

Enhancing Commercial Antibiotics with Trans-Cinnamaldehyde in Gram-Positive and Gram-Negative Bacteria: An In Vitro Approach

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Toxicity Study
2.2. Antimicrobial Susceptibility Test
- Antibiotics with a MIC for a given bacteria below 10 μg/mL were discarded. These results were considered difficult to improve in combination with a synergistic compound.
- Bacteria that were susceptible to DMSO at a concentration below 2.5% in the toxicity study were excluded because of the impossibility of solubilizing CIN in pure water.
2.3. Checkerboard Assay
2.4. Kinetic Growth Assay
3. Discussion
3.1. Antimicrobial Activity Analysis of Cinnamaldehyde and Commercial Antibiotics
3.1.1. Gram-Positive
3.1.2. Gram-Negative
3.2. Assessment of the Combination
3.2.1. Beta-Lactams
3.2.2. Aminoglycosides
3.2.3. Macrolides and Amphenicoles
3.3. Kinetics
4. Materials and Methods
4.1. Bacterial Strains
4.2. Compounds to Be Tested and Stock Solutions
4.3. Toxicity and Antimicrobial Susceptibility Tests
4.4. Checkerboard Assay
4.5. Kinetic Growth Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Action Plan on Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2015.
- Bell, B.G.; Schellevis, F.; Stobberingh, E.; Goossens, H.; Pringle, M. A systematic review and meta-analysis of the effects of antibiotic consumption on antibiotic resistance. BMC Infect. Dis. 2014, 14, 13. [Google Scholar] [CrossRef]
- Zhang, L.; Song, J.K.; Kong, L.L.; Yuan, T.Y.; Li, W.; Zhang, W.; Hou, B.Y.; Lu, Y.; Du, G.H. The strategies and techniques of drug discovery from natural products. Pharmacol. Ther. 2020, 216, 107686. [Google Scholar] [CrossRef] [PubMed]
- Neelam; Khatkar, A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. Nutr. 2020, 60, 2655–2675. [Google Scholar] [CrossRef]
- Chopra, B.; Dhingra, A.K. Natural products: A lead for drug discovery and development. Phytother. Res. 2021, 35, 4660–4702. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, R.; Moser, H.E. Physicochemical properties of antibacterial compounds: Implications for drug discovery. J. Med. Chem. 2008, 51, 2871–2878. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.G.; Lister, T.; May-Dracka, T.L. New natural products as new leads for antibacterial drug discovery. Bioorgan. Med. Chem. Lett. 2014, 24, 413–418. [Google Scholar] [CrossRef]
- Allemailem, K.S. Antimicrobial Potential of Naturally Occurring Bioactive Secondary Metabolites. J. Pharm. Bioallied Sci. 2021, 13, 155–162. [Google Scholar] [CrossRef]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Abreu, A.C.; McBain, A.J.; Simoes, M. Plants as sources of new antimicrobials and resistance-modifying agents. Nat. Prod. Rep. 2012, 29, 1007–1021. [Google Scholar] [CrossRef]
- Thirapanmethee, K.; Kanathum, P.; Khuntayaporn, P.; Huayhongthong, S.; Surassmo, S.; Chomnawang, M.T. Cinnamaldehyde: A plant-derived antimicrobial for overcoming multidrug-resistant Acinetobacter baumannii infection. Eur. J. Integr. Med. 2021, 48, 101376. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G.; Sucher, N.; Wang, Z.J.; Awang, D.V.C. Synergy Research: Natural Products for rational comedication with Chemotherapeutics and Antibiotics. Phytomedicine 2010, 18, 1. [Google Scholar] [CrossRef]
- Yang, S.K.; Yusoff, K.; Mai, C.W.; Lim, W.M.; Yap, W.S.; Lim, S.H.E.; Lai, K.S. Additivity vs. Synergism: Investigation of the Additive Interaction of Cinnamon Bark Oil and Meropenem in Combinatory Therapy. Molecules 2017, 22, 1733. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.W. Alternative natural sources for a new generation of antibacterial agents. Int. J. Antimicrob. Agents 2013, 42, 195–201. [Google Scholar] [CrossRef]
- Ma, J.K.C.; Chikwamba, R.; Sparrow, P.; Fischer, R.; Mahoney, R.; Twyman, R.M. Plant-derived pharmaceuticals-the road forward. Trends Plant Sci. 2005, 10, 580–585. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO Expert Committee on Food Additives (JECFA); Nutrition and Food Safety (NFS). Specifications for the Identity and Purity of Food Addictives and Their Toxicological Evaluation: Some Flavoring Substances and Non-Nutritive Sweetening Agents; World Health Organization: Geneva, Switzerland, 1967.
