Abstract
Dicranum Hedw. is a highly diverse and widely distributed genus within Dicranaceae. The species diversity and distribution of this genus in China, however, remain not well known. A new revision of Dicranum in China using morphological and molecular phylogenetic methods confirms that China has 39 species, including four newly reported species, D. bardunovii Tubanova & Ignatova, D. dispersum Engelmark, D. schljakovii Ignatova & Tubanova, and D. spadiceum J.E.Zetterst. Dicranum psathyrum Klazenga is transferred to Dicranoloma (Renauld) Renauld as a new synonym of Dicranoloma fragile Broth. Two species, Dicranum brevifolium (Lindb.) Lindb. and D. viride (Sull. & Lesq.) Lindb. are excluded from the bryoflora of China. A key to the Chinese Dicranum species is also provided. These results indicate an underestimation of the distribution range of numerous Dicranum species, underscoring the need for further in-depth investigations into the worldwide Dicranum diversity.
1. Introduction
Dicranaceae are a family of mosses (Bryophyta). Dicranum Hedw. (including Orthodicranum (Bruch & Schimp.) Loeske) stands out as one of the most diverse genera within Dicranaceae, comprising approximately 98 accepted species [1]. The taxonomy of this genus presents significant challenges owing to the considerable variation of gametophytic characteristics influenced by environmental conditions [2,3,4,5]. Recent advancements in molecular phylogeny have led to a more precise delineation of Dicranum, including the identification of distinct groups such as Dicranum species with fragile leaves [6,7], D. fuscescens Turner complexes [8,9], D. scoparium Hedw. complexes [4,10], arctic Dicranum species [11], D. acutifolium (Lindb. & Arnell) C.E.O.Jensen complexes [12,13], and European Dicranum species [14]. Despite these valuable insights, it is essential to recognize the geographical limitations of these studies, which have predominantly focused on Europe and Asian Russia, thus restricting our comprehensive understanding of Dicranum species diversity in other regions worldwide. Particularly in China, the delineation and relationships among morphological species of Dicranum remain challenging to ascertain.
In 1890–1895, Italian missionary Giuseppe Giraldi collected numerous bryophyte specimens in Shaanxi Province, China. These specimens were studied by Carl Müller, and the findings were published in the work titled Bryologia provinciae Schen-si Sinensis I-III [15,16,17]. The discovery of Dicranum rectifolium Müll.Hal. and D. drummondii Müll.Hal. (reported as D. thelinotum Müll.Hal.) marked the first known presence of Dicranum in China in 1896 [15]. Subsequently, additional researchers such as Salmon [18], Paris [19], Brotherus [20,21,22], Reimers [23], and Bartram [24] documented various Dicranum species in China (Table 1). By the end of World War II, a total of sixteen Dicranum species had been recorded in China. Significantly, three of these, including D. rectifolium [15], D. papillidens Broth. [20], and D. cheoi E.B.Bartram [24], were newly described to science. Regrettably, no Chinese botanists had contributed to the early reports on Dicranum species in China.
After World War II, a research group led by Pan-Chieh Chen, a pioneer in Chinese modern bryology [25], embarked on exploring the Chinese bryoflora. The results of these investigations were published in various Chinese floras, offering detailed descriptions and illustrations of Dicranum species. The earliest regional moss flora study was completed in 1963 [26], documenting seven previously undocumented Dicranum species in China: D. fulvum Hook., D. elongatum Schleich. ex Schwägr., D. groenlandicum Brid., D. fragilifolium Lindb., D. fuscescens Turner, D. undulatum Schrad. ex Brid., and D. spurium Hedw. Subsequently, several species were reported new to China by other Chinese bryologists [27,28,29,30,31,32], resulting in a total of 43 recorded Dicranum species listed in the Moss Flora of China [33].
Notably, Gao et al. [34] raised Dicranum subgenus Pseudochorisodontium Broth. as a new genus, Pseudochorisodontium (Broth.) C.Gao, and recent studies [3,7,14,35,36,37,38] have embraced a broader concept of this genus, merging Orthodicranum into Dicranum. Following these treatments, 34 Dicranum species have been cataloged in the Species Catalogue of China [35,36]. Nevertheless, there are still numerous omissions in this checklist that necessitate further study and updating. For example, more recently, Huang et al. [7,38] described two new species and a newly recorded species in China: D. baicalense Tubanova, D. hengduanense W.Z.Huang & R.L.Zhu, and D. shennongjiaense W.Z.Huang & R.L.Zhu (Table 1).

Table 1.
A list in chronological order of the Dicranum species described and recorded from China. Names in bold indicate being newly reported to science based on the specimens collected from China. Dicranum psathyrum (≡Dicranoloma fragile Klazenga) is not included.
Table 1.
A list in chronological order of the Dicranum species described and recorded from China. Names in bold indicate being newly reported to science based on the specimens collected from China. Dicranum psathyrum (≡Dicranoloma fragile Klazenga) is not included.
| No. | Species | Year | Reference and Notes |
|---|---|---|---|
| 1 | Dicranum rectifolium | 1896 | Müller [15] |
| 2 | Dicranum drummondii | 1896 | Müller [16], reported as Dicranum thelinotum |
| 3 | Dicranum japonicum | 1897 | Müller [17], reported as Dicranum schensianum |
| 4 | Dicranum montanum | 1898 | Müller [17] |
| 5 | Dicranum hamulosum | 1900 | Salmon [18], reported as Dicranum crispofalcatum |
| 6 | Dicranum lorifolium | 1900 | Salmon [18] |
| 7 | Dicranum nipponense | 1908 | Paris [19], reported as Dicranum rufescens |
| 8 | Dicranum papillidens | 1924 | Brotherus [20] |
| 9 | Dicranum scoparium | 1929 | Brotherus [21], reported as Dicranum orthophyllum |
| 10 | Dicranum muehlenbeckii | 1929 | Brotherus [21] |
| 11 | Dicranum crispifolium | 1929 | Brotherus [21] |
| 12 | Dicranum kashmirense | 1929 | Brotherus [21] |
| 13 | Dicranum majus | 1929 | Brotherus [21] |
| 14 | Dicranum mayrii | 1929 | Brotherus [22], reported as Dicranum formosicum |
| 15 | Dicranum flagellare | 1931 | Reimers [23] |
| 16 | Dicranum cheoi | 1935 | Bartram [24] |
| 17 | Dicranum fulvum | 1963 | Chen [26] |
| 18 | Dicranum elongatum | 1963 | Chen [26] |
| 19 | Dicranum groenlandicum | 1963 | Chen [26] |
| 20 | Dicranum fragilifolium | 1963 | Chen [26] |
| 21 | Dicranum fuscescens | 1963 | Chen [26] |
| 22 | Dicranum undulatum | 1963 | Chen [26] |
| 23 | Dicranum spurium | 1963 | Chen [26] |
| 24 | Dicranum bonjeanii | 1963 | Li [27] |
| 25 | Dicranum polysetum | 1977 | Gao [28] |
| 26 | Dicranum scottianum | 1977 | Gao [28] |
| 27 | Dicranum linzianum | 1979 | Gao and Chang [29] |
| 28 | Dicranum himalayanum | 1985 | Li [30] |
| 29 | Dicranum assimicum | 1985 | Li [30] |
| 30 | Dicranum setifolium | 1992 | Gao and Cao [31] |
| 31 | Dicranum leiodontum | 1992 | Gao and Cao [31] |
| 32 | Dicranum hakkodense | 2008 | Ignatova and Fedosov [6] |
| 33 | Dicranum hengduanense | 2023 | Huang et al. [7] |
| 34 | Dicranum shennongjiaense | 2024 | Huang et al. [38] |
| 35 | Dicranm baicalense | 2024 | Huang et al. [38] |
| 36 | Dicranum bardunovii | — | Present study |
| 37 | Dicranum dispersum | — | Present study |
| 38 | Dicrnaum schljakovii | — | Present study |
| 39 | Dicranum spadiceum | — | Present study |
To advance our understanding of Dicranum diversity in China, we conducted a taxonomic revision of Dicranum in China over the last two years [7,38]. Throughout this period, we encountered some intriguing specimens that did not correspond to any previously documented Dicranum species in China. To clarify the taxonomic classification of these Chinese mosses, we conducted a phylogenetic analysis by sequencing five chloroplast markers (rpoB, rps4-trnT, rps19-rpl2, trnH-psbA, and trnL-trnF) and one nuclear marker (ITS region). Through a meticulous examination of morphological characteristics and molecular phylogeny analysis, we confirmed the existence of four new records of Dicranum species in China, including D. bardunovii, D. dispersum, D. schljakovii, and D. spadiceum.
The primary objectives of this study are (1) to confirm the species diversity of Dicranum in China based on literature sources and our findings, (2) to provide photographs and descriptions of four new national records of Dicranum species, (3) to delineate the distribution of each Dicranum species within China and elucidate the differences between each species and those commonly confused species, and (4) to provide an updated key to Dicranum species in China.
2. Results
2.1. Results of Phylogenetic Analysis
The aligned six-loci dataset comprised 2692 characters, consisting of the following segments and lengths: nrITS1-5.8S-ITS2 (784 bp), rps19-rpl2 (352 bp), rpoB (457 bp), rps4-trnT (510 bp), trnH-psbA (139 bp), and trnL-trnF (450 bp). Of the 2692 aligned nucleotides in the 309 accessions analyzed, 1662 were constant sites, 400 were singleton sites, and 630 were parsimony-informative. Maximum likelihood (ML) and Bayesian inference (BI) analyses produced congruent trees with robust support for the majority of nodes. The ML tree topology with bootstrap values (BSML and PPBI) is shown in Figure 1.
