


Device Structure, Light Source Height, and Sunset Time Affect the Light-Trap Catching of Tea Leafhoppers
Abstract
1. Introduction
2. Results
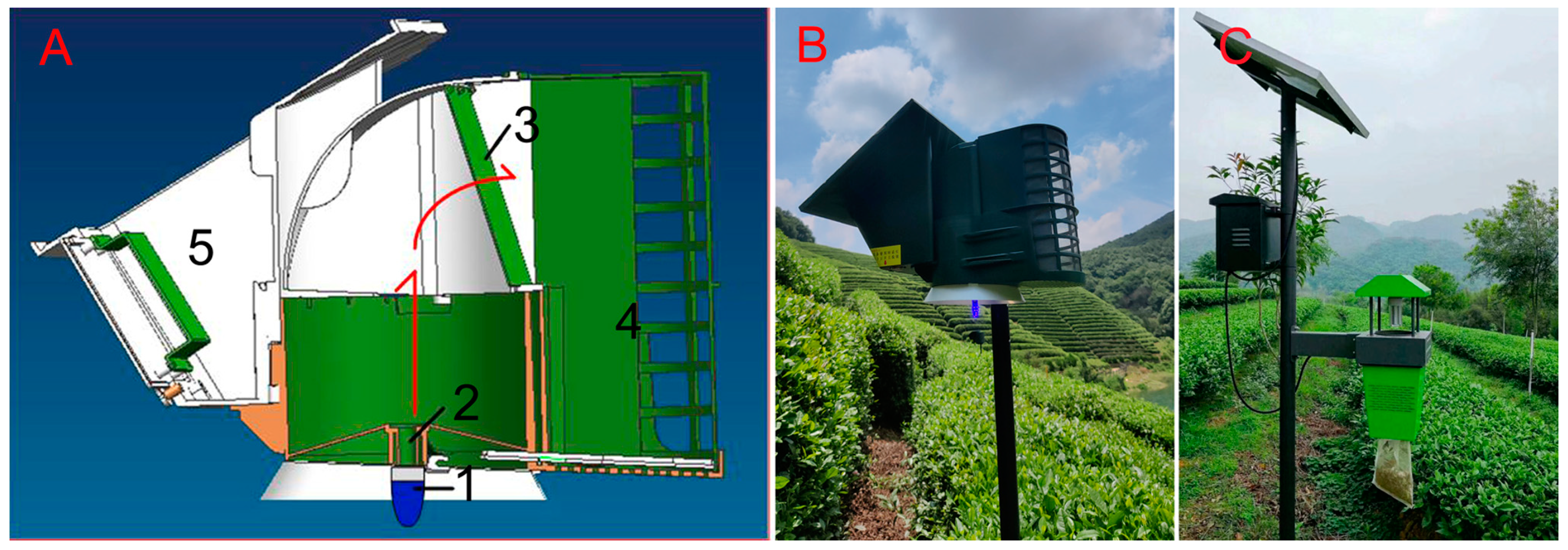
2.1. Improvements in Device Structure Increase Leafhopper Catches
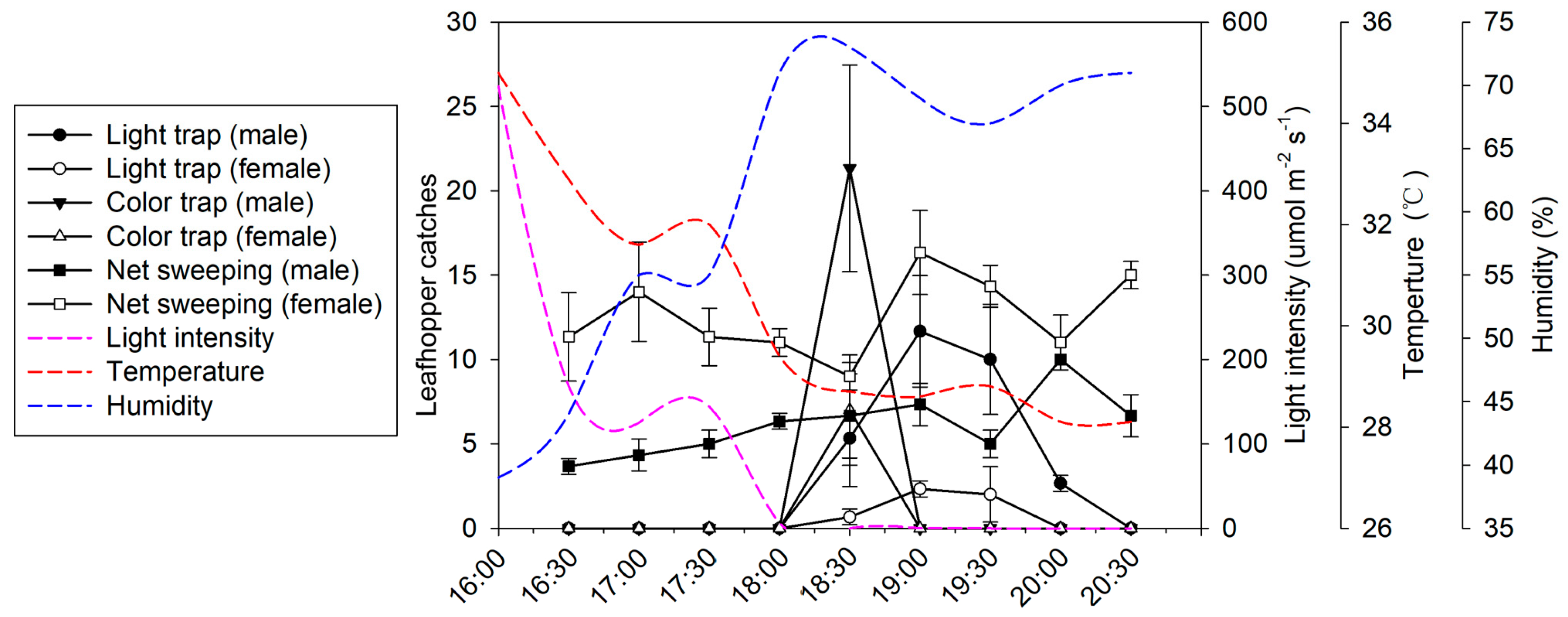
2.2. Trapping Window of Leafhoppers at Dusk
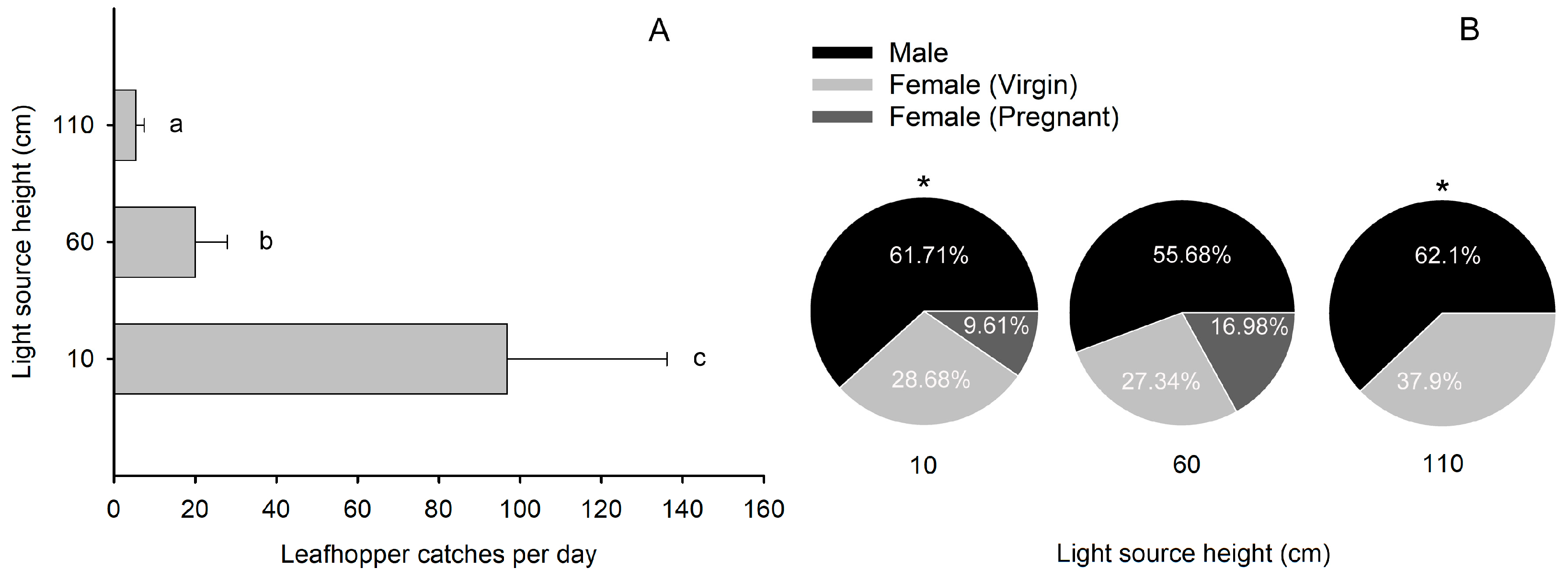
2.3. The Number of Leafhopper Catches Sharply Declined When the Height of the Light Source Exceeded Their Flight Height Range
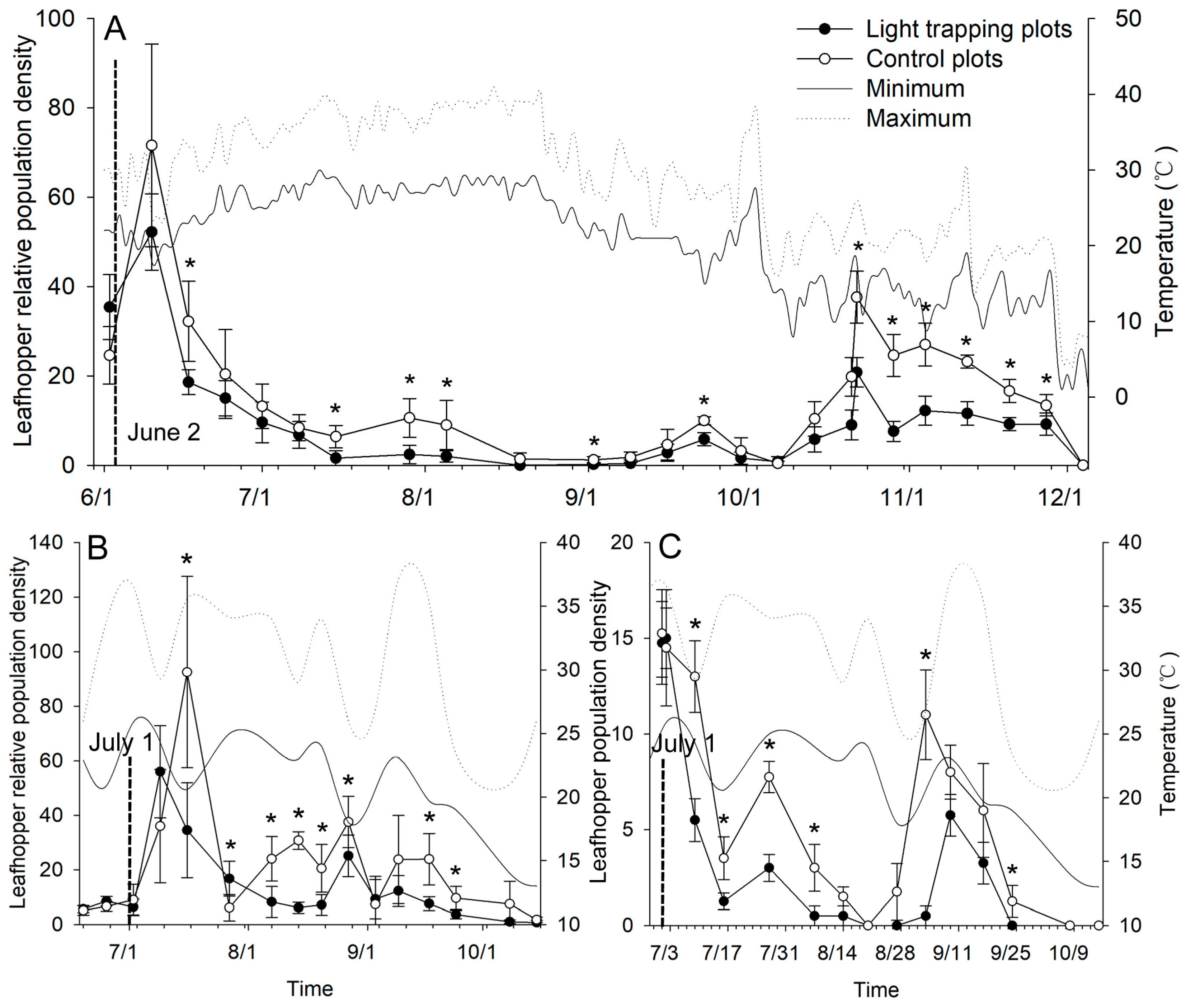
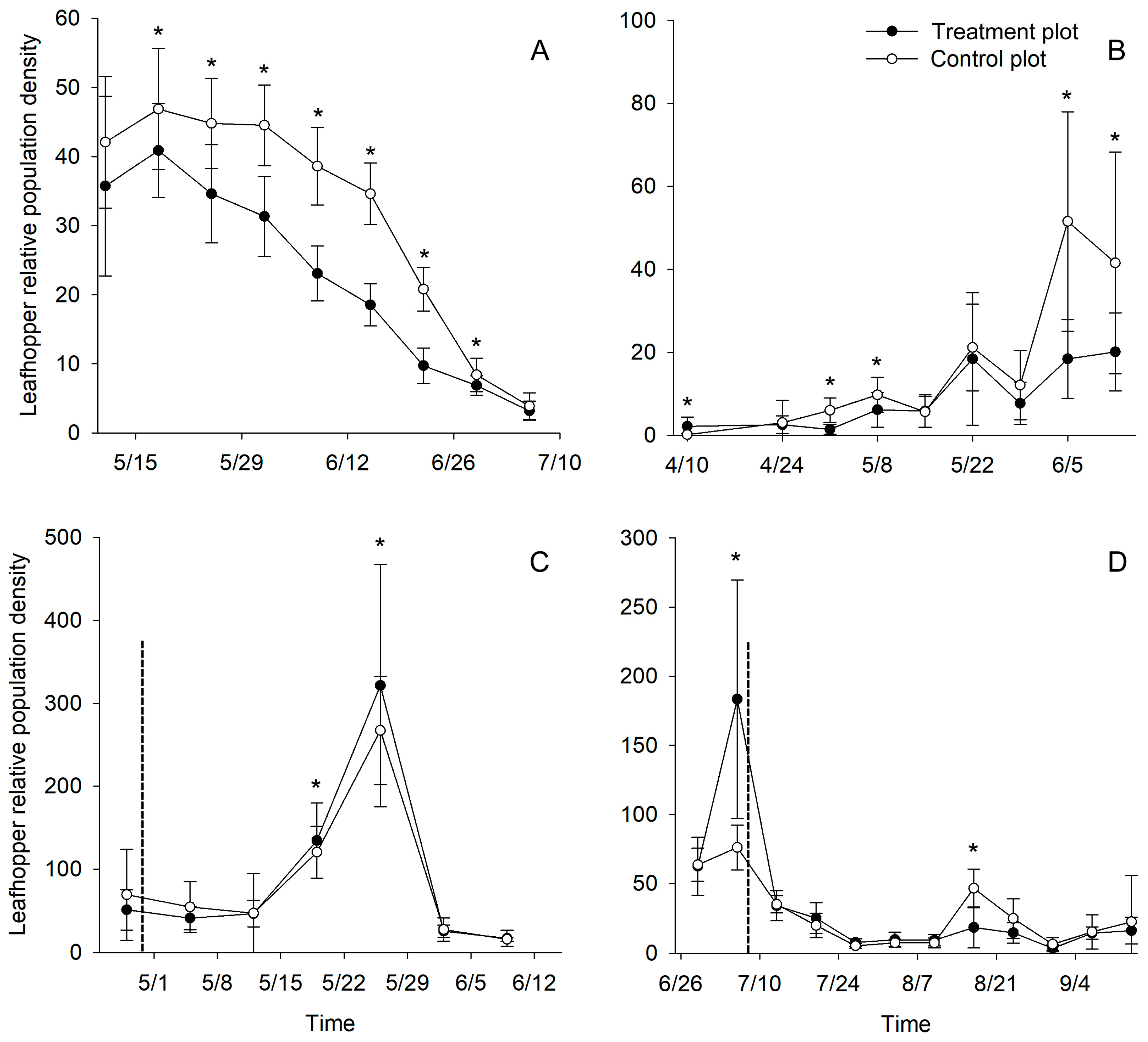
2.4. Efficacy of the Newly Designed Light Trap on E. onukii Control
3. Discussion
3.1. Improved Trapping Efficacy
3.2. Factors Affecting the Trapping Efficacy of the Light Trap
4. Materials and Methods
4.1. Light Traps
4.2. Device Structure
4.3. Trapping Window
4.4. Light Source Height
4.5. Efficacy of the Newly Designed Light Trap on E. onukii Control
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shimoda, M.; Honda, K. Insect reactions to light and its applications to pest management. Appl. Entomol. Zool. 1990, 48, 413–421. [Google Scholar] [CrossRef]
- Kim, K.N.; Huang, Q.Y.; Lei, C.L. Advances in insect phototaxis and application to pest management: A review. Pest Manag. Sci. 2019, 75, 3135–3143. [Google Scholar] [CrossRef]
- Gao, Y.; Li, G.H.; Li, K.B.; Lei, C.L.; Huang, Q.Y. Comparison of the trapping effect and antioxidant enzymatic activities using three different light sources in cockchafers. Environ. Sci. Pollut. Res. 2017, 24, 27855–27861. [Google Scholar] [CrossRef]
- Wilke, A.B.B.; Vasquez, C.; Carvajal, A.; Moreno, M.; Petrie, W.D.; Beier, J.C. Evaluation of the effectiveness of BG-Sentinel and CDC light traps in assessing the abundance, richness, and community composition of mosquitoes in rural and natural areas. Parasites Vectors 2022, 15, 51. [Google Scholar] [CrossRef]
