

Identification and Validation of the miR156 Family Involved in Drought Responses and Tolerance in Tea Plants (Camellia sinensis (L.) O. Kuntze)


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
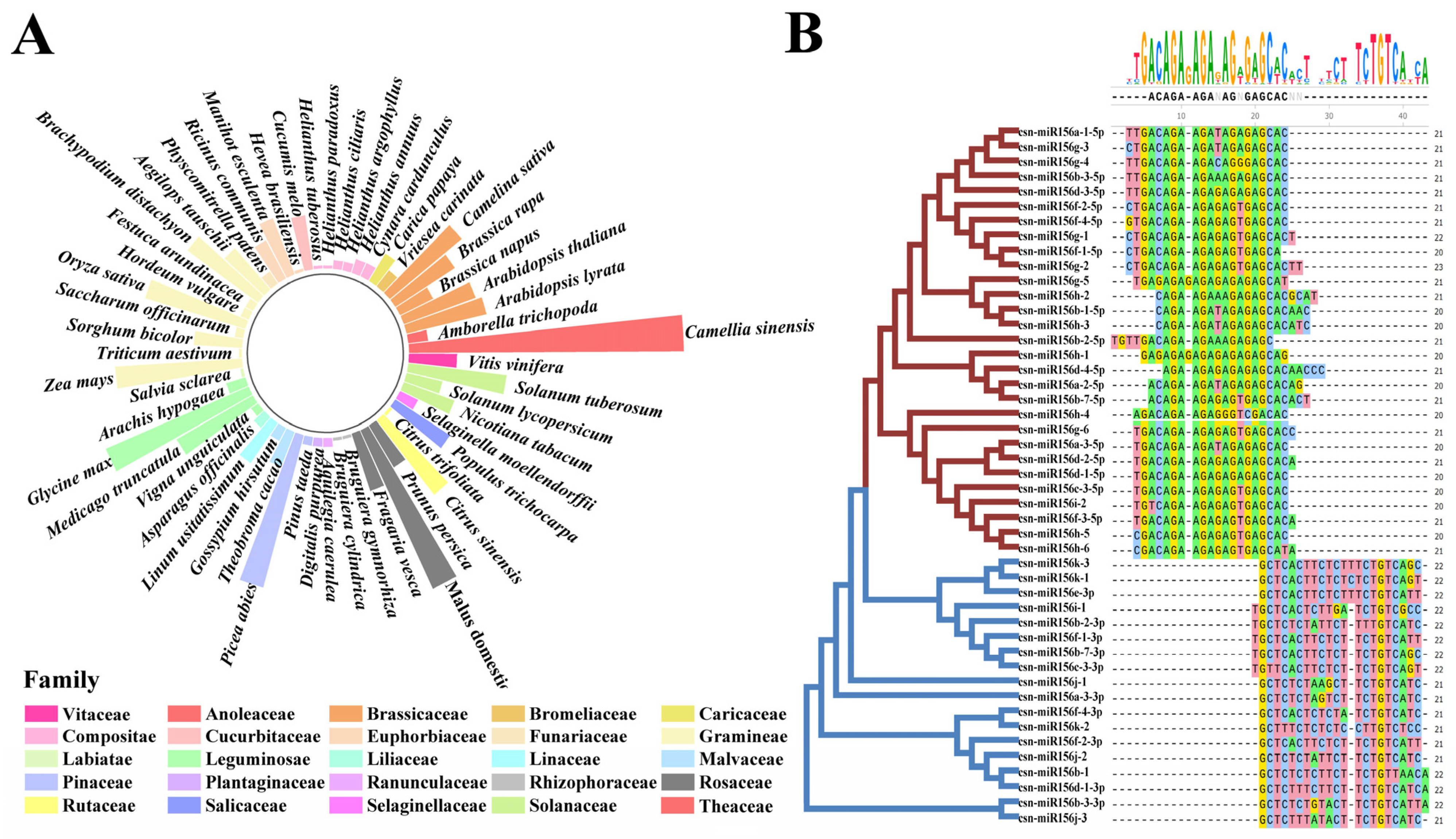
2.1. Identification and Analysis of miR156 in Plants
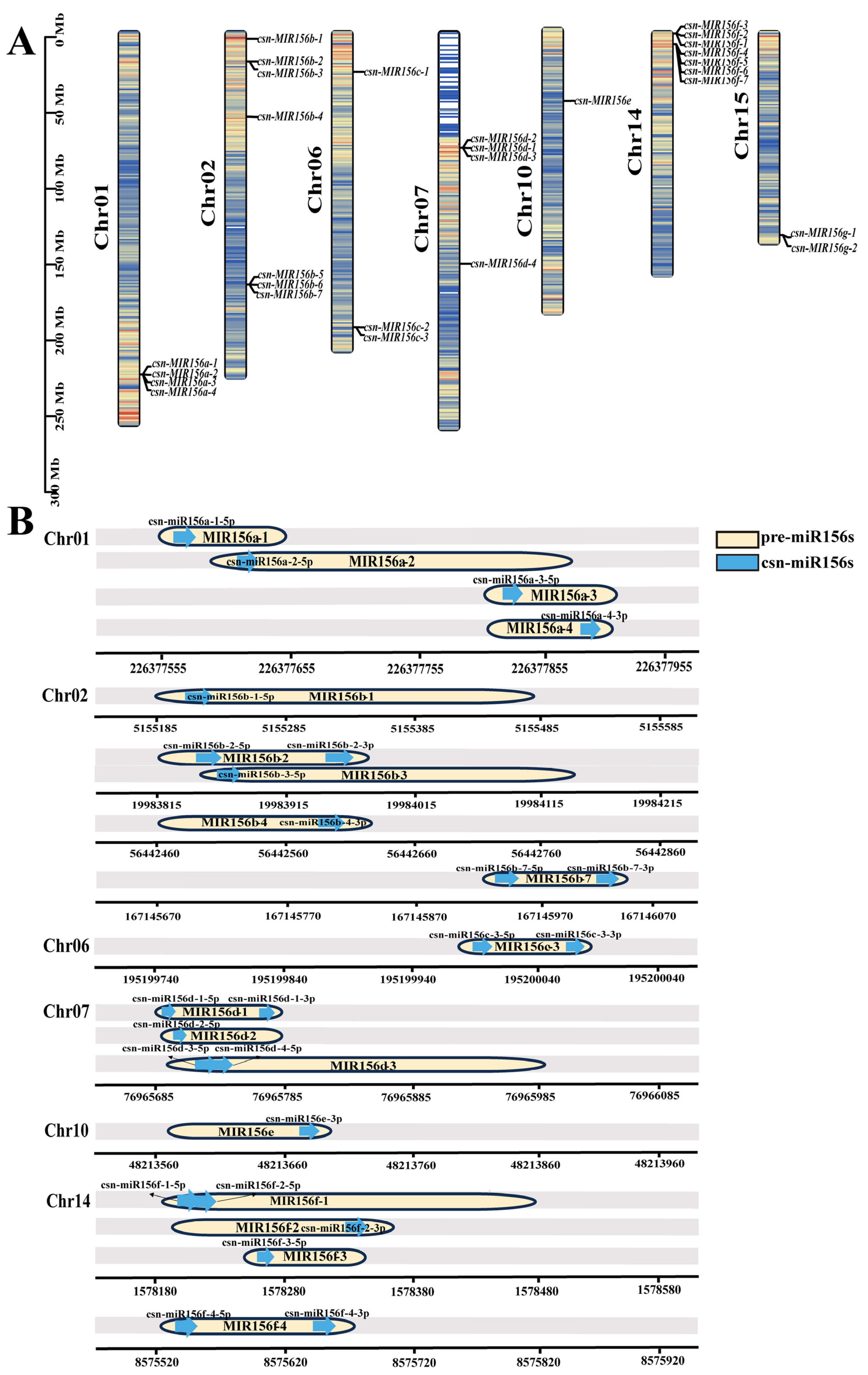
2.2. Sequence Analysis and Chromosomal Localization of csn-MIR156s in the Tea Plant Genome
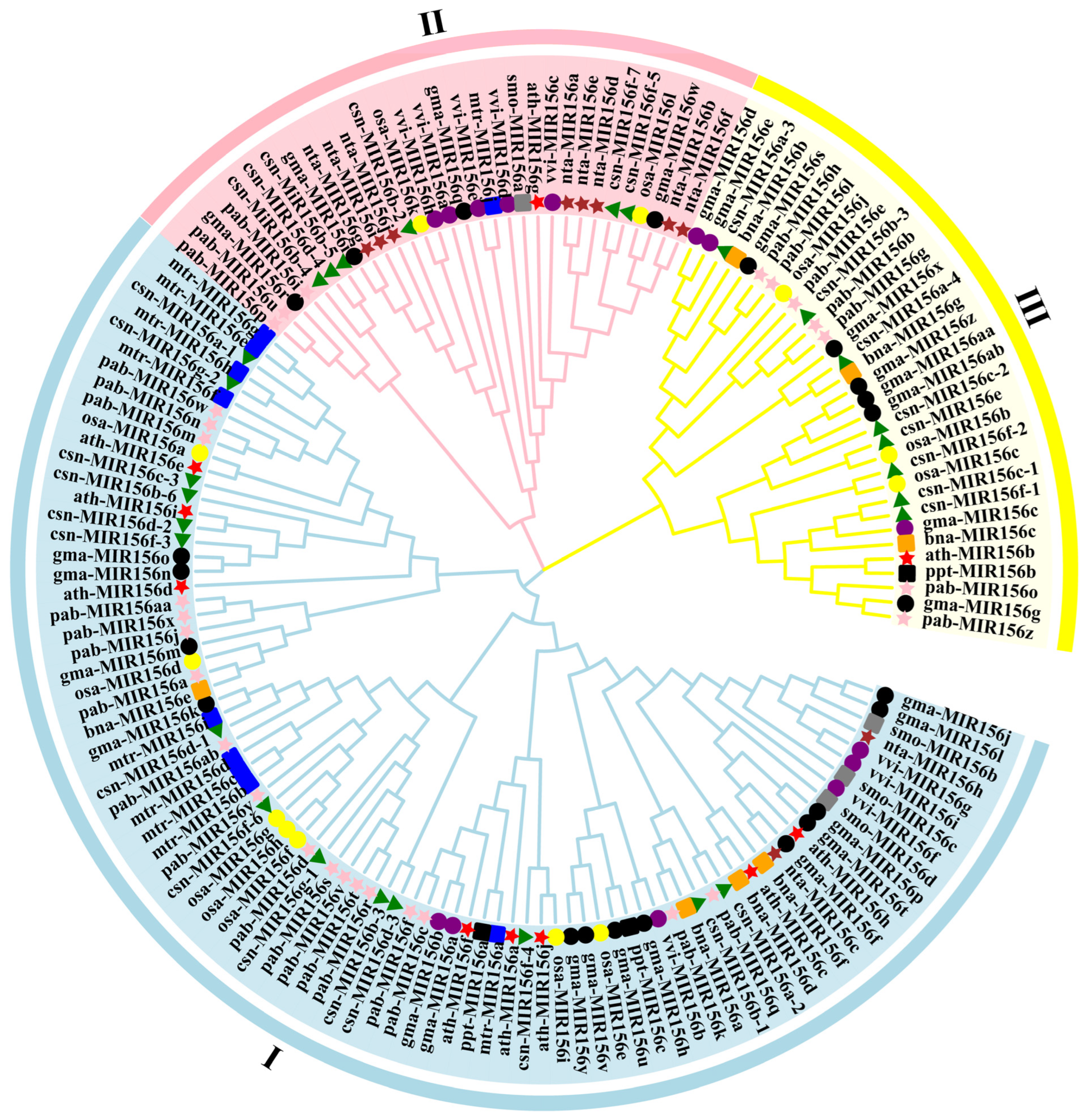
2.3. Predicted Secondary Structure Analysis and Phylogenetic Analysis of csn-MIR156s in the Tea Plant Genome
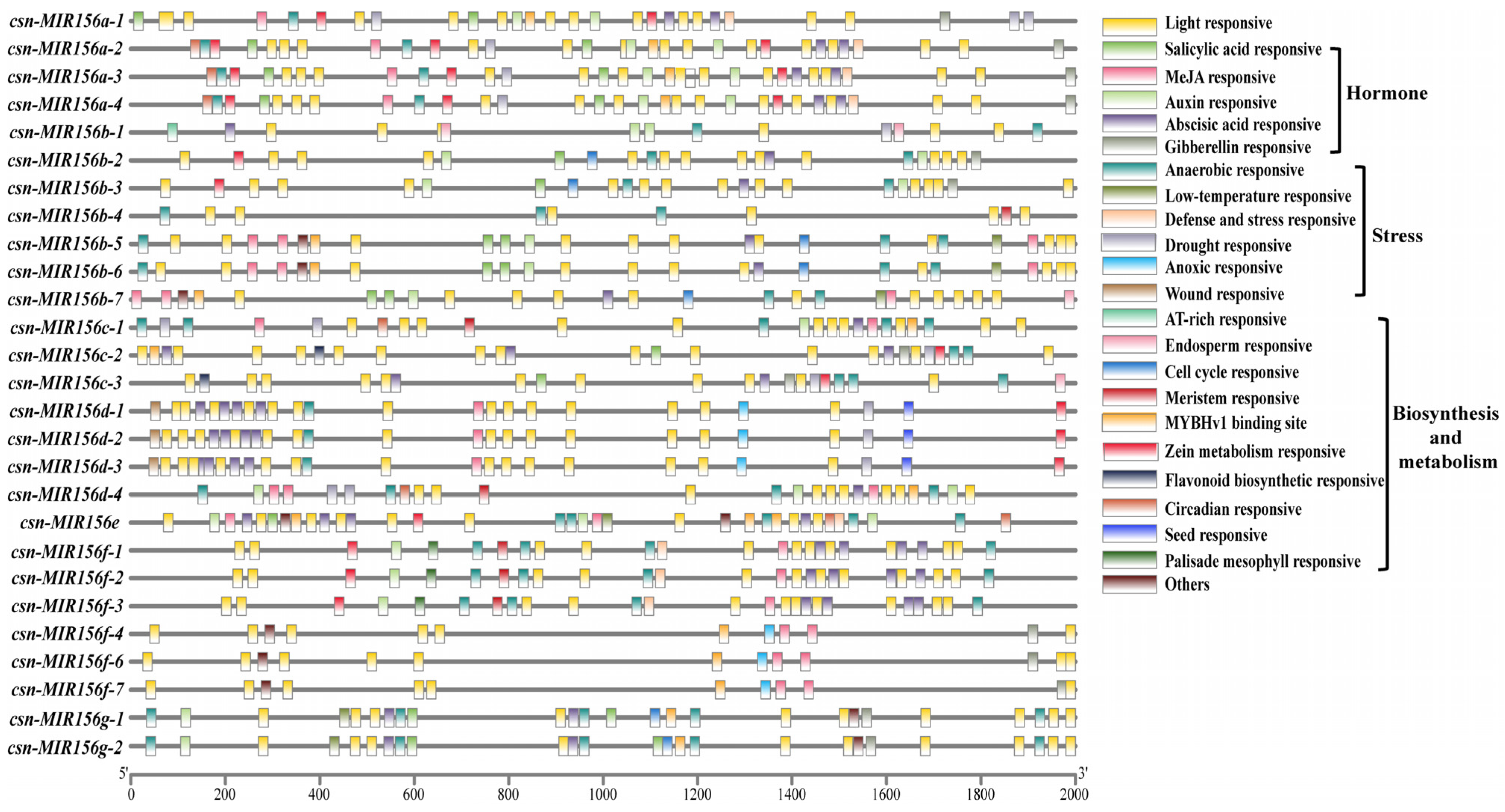
2.4. Analysis of Cis-Acting Elements in CsMIR156s Promoter Regions
2.5. Expression Analysis of csn-miR156s under Drought Stress
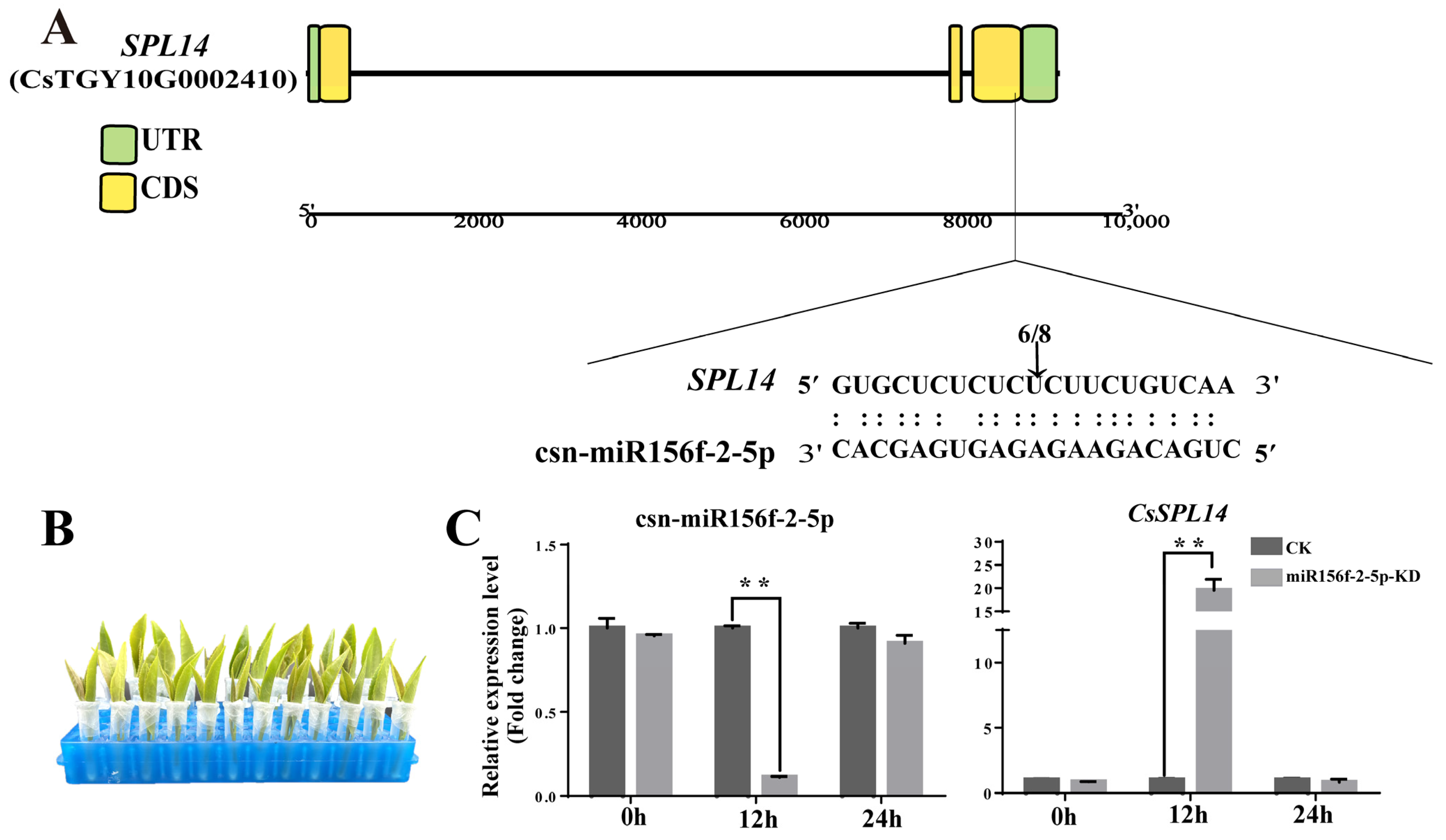
2.6. csn-miR156f-2-5p Participates in Drought Stress Response in Tea Plants
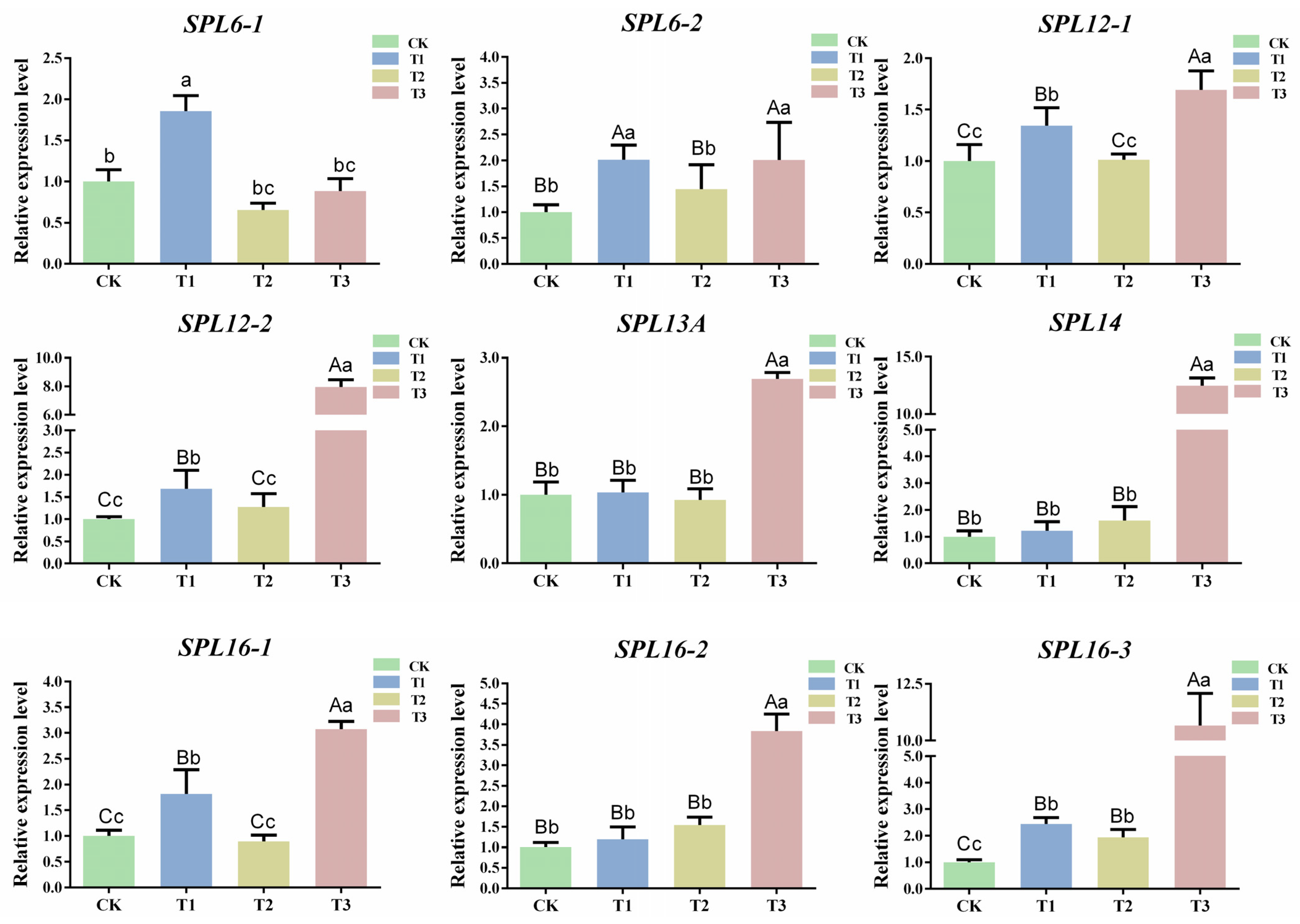
2.7. Prediction and Expression Analysis of csn-miR156f-2-5p Target Genes in Tea Plants
2.8. Csn-miR156f-2-5p Suppresses the Expression of CsSPL14 in Tea Plants
3. Discussion
3.1. Evolutionary Characteristics of the csn-miR156s Family
3.2. Different csn-miR156 Members May Respond to Drought Stress with Different Strategies
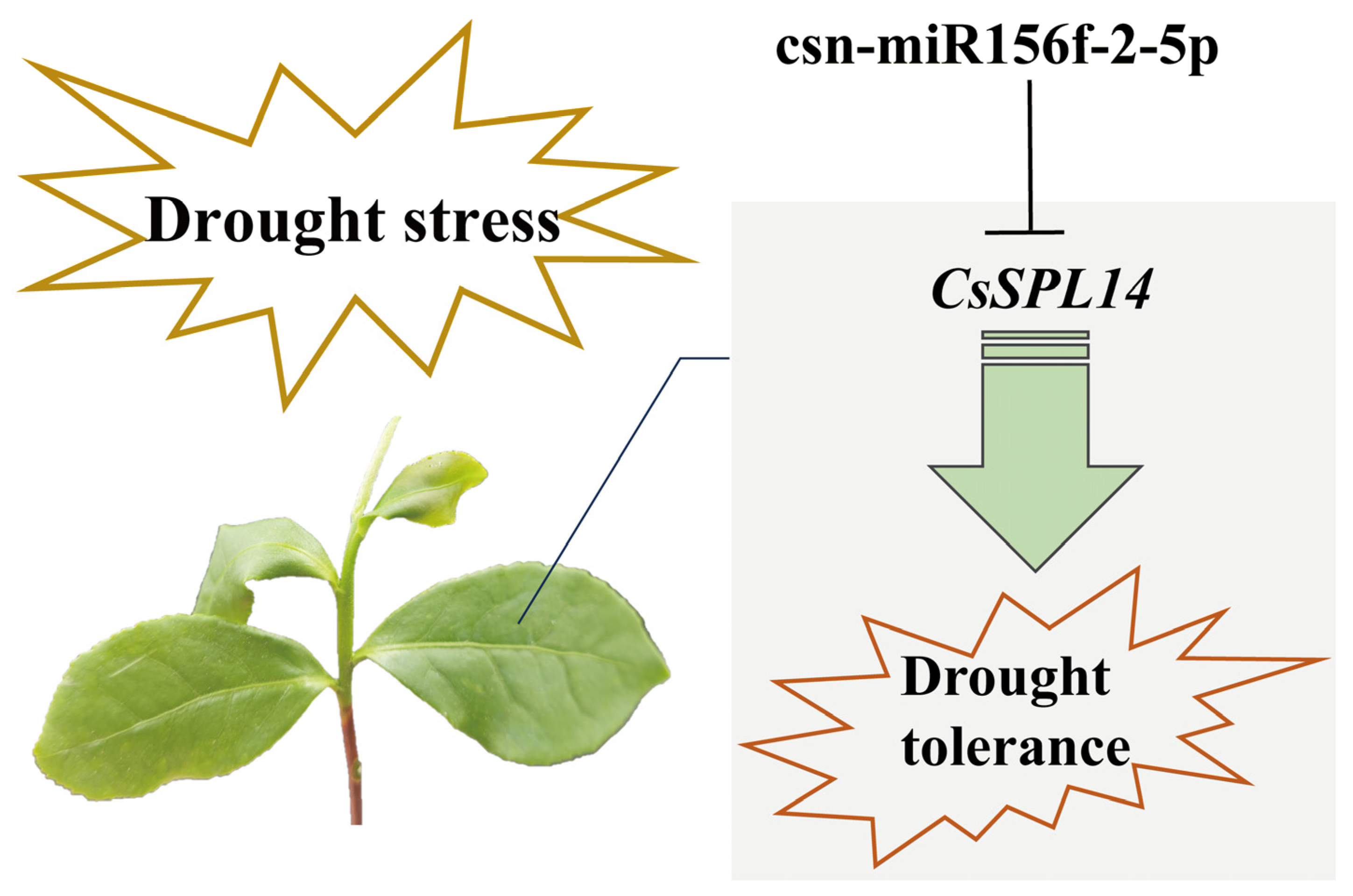
3.3. The csn-miR156 Family May Be Play a Role in the Drought Tolerance Mechanism by Cleaving CsSPLs
4. Materials and Methods
4.1. Plant Materials
4.2. Bioinformatic Analyses of csn-miR156s and Target Gene Prediction
4.3. Suppression of csn-miR156f-2-5p Expression in Tea Leaves Treated with AsODN
4.4. DAB and NBT Staining
4.5. Determination of Fv/Fm, Chlorophyll, and Proline Content
4.6. RNA Extraction and RT-qPCR Analysis
4.7. Cleavage Site Identification with Modified 5′ RLM-RACE
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Christian, J.I.; Basara, J.B.; Hunt, E.D.; Otkin, J.A.; Furtado, J.C.; Mishra, V.; Xiao, X.; Randall, R.M. Global distribution, trends, and drivers of flash drought occurrence. Nat. Commun. 2021, 12, 6330. [Google Scholar] [CrossRef] [PubMed]