- Council of Europe. Natural Flavouring Substances, Their Sources, and Added Artificial Flavouring Substances; The Council of Europe: Strasbourg, France, 1974. [Google Scholar]
- Flavoring Extract Manufacturers’ Association. Survey of flavoring ingredients usage levels. Fd Technol. Champaign 1965, 19, 155. [Google Scholar]
- Vasconcelos, N.G.; Croda, J.; Simionatto, S. Antibacterial mechanisms of cinnamon and its constituents: A review. Microb. Pathog. 2018, 120, 198–203. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, L.D.; de Moraes, A.A.B.; da Costa, K.S.; Galucio, J.M.P.; Taube, P.S.; Costa, C.M.L.; Cruz, J.N.; Andrade, E.H.D.; de Faria, L.J.G. Bioactive Natural Compounds and Antioxidant Activity of Essential Oils from Spice Plants: New Findings and Potential Applications. Biomolecules 2020, 10, 988. [Google Scholar] [CrossRef]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.F.; Hsu, H.Y.; Cheng, S.S.; Lin, I.F.; Chen, C.J.; Chen, S.T.; Chang, S.T. Cinnamaldehyde inhibits pro-inflammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signaling. Food Chem. Toxicol. 2008, 46, 220–231. [Google Scholar] [CrossRef]
- Li, R.; Liang, T.; Xu, L.Y.; Li, Y.W.; Zhang, S.J.; Duan, X.Q. Protective effect of cinnamon polyphenols against STZ-diabetic mice fed high-sugar, high-fat diet and its underlying mechanism. Food Chem. Toxicol. 2013, 51, 419–425. [Google Scholar] [CrossRef]
- Anand, P.; Murali, K.Y.; Tandon, V.; Murthy, P.S.; Chandra, R. Insulinotropic effect of cinnamaldehyde on transcriptional regulation of pyruvate kinase, phosphoenolpyruvate carboxykinase, and GLUT4 translocation in experimental diabetic rats. Chem.-Biol. Interact. 2010, 186, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, K.; Holley, R.A. Use of natural antimicrobials to increase antibiotic susceptibility of drug resistant bacteria. Int. J. Food Microbiol. 2010, 140, 164–168. [Google Scholar] [CrossRef]
- Sim, J.X.F.; Khazandi, M.; Pi, H.; Venter, H.; Trott, D.J.; Deo, P. Antimicrobial effects of cinnamon essential oil and cinnamaldehyde combined with EDTA against canine otitis externa pathogens. J. Appl. Microbiol. 2019, 127, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Topa, S.H.; Palombo, E.A.; Kingshott, P.; Blackall, L.L. Activity of Cinnamaldehyde on Quorum Sensing and Biofilm Susceptibility to Antibiotics in Pseudomonas aeruginosa. Microorganisms 2020, 8, 455. [Google Scholar] [CrossRef]
- Ferro, T.A.F.; Souza, E.B.; Suarez, M.A.M.; Rodrigues, J.F.S.; Pereira, D.M.S.; Mendes, S.J.F.; Gonzaga, L.F.; Machado, M.C.A.M.; Bomfim, M.R.Q.; Calixto, J.B.; et al. Topical Application of Cinnamaldehyde Promotes Faster Healing of Skin Wounds Infected with Pseudomonas aeruginosa. Molecules 2019, 24, 1627. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Fornari, F.; Riboni, N.; Spadini, C.; Cabassi, C.S.; Iannarelli, M.; Carraro, C.; Mazzeo, P.P.; Bacchi, A.; Orlandini, S.; et al. Development of novel cocrystal-based active food packaging by a Quality by Design approach. Food Chem. 2021, 347, 129051. [Google Scholar] [CrossRef]
- Institute CaLS. M100. Performance Standards for Antimicrobial Susceptibility Testing; Institute CaLS: Gainesville, FL, USA, 2020. [Google Scholar]
- Oliva, A.; Tafin, U.F.; Maiolo, E.M.; Jeddari, S.; Betrisey, B.; Trampuz, A. Activities of Fosfomycin and Rifampin on Planktonic and Adherent Enterococcus faecalis Strains in an Experimental Foreign-Body Infection Model. Antimicrob. Agents Chemother. 2014, 58, 1284–1293. [Google Scholar] [CrossRef]
- Chai, W.C.; Whittall, J.J.; Polyak, S.W.; Foo, K.; Li, X.; Dutschke, C.J.; Ogunniyi, A.D.; Ma, S.T.; Sykes, M.J.; Semple, S.J.; et al. Cinnamaldehyde derivatives act as antimicrobial agents against Acinetobacter baumannii through the inhibition of cell division. Front. Microbiol. 2022, 13, 967949. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of Minimum Inhibitory Concentrations (MICs) of Antibacterial Agents by Broth Dilution; Wiley Online Library: Hoboken, NJ, USA, 2003. [Google Scholar]
- Stefanovic, O.D.; Stanojevic, D.D.; Comic, L.R. Synergistic Antibacterial Activity of Salvia Officinalis and Cichorium Intybus Extracts And Antibiotics. Acta Pol. Pharm. 2012, 69, 457–463. [Google Scholar]