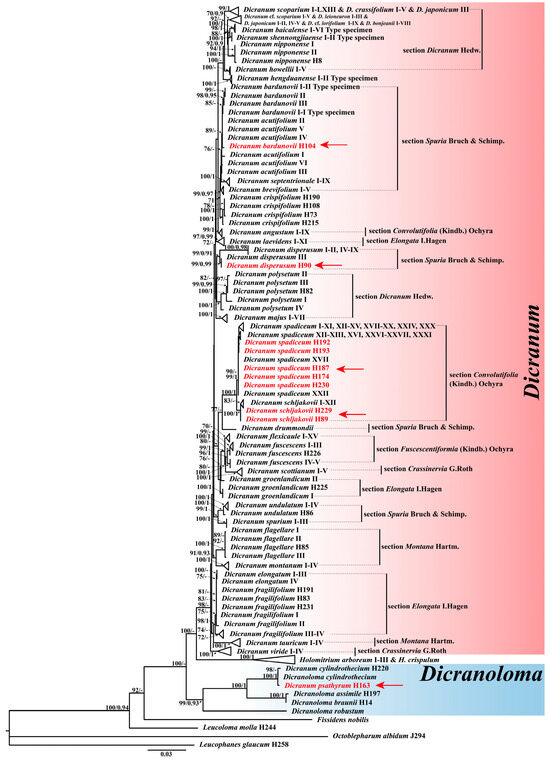
Figure 1.
Phylogeny of Dicranum species inferred from the combined dataset (trnH-psbA, rps4-trnT, trnL-trnF, rps19-rpl2, rpoB, and ITS). The topology derived from the best-scoring ML tree in IQtree is shown. ML bootstrap values BS ≥ 70 are shown on the left and Bayesian posterior probabilities values PP ≥ 0.90 on the right. The subject species are in red and indicated by arrows. Section division refers to Hodgetts et al. [37].
The phylogeny positions of Dicranum samples collected in China are as follows (Figure 1): (1) D. bardunovii, D. angustum, D. acutifolium, D. brevifolium, D. septentrionale, and D. crispifolium constitute a weakly supported clade. Within this clade, nine accessions of D. angustum (BSML = 99, PPBI = 1), five accessions of D. brevifolium (BSML = 99, PPBI = 0.97), four accessions of D. crispifolium (BSML = 100, PPBI = 1), and nine accessions of D. septentrionale (BSML = 99, PPBI = 0.97) form highly supported subclades, respectively. However, the accession of D. bardunovii from China is nested deeply within D. acutifolium. (2) Ten accessions of D. dispersum form a strongly supported clade (BSML = 99, PPBI = 0.91). Within this clade, one accession from China and one accession from Russia form a single subclade with high support values (BSML = 99, PPBI = 0.99). (3) Two accessions of D. schljakovii collected from China form a well-supported clade with twelve other accessions (BSML = 100, PPBI = 1). (4) Five accessions of D. spadiceum collected from China (Qinghai and Xinjiang) are nested deeply within the other accessions from Europe and Russia (BSML = 100, PPBI = 0.99). (5) The accession of D. psathyrum is deeply nested within the genus Dicranoloma (Renauld) Renauld (BSML = 99, PPBI = 0.93).
Based on the phylogenetic analysis, we conclusively confirm the newly recorded distribution of Dicranum dispersum, D. schljakovii, and D. spadiceum in China, and propose transferring D. psathyrum to the genus Dicranoloma. In addition, the tree topology allows for considering the accession of Dicranum bardunovii from China within D. acutifolium (Figure 1). Despite this, significant morphological distinctions exist between these two species (see note under D. bardunovii for details), supporting that our collection in China represents D. bardunovii rather than D. acutifolium.
2.2. Newly Recorded Species for China

Figure 2.
Plants of Dicranum species new to China. (A,B) D. bardunovii Tubanova & Ignatova, from R.L. Zhu et al. 20220830-30B (HSNU); (C,D) D. dispersum Engelmark, from S.B. Zhang 20220709-41 (HSNU); (E,F) D. schljakovii Ignatova & Tubanova, from R.L. Zhu et al. 20220803-314A (HSNU); (G,H) D. spadiceum J.E. Zetterst., from L. Shu & W.Z. Huang 20220817-38 (HSNU).

Figure 3.
Dicranum bardunovii Tubanova & Ignatova. All from R.L. Zhu et al. 20220830-30B (HSNU). (A) Leaves, C*, D*, E*, F* indicates the position of C, D, E, F on the leaf, respectively; (B) Leaf apex; (C) Upper laminal cells of leaf; (D) Middle laminal cells of leaf; (E) Basal lamianl cells of leaf; (F) Alar cells of leaf; (G) Costa surface and laminal cells in the upper portion of leaf (in dorsal view), arrows shows papillose; (H) cross-sections of leaf; (I) cross-section of stem.
Distribution in China: Yunnan (present report), new to China.
Specimens examined: China. Yunnan. Diqingzangzu Autonomous Prefecture, Deqin country, Baimaxueshan National Nature Reserve, at Baimaxueshan Pass, 28°22′52.68″ N, 99°1′12.03″ E, 4365 m, on soil near the base of azalea, 30 August 2022, R.L. Zhu et al. 20220830-30B (HSNU).
Description: Plants robust, in loose tufts, green or yellow-green in the upper part, brownish below. Stems 5–8 cm, matted with white tomentum in the upper part, rusty tomentum and rhizoids at the base. Cross-section of stem is rounded, consisting of 1–2 layers of smaller, obviously thick-walled outer cortical cells, numerous rows of larger and thinner-walled inner cortical cells, and 6–9 rows of central strand cells. Leaves straight and erect-spreading when wet, slightly curved when dry, 5.8–7.5 × 0.75–0.86 mm, lanceolate, from ovate base gradually acuminate. Leaf margins are serrated in the upper 3/5, strongly serrated in the distal part, and entirely smooth below. Costa ca. 200 µm wide at the leaf base, about 1/5 of leaf width at the base, the dorsal side of the costa is sharply and densely scabrose in the upper part, with one row of guide cells in the transverse section, two stereid bands, ventral epidermis not differentiated, and well differentiated dorsal epidermis. Leaf lamina is unistratose, with bistratose regions on or near the upper margins. Upper and median laminal cells are 7–18 × 7–9 µm, quadrate, irregularly angled, or short rectangular, a lot of which are strongly prorulose on the upper ends on the dorsal side of the leaf. Basal laminal cells are elongated rectangular to linear, 43–100 × 7–10 µm, and pitted. Alar cells are differentiated, brownish, decurrent, and not extending to the costa, with a group of thin-walled hyaline cells between the alar cells and costa, with 2–3 layers of cells in the transverse section. Sporophyte not seen.
Note: This species was previously known only from Russia (including Asiatic Russia east of Yenissey River and the European part of the Urals) [9,37]. In China, it grows on stone at high altitudes up to 4365 m, usually associated with Brachythecium coarctum (Müll.Hal.) Ignatov & Huttunen, Paraleucobryum enerve (Thed.) Loeske, and Symblepharis vaginata (Hook. ex Harv.) Wijk & Margad.
Dicranum bardunovii is highly similar to D. acutifolium, but can be differentiated by its sharply and densely scabrose laminal cells and costa in the distal part of the leaf, whereas the latter species has smooth cells and costa [9]. Although Tubanova and Ignatova [9], Lang et al. [12], and Kiebacher and Szövényi [13] provided molecular evidence supporting distinctions between the two taxa, the phylogeny trees in their results exhibited weak clade support. Lang et al. [12] also found that one of the two accessions from the holotype specimen of D. bardunovii nested deeply within the clade of D. acutifolium and attributed it to mixed collections of these two species in the holotype specimen. However, the morphological characteristics of the type of specimen do not support their hypothesis (refer to Supplementary Table S2 in Lang et al. [12]). The phylogenetic tree in our analysis showed low branch support in this complex, with D. bardunovii collected from China deeply nested within D. acutifolium (Figure 1). However, morphologically, we are highly confident that the Chinese specimen belongs to D. bardunovii, as evidenced by the sharply and densely scabrose laminal cells and costa in the distal part of the leaf (Figure 3G).
Dicranum bardunovii is well characterized by (1) its ventral stereid band being exposed and surface cells not differentiated (Figure 3H), (2) a transverse section of leaf in the upper part like a pair of tongs (Figure 3H), (3) a sharply and densely scabrose dorsal side of laminal cells and costa in the distal part of the leaf (Figure 3G), (4) strongly serrated margins in the distal part (Figure 3C), and (5) bistratose alar cells (Figure 3H). There are no obvious differences between the morphological characteristics mentioned above from China and the illustrations based on the type of specimen in Tubanova and Ignatova [9].
Dicranum dispersum Engelmark, Stuttgarter Beitr. Naturk., A 592: 4. 1999. (Figure 2C,D and Figure 4)
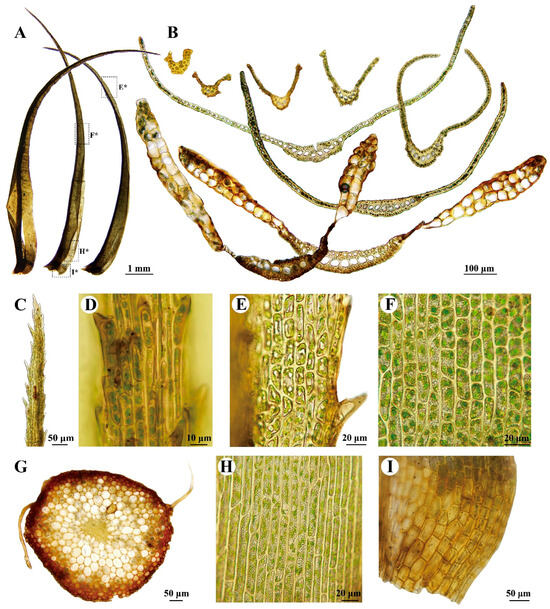
Figure 4.