- Ault, T.R. On the essentials of drought in a changing climate. Science 2020, 368, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Nadarajah, K.; Kumar, I.S. Drought response in rice: The miRNA story. Int. J. Mol. Sci. 2019, 20, 3766. [Google Scholar] [CrossRef]
- Jin, J.; Zhao, M.; Jing, T.; Wang, J.; Lu, M.; Pan, Y.; Du, W.; Zhao, C.; Bao, Z.; Zhao, W.; et al. (Z)-3-Hexenol integrates drought and cold stress signaling by activating abscisic acid glucosylation in tea plants. Plant Physiol. 2023, 193, 1491–1507. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Y.; Zhang, Y.; Dong, Y.; Liu, Y.; Liu, L.; Wan, S.; He, J.; Yu, Y. Accumulation of galactinol and ABA is involved in exogenous EBR-induced drought tolerance in tea plants. J. Agric. Food Chem. 2022, 70, 13391–13403. [Google Scholar] [CrossRef]
- Zhao, M.; Jin, J.; Wang, J.; Gao, T.; Luo, Y.; Jing, T.; Hu, Y.; Pan, Y.; Lu, M.; Schwab, W.; et al. Eugenol functions as a signal mediating cold and drought tolerance via UGT71A59-mediated glucosylation in tea plants. Plant J. 2022, 109, 1489–1506. [Google Scholar] [CrossRef]
- Zhang, Y.; He, J.; Xiao, Y.; Zhang, Y.; Liu, Y.; Wan, S.; Liu, L.; Dong, Y.; Liu, H.; Yu, Y. CsGSTU8, a glutathione S-transferase from Camellia sinensis, is regulated by CsWRKY48 and plays a positive role in drought tolerance. Front. Plant Sci. 2021, 12, 795919. [Google Scholar] [CrossRef]
- Zhang, Y.; Wan, S.; Liu, X.; He, J.; Cheng, L.; Duan, M.; Liu, H.; Wang, W.; Yu, Y. Overexpression of CsSnRK2.5 increases tolerance to drought stress in transgenic Arabidopsis. Plant Physiol. Bioch. 2020, 150, 162–170. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Y.; Chen, X.; Chen, Y. Plant noncoding RNAs: Hidden players in development and stress responses. Annu. Rev. Cell Dev. Biol. 2019, 35, 407–431. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Cao, X.; Qi, Y. MicroRNAs and their regulatory roles in plant-environment interactions. Annu. Rev. Plant Biol. 2019, 70, 489–525. [Google Scholar] [CrossRef]