- Bian, L.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Chen, Z.M. Decreased capture of natural enemies of pests in light traps with light-emitting diode technology. Ann. Appl. Biol. 2018, 173, 3. [Google Scholar] [CrossRef]
- Shi, L.Q.; Zeng, Z.H.; Huang, H.S.; Zhou, Y.M.; Vasseur, L.; You, M.S. Identification of Empoasca onukii (Hemiptera: Cicadellidae) and monitoring of its populations in the tea plantations of south China. J. Econ. Entomol. 2015, 108, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Jiang, R.; Zhang, C.; Gao, T.; Wang, Y.; Liu, C.; Long, Y.H.; Zhang, Y.L.; Yang, Y.Q. Wolbachia strain wGri from the tea geometrid moth Ectropis grisescens contributes to its host’s fecundity. Front. Microbiol. 2021, 12, 694466. [Google Scholar] [CrossRef]
- Wang, Z.B.; Ma, T.; Mao, T.F.; Guo, H.W.; Zhou, X.G.; Wen, X.J.; Xiao, Q. Application technology of the sex pheromone of the tea geometrid Ectropis grisescens (Lepidoptera: Geometridae). Int. J. Pest Manag. 2018, 64, 372–378. [Google Scholar] [CrossRef]
- Tan, R.R.; Chen, X.; Huang, D.J.; Wang, H.J.; Mao, Y.X. Pathogenicity of Ectropis grisescens Nucleopolyhedrovirus on Ectropis grisescens Warren and Ectropis obliqua Prout. Chin. J. Biol. Control 2023, 39, 684–689. (In Chinese) [Google Scholar]
- Shi, L.Q.; He, H.F.; Yang, G.; Huang, H.S.; Vasseur, L.; You, M.S. Are yellow sticky cards and light traps effective on tea green leafhoppers and their predators in Chinese tea plantations? Insects 2021, 12, 14. [Google Scholar] [CrossRef]
- Zhao, Q.; Shi, L.Q.; He, W.Y.; Li, J.Y.; You, S.J.; Chen, S.; Lin, J.; Wang, Y.B.; Zhang, L.W.; Yang, G.; et al. Genomic variations in the tea leafhopper reveal the basis of its adaptive evolution. Genom. Proteomics Bioinf. 2023, 20, 1092–1105. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.R. An improved suction trap for insects. Ann. Appl. Biol. 1951, 38, 582–591. [Google Scholar] [CrossRef]