Dicranum dispersum Engelmark. All from S.B. Zhang 20220709-41 (HSNU). (A) Leaves, E*, F*, H*, I* indicates the position of E, F, H, I on the leaf, respectively; (B) Cross-sections of leaf; (C) Leaf apex; (D) Costa in the upper portion of leaf (dorsal view); (E) Upper laminal cells of leaf; (F) Middle laminal cells of leaf; (G) Cross-section of stem; (H) Basal laminal cells of leaf; (I) Alar cells of leaf.
Distribution in China: Qinghai (present report), new to China.
Specimens examined: China. Qinghai. Tongren City. Zhamao Country, Maixiu area of Sanjiangyuan National Nature Reserve, 35°22′2″ N, 101°50′8″ E, 2942 m, southwest-facing slope 50°, on stone in a valley, 9 July 2022. S.B. Zhang 20220709-41 (HSNU).
Description: Plants rather robust, in loose tufts, green in the upper part, and brownish below. Stems 4–8 cm, matted with white tomentum in the upper part, and rusty tomentum and rhizoids at the base. Cross-section of stem is rounded, consisting of 2–4 layers of smaller, obviously thick-walled outer cortical cells, numerous rows of larger and thinner-walled inner cortical cells, and a number of central strand cells. Leaves somewhat flexuous, erect-spreading when wet, loosely curved when dry, usually keeled in the upper part, and appearing V-shaped or looking like a pair of tongs in the cross-section. Leaves gradually narrowed from an ovate–lanceolate base to a long acuminate apex, 9.5–12 × 1–1.2 mm. Leaf margins are serrated in the upper 3/5 to 1/2, and entirely smooth below. Costa is 1/5–1/4 of leaf width at the base and 200–300 µm wide at the leaf base, with teeth on the dorsal surface in the upper part, with one row of guide cells in the transverse section, two stereid bands, and differentiated dorsal and ventral layers of cells. Leaf laminal is unistratose, with bistratose regions on or near the upper margins. Upper and median laminal cells are 13–25(–34) × 9–11 µm, quadrate, irregularly angled, short rectangular, smooth, or slightly mammillose. Basal laminal cells are elongated–rectangular to linear, 65–120 × 7–12 µm, and slightly pitted or not. Alar cells are differentiated, brownish, extending to the costa, with 2–3(–4) layers of cells in the transverse section. Sporophyte not seen.
Note: Dicranum dispersum is predominantly found in various regions across Europe, (e.g., France, Germany, and Switzerland), Russia, Iran, Mongolia, and North America [39]. In China, it grows on rocky surfaces, associated with Thuidium cymbifolium (Dozy & Molk.) Dozy & Molk.
Dicranum dispersum has been classified as an endangered species in the Red Lists of Germany and France [40] and assessed as endangered under criteria D in The IUCN Red List of Threatened Species [39]. In recent years, new distribution sites of this rare species have been discovered in Europe, Asiatic Russia, and Alaska, U.S.A. [5,41,42], and our study confirms its presence in Qinghai Province, China. To date, the known populations are highly fragmented due to habitat fragmentation, resulting in small, isolated patches that are unable to sustain minimum viable populations and are at risk of entering an extinction vortex [43,44]. Furthermore, moss species, which prefer cold environments, may be more vulnerable to the global warming climate [45,46]. Therefore, we recommend including this rare species on the China Species Red List and advocating for further research into the distribution and biology of high-altitude distributed Dicranum species.
The important features of Dicranum dispersum include the (1) differentiated dorsal and ventral epidermal layers of costal cells (Figure 4B), (2) leaves that are channeled or keeled in the upper part, appearing V-shaped or tong-like in cross-sectional appearance (Figure 4B), (3) the presence of 2–3(–4) layers of alar cells (Figure 4B), (4) laminal cells that are occasionally or not pitted in the lower portion of the leaf (Figure 4H), (5) long to shortly excurrent costa (Figure 4A), and (6) laminal cells that are bistratose at the margins in the upper portion of the leaf (Figure 4B,E). Additional illustrations can refer to Afonina and Breen [41], Engelmark [47], Otnyukova [48], Ignatov and Ignatova [49], and Lüth [50].
This species can be confused with D. muehlenbeckii Bruch & Schimp., D. spadiceum J.E.Zetterst., D. acutifolium, D. fuscescens, D. brevifolium, D. caesium Mitt., and D. crispifolium. The differences between these species have been well discussed by Afonina and Breen [41], Engelmark [47], and Otnyukova and Ochrya [51].
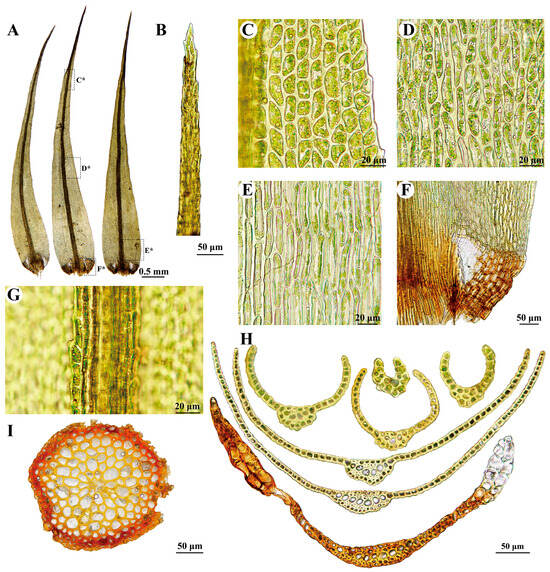
Figure 5.
Dicranum schljakovii Ignatova & Tubanova. All from R.L. Zhu et al. 20220803-314A (HSNU). (A) Leaves, C*, D*, E*, F* indicates the position of C, D, E, F on the leaf, respectively; (B) Leaf apex; (C) Upper lamianl cells of leaf; (D) Middle lamianl cells of leaf; (E) Basal lamianl cells of leaf; (F) Alar cells of leaf; (G) Costa in upper portion of leaf (dorsal view); (H) Cross-sections of leaf; (I) Cross-section of stem.
Distribution in China: inner Mongolia (present report), Xinjiang (present report), new to China.
Specimens examined: China. Inner Mongolia. Great Khingan Range, Genhe City, Alongshan Town, Mount Aokelidui (Mount Xianbei), 51°50′38.95′′ N, 122°2′49.08′′ E, 1447 m, on soil, 3 August 2022, R.L. Zhu et al. 20220803-314 (HSNU); Xinjiang. Altai region, Altai Mt., Dahe tree farm, 47°5′16′′ N, 87°02′24′′ E, 1900 m, on soil, 30 June 1985, M. Sulayman 1820A (XJU); Altai region, Borjin County, Kanas National Nature Reserve, 48°41′17.50′′ N, 87°15′12.87′′ E, 3200 m, on decaying wood, 2 July 1998, M. Sulayman 9861 (XJU); ibid., M. Sulayman 14158 (XJU); ibid., 48°35′28′′ N, 87°52′47.69′′ E, 2321 m, on stone, 27 September 2016, M. Sulayman 29199 (XJU); ibid., 48°41′14.42′′ N, 87°14′17.70′′ E, 2225 m, on soil, 10 July 2016, M. Sulayman 28070 (XJU); Altai region, Altai Mt., Qinghe County, Daqinghe, 46°55′48.46′′ N, 90°23′26.13′′ E, 1860 m, on stone, 16 June 2016, M.Sulayman 28281 (XJU); ibid., Bianhaizi, 46°49′25.55′′ N, 90°42′43.75′′ E, 2657 m, on stone, 4 August 2015, M. Sulayman 26705 (XJU).
Description: Plants in loose tufts, yellowish to light green. Stems are 2–2.5 cm, matted with white tomentum in the upper part, brownish below. Cross-section of stem is rounded, consisting of 2–3 layers of smaller, obviously thick-walled outer cortical cells, numerous rows of larger and thinner-walled inner cortical cells, and 5–7 rows of central strand cells. Leaves are erect-spreading when wet, loosely appressed and slightly flexuous when dry, 4.5–5.7 × 0.73–0.83 mm, and lanceolate. Leaf margins are subentire, slightly serrulate distally, and unistratose. Costa is ca. 100 µm wide at the leaf base, about 1/7 of leaf width at the base, slightly mammillose in the upper part on abaxial surface, with one row of guide cells in the transverse section, two stereid bands, and differentiated ventral and dorsal epidermis. Leaf laminal is unistratose and smooth. Upper laminal cells are (11–)15–25(–30) × 5–11 µm, irregular in shape, and moderately thick-walled; middle laminal cells are 15–33(–46) × 6–11 µm, rectangular, moderately thick-walled, and slightly porose. Basal laminal cells are elongated–rectangular, 46–80 × 5–13 µm, and pitted. Alar cells are well differentiated, bistratose, brownish, and not extending to the costa, with a group of thin-walled hyaline cells between the alar cells and costa. Setae are 1.0–1.5 cm. Capsules are curved, furrowed, and ca. 2 mm long. Annuli not seen; operculum with a long, straight beak. Spores are 20–25 μm and papillose.