- Feyissa, B.A.; Arshad, M.; Gruber, M.Y.; Kohalmi, S.E.; Hannoufa, A. The interplay between miR156/SPL13 and DFR/WD40-1 regulate drought tolerance in alfalfa. Bmc Plant Biol. 2019, 19, 434. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Liu, J.; Huang, J.; Wang, C.; Zhang, L.; Feng, S. Genome-wide identification of miR156 and SPL family genes and phenotypic analysis of vegetative phase change in pepper (Capsicum annuum L.). Gene 2023, 877, 147542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Su, L.Y.; Zhang, S.T.; Xu, X.P.; Chen, X.H.; Li, X.; Jiang, M.Q.; Huang, S.Q.; Chen, Y.K.; Zhang, Z.H.; et al. Analyses of microRNA166 gene structure, expression, and function during the early stage of somatic embryogenesis in Dimocarpus longan Lour. Plant Physiol. Bioch. 2020, 147, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, H.; Zhu, J.; Tao, L.; Wei, C. miR319a targeting of CsTCP10 plays an important role in defense against gray blight disease in tea plant (Camellia sinensis). Tree Physiol. 2022, 42, 1450–1462. [Google Scholar] [CrossRef]
- Hajyzadeh, M.; Turktas, M.; Khawar, K.M.; Unver, T. miR408 overexpression causes increased drought tolerance in chickpea. Gene 2015, 555, 186–193. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, S.; Yu, K.; Wang, H.L.; Xu, H.; Song, C.; Zhao, Y.; Wen, J.; Fu, C.; Li, Y.; et al. Manipulating microRNA miR408 enhances both biomass yield and saccharification efficiency in poplar. Nat. Commun. 2023, 14, 4285. [Google Scholar] [CrossRef]