- Bian, L.; Sun, X.L.; Chen, Z.M. Studies on daily flight activity and adult flight capacity of Empoasca vitis Gothe. J. Tea Sci. 2014, 34, 248–252. (In Chinese) [Google Scholar]
- Mazzoni, V.; Nieri, R.; Eriksson, A.; Virant-Doberlet, M.; Polajnar, J.; Anfora, G.; Lucchi, A. Mating disruption by vibrational signals: State of the field and perspectives. In Biotremology: Studying Vibrational Behavior; Animal Signals and Communication; Hill, P.S.M., Lakes-Harlan, R., Mazzoni, V., Narins, P., Virant-Doberlet, M., Wessel, A., Eds.; Springer: Cham, Swizerland, 2019; Volume 6, pp. 331–354. [Google Scholar]
- Wilton, D.P.; Fay, R.W. Air flow direction and velocity in light trap design. Entomol. Exp. Appl. 1972, 15, 377–386. [Google Scholar] [CrossRef]
- Shan, Y.; Zhou, X.S.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Xiu, C.L.; Chen, Z.M.; Bian, L. Mating and post-copulation behavior in the tea leafhopper, Empoasca onukii (Hemiptera: Cicadellidae). Front. Plant Sci. 2023, 14, 1273718. [Google Scholar] [CrossRef]
- Kersting, U.; Başpmar, H. Seasonal and diurnal flight activity of Circulifer haematoceps (Hom., Cicadellidae), an important leafhopper vector in the Mediterranean area and the Near East. J. Appl. Entomol. 1995, 119, 533–537. [Google Scholar] [CrossRef]
- Lanier, G.N. Principle of attraction-annihilation: Mass trapping and other means. In Behavior-Modifying Chemicals for Insect Management, Applications of Pheromones and other Attractants; Ridgway, R.L., Silverstein, R.M., Inscoe, M.N., Eds.; Marcel Dekker: New York, NY, USA, 1990; pp. 25–45. [Google Scholar]
- Polajnar, J.; Eriksson, A.; Lucchi, A.; Anfora, G.; Virant-Doberlet, M.; Mazzoni, V. Manipulating behaviour with substrate-borne vibrations—Potential for insect pest control. Pest Manag. Sci. 2014, 71, 15–23. [Google Scholar] [CrossRef]
- Zaffaroni-Caorsi, V.; Nieri, R.; Pugno, N.M.; Mazzoni, V. Effect of vibrational mating disruption on flight activity and oviposition to control the grapevine pest, Scaphoideus titanus. Arthropod. Struct. Dev. 2022, 69, 101173. [Google Scholar] [CrossRef]