Note: In China, Dicranum schjakovii grows on soil or decaying wood at altitudes ranging from 773 m to 3200 m. It is usually associated with Pleurozium schreberi (Willd. ex Brid.) Mitt. and Ptilidium pulcherrimum (Weber) Vain. D. schjakovii is easily confused with D. spadiceum owing to the irregular upper laminal cells in both species, but can be separated by the size of their leaves: D. schljakovii has leaves measuring 4.5–5.7 × 0.73–0.83 mm, while D. spadiceum has larger leaves measuring 7–8 × 1.3–1.5 mm.
Dicranum spadiceum J.E.Zetterst., Kongl. Svenska Vetensk. Acad. Handl., n.s. 5(10): 20. 1865 (Figure 2G,H and Figure 6).
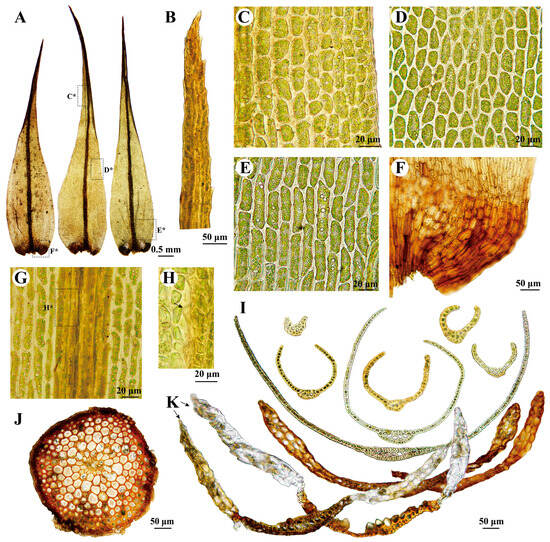
Figure 6.
Dicranum spadiceum J.E. Zetterst. (A–G,I,J) from L. Shu & W.Z. Huang 20220817-38 (HSNU); (H,K) from M. Sulayman 16887 (XJU). (A) Leaves, C*, D*, E*, F* indicates the position of C, D, E, F on the leaf, respectively; (B) Leaf apex; (C) Upper laminal cells of leaf; (D) Middle laminal cells of leaf; (E) Basal lamianl cells of leaf; (F) Alar cells of leaf; (G,H) Costa in the upper portion of leaf (dorsal view), H* indicates the position of H, arrow in H shows weakly mammillose; (I,K) Cross-sections of leaf; (J) Cross-section of stem.
Distribution in China: Qinghai (present report), Xinjiang (present report), new to China.
Specimens examined: China. Xinjiang. Urumqi City, Urumqi County, Mount Nanshan, Xiaoquzi forestry farm, 43°26′63′′ N, 87°05′22′′ E, 2420 m, on soil, 2 July 2011, Horyat Abliz 450 (XJU); Xinjiang, Altai region, Altai city, Xiaodonggou Forest Park, 48°00′20.4′′ N, 88°18′33.5′′ E, 1904 m, on rock, 7 August 2011, M. Sulayman 16887 (XJU); Xinjiang, Mount Altai, Qiinggil County, Qibahalegai, 47°04′14.00′′ N, 90°13′59.37′′ E, 1600 m, on decaying wood, 2 August 2015, M. Sulayman 26582 (XJU); Qoqak, the Barluk Mountain National Nature Reserve, 45°47′58′′ N, 82°59′50′′ E, 1781 m, on soil, 22 July 2022, M. Sulayman 37449 (XJU). Qinghai. Haidong City, Huzhutuzu Autonomous County, Huzhubeishan National Forest Park, near the Shengmutianchi, 36°53′0.94″N, 102°19′39.81″E, 3737 m, on soil, 17 August 2022, L. Shu & W.Z. Huang 20220817-34 (HSNU), L. Shu & W.Z. Huang 20220817-38 (HSNU), L. Shu & W.Z. Huang 20220817-39 (HSNU).
Description: Plants in loose tufts, light brownish green. Stems are 3–5 cm and moderately tomentose. Cross-section of stem is rounded, consisting of 2–4 layers of smaller, obviously thick-walled outer cortical cells, numerous rows of larger and thinner-walled inner cortical cells, and 5–7 rows of central strand cells. Leaves are straight and erect when wet, loosely appressed when dry, 7–8 × 0.13–0.15 mm, gradually to abruptly narrowed into long and narrow tubular acumen from the ovate base, semicircular in the transverse section. Leaf margins are subentire, slightly serrulate distally, and unistratose. Costa is ca. 120 µm wide at the leaf base, about 1/13 to 1/10 of leaf width at the base, smooth or weakly mammillose in the upper part on the abaxial surface, with one row of guide cells in transverse section, two stereid bands, and differentiated ventral and dorsal epidermis. Leaf lamina is unistratose and smooth. Upper and middle laminal cells are 20–41 × 9–13 µm, irregular in shape, moderately thick-walled, and slightly porose. Basal laminal cells are elongated–rectangular, 45–85 × 8–12 µm, and pitted. Alar cells are well differentiated, 2-4-stratose, brownish, and hyaline, with few or no thin-walled cells between the costa and alar groups. Sporophyte not seen.
Note: This species is widely distributed in Europe [2], North America [3], and Russia [5]. In China, we discovered this species in an alpine meadow in Qinghai, along with three specimens from Xinjiang. It grows on soil, rock, or decaying wood, from 1600 m to 3737 m, usually associated with Cirriphyllum cirrosum (Schwägr.) Grout, Sanionia uncinata (Hedw.) Loeske, and Distichium capillaceum (Hedw.) Bruch & Schimp.
According to the description and illustration of the lectotype specimen of Dicranum spadiceum by Ignatova et al. [5], only two layers of alar cells are present in this species. This characteristic was also noted and confirmed by Hedenäs and Bisang [2], Ireland Jr. [3], Ignatov and Ignatova [49], and Lüth [50]. In our samples, three specimens from Xinjiang also exhibit bistratose alar cells (Figure 6K), while two specimens from Qinghai show partially 3–4-layer alar cells (Figure 6I). Our phylogenetic analyses indicate that these two intriguing samples from Qinghai are highly nested within the other samples (BSML = 99, PPBI = 1; Figure 1), suggesting that they belong to D. spadiceum. We propose that this characteristic may be influenced by environmental factors, as the two specimens from Qinghai were found at high altitudes of 3737 m, whereas specimens from Xinjiang (this study), Europe, and Asian Russia were found at lower altitudes ranging from 704 m to 2430 m [5]. The differences between D. spadiceum and D. schljakovii are discussed under the latter species.
2.3. Taxonomy Treatment
Dicranoloma fragile Broth. Nat. Pflanzenfam. (ed. 2) 10: 209. 1924.
≡Dicranum psathyrum Klazenga, J. Hattori Bot. Lab. 87: 118. 1999. Type: Nepal. In Nepaul legit Hon. D. Gardner, et ad J. Banks Baronetum communicavit Gul. Wallich (BM). syn. nov.
Distribution in China: Dicranoloma fragile is common in the south of China [33,34,35,36], including Anhui, Fujian, Guangdong, Guangxi, Guizhou, Hainan, Hunan, Sichuan, Xizang, Yunnan, and Zhejiang [33,34].
Notes: This species was initially described as Dicranum fragile Hook., nom. illeg., by Hooker [52]. Later, Brotherus [53] formally published the species as Dicranoloma fragile Broth. However, Klazenga [54] moved this species to the genus Dicranum due to the absence of limbidium and differences in the transverse section of the costa from all Dicranoloma species. Nevertheless, the epithet “fragile” was already in use for Dicranum fragile Brid. (≡Campylopus fragile (Brid.) Bruch & Schimp.) at that time. Consequently, Klazenga [54] proposed using “psathyrum” as an alternative.
According to our phylogenetic analysis, Dicranum psathyrum was found deeply nested within the genus Dicranoloma (BSML = 99, PPBI = 0.93) (Figure 1). Consequently, Klazenga’s [54] taxonomic classification should be rejected, the taxonomic status of Dicranoloma fragile needs to be reinstated, and Dicranum psathyrum should be treated as a synonym of Dicranoloma fragile.
2.4. Chinese Dicranum Species and Annotation
Dicranum assamicum Dixon, J. Bombay Nat. Hist. Soc. 39: 774. 1937.
Distribution in China: Distributed only in the Himalayan region in China [35,36,55], including Chongqing, Sichuan [33,34], Yunnan [56], and Xizang [30,33,34].
Note: Dicranum assamicum may be confused with D. japonicum Mitt. because both species possess 2–4 layers of alar cells. However, D. assamicum can be well distinguished by the serrated costa on the upper portion of the leaf, straight capsules, smooth peristome teeth above, and the absence of minute and longitudinal point striations below [30,56,57], whereas D. japonicum is characterized by 2–3(–4) serrated ridges on the upper portion of the costa, slightly curved capsules, densely papillose peristome teeth above, and minute and longitudinal point striations below [58].
Dicranum baicalense Tubanova, Arctoa 31(2): 147. 2022.
Distribution in China: inner Mongolia [38].
Note: This species has been known from Russia (southern Siberia and the southern part of the Russian Far East) [10] and China [38].