- Ma, Y.; Xue, H.; Zhang, F.; Jiang, Q.; Yang, S.; Yue, P.; Wang, F.; Zhang, Y.; Li, L.; He, P.; et al. The miR156/SPL module regulates apple salt stress tolerance by activating MdWRKY100 expression. Plant Biotechnol. J. 2021, 19, 311–323. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Guo, Q.; Zhang, X.; Zhou, L.; Zhang, Y.; Zhang, C. Conservation and diversity of miR166 family members from highbush blueberry (Vaccinium corymbosum) and their potential functions in abiotic stress. Front. Genet. 2022, 13, 919856. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Yan, J.; Wang, K.; Luo, H.; Zhang, W. MiR319-mediated ethylene biosynthesis, signalling and salt stress response in switchgrass. Plant Biotechnol. J. 2019, 17, 2370–2383. [Google Scholar] [CrossRef] [PubMed]
- Hang, N.; Shi, T.; Liu, Y.; Ye, W.; Taier, G.; Sun, Y.; Wang, K.; Zhang, W. Overexpression of Os-microRNA408 enhances drought tolerance in perennial ryegrass. Physiol. Plantarum. 2021, 172, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.R.; Baurle, I. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef]
- Visentin, I.; Pagliarani, C.; Deva, E.; Caracci, A.; Tureckova, V.; Novak, O.; Lovisolo, C.; Schubert, A.; Cardinale, F. A novel strigolactone-miR156 module controls stomatal behaviour during drought recovery. Plant Cell Environ. 2020, 43, 1613–1624. [Google Scholar] [CrossRef] [PubMed]
- Brun, G. At the crossroads of strigolactones and abscisic acid pathways: A role for miR156. Plant Cell Environ. 2020, 43, 1609–1612. [Google Scholar] [CrossRef]
- Jeyaraj, A.; Zhang, X.; Hou, Y.; Shangguan, M.; Gajjeraman, P.; Li, Y.; Wei, C. Genome-wide identification of conserved and novel microRNAs in one bud and two tender leaves of tea plant (Camellia sinensis) by small RNA sequencing, microarray-based hybridization and genome survey scaffold sequences. Bmc Plant Biol. 2017, 17, 212. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, X.; Chen, X.; Song, C.; Zou, Z.; Wang, Y.; Wang, M.; Fang, W.; Li, X. Identification and characterization of cold-responsive microRNAs in tea plant (Camellia sinensis) and their targets using high-throughput sequencing and degradome analysis. Bmc Plant Biol. 2014, 14, 271. [Google Scholar] [CrossRef]