- Mazzoni, V.; Lucchi, A.; Čokl, A.; Prešern, J.; Virant-Doberlet, M. Disruption of the reproductive behaviour of Scaphoideus titanus by playback of vibrational signals. Entomol. Exp. Appl. 2009, 133, 174–185. [Google Scholar] [CrossRef]
- Zapponi, L.; Nieri, R.; Zafaroni-Caorsi, V.; Pugno, N.M.; Mazzoni, V. Vibrational calling signals improve the efficacy of pheromone traps to capture the brown marmorated stink bug. J. Pest Sci. 2023, 96, 587–597. [Google Scholar] [CrossRef]
- Bian, L.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Xin, Z.J.; Chen, Z.M. Design of an attractant for Empoasca onukii (Hemiptera: Cicadellidae) based on the volatile components of fresh tea leaves. J. Econ. Entomol. 2018, 111, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Hegazi, E.; Khafagi, W.E.; Konstantopoulou, M.; Raptopoulos, D.; Tawfik, H.; El-Aziz, G.M.A.; El-Rahman, S.M.A.; Atwa, A.; Aggamy, E.; Showeil, S. Efficient mass-trapping method as an alternative tactic for suppressing populations of leopard moth (Lepidoptera: Cossidae). Ann. Entomol. Soc. Am. 2009, 102, 809–818. [Google Scholar] [CrossRef]
- Byers, J.A. Analysis of vertical distributions and effective flight layers of insects: Three-dimensional simulation of flying insects and catch at trap heights. Environ. Entomol. 2011, 40, 1210–1222. [Google Scholar] [CrossRef]
- Gaglio, G.; Brianti, E.; Napoli, E.; Falsone, L.; Dantas-Torres, F.; Tarallo, V.D.; Otranto, D.; Giannetto, S. Effect of night time-intervals, height of traps and lunar phases on sand fly collection in a highly endemic area for canine leishmaniasis. Acta Trop. 2014, 133, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Venter, G.J.; Hermanides, K.G.; Boikanyo, S.N.B.; Majatladi, D.M.; Morey, L. The effect of light trap height on the numbers of Culicoides midges collected under field conditions in South Africa. Vet. Parasitol. 2009, 166, 343–345. [Google Scholar] [CrossRef]