Dicranum baicalense may be mistakenly identified as D. leioneuron Kindb., D. nipponense Besch., and D. japonicum in China. However, they can be distinguished by the following characteristics: (1) D. leioneuron is characterized by a straight capsule, while D. baicalense has an arcuate capsule; (2) D. japonicum features alar cells that are 2–4-layered, with the capsule becoming striated when dry; in contrast, D. baicalense has 2-layered alar cells and a smooth capsule when dry; (3) D. nipponense has distal leaf cells measuring 50–80 µm, with short cells above the alar cells extending upwards along the leaf margins, whereas D. baicalense has distal leaf cells measuring 65–100 µm and lacks short cells above the alar cells.
Note: New to China (present study).
Dicranum bonjeanii De Not., Elenc. Musch.: 29. 1837.
Distribution in China: Heilongjiang [33,34], Jilin [33,34], inner Mongolia [33,34,59], Shaanxi [60], and Yunnan [38].
Note: Dicranum bonjeanii can be recognized by its elongated–rectangular upper cells, weakly to strongly transversely undulate upper leaf portions, and comparable orientation of apical and lower leaves. This species could only be confused with D. polysetum Sw. in China, and the differences between them are discussed under D. polysetum.
Dicranum cheoi E.B.Bartram, Ann. Bryol. 8: 8. 1936.
Distribution in China: Endemic to China, known only from Guizhou [33,34,61], and Xizang [33,34].
Note: Dicranum cheoi may be confused with D. fuscescens, D. hamulosum Mitt., D. bardunovii, and D. muehlenbeckii Bruch & Schimp. However, the epidermal cells on the ventral side of the costa are not differentiated in D. fuscescens, D. hamulosum, and D. bardunovii, but well differentiated in D. cheoi. In addition, the upper leaf lamina is bistratose along the margins in D. cheoi, but unistratose in D. muehlenbeckii.
Dicranum crispifolium Müll.Hal., Bot. Zeitung (Berlin) 22: 349. 1864.
Distribution in China: Sichuan [33,34], Yunnan [33,34], Xizang [33,34].
Note: Both Dicranum drummondii and D. dispersum exhibit multilayered alar cells, and quadrate or rectangular laminal cells in the upper portion of the leaf, resembling those of D. crispifolium. However, epidermal cells on the ventral side of the costa are differentiated in D. dispersum but not differentiated in D. crispifolium. Differences between D. crispifolium and D. drummondii are discussed under the latter species.
Dicranum dispersum Engelmark, Stuttgarter Beitr. Naturk., A 592: 4. 1999 (Figure 2C,D and Figure 4).
Note: New to China (present study).
Dicranum drummondii Müll.Hal., Syn. Musc. Frond. 1: 356. 1848.
Distribution in China: Mainly distributed in the northeast and southwest of China [35,36,55], including Guizhou [33,34,61], Heilongjiang [28,59], inner Mongolia [59], Jilin [28,33,34], Shaanxi [33,34], Sichuan [33,34], and Xizang [33,34].
Note: Dicranum drummondii, a member of the large-sized Dicranum species, is frequently confused with D. crispifolium in China. These two species share 3–5-layered alar cells and papillose laminal cells on the dorsal surface in the upper portion of the leaf. However, D. drummondii can be easily separated by its unistratose laminal cells along the margins, while they are bistratose in D. crispifolium. In addition, the upper leaf laminal cells are rugose in D. drummondii, contrasting with the non-rugose leaves of D. crispifolium.
Dicranum elongatum Schleich. ex Schwägr., Sp. Musc. Frond., Suppl. 1, 1: 171. 1811.
Distribution in China: Guizhou [61], Hebei [33,34], inner Mongolia [33,34,59], Jilin [33,34], Liaoning [34], Sichuan [33,34], Xinjiang [34], Yunnan [33,34,56].
Note: Dicranum elongatum is closely related to D. setifolium Cradot and D. groenlandicum, but can be distinguished by the presence of short, smooth laminal cells on the upper portion of the leaf. In contrast, D. setifolium exhibits elongated–rhomboidal cells, and D. groenlandicum exhibits long, porose cells throughout the leaf.
Dicranum flagellare Hedw., Sp. Musc. Frond.: 130. 1801.
Distribution in China: Common in the northeast and southwest of China [35,36,55], including Guangxi, Heilongjiang [33,34], Hubei, inner Mongolia [59], Jilin [33,34], Shandong [62], Taiwan, Xizang [30], and Yunnan (present report).
Note: Dicranum flagellare is easily recognized by its small size and universal occurrence of numerous flagellae that arise in the upper leaf axils. This species bears a resemblance to D. mayrii Broth. and D. ignatovii Tubanova & Fedosov in terms of flagelliform branchlets but differs from D. ignatovii by its unistratose alar cells (in contrast to the bistratose alar cells in D. ignatovii) and differs from D. mayrii by its almost smooth laminal cells and costa in the distal portion of leaf (which are sharply and densely scabrose in D. mayri).
Dicranum fragilifolium Lindb., Öfvers. Kongl. Vetensk.-Akad. Förh. 14(4): 125. 1857.
Distribution in China: Common in the north and southwest of China [35,36,55], including Chongqing, Guizhou [61], Heilongjiang [28,33], Hubei, inner Mongolia [33], Jilin [28], Shaanxi, Sichuan, Taiwan [33], Yunnan [56], Xinjiang [63], and Zhejiang [64].
Note: Dicranum fragilifolium is characterized by its straight and erect, stiff, and fragile leaves. This species differs from D. hakkodense Cardot and Dicranoloma fragile, with few blunt teeth at the apices (in contrast to sharply denticulate or serrated in Dicranum hakkodense and Dicranoloma fragile). Furthermore, it can be distinguished from Dicranum hengduanense by the distinct stereid bands present in the lower part of its costa.
Dicranum fulvum Hook., Musci Exot. 2: 149. 1819.
Distribution in China: Common in the northeast and southwest of China [35,36,55], including Chongqing [65], Guizhou [33,34], Heilongjiang [28], Jilin [26], Liaoning [26], and Yunnan [56].
Note: Dicranum fulvum can be distinguished from all Chinese Dicranum species by its larger proportion of bistratose leaf lamina, except for D. shennongjiaense and D. majus Turner. However, laminal cells in the upper leaf portion are regularly quadrate to short rectangular in D. fulvum, contrasting with the rectangular and elongated cells in D. shennongjiaense and D. majus.
Dicranum fuscescens Turner, Muscol. Hibern. Spic. 60. pl. 5: f. 1804.
Distribution in China: Guizhou [33,34], Heilongjiang [33,34], Jilin [33,34], Liaoning [33,34], inner Mongolia [33,34], Shaanxi [34], Sichuan [34], Xizang [33,34], and Zhejiang [66].
Note: Dicranum fuscescens might be confused with D. muehlenbeckii, D. cheoi, and D. hamulosum, and the differences are annotated under the latter species, respectively.
Dicranum groenlandicum Brid., Muscol. Recent. Suppl. 4: 68. 1819.
Distribution in China: Mainly distributed in the north and southwest of China [35,36,55], including Heilongjiang [33,34], inner Mongolia [33,34,59], Jilin [33,34], Xinjiang [63], Xizang [30], and Yunnan [33,34].
Note: Dicranum groenlandicum, D. elongatum, D. himalayanum Mitt., and D. setifolium are closely related and can be easily confused; the distinctions among them are annotated under the latter three species, respectively.
Dicranum hakkodense Cardot, Bull. Herb. Boissier, sér. 2, 7: 714. 1907.
Distribution in China: Sichuan [6].
Note: Dicranum hakkodense has long been classified as D. viride var. hakkodense (Cardot) Takaki, but was subsequently reclassified as a distinct species based on molecular evidence by Ignatova and Fedosov [6]. The presence of sharply denticulate leaf apices serves as a key feature for distinguishing between D. hakkodense and D. viride; while the former exhibits sharp denticulation apices, the latter has only a few blunt teeth or entirely smooth apices. Variations in comparison to D. fragilifolium and D. hengduanense are individually discussed within the context of the latter species.
Dicranum hamulosum Mitt., Trans. Linn. Soc. London, Bot. 3: 156. 1891.
Distribution in China: Guangxi [34], Hainan [34], Heilongjiang [26], Jilin [33,34], Liaoning [26], Sichuan [34], Taiwan [33,34], Xizang [34], Yunnan [34], and Zhejiang [33,34].
Note: Dicranum hamulosum might be confused with D. cheoi and D. fuscescens, but can be distinguished by the unistratose alar cells, contrasting with the bistratose alar cells found in D. cheoi and D. fuscescens. In addition, the epidermal cells on the ventral side of the costa are undifferentiated in D. hamulosum but show differentiation in D. cheoi.
Dicranum hengduanense W.Z.Huang & R.L.Zhu, Bryologist 126(2): 229. 2023.
Distribution in China: Endemic to China. At present, Dicranum hengduanense is known only in two localities in Yunnan province, southwestern China [7].
Note: Dicranum hengduanensis is one of the Dicranum species with fragile leaves. In contrast to D. hakkodense and D. psathyrum (≡Dicranoloma fragile), this species exhibits entirely smooth leaf apices instead of sharply denticulate ones. Differences from D. fragilifolium are discussed under the latter species. In addition, D. hengduanense can be distinguished from other Dicranum species with delicate leaves by the absence of stereid bands and the presence of only one layer of cells above and below the guide cells [7].
Dicranum himalayanum Mitt., J. Proc. Linn. Soc., Bot., Suppl. 1: 14. 1859.
Distribution in China: Mainly distributed in the southwest of China [35,36,55], including Sichuan, Xizang, and Yunnan [30,33,34,59].
