
Effect of Bacteria Inoculation on Colonization of Roots by Tuber melanosporum and Growth of Quercus ilex Seedlings

 ,
, 
 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
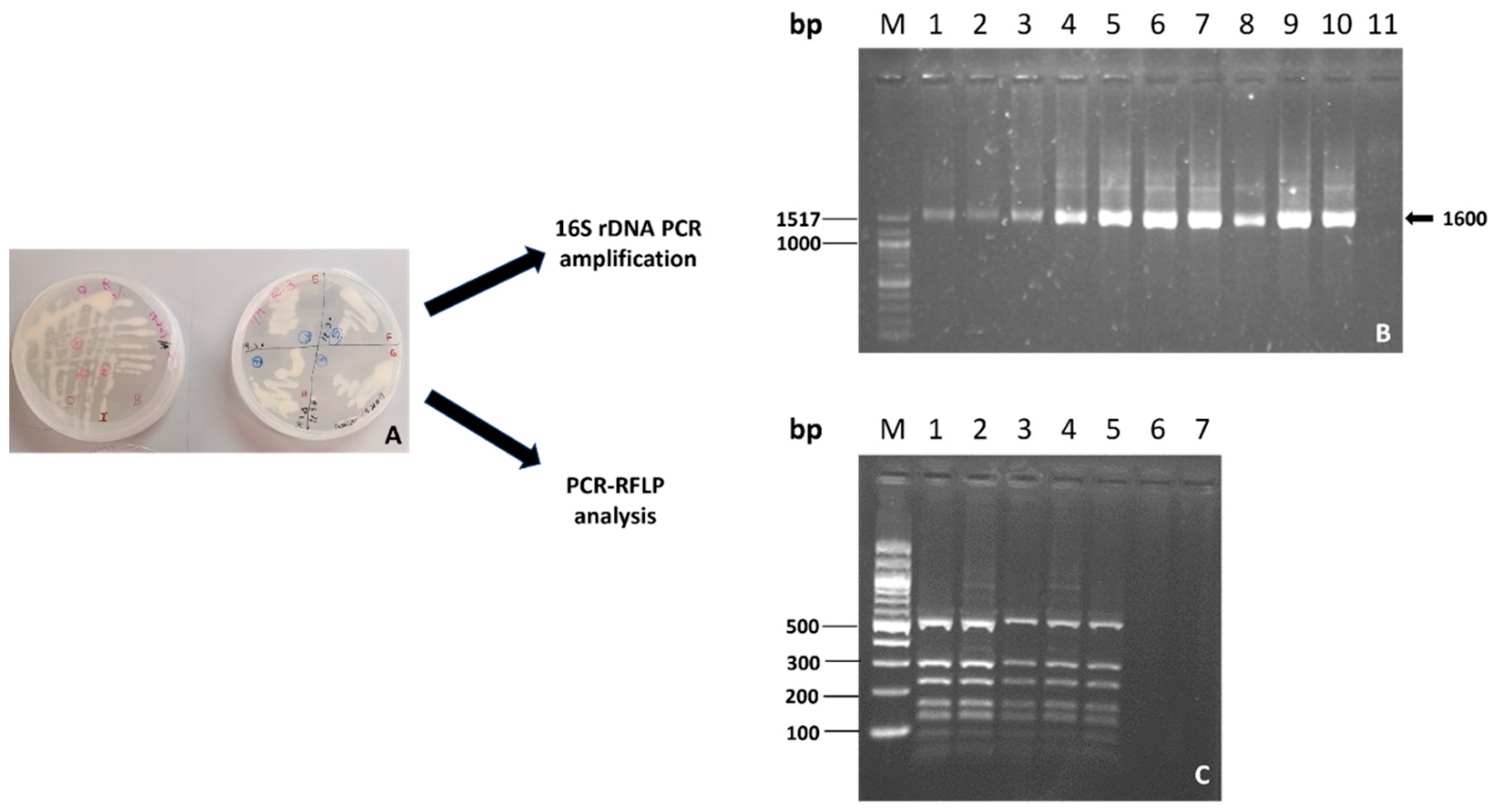
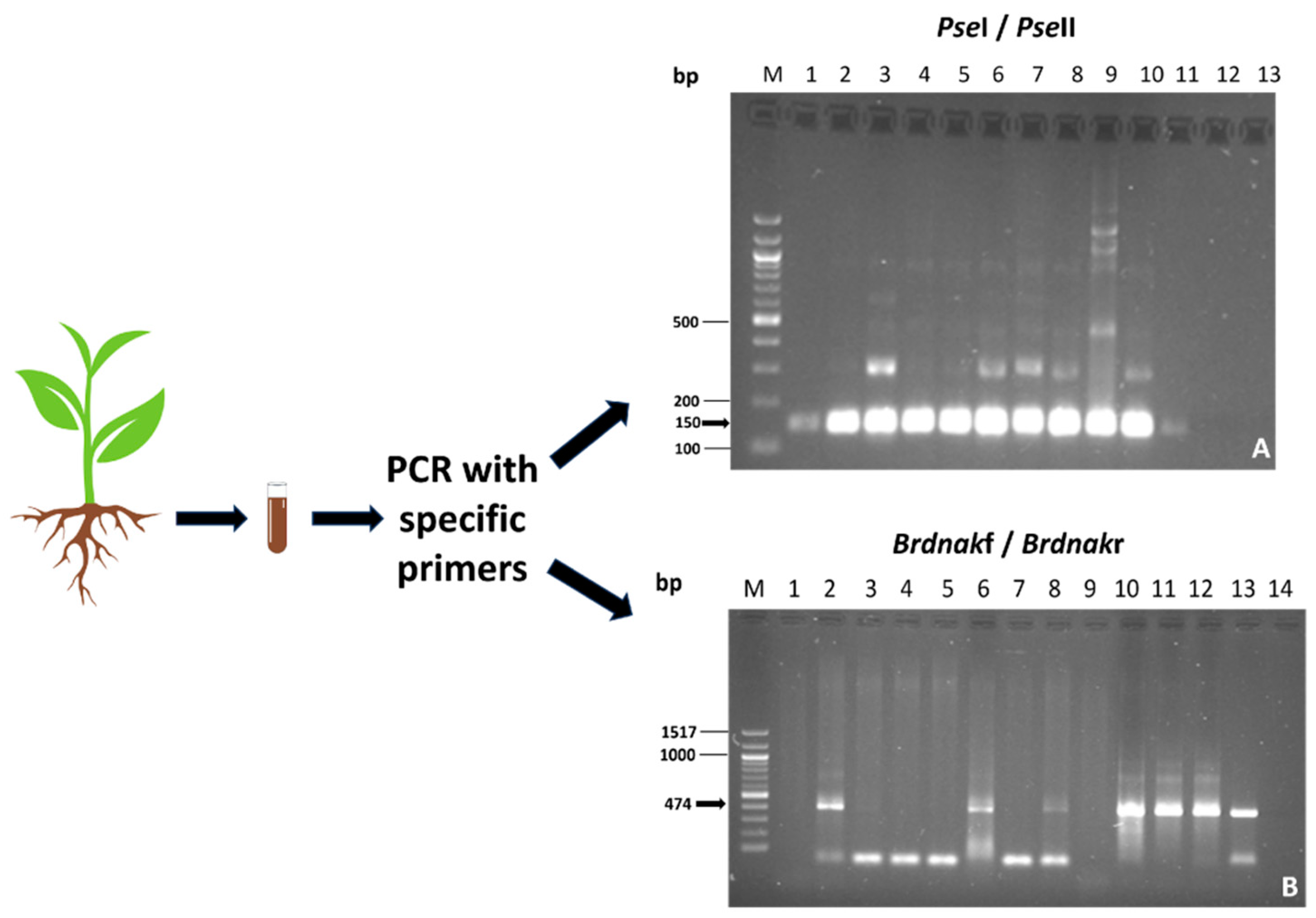
2.1. Evaluation of the Identity and the Presence of Inoculated Bacterial Strains
2.2. Mycorrhizae Analysis
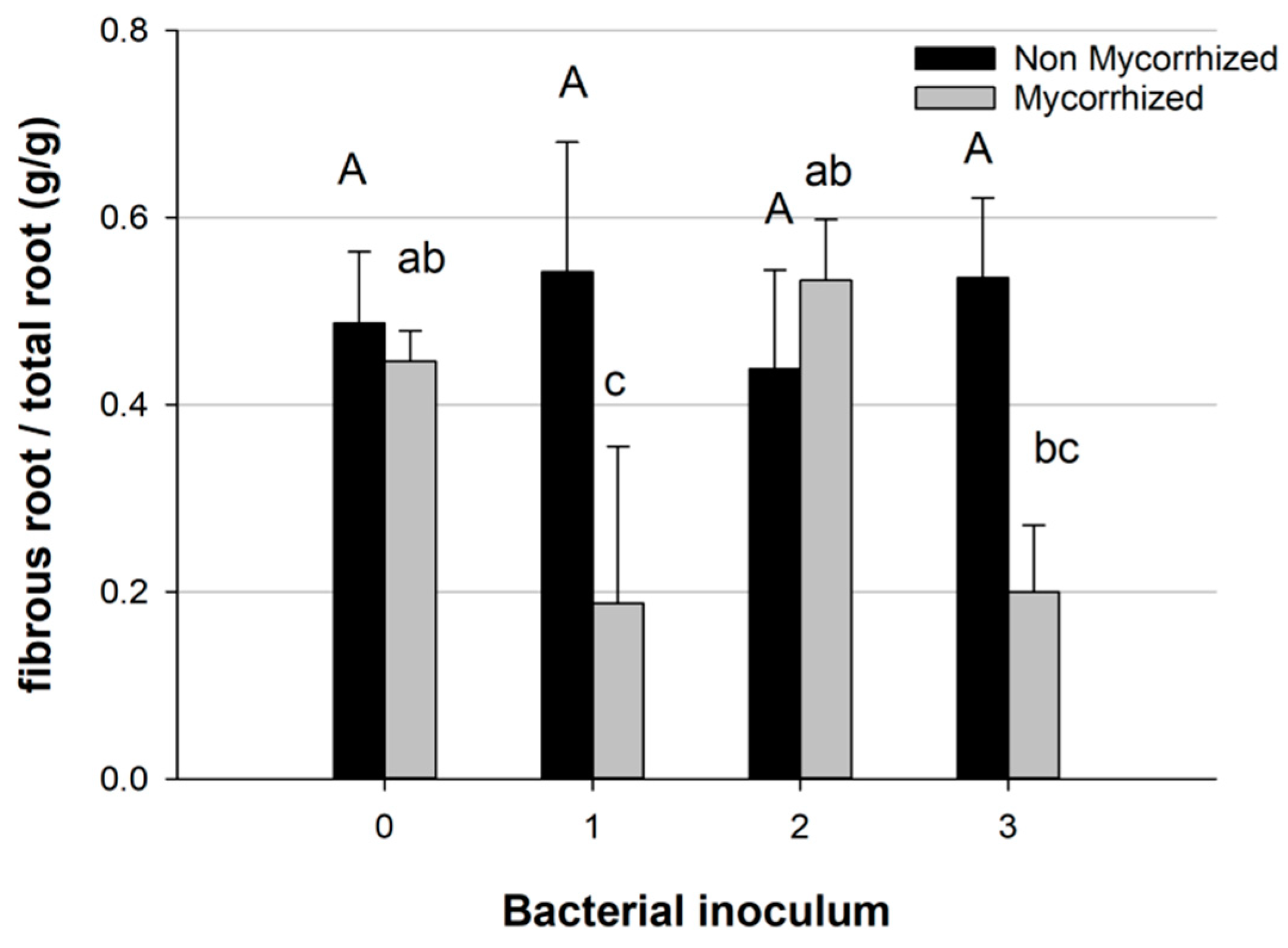
2.3. Plant and Root Architecture Analysis
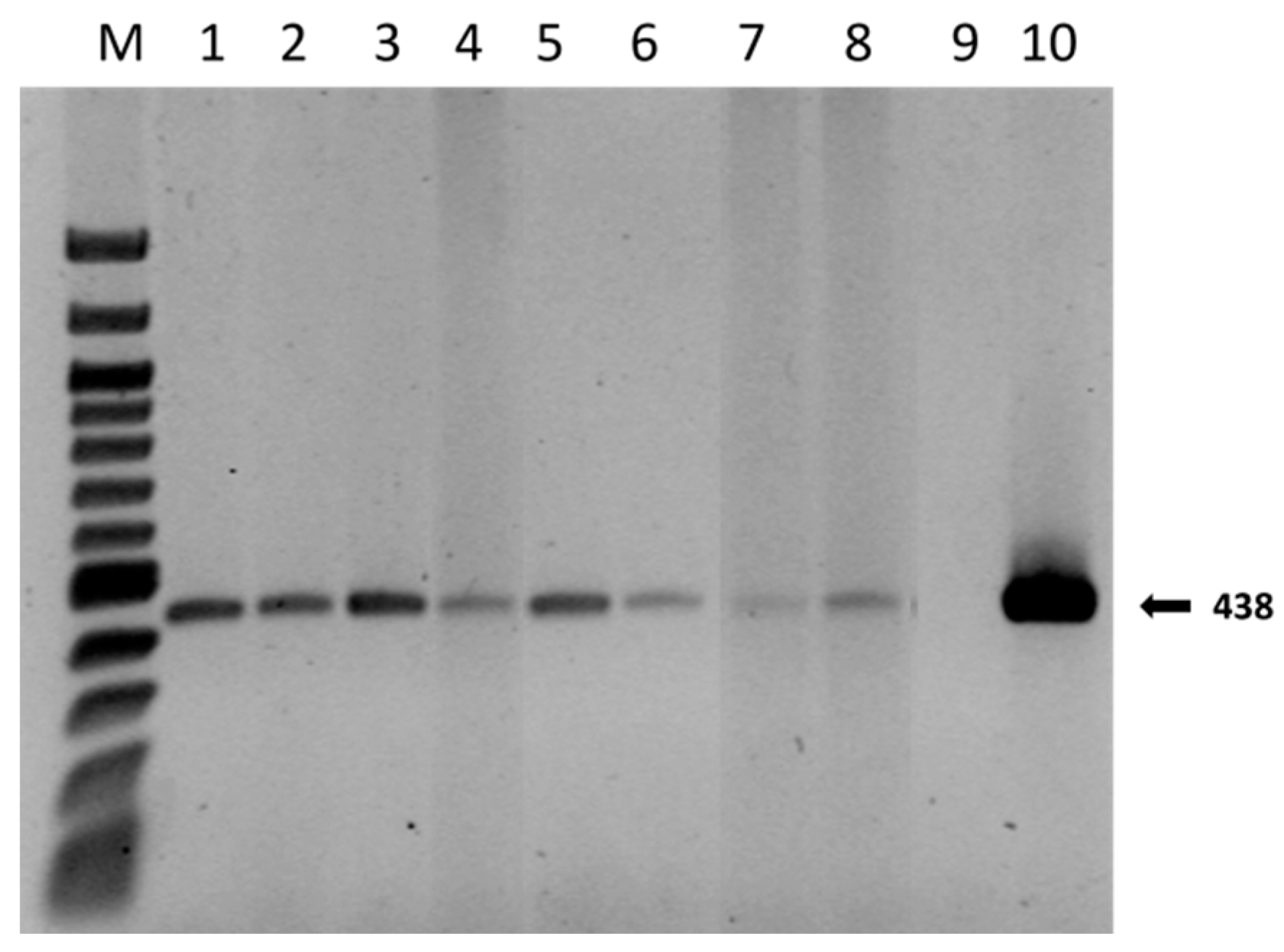
2.4. Mycorrhization Degree
3. Discussion
4. Materials and Methods
4.1. Seedling Cultivation
4.2. T. melanosporum Inoculation
4.3. Bacterial Inocula Preparation
4.4. Experimental Setup
4.5. Molecular Identification of Bacterial Species
4.6. Morphological and Molecular Identification of T. melanosporum Root Colonization