- Silva, F.S.; Lobo, S.E.P.D.; da Silva, A.A. Seasonal abundance and the effect of light trap height on the captures of the vesicating beetles of the Genus Paederus (Coleoptera: Staphylinidae) in Brazil. Entomol. News 2017, 127, 21–27. [Google Scholar] [CrossRef]
- de Araújo, E.C.; Neta, B.M.C.; Brito, J.M.; Silva, F.S. Effect of ultraviolet LED and trap height on catches of host-seeking anopheline mosquitoes by using a low-cost passive light trap in northeast Brazil. Parasitol. Res. 2023, 122, 1343–1349. [Google Scholar] [CrossRef]
- Braverman, Y.; Linley, J.R. Effect of light trap height on catch of Culicoides (Diptera: Ceratopogonidae) in Israel. J. Med. Entomol. 1993, 30, 1060–1063. [Google Scholar] [CrossRef]
- Bian, L.; Cai, X.M.; Chen, Z.M. Effects of light-emitting diode light traps with a suction fan on the population dynamics of the tea leafhopper Empoasca onukii (Hemiptera:Cicadellidae) within the effective distance. J. Plant Protect. 2019, 46, 902–909. (In Chinese) [Google Scholar]
- Wang, Q.S.; Li, H.L.; Zhang, H.; Wang, D.F.; Liu, F.J.; Li, L.D. Seasonal effect of hanging height for yellow sticky traps on trapping adult Empoasca vitis (göthe) at tea plantations. Acta Tea Sin. 2015, 56, 121–125. (In Chinese) [Google Scholar]
- Jarrell, K.R.; Rebek, E.J.; Wayadande, A.C.; Giles, K.L. Biology, ecology, and management of eastern grape leafhopper (Hemiptera: Cicadellidae), a key pest of vineyards in north America. J. Integr. Pest Manag. 2020, 11, 6. [Google Scholar] [CrossRef]
- Matsukura, K.; Yoshida, K.; Matsumura, M. Efficient monitoring of maize orange leafhopper, Cicadulina bipunctata (Hemiptera: Cicadellidae), and small brown planthopper, Laodelphax striatellus (Hemiptera: Delphacidae), in forage maize fields using yellow sticky traps. Appl. Entomol. Zool. 2011, 46, 585–591. [Google Scholar] [CrossRef]