Note: Dicranum himalayanum can be confused with D. groenlandicum due to the smooth leaf margin and prosenchymatous and porose laminal cells in the upper portion of the leaf. However, these two species differ in their growth patterns: D. himalayanum forms loose tufts with branching and possesses 2–3-layered alar cells, whereas D. groenlandicum typically creates very dense cushions with minimal or no branching and has unistratose or occasionally bistratose alar cells [30,34].
Dicranum japonicum Mitt., Trans. Linn. Soc. London, Bot. 3: 155. 1891.
Distribution in China: Widely distributed in China [35,36,55].
Note: Dicranum japonicum may be confused with D. baicalense, D. bonjeanii, D. lorifolium, D. nipponense, and D. scoparium in China. In its typical forms, D. japonicum presents 2–4 layers of alar cells [10]; however, some specimens may exhibit only 2-stratose alar cells [10,58]. Dicranum japonicum can be distinguished by the combination of the following characteristics: (1) inclined and asymmetrical capsules, (2) costa with two lamellae or ridges on its back, (3) leaves 8–9(–11) mm long, being about 10 times as long as they are wide, and (4) the absence of short cells above the alar cells. In addition, confusion between D. japonicum and D. assamicum is possible, with distinct characteristics detailed under D. assamicum.
Dicranum kashmirense Broth., Acta Soc. Sci. Fenn. 24(2): 9. 1899.
Distribution in China: Mainly distributed in the south of China [35,36,55], including Chongqing, Guangxi [33,34,59], Guizhou [61], Hubei, Hunan [33,34,59], Jiangxi [67], Sichuan [33,34,59], and Yunnan [56].
Note: Dicranum kashmirense may be confused with D. scoparium but can be distinguished by the unistratose alar cells in D. kashmirense, while they are bistratose in D. scoparium.
Dicranum leiodontum Cardot, Bull. Herb. Boissier, sér. 2, 7: 714. 1907.
Distribution in China: Jilin, Xinjiang, and Xizang [31,33,34,35,36,55,63].
Note: Dicranum leiodontum may be confused only with D. papillidens, with the distinctions between them detailed under the latter species.
Dicranum linzianum C.Gao, Acta Phytotax. Sin. 17: 115. 1979.
Distribution in China: Endemic to China, known only from Xizang [33,34,35,36,55].
Note: Dicranum linzianum may be confused exclusively with D. spurium due to both having ovate–lanceolate leaves, and the distinguishing characteristics are discussed under the latter species.
Dicranum lorifolium Mitt., J. Proc. Linn. Soc., Bot., Suppl. 1: 15. 1859.
Distribution in China: Mainly distributed in the south of China [35,36,55], including Chongqing, Fujian [33,34], Gansu [33,34], Guizhou [33,34], Jiangxi [67], Taiwan, Xizang [33,34], Yunnan [33,34], and Zhejiang [68].
Note: Dicranum lorifolium may be confused with D. assamicum, D. baicalense, D. bonjeanii, D. japonicum, D. nipponense, and D. scoparium in China, and it is difficult to distinguish them based solely on gametophyte. However, D. lorifolium can be well distinguished by the following unique combination of characteristics: (1) bistratose alar cells, (2) costa above with two lamellae or ridges on its back, (3) the presence of short cells above the alar cells, extending upwards along the leaf margins, and (4) always erect capsules.
Dicranum majus Turner, Muscol. Hibern. Spic.: 58. 1804.
Distribution in China: Widely distributed in China [35,36,66].
Note: Dicranum majus is one of the largest members of the genus when well developed. This is a very variable species [2,69], in terms of leaf curvature and length, the upper leaf lamina being partly bistratose or not, and the upper laminal cells spinosely projecting or not [2]. D. majus may be confused with D. scoparium, but D. scoparium has four distinct ridges on the upper back of the costa rather than furrows and serrations in D. majus. In addition, setae generally aggregate in D. majus but are solitary in D. scoparium [69].
Dicranum mayrii Broth., Hedwigia 38: 207. 1899.
Distribution in China: Chongqing [65], Heilongjiang [33,34], and Taiwan [33,34].
Note: Dicranum mayrii is easily recognized by its universal occurrence of numerous flagellae and mammillose upper laminal cells on the dorsal side. This species could only be confused with D. flagellare in China, and differences between these two species are discussed under D. flagellare.
Dicranum montanum Hedw., Sp. Musc. Frond.: 143. 1801.
Distribution in China: Widely distributed in China [35,36,55], including Guizhou [61], Hainan [33,34], Hebei [33,34], Heilongjiang [33,34], Hubei, inner Mongolia [33,34], Jilin [33,34], Liaoning, Shaanxi [60], Sichuan [60], Xizang [30,33,34], Yunnan [56,60], and Zhejiang [60].
Note: Dicranum montanum is one of the smallest species within the genus in China and is easily recognized because it looks more like Dicranoweisia cirrata (Hedw.) Lindb. than a Dicranum species. In addition, another aid in its identification is the occasional presence of small, delicate, clustered branchlets with linear leaves that are readily detachable (probably a means of asexual reproduction like Brothera leana (Sull.) Müll.Hal.), which occur near the stem apices.
Dicranum muehlenbeckii Bruch & Schimp., Bryol. Eur. 1: 142. 1847.
Distribution in China: Guizhou [28,61], Heilongjiang [28], inner Mongolia [34], Jilin [33,34], Liaoning [28], Shaanxi [55], Sichuan [33,34], Taiwan [55], Xinjiang [28,30,63], Xizang [33,34], Yunnan [56], and Zhejiang [33,34].
Note: Dicraunm muehlenbeckii may be confused with D. fuscescens; however, the epidermal cells on the ventral side of the costa are not differentiated in D. fuscescens, whereas they are well differentiated in D. muehlenbeckii.
Dicranum nipponense Besch., Ann. Sci. Nat., Bot., sér. 7, 17: 332. 1893.
Distribution in China: Widely distributed in China [35,36,55].
Note: Dicranum nipponense may be confused with D. assamicum, D. baicalense, D. bonjeanii, D. japonicum, D. lorifolium, and D. scoparium in China. However, it can be distinguished by the following combination of characteristics: (1) bistratose alar cells, (2) costa above with two lamellae or ridges on its back, (3) the presence of short cells above the alar cells, extending upwards along the leaf margins, (4) capsules inclined and asymmetrical, (5) dark green plants, and (6) leaves 3–7 mm long.
Dicranum papillidens Broth., Sitzungsber. Akad. Wiss. Wien, Math.-Naturwiss. Kl., Abt. 1, 133: 561. 1924.
Distribution in China: Endemic to China, known only from Sichuan [33,34,55].
Note: Dicranum papillidens was first reported by Brotherus [20] in 1924 and has remained elusive for almost a century. This species is characterized by (1) unistratose alar cells, (2) quadrate or short rectangular cells in the upper leaf, (3) a smooth abaxial surface of the upper laminal cells and costa, (4) being strongly porose in the basal and middle leaf cells, and (4) having densely papillose peristome teeth throughout. Dicranum papillidens can be easily separated from D. leiodontum by its strongly porose basal and middle laminal cells as well as its densely papillose peristome teeth.
Dicranum polysetum Sw., Monthly Rev. 34: 538. 1801.
Distribution in China: Mainly distributed in the north and southwest of China [35,36,55], including Heilongjiang [33,34], Henna [69], inner Mongolia [33,34,59], Jilin [33,34], Xinjiang [63], Xizang [33,34], and Yunnan.
Note: Dicranum polysetum is readily distinguishable in the field due to its distinctive leaf orientation, with erect uppermost leaves and spreading lower leaves, as well as strongly undulate leaves. This species can only be confused with D. bonjeanii; however, the orientation of the apical leaves and leaves further down are not clearly different in the latter species.
Dicranum rectifolium Müll.Hal., Nuovo Giorn. Bot. Ital., n.s. 3: 98. 1896.
Distribution in China: Endemic to China, known only from Shaanxi [15].
Note: This species was initially reported by Müller in Shaanxi [15] but has not been rediscovered for over 125 years. Regrettably, there is no available information regarding illustrations or descriptions of this species. This species was classified as an insufficiently known species in the earlier Chinese Checklist of the Mosses [70].
Note: New to China (present study).
Dicranum scoparium Hedw., Sp. Musc. Frond. 126. 1801.
Distribution in China: Widely distributed in China [35,36,55].
Note: Dicranum scoparium is one of the most variable Dicranum species, with both straight and falcate leaves commonly observed, and the presence of both dwarf and large male plants. In China, this species can be distinguished from other members of the D. scoparium complex (including D. baicalense, D. bonjeanii, D. japonicum, D. lorifolium, D. nipponense, and D. polysetum) by the four ridges on the dorsal surface of its costa, as opposed to the typical two ridges found in other species.
Dicranum scottianum Turner, Muscol. Hibern. Spic.: 75. 1804.
Distribution in China: Heilongjiang [28], inner Mongolia [59], Jilin [28], and Taiwan (present study).
Note: Dicranum scottianum is characterized by (1) its asymmetrical leaves, with one side of the leaf being wider than the other, (2) the difference in the angle of insertion of the lamina with respect to the costa, (3) the laminal cells that are thick-walled, and (4) the wide and deep costa that has two bands of stereids. In the only specimen we examined from Taiwan, the margins are dentate in the upper part of the leaf, which can be referrable to D. canariense Hampe ex Müll.Hal. [2]; however, Price et al. [71] synonymized it under D. scottianum in 2016.
Dicranum setifolium Cardot, Bull. Herb. Boissier, sér. 2, 7: 714. 1907.
Distribution in China: Jilin and Sichuan [33,34,35,36,55].