4.7. Effects of Bacterial Formulations on Quercus Ilex Development and Plant Root T. melanosporum Colonization
4.7.1. Plant Measurements
4.7.2. Estimation of T. melanosporum Colonization
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviation
| ECM | ectomycorrhizae |
References
- Pegler, D.N.; Spooner, B.M.; Young, T.W.K. British Truffles. A revision of British hypogeous fungi. Kew. Bull. 1993, 49, 167. [Google Scholar] [CrossRef]
- Bonito, G.M.; Smith, M.E. General systematic position of the truffles: Evolutionary theories. In True Truffle (Tuber spp.) in the World. Soil Biology; Springer: Cham, Switzerland, 2016; Volume 47, pp. 3–18. ISBN 978-3-319-31436-5. [Google Scholar] [CrossRef]
- Laliberté, E.; Lambers, H.; Burgess, T.I.; Wright, S.J. Phosphorus limitation, soil-borne pathogens and the coexistence of plant species in hyperdiverse forests and shrublands. New Phytol. 2015, 206, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Callot, G.; Daignieres y Fernandez, D.; Raymond, M.; Salducci, X. Incidences du sol sur la production de truffe noire du Perigord Tuber melanosporum. In Proceedings of the V Congress International: Science and Cultivation of Truffe; Corvoisier, M., Olivier, J., Chevalier, G., Eds.; Federation Francaise des Trufficulteurs: Aix en Provence, France, 1999. [Google Scholar]
- Zambonelli, A.; Iotti, M.; Hall, I. Current status of truffle cultivation: Recent results and future perspectives. Micol. Ital. 2015, 44, 31–40. [Google Scholar] [CrossRef]
- Leonardi, M.; Iotti, M.; Pacioni, G.; Hall, I.R.; Zambonelli, A. Truffles: Biodiversity, ecological significances, and biotechnological applications. In Industrially Important Fungi for Sustainable Development; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2021; pp. 107–146. [Google Scholar] [CrossRef]
- Ortego, J.; Bonal, R.; Munoz, A. Genetic Consequences of Habitat Fragmentation in Long-Lived Tree Species: The Case of the Mediterranean Holm Oak (Quercus Ilex, L.). J. Hered. 2010, 101, 717–726. [Google Scholar] [CrossRef] [PubMed]
- MIPAAF (Italian Ministry for Agriculture, Food and Forest Policies). DM 17/05/2022. Guidelines for Planning the Production and Use of Native Species of Forestry Interest. Available online: https://www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/18359 (accessed on 21 December 2023).