- Jugreet, B.S.; Mahomoodally, M.F. Essential oils from 9 exotic and endemic medicinal plants from Mauritius shows in vitro antibacterial and antibiotic potentiating activities. S. Afr. J. Bot. 2020, 132, 355–362. [Google Scholar] [CrossRef]
- Cardoso, K.; Gandra, R.F.; Wisniewski, E.S.; Osaku, C.A.; Kadowaki, M.K.; Felipach-Neto, V.; Haus, L.; Simao, R.D.G. DnaK and GroEL are induced in response to antibiotic and heat shock in Acinetobacter baumannii. J. Med. Microbiol. 2010, 59, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Cestellos-Blanco, S.; Inoue, K.; Zare, R.N. Miniaturized Antimicrobial Susceptibility Test by Combining Concentration Gradient Generation and Rapid Cell Culturing. Antibiotics 2015, 4, 455–466. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, P.G.G.; Lemos, T.L.G.; Bizerra, A.M.C.; Arriaga, A.M.C.; Ferreira, D.A.; Santiago, G.M.P.; Braz Filho, R.; Costa, J.G.M. Antibacterial and Antioxidant Activities of Ursolic Acid and Derivatives. Molecules 2014, 19, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Lv, P.C.; Xiao, Z.P.; Fang, R.Q.; Li, H.Q.; Zhu, H.L.; Liu, C.H. Synthesis, characterization and structure-activity relationship analysis of novel depsides as potential antibacterials. Eur. J. Med. Chem. 2009, 44, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.I.; Braga, T.M.; Silva, P.; Fernandes, J.A.; Ribeiro-Claro, P.; Lopes, M.D.S.; Paz, F.A.A.; Braga, S.S. Chloramphenicol center dot cyclodextrin inclusion compounds: Co-dissolution and mechanochemical preparations and antibacterial action. Crystengcomm 2013, 15, 2822–2834. [Google Scholar] [CrossRef]
- Wang, J.F.; Zhou, Z.H.; He, F.; Ruan, Z.; Jiang, Y.; Hua, X.T.; Yu, Y.S. The role of the type VI secretion system vgrG gene in the virulence and antimicrobial resistance of Acinetobacter baumannii ATCC 19606. PLoS ONE 2018, 13, e0192288. [Google Scholar] [CrossRef]
- Chollet, R.; Chevalier, J.; Bollet, C.; Pages, J.M.; Davin-Regli, A. RamA is an alternate activator of the multidrug resistance cascade in Enterobacter aerogenes. Antimicrob. Agents Chemother. 2004, 48, 2518–2523. [Google Scholar] [CrossRef]
- Yurttas, L.; Ozkay, Y.; Kaplancikli, Z.A.; Tunali, Y.; Karaca, H. Synthesis and antimicrobial activity of some new hydrazone-bridged thiazole-pyrrole derivatives. J. Enzym. Inhib. Med. Chem. 2013, 28, 830–835. [Google Scholar] [CrossRef]
- Maseda, H.; Hashida, Y.; Konaka, R.; Shirai, A.; Kourai, H. Mutational Upregulation of a Resistance-Nodulation-Cell Division-Type Multidrug Efflux Pump, SdeAB, upon Exposure to a Biocide, Cetylpyridinium Chloride, and Antibiotic Resistance in Serratia marcescens. Antimicrob. Agents Chemother. 2009, 53, 5230–5235. [Google Scholar] [CrossRef]
- Eliuz, E.A.E.; Ayas, D.; Goksen, G. In Vitro Phototoxicity and Antimicrobial Activity of Volatile Oil Obtained from Some Aromatic Plants. J. Essent. Oil Bear. Plants 2017, 20, 758–768. [Google Scholar] [CrossRef]
- Barbarossa, A.; Sblano, S.; Rosato, A.; Carrieri, A.; Corbo, F.; Clodoveo, M.L.; Fracchiolla, G.; Carocci, A. Synergistic Action of Cinnamomum verum Essential Oil with Sertraline. Antibiotics 2022, 11, 1617. [Google Scholar] [CrossRef]
- Kerekes, E.B.; Vidacs, A.; Tako, M.; Petkovits, T.; Vagvolgyi, C.; Horvath, G.; Balazs, V.L.; Krisch, J. Anti-Biofilm Effect of Selected Essential Oils and Main Components on Mono- and Polymicrobic Bacterial Cultures. Microorganisms 2019, 7, 345. [Google Scholar] [CrossRef]
- Liu, Q.; Niu, H.; Zhang, W.; Mu, H.; Sun, C.; Duan, J. Synergy among thymol, eugenol, berberine, cinnamaldehyde and streptomycin against planktonic and biofilm-associated food-borne pathogens. Lett. Appl. Microbiol. 2015, 60, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, L.; Han, J.; Dong, P.; Luo, X.; Zhang, Y.; Zhu, L. Inhibition of Biofilm Formation and Related Gene Expression of Listeria monocytogenes in Response to Four Natural Antimicrobial Compounds and Sodium Hypochlorite. Front. Microbiol. 2021, 11, 617473. [Google Scholar] [CrossRef] [PubMed]
- Neri, F.; Mari, M.; Brigati, S. Control of Penicillium expansum by plant volatile compounds. Plant Pathol. 2006, 55, 100–105. [Google Scholar] [CrossRef]
- Garcia-Salinas, S.; Elizondo-Castillo, H.; Arruebo, M.; Mendoza, G.; Irusta, S. Evaluation of the Antimicrobial Activity and Cytotoxicity of Different Components of Natural Origin Present in Essential Oils. Molecules 2018, 23, 1399. [Google Scholar] [CrossRef] [PubMed]