- Sabale, J.P.; Das, C.; Samui, R.P. Influence of weather factors on light trap catches of green leaf hopper at Pattambi, Kerala. J. Agrometeorol. 2010, 12, 108–110. [Google Scholar] [CrossRef]
- Skuhravy, V.; Zumr, V. Nocturnal and seasonal flight activity of the nun moth Lymantria monacha L. (Lepidoptera, Lymantriidae) as determined by pheromone and light traps. J. Appl. Entomol. 1981, 92, 315–319. [Google Scholar] [CrossRef]
- Rodriguez, C.M.; Madden, L.V.; Nault, L.R. Diel flight periodicity of Graminella nigrifrons (Homoptera: Cicadellidae). Ann. Entomol. Soc. Am. 1992, 85, 792–798. [Google Scholar] [CrossRef]
- Raju, N.; Rajendran, R.; Kareem, A.A. Ecological factors favouring green leafhopper (Nephotettix virescens) population in rice (Oryza sativa). Indian J. Agric. Sci. 1997, 67, 36–38. [Google Scholar]
- Feil, H.; Feil, W.S.; Purcell, A.H. Effects of temperature on the flight activity of Graphocephala atropunctata (Hemiptera: Cicadellidae). J. Econ. Entomol. 2000, 93, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Bian, L.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Chen, Z.M. Sticky card for Empoasca onukii with bicolor patterns captures less beneficial arthropods in tea gardens. Crop Prot. 2021, 149, 105761. [Google Scholar] [CrossRef]
- Pu, X.Y.; Feng, M.G.; Shi, C.H. Impact of three application methods on the field efficacy of a Beauveria bassiana-based mycoinsecticide against the false-eye leafhopper, Empoasca vitis (Homoptera: Cicadellidae) in the tea canopy. Crop Prot. 2005, 24, 167–175. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summer | Autumn | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Trap 1 | Trap 2 | Trap 3 | F | p | Trap 1 | Trap 2 | Trap 3 | F | p | ||
| Tea pest | |||||||||||
| Hemiptera | Cicadellidae | 1152.33 ± 127.49 a | 23.67 ± 6.43 c | 308.67 ± 53.15 b | 162.14 | <0.001 | 39.33 ± 8.74 a | 7.0 ± 2.0 c | 21.0 ± 5.57 b | 21.25 | 0.002 |
| Lepidoptera | Geometridae | 32.0 ± 6.08 b | 28.67 ± 3.06 b | 65.0 ± 8.89 a | 28.97 | 0.001 | 13.67 ± 4.51 a | 13.0 ± 7.21 a | 24.67 ± 8.74 a | 2.60 | 0.154 |