- Zhao, S.; Mi, X.; Guo, R.; Xia, X.; Liu, L.; An, Y.; Yan, X.; Wang, S.; Guo, L.; Wei, C. The biosynthesis of main taste compounds Is coordinately regulated by miRNAs and phytohormones in tea plant (Camellia sinensis). J. Agric. Food Chem. 2020, 68, 6221–6236. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, S.; Zhu, C.; Chang, X.; Yue, C.; Wang, Z.; Lin, Y.; Lai, Z. Identification of drought-responsive miRNAs and physiological characterization of tea plant (Camellia sinensis L.) under drought stress. Bmc Plant Biol. 2017, 17, 211. [Google Scholar] [CrossRef]
- Fan, K.; Fan, D.; Ding, Z.; Su, Y.; Wang, X. Cs-miR156 is involved in the nitrogen form regulation of catechins accumulation in tea plant (Camellia sinensis L.). Plant Physiol. Bioch. 2015, 97, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J.; Meyers, B.C. Revisiting criteria for plant microRNA annotation in the era of big data. Plant Cell 2018, 30, 272–284. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, Q.; Zeng, Y.; Chen, X.; Zhang, Z.; Chen, Y.; Lai, Z. Molecular evolution and spatiotemporal expression of longan miR166 family. J. Hortic. 2017, 44, 2285–2295. [Google Scholar]
- Xie, Q.; Wang, X.; He, J.; Lan, T.; Zheng, J.; Li, Y.; Pan, J.; Lin, L.; Zhao, J.; Li, J.; et al. Distinct evolutionary profiles and functions of microRNA156 and microRNA529 in land plants. Int. J. Mol. Sci. 2021, 22, 11100. [Google Scholar] [CrossRef] [PubMed]
- Pei, M.; Liu, H.; Wei, T.; Jin, H.; Yu, Y.; Ma, M.; Song, X.; Dai, R.; Guo, D. Identification, characterization, and verification of miR399 target gene in grape. Hortic. Plant J. 2023, 10, 91–102. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, H.; Jiang, H.L.; Wang, H.S.; Chen, K.X.; Duan, J.J.; Feng, S.J.; Wu, G. Regulation of cadmium tolerance and accumulation by miR156 in Arabidopsis. Chemosphere 2020, 242, 125168. [Google Scholar] [CrossRef] [PubMed]