- Rani, S.; Verma, S.; Singh, H.; Ram, C. Antibacterial activity and mechanism of essential oils in combination with medium-chain fatty acids against predominant bovine mastitis pathogens. Lett. Appl. Microbiol. 2022, 74, 959–969. [Google Scholar] [CrossRef]
- Ali, I.A.A.; Matinlinna, J.P.; Levesque, C.M.; Neelakantan, P. Trans-Cinnamaldehyde Attenuates Enterococcus faecalis Virulence and Inhibits Biofilm Formation. Antibiotics 2021, 10, 702. [Google Scholar] [CrossRef]
- Ferro, T.A.F.; Araujo, J.M.M.; Pinto, B.L.D.; dos Santos, J.S.; Souza, E.B.; da Silva, B.L.R.; Colares, V.L.P.; Novais, T.M.G.; Filho, C.M.B.; Struve, C.; et al. Cinnamaldehyde Inhibits Staphylococcus aureus Virulence Factors and Protects against Infection in a Galleria mellonella Model. Front. Microbiol. 2016, 7, 2052. [Google Scholar] [CrossRef]
- Huycke, M.M.; Gilmore, M.S.; Jett, B.D.; Booth, J.L. Transfer of Pheromone-Inducible Plasmids between Enterococcus-Faecalis in The Syrian-Hamster Gastrointestinal-Tract. J. Infect. Dis. 1992, 166, 1188–1191. [Google Scholar] [CrossRef]
- Huycke, M.M.; Joyce, W.; Wack, M.F. Augmented production of extracellular superoxide by blood isolates of Enterococcus faecalis. J. Infect. Dis. 1996, 173, 743–746. [Google Scholar] [CrossRef]
- Huycke, M.M.; Moore, D.; Joyce, W.; Wise, P.; Shepard, L.; Kotake, Y.; Gilmore, M.S. Extracellular superoxide production by Enterococcus faecalis requires demethylmenaquinone and is attenuated by functional terminal quinol oxidases. Mol. Microbiol. 2001, 42, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Huycke, M.; Moore, D. In vivo production of hydroxyl radical (OH) by Enterococcus faecalis (Ef) colonizing the intestinal tract. Free Radic. Biol. Med. 2001, 31, S144. [Google Scholar]
- Baskaran, S.A.; Kazmer, G.W.; Hinckley, L.; Andrew, S.M.; Venkitanarayanan, K. Antibacterial effect of plant-derived antimicrobials on major bacterial mastitis pathogens in vitro. J. Dairy Sci. 2009, 92, 1423–1429. [Google Scholar] [CrossRef] [PubMed]
- Didehdar, M.; Chegini, Z.; Tabaeian, S.P.; Razavi, S.; Shariati, A. Cinnamomum: The New Therapeutic Agents for Inhibition of Bacterial and Fungal Biofilm-Associated Infection. Front. Cell. Infect. Microbiol. 2022, 12, 930624. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, S.; Cho, K.-H.; Lee, J.-H.; Lee, J. Antibiofilm Activities of Cinnamaldehyde Analogs against Uropathogenic Escherichia coli and Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 7225. [Google Scholar] [CrossRef]
- Foss, M.H.; Eun, Y.J.; Grove, C.I.; Pauw, D.A.; Sorto, N.A.; Rensvold, J.W.; Pagliarini, D.J.; Shaw, J.T.; Weibel, D.B. Inhibitors of bacterial tubulin target bacterial membranes in vivo. Medchemcomm 2013, 4, 112–119. [Google Scholar] [CrossRef]
- Liu, X.; Liu, R.; Zhao, R.; Wang, J.; Cheng, Y.; Liu, Q.; Wang, Y.; Yang, S. Synergistic Interaction Between Paired Combinations of Natural Antimicrobials Against Poultry-Borne Pathogens. Front. Microbiol. 2022, 13, 811784. [Google Scholar] [CrossRef]
- Montagu, A.; Joly-Guillou, M.-L.; Rossines, E.; Cayon, J.; Kempf, M.; Saulnier, P. Stress Conditions Induced by Carvacrol and Cinnamaldehyde on Acinetobacter baumannii. Front. Microbiol. 2016, 7, 1133. [Google Scholar] [CrossRef]
- Vasconcelos, N.G.; Queiroz, J.; da Silva, K.E.; Vasconcelos, P.C.D.; Croda, J.; Simionatto, S. Synergistic effects of Cinnamomum cassia L. essential oil in combination with polymyxin B against carbapenemase-producing Klebsiella pneumoniae and Serratia marcescens. PLoS ONE 2020, 15, e0236505. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Nannapaneni, R.; Kiess, A.; Schilling, W.; Sharma, C.S. Reduction of Salmonella on Turkey Breast Cutlets by Plant-Derived Compounds. Foodborne Pathog. Dis. 2014, 11, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Wendakoon, C.N.; Sakaguchi, M. Inhibition of Amino-Acid Decarboxylase Activity of Enterobacter-Aerogenes By Active Components in Spices. J. Food Prot. 1995, 58, 280–283. [Google Scholar] [CrossRef]
- Wang, N.Y.; Luo, J.; Deng, F.; Huang, Y.S.; Zhou, H. Antibiotic Combination Therapy: A Strategy to Overcome Bacterial Resistance to Aminoglycoside Antibiotics. Front. Pharmacol. 2022, 13, 839808. [Google Scholar] [CrossRef]