- Grossnickle, S.C.; Macdonald, J.E. Seedling quality: History, application, and plant attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef]
- Murat, C. Forty years of inoculating seedlings with truffle fungi: Past and future perspectives. Mycorrhiza 2015, 25, 77–81. [Google Scholar] [CrossRef]
- Aspray, T.J.; Frey-Klett, P.; Jones, J.E.; Whipps, J.M.; Garbaye, J.; Bending, G.D. Mycorrhization helper bacteria: A case of specificity for altering ectomycorrhiza architecture but not ectomycorrhiza formation. Mycorrhiza 2006, 16, 533–541. [Google Scholar] [CrossRef]
- Reyna, S.; Garcia-Barreda, S. Black truffle cultivation: A global reality. For. Syst. 2014, 23, 317–328. [Google Scholar] [CrossRef]
- Taschen, E.; Sauve, M.; Taudiere, A.; Parlade, J.; Selosse, M.A.; Richard, F. Whose truffle is this? distribution patterns of ectomycorrhizal fungal diversity in Tuber melanosporum Brûlés developed in multi-host mediterranean plant communities. Environ. Microbiol. 2015, 17, 2747–2761. [Google Scholar] [CrossRef]
- Balestrini, R.; Kottke, I. Structure and development of ectomycorrhizal roots. In Mol. Mycorrhizal Symbiosis; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 47–61. [Google Scholar] [CrossRef]
- Zadworny, M.; Eissenstat, D.M. Contrasting the morphology, anatomy and fungal colonization of new pioneer and fibrous roots. New Phytol. 2011, 190, 213–221. [Google Scholar] [CrossRef]
- Chen, W.; Ye, T.; Sun, Q.; Niu, T.; Zhang, J. Arbuscular mycorrhizal fungus alters root system architecture in Camellia sinensis L. as revealed by RNA-Seq analysis. Front. Plant Sci. 2021, 12, 777357. [Google Scholar] [CrossRef] [PubMed]
- Hetrick, B.A.D. Mycorrhizas and root architecture. Experientia 1991, 47, 355–362. [Google Scholar] [CrossRef]
- Pruett, G.E.; Bruhn, J.N.; Mihail, J.D. Greenhouse Production of Burgundy truffle mycorrhizae on oak roots. New For. 2009, 37, 43–52. [Google Scholar] [CrossRef]
- Bruhn, J.N.; Mihail, J.D.; Pruett, G.E. Truffle seedling production method has long-term consequences for tree growth and root colonization. Agrofor. Syst. 2013, 87, 679–688. [Google Scholar] [CrossRef]
- Fischer, C.; Colinas, C. Methodology for certification of Quercus ilex seedlings inoculated with Tuber melanosporum for commercial application. In Proceedings of the First International Conference on Mycorrhiza, Berkeley, CA, USA, 4–9 August 1996. [Google Scholar]
- Alvarado, P.; Manjon, J.L. A quantitative and molecular examination of Tuber melanosporum mycorrhizae in Quercus ilex seedlings from different suppliers in Spain. For. Syst. 2013, 22, 159–169. [Google Scholar] [CrossRef][Green Version]
- Andrés-Alpuente, A.; Sánchez, S.; Martín, M.; Aguirre, Á.J.; Barriuso, J.J. Comparative analysis of different methods for evaluating quality of Quercus ilex seedlings inoculated with Tuber melanosporum. Mycorrhiza 2014, 24 (Suppl. S1), 29–37. [Google Scholar] [CrossRef]
- Garcia-Montero, L.G.; Moreno, D.; Monleon, V.J.; Arredondo-Ruiz, F. Natural production of Tuber aestivum in central Spain: Pinus spp. versus Quercus spp. Brûlés. For. Syst. 2014, 23, 394–399. [Google Scholar] [CrossRef]
- Bonet, J.A.; Fischer, C.R.; Colinas, C. Cultivation of black truffle to promote reforestation and land-use stability. Agron. Sustain. Dev. 2006, 26, 69–76. [Google Scholar] [CrossRef]