| Olethreutidae | 44.33 ± 5.03 b | 8.0 ± 3.61 b | 19.67 ± 8.14 a | 29.59 | 0.001 | 0 | 0 | 0 | |||
| Others | 12.67 ± 3.51 a | 10.67 ± 6.23 a | 32.67 ± 14.57 b | 5.01 | 0.053 | 8.33 ± 2.52 b | 6.67 ± 4.62 b | 34.33 ± 14.05 a | 9.63 | 0.013 | |
| Total | 89.0 ± 10.39 a | 47.33 ± 4.93 b | 117.33 ± 27.47 a | 12.59 | 0.007 | 22.0 ± 6.25 b | 19.67 ± 11.72 b | 59.0 ± 20.66 a | 7.26 | 0.025 | |
| Diptera | Cecidomyiidae | 249.33 ± 61.09 a | 218.33 ± 72.22 a | 277.67 ± 77.41 a | 0.53 | 0.614 | 4084.0 ± 651.75 a | 2916.67 ± 940.69 a | 4560.0 ± 799.02 a | 3.30 | 0.108 |
| Simuliidae | 450.0 ± 53.51 a | 23.67 ± 16.62 c | 200.0 ± 74.18 b | 47.78 | <0.001 | 0 | 0 | 0 | |||
| Others | 28.0 ± 14.18 a | 32.0 ± 15.52 a | 58.67 ± 33.83 a | 1.58 | 0.282 | 57.0 ± 19.29 b | 41.67 ± 22.5 b | 123.0 ± 39.13 a | 6.98 | 0.027 | |
| Total | 727.33 ± 57.84 a | 274.0 ± 55.46 b | 536.33 ± 172.4 a | 12.9 | 0.007 | 4141.0 ± 670.84 ab | 2958.33 ± 962.53 b | 4683.0 ± 837.19 a | 3.37 | 0.104 | |
| Natural enemy | |||||||||||
| Coleoptera | Staphylinidae | 1.0 ± 1.0 b | 0.67 ± 0.58 b | 6.67 ± 2.08 a | 18.06 | 0.003 | 0.67 ± 0.58 a | 0.67 ± 0.58 a | 3.33 ± 2.31 a | 3.56 | 0.096 |
| Hymenoptera | Formicidae | 2.33 ± 0.58 b | 0 b | 11.0 ± 2.31 a | 18.51 | 0.003 | 0.67 ± 0.58 b | 0.33 ± 0.58 b | 8.67 ± 2.08 a | 40.07 | <0.001 |
| Others | 0 b | 0 b | 1.33 ± 0.58 a | 16.0 | 0.004 | 0.67 ± 1.15 a | 0.33 ± 0.58 a | 1.67 ± 0.58 a | 2.17 | 0.196 | |
| Total | 3.33 ± 0.58 b | 0.67 ± 0.58 b | 19.0 ± 2.65 a | 115.17 | <0.001 | 2.0 ± 1.0 b | 1.33 ± 1.53 b | 13.67 ± 6.52 a | 16.87 | 0.003 | |
| Date | 28 July | 8 August | 9 August | 31 August | 14 September | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ts | 18:56 | 18:48 | 18:47 | 18:36 | 18:07 | |||||
| Light trap | t1 | t2 | t1 | t2 | t1 | t2 | t1 | t2 | t1 | t2 |
| 18:30 | 20:00 | 18:00 | 20:00 | 18:00 | 20:00 | 18:00 | 20:00 | 17:30 | 20:00 | |