- Cava-Roda, R.; Taboada-Rodriguez, A.; Lopez-Gomez, A.; Martinez-Hernandez, G.B.; Marin-Iniesta, F. Synergistic Antimicrobial Activities of Combinations of Vanillin and Essential Oils of Cinnamon Bark, Cinnamon Leaves, and Cloves. Foods 2021, 10, 1406. [Google Scholar] [CrossRef] [PubMed]
- El Atki, Y.; Aouam, I.; El Kamari, F.; Taroq, A.; Nayme, K.; Timinouni, M.; Lyoussi, B.; Abdellaoui, A. Antibacterial activity of cinnamon essential oils and their synergistic potential with antibiotics. J. Adv. Pharm. Technol. Res. 2019, 10, 63–67. [Google Scholar] [CrossRef]
- Utchariyakiat, I.; Surassmo, S.; Jaturanpinyo, M.; Khuntayaporn, P.; Chomnawang, M.T. Efficacy of cinnamon bark oil and cinnamaldehyde on anti-multidrug resistant Pseudomonas aeruginosa and the synergistic effects in combination with other antimicrobial agents. BMC Complement. Altern. Med. 2016, 16, 158. [Google Scholar] [CrossRef]
- Karumathil, D.P.; Nair, M.S.; Gaffney, J.; Kollanoor-Johny, A.; Venkitanarayanan, K. Trans-Cinnamaldehyde and Eugenol Increase Acinetobacter baumannii Sensitivity to Beta-Lactam Antibiotics. Front. Microbiol. 2018, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kang, O.H.; Kwon, D.Y. Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2021, 22, 2752. [Google Scholar] [CrossRef]
- Masi, M.; Refregiers, M.; Pos, K.M.; Pages, J.M. Mechanisms of envelope permeability and antibiotic influx and efflux in Gram-negative bacteria. Nat. Microbiol. 2017, 2, 17001. [Google Scholar] [CrossRef]
- Calvo, J.; Martínez-Martínez, L. Mecanismos de Acción de Los Antimicrobianos; Enfermedades Infecciosas y Microbiología Clínica; Elsevier: Amsterdam, The Netherlands, 2009; Volume 27, pp. 44–52. [Google Scholar]
- Fernandes, R.; Amador, P.; Prudencio, C. Beta-Lactams: Chemical structure, mode of action and mechanisms of resistance. Rev. Med. Microbiol. 2013, 24, 7–17. [Google Scholar] [CrossRef]
- Subhadra, B.; Kim, D.H.; Woo, K.; Surendran, S.; Choi, C.H. Control of Biofilm Formation in Healthcare: Recent Advances Exploiting Quorum-Sensing Interference Strategies and Multidrug Efflux Pump Inhibitors. Materials 2018, 11, 1676. [Google Scholar] [CrossRef]
- Chadha, J.; Singh, J.; Chhibber, S.; Harjai, K. Gentamicin Augments the Quorum Quenching Potential of Cinnamaldehyde In Vitro and Protects Caenorhabditis elegans From Pseudomonas aeruginosa Infection. Front. Cell. Infect. Microbiol. 2022, 12, 899566. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Blanchard, J.S. Molecular insights into aminoglycoside action and resistance. Chem. Rev. 2005, 105, 477–497. [Google Scholar] [CrossRef]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of resistance to aminoglycoside antibiotics: Overview and perspectives. Medchemcomm 2016, 7, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Tiwari, M.; Tiwari, V. Efflux pumps in multidrug-resistant Acinetobacter baumannii: Current status and challenges in the discovery of efflux pumps inhibitors. Microb. Pathog. 2021, 152, 104766. [Google Scholar] [CrossRef] [PubMed]
- Visvalingam, J.; Palaniappan, K.; Holley, R.A. In vitro enhancement of antibiotic susceptibility of drug resistant Escherichia coli by cinnamaldehyde. Food Control 2017, 79, 288–291. [Google Scholar] [CrossRef]
- Gaynor, M.; Mankin, A.S. Macrolide antibiotics: Binding site, mechanism of action, resistance. Curr. Top. Med. Chem. 2003, 3, 949–960. [Google Scholar] [CrossRef]
- Prajapati, J.D.; Kleinekathofer, U.; Winterhalter, M. How to Enter a Bacterium: Bacterial Porins and the Permeation of Antibiotics. Chem. Rev. 2021, 121, 5158–5192. [Google Scholar] [CrossRef]
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef]
- Tetard, A.; Zedet, A.; Girard, C.; Plesiat, P.; Llanes, C. Cinnamaldehyde Induces Expression of Efflux Pumps and Multidrug Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e01081-19. [Google Scholar] [CrossRef]
- Oselusi, S.O.; Christoffels, A.; Egieyeh, S.A. Cheminformatic Characterization of Natural Antimicrobial Products for the Development of New Lead Compounds. Molecules 2021, 26, 3970. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 637511, Cinnamaldehyde. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Cinnamaldehyde (accessed on 6 June 2023).
- Base, H.M. Cinnamaldehyde. Available online: https://hmdb.ca/metabolites/HMDB0003441#references (accessed on 10 October 2023).