- Büntgen, U.; Egli, S.; Schneider, L.; von Arx, G.; Rigling, A.; Camarero, J.J.; Sangüesa-Barreda, G.; Fischer, C.R.; Oliach, D.; Bonet, J.A.; et al. Long-term irrigation effects on Spanish holm oak growth and its black truffle symbiont. Agric. Ecosyst. Environ. 2015, 202, 148–159. [Google Scholar] [CrossRef]
- Taschen, E.; Callot, G.; Savary, P.; Sauve, M.; Penuelas-samaniego, Y.; Rousset, F.; Parlade, X.; Selosse, M.A.; Richard, F. Efficiency of the traditional practice of traps to stimulate black truffle production, and its ecological mechanisms. Sci. Rep. 2022, 12, 16201. [Google Scholar] [CrossRef]
- Bonfante, P.; Anca, I.A. Plants, mycorrhizal fungi, and bacteria: A network of interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Splivallo, R.; Bossi, S.; Maffei, M.; Bonfante, P. Discrimination of truffle fruiting body versus mycelial aromas by stir bar sorptive extraction. Phytochemistry 2007, 68, 2584–2598. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Gamalero, E.; Glick, B.R. Mycorrhizal-bacterial amelioration of plant abiotic and biotic stress. Front. Sustain. Food Syst. 2021, 5, 672881. [Google Scholar] [CrossRef]
- Dominguez, J.A.; Martin, A.; Anriquez, A.; Albanesi, A. The combined effects of Pseudomonas fluorescens and Tuber melanosporum on the quality of Pinus halepensis seedlings. Mycorrhiza 2012, 22, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Piñuela, Y.; Alday, J.G.; Oliach, D.; Bolaño, F.; Colinas, C.; Bonet, J.A. Use of inoculator bacteria to promote Tuber melanosporum root colonization and growth on Quercus faginea saplings. Forests 2020, 11, 792. [Google Scholar] [CrossRef]
- Barbieri, E.; Guidi, C.; Bertaux, J.; Frey-Klett, P.; Garbaye, J.; Ceccaroli, P.; Saltarelli, R.; Zambonelli, A.; Stocchi, V. Occurrence and diversity of bacterial communities in Tuber magnatum during truffle maturation. Environ. Microbiol. 2007, 9, 2234–2246. [Google Scholar] [CrossRef] [PubMed]
- Antony-Babu, S.; Deveau, A.; Van Nostrand, J.D.; Zhou, J.; Le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Black truffle associated bacterial communities during the development and maturation of Tuber melanosporum ascocarps and putative functional roles. Environ. Microbiol. 2014, 16, 2831–2847. [Google Scholar] [CrossRef]
- Vahdatzadeh, M.; Deveau, A.; Splivallo, R. The Role of the microbiome of truffles in aroma formation: A meta-analysis approach. Appl. Environ. Microbiol. 2015, 81, 6946–6952. [Google Scholar] [CrossRef]
- Le Roux, C.; Tournier, E.; Lies, A.; Sanguin, H.; Chevalier, G.; Duponnois, R.; Mousain, D.; Prin, Y. Bacteria of the genus Rhodopseudomonas (Bradyrhizobiaceae): Obligate symbionts in mycelial cultures of the black truffles Tuber melanosporum and Tuber brumale. Springerplus 2016, 5, 1085. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Deveau, A.; Palin, B.; Delaruelle, C.; Peter, M.; Kohler, A.; Pierrat, J.C.; Sarniguet, A.; Garbaye, J.; Martin, F.; Frey-Klett, P. The mycorrhiza helper Pseudomonas fluorescens BBc6R8 has a specific priming effect on the growth, morphology and gene expression of the ectomycorrhizal fungus Laccaria bicolor S238N. New Phytol. 2007, 175, 743–755. [Google Scholar] [CrossRef]
- Rincón, A.; de Felipe, M.R.; Fernández-Pascual, M. Inoculation of Pinus halepensis Mill. with selected ectomycorrhizal fungi improves seedling establishment 2 years after planting in a degraded gypsum soil. Mycorrhiza 2007, 18, 23–32. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, X.; Smith, D.L. Enhanced soybean plant growth resulting from coinoculation of Bacillus strains with Bradyrhizobium japonicum. Crop Sci. 2003, 43, 1774–1781. [Google Scholar] [CrossRef]