| Male | 12.89 ± 6.29 | 1.75 ± 1.22 | 5.17 ± 3.86 | 7.41 ± 4.72 | 12.07 ± 10.15 | |||||
| Female | 5.56 ± 4.90 | 0.17 ± 0.39 | 1.33 ± 2.02 | 1.25 ± 1.36 | 2.73 ± 3.47 | |||||
| t | 11.36 | 5.06 | 5.90 | 4.68 | 4.91 | |||||
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |||||
| Sticky card | t1 | t2 | t1 | t2 | t1 | t2 | t1 | t2 | t1 | t2 |
| 18:30 | 19:30 | 18:30 | 19:00 | 18:30 | 19:00 | 18:00 | 18:30 | 17:30 | 18:30 | |
| Male | 6.66 ± 5.85 | 2.33 ± 0.58 | 11.67 ± 3.51 | 21.33 ± 7.51 | 21.5 ± 20.15 | |||||
| Female | 5 ± 6.03 | 0 | 0 | 7.0 ± 4.0 | 11.0 ± 7.75 | |||||
| t | 7.91 | 7 | 5.75 | 7.07 | 1.29 | |||||
| p | 0.001 | 0.02 | 0.029 | 0.019 | 0.253 | |||||
| Net sweeping | ||||||||||
| Male | 4.25 ± 2.74 | 3.89 ± 1.37 | 4.15 ± 2.25 | 6.11 ± 2.17 | 6.19 ± 2.68 | |||||
| Female | 12.37 ± 4.46 | 9.48 ± 2.87 | 10.78 ± 3.04 | 12.59 ± 2.98 | 22.74 ± 4.61 | |||||
| t | 10.45 | 8.46 | 14.42 | 8.92 | 19.41 | |||||
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, L.; Ji, H.; Cai, X.; Cheng, G.; Xie, X.; Duan, X.; Chen, Z. Device Structure, Light Source Height, and Sunset Time Affect the Light-Trap Catching of Tea Leafhoppers. Plants 2024, 13, 241. https://doi.org/10.3390/plants13020241
Bian L, Ji H, Cai X, Cheng G, Xie X, Duan X, Chen Z. Device Structure, Light Source Height, and Sunset Time Affect the Light-Trap Catching of Tea Leafhoppers. Plants. 2024; 13(2):241. https://doi.org/10.3390/plants13020241
Chicago/Turabian StyleBian, Lei, Huihua Ji, Xiaoming Cai, Guo Cheng, Xiaoqun Xie, Xiaofeng Duan, and Zongmao Chen. 2024. "Device Structure, Light Source Height, and Sunset Time Affect the Light-Trap Catching of Tea Leafhoppers" Plants 13, no. 2: 241. https://doi.org/10.3390/plants13020241
APA StyleBian, L., Ji, H., Cai, X., Cheng, G., Xie, X., Duan, X., & Chen, Z. (2024). Device Structure, Light Source Height, and Sunset Time Affect the Light-Trap Catching of Tea Leafhoppers. Plants, 13(2), 241. https://doi.org/10.3390/plants13020241