- Panda, L.; Duarte-Sierra, A. Recent Advancements in Enhancing Antimicrobial Activity of Plant-Derived Polyphenols by Biochemical Means. Horticulturae 2022, 8, 401. [Google Scholar] [CrossRef]
- Rolfe, M.D.; Rice, C.J.; Lucchini, S.; Pin, C.; Thompson, A.; Cameron, A.D.S.; Alston, M.; Stringer, M.F.; Betts, R.P.; Baranyi, J.; et al. Lag Phase Is a Distinct Growth Phase That Prepares Bacteria for Exponential Growth and Involves Transient Metal Accumulation. J. Bacteriol. 2012, 194, 686–701. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of gram-positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Diekema, D.J.; Messer, S.A.; Pfaller, M.A.; Klepser, M.E. Comparison of Etest, chequerboard dilution and time-kill studies for the detection of synergy or antagonism between antifungal agents tested against Candida species. J. Antimicrob. Chemother. 2002, 49, 345–351. [Google Scholar] [CrossRef]
- Rand, K.H.; Houck, H.J.; Brown, P.; Bennett, D. Reproducibility of the Microdilution Checkerboard Method for Antibiotic Synergy. Antimicrob. Agents Chemother. 1993, 37, 613–615. [Google Scholar] [CrossRef]
- Bonapace, C.R.; White, R.L.; Friedrich, L.V.; Bosso, J.A. Evaluation of antibiotic synergy against Acinetobacter baumannii: A comparison with Etest, time-kill, and checkerboard methods. Diagn. Microbiol. Infect. Dis. 2000, 38, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.H.; Yu, C.M.; Yu, V.L.; Chow, J.W. Synergy Assessed by Checkerboard-A Critical Analysis. Diagn. Microbiol. Infect. Dis. 1993, 16, 343–349. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.J.; Middleton, R.F.; Westmacott, D. The Fractional Inhibitory Concentration (Fic) Index as a Measure of Synergy. J. Antimicrob. Chemother. 1983, 11, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Siriwong, S.; Teethaisong, Y.; Thumanu, K.; Dunkhunthod, B.; Eumkeb, G. The synergy and mode of action of quercetin plus amoxicillin against amoxicillin-resistant Staphylococcus epidermidis. BMC Pharmacol. Toxicol. 2016, 17, 39. [Google Scholar] [CrossRef] [PubMed]
- Gan, C.; Langa, E.; Valenzuela, A.; Ballestero, D.; Pino-Otín, M.R. Synergistic Activity of Thymol with Commercial Antibiotics against Critical and High WHO Priority Pathogenic Bacteria. Plants 2023, 12, 1868. [Google Scholar] [CrossRef] [PubMed]
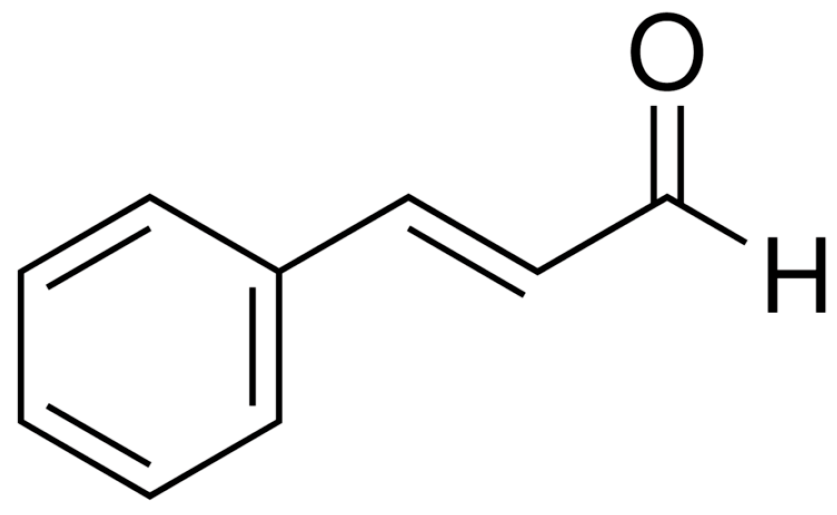

{kind=link}
| MICalone (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Bacteria | AMP | AMO | STM | GTM | ERY | TC | CHL | CIN | |
| Bacillus subtilis (ATCC 6633) | Exp. | 0.3 | 0.3 | 8 | 8 | <0.5 | 5 | 1.5 | 500 |
| Lit. | 1 f | 1.56 j | 1.562 n | 1.56 j | |||||
| Enterococcus faecalis (ATCC 19433) | Exp. | 500 | 500 | 250 | 25 | 100 | 12.5 | 125 | 1000 |
| Lit. | |||||||||
| Listeria monocytogenes (ATCC 7644) | Exp. | 125 | 62.5 | 200 | 12.5 | 125 | 12.5 | 10 | 500 |
| Lit. | 64 o | ||||||||
| Streptococcus agalactiae (ATCC 12386) | Exp. | 62.5 | 62.5 | 100 | 18.75 | 125 | 0.19 | 50 | 500 |
| Lit. | |||||||||
| Staphylococcus aureus (ATCC 9144) | Exp. | 0.15 | 0.625 | 100 | 25 | 0.62 | 12.5 | 30 | 500 |
| Lit. | |||||||||
| Acinetobacter baumannii (ATCC 19606) | Exp. | 125 | 62.5 | 75 | 25 | 10 | 0.75 | 62.5 | 500 |
| Lit. | 64 g | 256 k | 16 g | 256 p | |||||
| Escherichia coli (ATCC 25922) | Exp. | 7.5 | 7.5 | 15.65 | 15.65 | 62.5 | 0.75 | 7.5 | 500 |
| Lit. | 7.5 e | 2–8 h | 32 l | 64 m | 32 a | 7.5 e | 256 d | ||
| 157 b | |||||||||
| Klebsiella aerogenes (ATCC 13048) | Exp. | 1500 | 1000 | 4.5 | 2.5 | 31.25 | 1.9 | 10 | 500 |
| Lit. | 8 q | ||||||||