- Leggett, M.; Diaz-Zorita, M.; Koivunen, M.; Bowman, R.; Pesek, R.; Stevenson, C.; Leister, T. Soybean response to inoculation with Bradyrhizobium japonicum in the United States and Argentina. Agron. J. 2017, 109, 1031–1038. [Google Scholar] [CrossRef]
- Barbieri, E.; Ceccaroli, P.; Saltarelli, R.; Guidi, C.; Potenza, L.; Basaglia, M.; Fontana, F.; Baldan, E.; Casella, S.; Ryahi, O.; et al. New evidence for nitrogen fixation within the Italian white truffle Tuber magnatum. Fungal Biol. 2010, 114, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Pinkas, Y.; Maimon, M.; Shabi, E.; Elisha, S.; Shmulewich, Y.; Freeman, S. Inoculation, isolation and identification of Tuber melanosporum from old and new oak hosts in Israel. Mycol. Res. 2000, 104, 472–477. [Google Scholar] [CrossRef]
- Herrero de Aza, C.; Armenteros, S.; McDermott, J.; Mauceri, S.; Olaizola, J.; Hernández-Rodríguez, M.; Mediavilla, O. Fungal and bacterial communities in Tuber melanosporum plantations from Northern Spain. Forests 2022, 13, 385. [Google Scholar] [CrossRef]
- Kang, S.M.; Asaf, S.; Kim, S.J.; Yun, B.W.; Lee, I.J. Complete genome sequence of plant growth-promoting bacterium Leifsonia xyli SE134, a possible gibberellin and auxin producer. J. Biotechnol. 2016, 239, 34–38. [Google Scholar] [CrossRef]
- Garbaye, J. Tansley Review No. 76 Helper Bacteria: A new dimension to the mycorrhizal symbiosis. New Phytol. 1994, 128, 197–210. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Varma, A.; Tuteja, N. Mycorrhizal helper bacteria: Sustainable approach. In Mycorrhiza—Function, Diversity, State of the Art; Springer International Publishing: Cham, Switzerland, 2017; pp. 61–74. [Google Scholar] [CrossRef]
- Meena, V.S.; Kashyap, P.; Nangare, D.D.; Singh, J. Effect of coloured shade nets on yield and quality of pomegranate (Punica granatum) cv. Mridula in semi-arid region of Punjab. Indian J. Agri. Sci. 2016, 86, 500–505. [Google Scholar] [CrossRef]
- Musyoka, D.M.; Njeru, E.M.; Nyamwange, M.M.; Maingi, J.M. Arbuscular mycorrhizal fungi and Bradyrhizobium co-inoculation enhances nitrogen fixation and growth of green grams (Vigna radiata L.) under water stress. J. Plant Nutr. 2020, 43, 1036–1047. [Google Scholar] [CrossRef]
- Splivallo, R.; Vahdatzadeh, M.; MacIá-Vicente, J.G.; Molinier, V.; Peter, M.; Egli, S.; Uroz, S.; Paolocci, F.; Deveau, A. Orchard conditions and fruiting body characteristics drive the microbiome of the black truffle Tuber aestivum. Front. Microbiol. 2019, 10, 439462. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.Academic Press, Elsevier: Amsterdam, The Netherlands, 2008; pp. 1–787. [Google Scholar] [CrossRef]
- Le Tacon, F.; Rubini, A.; Murat, C.; Riccioni, C.; Robin, C.; Belfiori, B.; Zeller, B.; De la Varga, H.; Akroume, E.; Deveau, A.; et al. Certainties and uncertainties about the life cycle of the Périgord black truffle (Tuber melanosporum Vittad.). Ann. For. Sci. 2016, 73, 105–117. [Google Scholar] [CrossRef]
- Kües, U.; Martin, F. On the road to understanding truffles in the underground. Fungal Genet. Biol. 2011, 48, 555–560. [Google Scholar] [CrossRef]