- Gramzow, L.; Theissen, G. Plant miRNA conservation and evolution. Methods Mol. Biol. 2019, 1932, 41–50. [Google Scholar]
- Wang, H.; Lu, Z.; Xu, Y.; Kong, L.; Shi, J.; Liu, Y.; Fu, C.; Wang, X.; Wang, Z.Y.; Zhou, C.; et al. Genome-wide characterization of SPL family in Medicago truncatula reveals the novel roles of miR156/SPL module in spiky pod development. Bmc Genom. 2019, 20, 552. [Google Scholar] [CrossRef]
- Naya, L.; Khan, G.A.; Sorin, C.; Hartmann, C.; Crespi, M.; Lelandais-Briere, C. Cleavage of a non-conserved target by a specific miR156 isoform in root apexes of Medicago truncatula. Plant Signal Behav. 2010, 5, 328–331. [Google Scholar] [CrossRef]
- Ding, X.; Guo, J.; Lv, M.; Wang, H.; Sheng, Y.; Liu, Y.; Gai, J.; Yang, S. The miR156b-GmSPL2b module mediates male fertility regulation of cytoplasmic male sterility-based restorer line under high-temperature stress in soybean. Plant Biotechnol. J. 2023, 21, 1542–1559. [Google Scholar] [CrossRef]
- Yin, H.; Hong, G.; Li, L.; Zhang, X.; Kong, Y.; Sun, Z.; Li, J.; Chen, J.; He, Y. miR156/SPL9 regulates reactive oxygen species accumulation and immune response in Arabidopsis thaliana. Phytopathology 2019, 109, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ming, L.; Liao, K.; Xia, C.; Sun, S.; Chang, Y.; Wang, H.; Fu, D.; Xu, C.; Wang, Z.; et al. Bract suppression regulated by the miR156/529-SPLs-NL1-PLA1 module is required for the transition from vegetative to reproductive branching in rice. Mol. Plant 2021, 14, 1168–1184. [Google Scholar] [CrossRef]
- Ellis, J.; Dodds, P.; Pryor, T. Structure, function and evolution of plant disease resistance genes. Curr. Opin. Plant Biol. 2000, 3, 278–284. [Google Scholar] [CrossRef]