Note: Dicranum setifolium shows a close relation to D. elongatum and D. groenlandicum but is distinguished from the latter two species by the characteristics of its upper and middle laminal cells. In D. groenlandicum, these cells are porose, while in D. setifolium, they are smooth. The differences between D. setifolium and D. elongatum are discussed under the latter species.
Dicranum shennongjiaense W.Z.Huang & R.L.Zhu
Distribution in China: Hubei [38].
Note: Refer to Huang et al. [38] for details.
Dicranum spadiceum J.E.Zetterst., Kongl. Svenska Vetensk. Acad. Handl., n.s. 5(10): 20. 1865 (Figure 2G,H and Figure 6).
Note: New to China (present study).
Dicranum spurium Hedw., Sp. Musc. Frond.: 141. 1801.
Distribution in China: Mainly distributed in the northeast of China [35,36,55], including Heilongjiang [28], inner Mongolia [33,34,59], and Jilin [33,34].
Note: Dicranum spurium may be confused with D. linzianum in China as both species exhibit ovate–lanceolate leaves (relatively broad basal portion, becoming suddenly narrow to a short acumen). However, these species can be differentiated by the upper laminal cells: those of D. spurium are irregular, triangular, or quadrate, with prorulae on the dorsal surface, whereas D. linzianum has oval and smooth upper laminal cells.
Dicranum undulatum Schrad. ex Brid., J. Bot. (Schrader) 1800. 1(2): 294. 1801.
Distribution in China: Mainly distributed in the northeast of China [35,36,55], including Heilongjiang [33,34], inner Mongolia [33,34], Jilin [33,34], Liaoning [28], and Xinjiang [28].
Note: Dicranum undulatum may be confused with D. polysetum and D. bonjeanii in China due to their similarly transversely undulating leaves but can be distinguished by the irregular oval shape of the upper laminal cells, which differ from the elongated–rectangular cells found in the latter two species.
2.5. Excluded Species
Dicranum brevifolium (Lindb.) Lindb., Musci Scand.: 24. 1879.
Note: Sulayman et al. [72] reported the presence of this species in the Altai Mountains, Xinjiang. Although we have not seen the voucher specimen directly, based on their illustrations, it is clearly an incorrect identification: (1) the transverse section of the upper leaf looks like a pair of tongs in D. brevifolium [2,12,47], but it looks more or less like a circular outline in their illustration; (2) the ventral epidermal layer of cells, which are usually poorly differentiated in D. brevifolium [2,8,47], are distinctly differentiated in their illustration. In addition, (3) while the leaf lamina is bistratose along the margins in D. brevifolium, it is unistratose in their illustrations. In conclusion, D. brevifolium should be excluded from the moss flora of China.
Dicranum viride (Sull. & Lesq.) Lindb., Hedwigia 2(11): 70. 1863.
Note: When Ignatova and Fedosov revised Dicranum species with fragile leaves in Russia [6], they found that all records of D. viride in the Russian Far East actually belong to D. hakkodense, and its occurrence in Japan, Korea, and China, as well as in Alaska seems to be doubtful. In China, this species has been recorded in Chongqing [35,36], Guizhou [33,34,61], Hubei [73], Sichuan [33,34], Xinjiang [74], and Yunnan [33,34,56]. However, all illustrations and descriptions of this species in China [33,34,56,61], characterized by serrated leaf apices and elongated–rectangular basal laminal cells, align with D. hakkodense. In addition, our examination of specimens previously labeled as D. viride in China, including M. Zang-5276, L.S. Wang-879, and X.J. Li-4329 from Yunnan (KUN), G.L. Zheng 231 from Hubei (CCNU), M. Sulayman 13008, 25902, 17145 from Xinjiang (XJU), Q. Gao et al. 3186 from inner Mongolia (PE), Huang & Li 2248 from Shaanxi (PE), Y. Liu 320 from Sichuan (PE), and F081116, L. He 050912425 from Guizhou (GACP), revealed that none of them correspond to D. viride.
2.6. Key to Dicranum Species in China
Note: Dicranum rectifolium is not included because we have no information, illustrations, or descriptions of this species, which was merely relisted without additional specimens in a checklist from 1896 and was listed as an insufficiently known species in China [70].
- 1. Leaves stiff and fragile, upper leaf portions very narrow, and tips frequently broken….2
- 1. Leaves usually crisped or falcate, tips mostly present...........................................................6
- 2. Leaf apices entirely smooth or with few blunt teeth..............................................................3
- 2. Leaf apices sharply denticulate or serrulate............................................................................4
- 3. Costa in lower portion of the leaf with stereid bands, sometimes weak, with up to 2–3(–4) layers of cells above and below guide cells...........................................D. fragilifolium Lindb.
- 3. Costa in lower portion of the leaf lacking stereid bands, with up to (0–)1–2 layers of cells above and below guide cells.......................................D. hengduanense W.Z.Huang & R.L.Zhu
- 4. Margin serrated in the upper 1/2–3/4(–5/6), entirely smooth below. Basal laminal cells elongated–rectangular, 30–90(–120) µm long…………………………………...Dicranoloma fragile Broth. (≡ Dicranum psathyrum Klazenga)
- 4. Margin serrated only near apex. Basal laminal cells short rectangular, 20–50 µm long….5
- 5. Leaves only occasionally fragile, crisped when dry; basal laminal cells quadrate to short rectangular, 20–35 µm long; costa broader, occupying 1/3 or more of the total leaf base width; leaf lamina mostly 2-stratose above.........................................D. fulvum Hook. (in part)
- 5. Leaves moderately fragile, straight, or slightly falcate–secund when dry; basal laminal cells elongated–rectangular, 25–50 µm long; costa narrower, occupying less than 1/3 the leaf base width; leaf lamina mostly 1-stratose above...............................D. hakkodense Cardot
- 6. Plant with flagelliform branchlets (rigid and terete branches with appressed leaves) in the distal leaf axils...........................................................................................................................7
- 6. Plant without flagelliform branchlets......................................................................................8
- 7. Upper laminal cells smooth on dorsal surface...........................................D. flagellare Hedw.
- 7. Upper laminal cells mammillose on dorsal surface........................................D. mayrii Broth.
- 8. Laminal cells irregularly bistratose in the upper portion of the leaf…………...………..…9
- 8. Laminal cells unistratose in the upper portion of the leaf, or bistratose only along margins….............................................................................................................................................11
- 9. Laminal cells in the upper leaf portion are regularly quadrate to short rectangular…………………………….....................….......…...………….D. fulvum Hook. (in part)
- 9. Laminal cells in the upper leaf portion are rectangular and elongate….…….….……….10
- 10. Costa with 2 rows of guide cells……………………………....……….…...D. majus Turner
- 10. Costa with 1 row of guide cells…………..……D. shennongjiaense W.Z.Huang & R.L.Zhu
- 11. Cells in upper leaf prosenchymatous and porose..............................................................12
- 11. Cells in upper leaf quadrate or rectangular, rarely elongated–rectangular, with or without pores........................................................................................................................................23
- 12. Leaves straight, in upper part more or less tubular, apex rounded, margin entirely smooth throughout or sometimes with a few obtuse denticles near apex.............................13
- 12. Leaves falcate or straight, in upper part flat or channeled, apex acuminate, margin above finely to coarsely denticulate...........................................................................................14
- 13. Plants in loose tufts, alar cells 2–3-layered............................D. himalayanum Mitt. (in part)
- 13. Plants in dense cushions, alar cells unistratose or rarely bistratose...........................................................................................................................................................D. groenlandicum Brid.
- 14. Costa above with at least four lamellae or numerous ridges or furrows on back (at least in some leaves).................................................................................................D. scoparium Hedw.
- 14. Costa above with two lamellae, or ridges, or smooth on back...........................................15
- 15. Leaf lamina transversely undulate.......................................................................................16
- 15. Leaf lamina not undulate.......................................................................................................17
- 16. Leaf lamina strongly transversely undulate; margin in upper part spinosely denticulate or dentate. Apical leaves erect, leaves below spreading...........................D. polysetum Sw.
- 16. Leaf lamina more or less strongly transversely undulate (at least in some leaves); margin obtusely to sharply denticulate. Orientation of apical leaves and leaves further down not clearly different.........................................................................................D. bonjeanii De Not
- 17. Alar cells unistratose or bistratose.......................................................................................18
- 17. Alar cells 2–4-layered.............................................................................................................42
- 18. Alar cells unistratose...............................................................................D. kashmirense Broth.
- 18. Alar cells bistratose.................................................................................................................19
- 19. Costa above smooth on back...................................................D. himalayanum Mitt. (in part)
- 19. Costa above with two lamellae or ridges on back...............................................................20
- 20. Capsules straight...........................................................................................D. lorifolium Mitt.
- 20. Capsules arcuate.....................................................................................................................21
- 21. Leaves 8–9(–11) mm long..............................................................D. japonicum Mitt. (in part)
- 21. Leaves 3–7.5 mm long............................................................................................................22
- 22. Plants dark green, presence of short cells above the alar cells, extending upwards along the leaf margins...............................................................................................D. nipponense Besch.
- 22. Plants yellowish to light green, no presence of short cells above the alar cells.....................................................................................................................................D. baicalense Tubanova
- 23. Upper leaf lamina in at least some leaves rugose, or more or less transversely undulate (dry and wet).................................................................................................................................24
- 23. Upper leaf lamina neither rugose nor undulate..................................................................26
- 24. Leaf lamina transversely undulate, in upper part with smooth or lowly mammillose cells; basal leaf portion lanceolate, leaf apex more or less obtuse.............................................................................................................................................D. undulatum Schrad. ex Brid.