- Zampieri, E.; Rizzello, R.; Bonfante, P.; Mello, A. The detection of mating type genes of Tuber melanosporum in productive and non productive soils. Appl. Soil Ecol. 2012, 57, 9–15. [Google Scholar] [CrossRef]
- Iotti, M.; Piattoni, F.; Zambonelli, A. Techniques for host plant inoculation with truffles and other edible ectomycorrhizal mushrooms. In Edible Ectomycorrhizal Mushrooms—Current Knowledge and Future Prospects; Springer: Berlin/Heidelberg, Germany, 2012; pp. 145–161. [Google Scholar] [CrossRef]
- Amicucci, A.; Barbieri, E.; Sparvoli, V.; Gioacchini, A.M.; Calcabrini, C.; Palma, F.; Stocchi, V.; Zambonelli, A. Microbial and pigment profile of the reddish patch occurring within Tuber magnatum ascomata. Fungal Biol. 2018, 122, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Amicucci, A.; Rossi, I.; Potenza, L.; Zambonelli, A.; Agostini, D.; Palma, F.; Stocchi, V. Identification of ectomycorrhizae from Tuber species by RFLP analysis of the Its region. Biotechnol. Lett. 1996, 18, 821–826. [Google Scholar] [CrossRef]
- Purohit, H.; Raje, D.; Kapley, A. Identification of signature and primers specific to genus Pseudomonas using mismatched patterns of 16S rDNA sequences. BMC Bioinform. 2003, 4, 19. [Google Scholar] [CrossRef]
- Menna, P.; Barcellos, F.G.; Hungria, M. Phylogeny and Taxonomy of a Diverse Collection of Bradyrhizobium Strains Based on Multilocus Sequence Analysis of the 16S rRNA Gene, ITS Region and glnII, recA, atpD and dnaK Genes. Int. J. Syst. Evol. Microbiol. 2009, 59, 2934–2950. [Google Scholar] [CrossRef]
- Zambonelli, A.; Salomoni, S.; Pisi, A. Caratterizzazione anatomo-morfologica delle micorrize di Tuber spp. su Quercus pubescens Willd. Micol. Ital. 1993, 73–90. [Google Scholar]
- Paolocci, F.; Rubini, A.; Granetti, B.; Arcioni, S. Rapid molecular approach for a reliable identification of Tuber spp. ectomycorrhizae. FEMS Microbiol. Ecol. 1999, 28, 23–30. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Polverigiani, S.; McCormack, M.L.; Mueller, C.W.; Eissenstat, D.M. Growth and physiology of olive pioneer and fibrous roots exposed to soil moisture deficits. Tree Physiol. 2011, 31, 1228–1237. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorgi, V.; Amicucci, A.; Landi, L.; Castelli, I.; Romanazzi, G.; Peroni, C.; Ranocchi, B.; Zambonelli, A.; Neri, D. Effect of Bacteria Inoculation on Colonization of Roots by Tuber melanosporum and Growth of Quercus ilex Seedlings. Plants 2024, 13, 224. https://doi.org/10.3390/plants13020224
Giorgi V, Amicucci A, Landi L, Castelli I, Romanazzi G, Peroni C, Ranocchi B, Zambonelli A, Neri D. Effect of Bacteria Inoculation on Colonization of Roots by Tuber melanosporum and Growth of Quercus ilex Seedlings. Plants. 2024; 13(2):224. https://doi.org/10.3390/plants13020224
Chicago/Turabian StyleGiorgi, Veronica, Antonella Amicucci, Lucia Landi, Ivan Castelli, Gianfranco Romanazzi, Cristiano Peroni, Bianca Ranocchi, Alessandra Zambonelli, and Davide Neri. 2024. "Effect of Bacteria Inoculation on Colonization of Roots by Tuber melanosporum and Growth of Quercus ilex Seedlings" Plants 13, no. 2: 224. https://doi.org/10.3390/plants13020224
APA StyleGiorgi, V., Amicucci, A., Landi, L., Castelli, I., Romanazzi, G., Peroni, C., Ranocchi, B., Zambonelli, A., & Neri, D. (2024). Effect of Bacteria Inoculation on Colonization of Roots by Tuber melanosporum and Growth of Quercus ilex Seedlings. Plants, 13(2), 224. https://doi.org/10.3390/plants13020224