Ikkonen, E., Rubaeva, A. A., Levkin, I. A., & Titov, A. F. (2024). Effects of Extended Light/Dark Cycles on Solanaceae Plants. Plants, 13(2), 244. https://doi.org/10.3390/plants13020244