| Klebsiella pneumoniae (C6) | Exp. | 500 | 125 | 10 | 0.78 | 62.5 | 1.17 | 15 | 500 |
| Lit. | |||||||||
| Pasteurella aerogenes (ATCC 27883) | Exp. | 125 | 500 | 15.6 | 6.25 | 62.5 | 10 | 8 | 500 |
| Lit. | |||||||||
| Pseudomonas aeruginosa (ATCC 27853) | Exp. | 1000 | >1000 | 62.5 | 12.5 | 500 | 25 | 250 | 1000 |
| Lit. | >64 b | 0.12 i | 12.5 j | 0.5–2 e | 8–32 e | 100 r | 1000 c | ||
| 630 b | |||||||||
| Salmonella enterica (ATCC 13311) | Exp. | <0.15 | <0.15 | 50 | 0.75 | 31.25 | 1.17 | 3.9 | 500 |
| Lit. | |||||||||
| Serratia marcescens (ATCC 13880) | Exp. | 125 | 125 | 0.4 | 5 | 250 | 250 | 125 | 500 |
| Lit. | 6.25 s | ||||||||
| Proteus mirabilis (ATCC 35659) | Exp. | 0.08 | 0.62 | 12.5 | 25 | <0.08 | 0.08 | 7.8 | - |
| Lit. | |||||||||
| Gram+/− | Bacteria | Compounds | MICcomb (mg/mL) | FIC | ΣFIC | Activity | MIC Reduction (%) |
|---|---|---|---|---|---|---|---|
| GRAM+ | E. faecalis | CIN | 250 | 0.25 | 0.5 | Synergy | 75 |
| STM | 62.5 | 0.25 | 75 | ||||
| CIN | 250 | 0.25 | 0.37 | 75 | |||
| GTM | 3 | 0.12 | 88 | ||||
| CIN | 500 | 0.5 | 0.75 | Addition | 50 | ||
| AMP | 125 | 0.25 | 75 | ||||
| CIN | 500 | 0.5 | 0.75 | 50 | |||
| AMO | 125 | 0.25 | 75 | ||||
| CIN | 500 | 0.5 | 1.5 | 50 | |||
| ERY | 100 | 1 | 0 | ||||
| CIN | 500 | 0.5 | 2.5 | 50 | |||
| TC | 25 | 2 | −100 | ||||
| CIN | 500 | 0.5 | 4.5 | Antagonism | 50 | ||
| CHL | 500 | 4 | −300 | ||||
| L. monocytogenes | CIN | 125 | 0.25 | 0.5 | Synergy | 75 | |
| AMP | 31.25 | 0.25 | 75 | ||||
| CIN | 125 | 0.25 | 0.5 | 75 | |||
| STM | 50 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 2.5 | Addition | 50 | ||
| AMO | 125 | 2 | −100 | ||||
| CIN | 250 | 0.5 | 0.62 | 50 | |||
| GTM | 1.56 | 0.12 | 88 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| ERY | 62.5 | 0.5 | 50 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| TC | 6.25 | 0.5 | 50 | ||||
| CIN | 250 | 0.5 | 8.5 | Antagonism | 50 | ||
| CHL | 62.5 | 8 | −525 | ||||
| S. agalactiae | CIN | 125 | 0.25 | 0.5 | Synergy | 75 | |
| STM | 25 | 0.25 | 75 | ||||
| CIN | 125 | 0.12 | 0.37 | 88 | |||
| GTM | 4.6 | 0.25 | 75 | ||||
| CIN | 31.5 | 0.25 | 0.5 | 75 | |||
| CHL | 12.5 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 1 | Addition | 50 | ||
| AMP | 31.25 | 0.5 | 50 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| AMO | 31.25 | 0.5 | 50 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| ERY | 62.5 | 0.5 | 50 | ||||
| S. aureus | CIN | 125 | 0.25 | 0.31 | Synergy | 75 | |
| STM | 6.25 | 0.06 | 94 | ||||
| CIN | 31.25 | 0.06 | 0.19 | 94 | |||
| GTM | 3 | 0.13 | 88 | ||||
| CIN | 250 | 0.5 | 1.5 | Addition | 50 | ||
| TC | 12.5 | 1 | 0 | ||||
| CIN | 500 | 1 | >3 | 0 | |||
| CHL | >60 | 2 | <−100 | ||||
| GRAM− | A. baumannii | CIN | 125 | 0.5 | 0.75 | Addition | 75 |
| AMP | 62.5 | 0.25 | 50 | ||||
| CIN | 500 | 1 | 2 | 0 | |||
| AMO | 62.5 | 1 | 0 | ||||
| CIN | 250 | 0.5 | 1.5 | 50 | |||
| ERY | 10 | 1 | 0 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| CHL | 31.25 | 0.5 | 50 | ||||
| CIN | 125 | 0.25 | >2.25 | 75 | |||
| GTM | >50 | 2 | <−100 | ||||
| CIN | 250 | 0.5 | 4.5 | Antagonism | 50 | ||
| STM | 300 | 4 | −300 | ||||
| E. coli | CIN | 250 | 0.5 | 0.75 | Addition | 50 | |
| STM | 3.9 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 0.75 | 50 | |||
| GTM | 3.9 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 1.5 | 50 | |||
| ERY | 62.5 | 1 | 0 | ||||
| K. aerogenes | CIN | 250 | 0.5 | 0.62 | Addition | 50 | |
| AMP | 187.5 | 0.12 | 87 | ||||
| CIN | 250 | 0.5 | 0.62 | 50 | |||
| AMO | 125 | 0.12 | 87 | ||||
| CIN | 250 | 0.5 | 2.5 | 50 | |||