- Eldem, V.; Celikkol, A.U.; Ozhuner, E.; Bakir, Y.; Uranbey, S.; Unver, T. Genome-wide identification of miRNAs responsive to drought in peach (Prunus persica) by high-throughput deep sequencing. PLoS ONE 2012, 7, e50298. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Zhou, C.; Zhu, C.; Chen, L.; Shi, B.; Lin, Y.; Lai, Z.; Guo, Y. Genome-wide investigation of the miR166 family provides new insights into Its involvement in the drought stress responses of tea plants (Camellia sinensis (L.) O. Kuntze). Forests 2022, 13, 628. [Google Scholar] [CrossRef]
- Trindade, I.; Capitao, C.; Dalmay, T.; Fevereiro, M.P.; Santos, D.M. miR398 and miR408 are up-regulated in response to water deficit in Medicago truncatula. Planta 2010, 231, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, L.; Zhao, M.; Tian, Q.; Zhang, W.H. Identification of drought-responsive microRNAs in Medicago truncatula by genome-wide high-throughput sequencing. Bmc Genom. 2011, 12, 367. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Y.; Liu, X.; Duan, S.; Jiang, S.; Zhu, J.; Zhang, Y.; Hou, H. The mdmiR156n regulates drought tolerance and flavonoid synthesis in apple calli and Arabidopsis. Int. J. Mol. Sci. 2023, 24, 6049. [Google Scholar] [CrossRef]
- Feng, C.; Zhang, X.; Du, B.; Xiao, Y.; Wang, Y.; Sun, Y.; Zhou, X.; Wang, C.; Liu, Y.; Li, T.H. MicroRNA156ab regulates apple plant growth and drought tolerance by targeting transcription factor MsSPL13. Plant Physiol. 2023, 192, 1836–1857. [Google Scholar] [CrossRef]
- Cui, L.; Zheng, F.; Wang, J.; Zhang, C.; Xiao, F.; Ye, J.; Li, C.; Ye, Z.; Zhang, J. miR156a-targeted SBP-Box transcription factor SlSPL13 regulates inflorescence morphogenesis by directly activating SFT in tomato. Plant Biotechnol. J. 2020, 18, 1670–1682. [Google Scholar] [CrossRef]
- Li, S.; Cheng, Z.; Li, Z.; Dong, S.; Yu, X.; Zhao, P.; Liao, W.; Yu, X.; Peng, M. MeSPL9 attenuates drought resistance by regulating JA signaling and protectant metabolite contents in cassava. Theor. Appl. Genet. 2022, 135, 817–832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, S.; Shi, L.; Gong, D.; Zhang, S.; Zhao, Q.; Zhan, D.; Vasseur, L.; Wang, Y.; Yu, J.; et al. Haplotype-resolved genome assembly provides insights into evolutionary history of the tea plant Camellia sinensis. Nat. Genet. 2021, 53, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, S.; Liu, L.; Li, R.; Guo, R.; Xia, X.; Wei, C. miR477 targets the phenylalanine ammonia-lyase gene and enhances the susceptibility of the tea plant (Camellia sinensis) to disease during Pseudopestalotiopsis species infection. Planta 2020, 251, 59. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, S.; Zhou, C.; Tian, C.; Yang, N.; Zhang, C.; Zheng, A.; Chen, Y.; Lai, Z.; Guo, Y. Identification and Validation of the miR156 Family Involved in Drought Responses and Tolerance in Tea Plants (Camellia sinensis (L.) O. Kuntze). Plants 2024, 13, 201. https://doi.org/10.3390/plants13020201
Wen S, Zhou C, Tian C, Yang N, Zhang C, Zheng A, Chen Y, Lai Z, Guo Y. Identification and Validation of the miR156 Family Involved in Drought Responses and Tolerance in Tea Plants (Camellia sinensis (L.) O. Kuntze). Plants. 2024; 13(2):201. https://doi.org/10.3390/plants13020201
Chicago/Turabian StyleWen, Shengjing, Chengzhe Zhou, Caiyun Tian, Niannian Yang, Cheng Zhang, Anru Zheng, Yixing Chen, Zhongxiong Lai, and Yuqiong Guo. 2024. "Identification and Validation of the miR156 Family Involved in Drought Responses and Tolerance in Tea Plants (Camellia sinensis (L.) O. Kuntze)" Plants 13, no. 2: 201. https://doi.org/10.3390/plants13020201
APA StyleWen, S., Zhou, C., Tian, C., Yang, N., Zhang, C., Zheng, A., Chen, Y., Lai, Z., & Guo, Y. (2024). Identification and Validation of the miR156 Family Involved in Drought Responses and Tolerance in Tea Plants (Camellia sinensis (L.) O. Kuntze). Plants, 13(2), 201. https://doi.org/10.3390/plants13020201