| ERY | 62.5 | 2 | −100 | ||||
| CIN | 250 | 0.5 | 8.5 | Antagonism | 50 | ||
| CHL | 80 | 8 | −700 | ||||
| K. pneumoniae | CIN | 125 | 0.25 | 0.5 | Synergy | 75 | |
| AMP | 125 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 2.5 | Addition | 50 | ||
| AMO | 250 | 2 | −100 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| STM | 5 | 0.5 | 50 | ||||
| CIN | 125 | 0.25 | 2.5 | 75 | |||
| ERY | 125 | 2 | −100 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| CHL | 7.5 | 0.5 | 50 | ||||
| P. aerogenes | CIN | 125 | 0.25 | 0.5 | Synergy | 75 | |
| AMP | 31.25 | 0.25 | 75 | ||||
| CIN | 62.5 | 0.13 | 0.19 | 87 | |||
| AMO | 31.25 | 0.06 | 94 | ||||
| CIN | 250 | 0.5 | 1.5 | Addition | 50 | ||
| STM | 15.6 | 1 | 0 | ||||
| CIN | 250 | 0.5 | 1.5 | 50 | |||
| ERY | 62.5 | 1 | 0 | ||||
| CIN | 250 | 0.5 | 1.5 | 50 | |||
| TC | 10 | 1 | 0 | ||||
| P. aeruginosa | CIN | 500 | 0.5 | 0.62 | Addition | 50 | |
| AMP | 125 | 0.12 | 87 | ||||
| CIN | 250 | 0.25 | 0.75 | 75 | |||
| STM | 31.25 | 0.5 | 50 | ||||
| CIN | 250 | 0.25 | 1.5 | 75 | |||
| GTM | 12.5 | 1 | 0 | ||||
| CIN | 500 | 0.5 | 1.5 | 50 | |||
| ERY | 500 | 1 | 0 | ||||
| CIN | 500 | 0.5 | 1.5 | 50 | |||
| TC | 25 | 1 | 0 | ||||
| CIN | 500 | 0.5 | 4.5 | Antagonism | 50 | ||
| CHL | 1 | 4 | −300 | ||||
| S. enterica | CIN | 62.5 | 0.12 | 0.37 | Synergy | 87 | |
| STM | 12.5 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 0.56 | Addition | 50 | ||
| ERY | 1.9 | 0.06 | 94 | ||||
| S. marcescens | CIN | 125 | 0.25 | 0.26 | Synergy | 75 | |
| CHL | 1.9 | 0.01 | 98 | ||||
| CIN | 125 | 0.25 | 0.5 | 75 | |||
| ERY | 62.5 | 0.25 | 75 | ||||
| CIN | 250 | 0.5 | 1 | Addition | 50 | ||
| AMP | 62.5 | 0.5 | 50 | ||||
| CIN | 250 | 0.5 | 1 | 50 | |||
| AMO | 62.5 | 0.5 | 50 | ||||
| CIN | 250 | 0.5 | 0.62 | 50 | |||
| TC | 31.25 | 0.12 | 87 |
| Antibiotic | CAS | Provider | Purity/% |
|---|---|---|---|
| Ampicillin | 69-53-4 | Sigma Aldrich (St. Louis, MO, USA) | >96 |
| Amoxicillin | 26787-78-0 | ||
| Gentamycin (sulphate) | 1405-41-0 | Acofarma (Barcelona, Spain) | |
| Streptomycin (sulphate) | 3810-74-0 | ||
| Erythromycin | 114-07-8 | 97.5 | |
| Tetracycline (clorhidrate) | 64-75-5 | 99.2 | |
| Chloramphenicol | 56-75-7 | 99.6 | |
| Trans-cinnamaldehyde | 104-55-2 | Sigma Aldrich | >95 |
| DMSO | 67-68-5 | Fisher Bioreagents (Madrid, Spain) | >99.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrando, N.; Pino-Otín, M.R.; Ballestero, D.; Lorca, G.; Terrado, E.M.; Langa, E. Enhancing Commercial Antibiotics with Trans-Cinnamaldehyde in Gram-Positive and Gram-Negative Bacteria: An In Vitro Approach. Plants 2024, 13, 192. https://doi.org/10.3390/plants13020192
Ferrando N, Pino-Otín MR, Ballestero D, Lorca G, Terrado EM, Langa E. Enhancing Commercial Antibiotics with Trans-Cinnamaldehyde in Gram-Positive and Gram-Negative Bacteria: An In Vitro Approach. Plants. 2024; 13(2):192. https://doi.org/10.3390/plants13020192
Chicago/Turabian StyleFerrando, Natalia, María Rosa Pino-Otín, Diego Ballestero, Guillermo Lorca, Eva María Terrado, and Elisa Langa. 2024. "Enhancing Commercial Antibiotics with Trans-Cinnamaldehyde in Gram-Positive and Gram-Negative Bacteria: An In Vitro Approach" Plants 13, no. 2: 192. https://doi.org/10.3390/plants13020192
APA StyleFerrando, N., Pino-Otín, M. R., Ballestero, D., Lorca, G., Terrado, E. M., & Langa, E. (2024). Enhancing Commercial Antibiotics with Trans-Cinnamaldehyde in Gram-Positive and Gram-Negative Bacteria: An In Vitro Approach. Plants, 13(2), 192. https://doi.org/10.3390/plants13020192