- 24. Leaf lamina rugose, in upper leaf on back with spine-like or conically projecting cells; basal leaf portion ovate, leaf apex acuminate............................................................................25
- 25. Upper leaf laminal cells irregularly quadrate, walls esinuose; leaves gradually narrowed to long, falcate acumen...............................................................D. drummondii Müll.Hal.
- 25. Upper leaf laminal cells very irregular, triangular, quadrate, with sinuose walls; leaves suddenly narrowed to short acumen...............................................................D. spurium Hedw.
- 26. Alar cells unistratose..............................................................................................................27
- 26. Alar cells bistratose or multilayered.....................................................................................30
- 27. Upper laminal cells mammillose on abaxial surface..........................................................28
- 27. Upper laminal cells smooth on abaxial surface...................................................................29
- 28. Plant small, leaf laminal cells unistratose..............................................D. montanum Hedw.
- 28. Plant size variable, leaf laminal cells bistratose along margins.............D. hamulosum Mitt.
- 29. Peristome teeth densely papillose throughout......................................D. papillidens Broth.
- 29. Peristome teeth smooth above, faintly vertically striated below......D. leiodontum Cardot
- 30. Upper leaf laminal cells bistratose along margins..............................................................31
- 30. Leaf laminal cells unistratose................................................................................................36
- 31. Alar cells 2–3(–4)-stratose......................................................................................................32
- 31. Alar cells bistratose.................................................................................................................33
- 32. Epidermal cells on ventral side of costa differentiated.............................................................................................................................................D. dispersum Engelmark (in part)
- 32. Epidermal cells on ventral side of costa undifferentiated.............D. crispifolium Müll.Hal.
- 33. Laminal cells above strongly incrassate...............................................D. scottianum Turner
- 33. Laminal cells thin-walled or slightly incrassate..................................................................34
- 34. Epidermal cells on ventral side of costa differentiated.......................D. cheoi E.B.Bartram
- 34. Epidermal cells on ventral side of costa not differentiated................................................35
- 35. Leaves falcate–secund.............................................................................D. fuscescens Turner
- 35. Leaves flexuous when dry and erect-spreading when moist...................................................................................................................D. bardunovii Tubanova & Ignatova
- 36. Transverse section of upper leaf keeled...........................D. dispersum Engelmark (in part)
- 36. Transverse section concave...................................................................................................37
- 37. Leaves ovate–lanceolate, with a short, narrowly acuminate apex…..D. linzianum C.Gao
- 37. Leaves narrowly lanceolate, with a long apex.....................................................................38
- 38. Leaves strongly curled when dry. Leaf margin near apex irregularly and coarsely denticulate to dentate.....................................................................D. muehlenbeckii Bruch & Schimp.
- 38. Leaves straight or weakly curled in their upper portions when dry. Leaf margin weakly denticulate to almost entirely smooth........................................................................................39
- 39. Plants more loosely tufted. Upper leaf laminal cells thin-walled to moderately incrassate; leaf base ovate or ovate–lanceolate....................................................................................40
- 39. Plants in dense, compact tufts. Upper leaf laminal cells strongly incrassate; leaf base lanceolate.......................................................................................................................................41
- 40. Plants small, leaves 4.5–5.7 × 0.73–0.83 mm................D. schljakovii Ignatova & Tubanova
- 40. Plants medium-sized, leaves 7–8 × 1.3–1.5 mm.............................D. spadiceum J.E.Zetterst.
- 41. Distal laminal cells short rectangular, quadrate, rounded........................................................................................................................D. elongatum Schleich. ex Schwägr.
- 41. Distal laminal cells elongated–rhomboidal...........................................D. setifolium Cardot
- 42. Capsules straight, peristome teeth smooth above, without being minutely and longitudinally point-striated below......................................................................D. assamicum Dixon
- 42. Capsules slightly curved. peristome teeth densely papillose above, minutely and longitudinally point-striated below........................................................D. japonicum Mitt. (in part)
3. Materials and Methods
3.1. Taxon Sampling
During the course of the present study, some specimens from the following Chinese herbaria were examined, including China Agricultural University (BAU), Central China Normal University (CCNU), East China Normal University (HSNU), Hangzhou Normal University (HTC), Institute of Botany, Chinese Academy of Sciences (PE), Kunming Institute of Botany, Chinese Academy of Sciences (KUN), Lushan Botanical Garden, Chinese Academy of Science (LBG), and Xinjiang University (XJU).
The inter-tribe relationships in Dicranum have been well resolved based on the combined analysis of five chloroplast loci (rps4-trnT, trnL-trnF, trnH-psbA, rps19-rpl2, and rpoB) and the nrITS1-5.8S-ITS2 region [4,7,11,12,14]. To determine the species phylogeny position of Chinese specimens, 29 specimens representing 19 species from China were newly sequenced for all six gene markers (see Supplementary Table S1). A total of 273 published accessions of Dicranum, spanning 36 species from previous studies, were also utilized for the analysis [4,5,6,7,8,11,12,14,38,75,76]. In addition, four accessions of Holomitrium Brid., two accessions of Dicranoloma, and one accession of Fissidens Hedw. were also selected as outgroups. A total of 309 accessions were included in the analysis. A list of taxa with collection localities, vouchers, herbarium acronyms, and GenBank accession numbers are listed in Supplementary Table S2.
3.2. Morphological Study
The plant photos were taken using a digital camera (Canon M6; Canon, Tokyo, Japan). Specimens were examined using a Zeiss stereo zoom scope (Stemi DV4; Zeiss, Oberkochen, Germany) and an Olympus microscope (Olympus BX43; Olympus, Tokyo, Japan), and microscopic images were captured by a digital camera (Olympus DP71; Olympus, Tokyo, Japan) attached to the microscope.
3.3. DNA Extraction, Sequencing, Assembly, and Annotation
The sample preparation, total DNA extraction, and chloroplast genome and nuclear DNA assembly followed protocols previously used in earlier studies [77]. The whole chloroplast genomes were annotated using Geneious v.11.1.5 [78] based on the D. hengduanense W.Z.Huang & R.L.Zhu as the reference plastome (accession number: OQ401775) and then adjusted manually. The assembled nuclear DNA was aligned with published ITS data using D. scoparium as a reference (accession number: KF423564) in Geneious v.11.1.5 [78], and then annotated and extracted.
3.4. Morphological Study
Six datasets were aligned using MAFFT v7.311 [79] and ambiguous alignment regions were trimmed using trimAl v1.2 [80]. Six individual alignments were concatenated in Geneious v.11.1.5 [78]. Absent bases were coded as missing.
Phylogenetic analyses were conducted using maximum likelihood (ML) and Bayesian inference (BI) methods in IQtree v2.0.6 [81] and MrBayes 3.2.6 [82], respectively. IQtree was performed with the best-fitting substitution model for each DNA region (HKY+F+G4 for ITS-partition, TPM2u+F+I+G4 for rps19-rpl2-partition, rpoB-partition, rps4-trnT-partition, and trnL-trnF intergenic spacers, and TPM3+F+G4 for trnH-psbA-partition) selected by ModelFinder according to the Bayesian information criterion (BIC) [83,84], and the fast bootstrap option with 1000 replicates. For BI analyses, each DNA region was also assigned its own substitution model (HKY+G is the for best-fit model for ITS-partition; GTR+I+G for rpoB-partition and for trnL-trnF intergenic spacers; HKY+G for rps4-trnT-partition and for rps19-rpl2-partition; HKY+G for trnH-psbA-partition), as determined by the Akaike information criterion (AIC) [83,84]. Two independent analyses consisting of four Markov chain Monte Carlo (MCMC) chains were run for 5,000,000 generations, with one tree sampled for every 1000 generations. The posterior distribution of trees was summarized by a >50% majority-rule consensus tree after discarding the first 25% of samples as burn-in. Convergence was assessed by examining the likelihood plots in Tracer v.1.7 [85].
4. Conclusions
Four species of Dicranum are newly reported for China, including D. bardunovii, D. dispersum, D. schljakovii, and D. spadiceum. Two species of Dicranum, D. brevifolium and D. viride, are proposed for exclusion from the bryoflora of China. Additionally, Dicranum psathyrum is proposed as a synonym of Dicranoloma fragile. Currently, a total of 39 Dicranum species are known in China.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants13131759/s1, Table S1: Sequences newly generated in the study, including taxon, isolate, locality, vouchers, herbarium code, and GenBank accession numbers; Table S2: Sequences downloaded from Genbank, including taxa, localities, vouchers, herbarium codes, and GenBank accession numbers (rps4-trnT, trnL-trnF, trnH-psbA, rps19-rpl2, rpoB, and ITS).
Author Contributions
Conceptualization, W.-Z.H. and R.-L.Z.; methodology, W.-Z.H.; software, W.-Z.H.; validation, W.-Z.H.; formal analysis, W.-Z.H.; investigation, W.-Z.H., C.S., H.X., M.S., L.S., and R.-L.Z.; resources, Y.-H.W. and R.-L.Z.; data curation, W.-Z.H.; writing—original draft preparation, W.-Z.H.; writing—review and editing, W.-Z.H., R.-L.Z., Y.-H.W., and L.S.; visualization, W.-Z.H.; supervision, W.-Z.H.; project administration, W.-Z.H. and R.-L.Z.; funding acquisition, Y.-H.W. and R.-L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant numbers 32370218, 32270215, and 32060050.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We are grateful to Sheng-Bang Zhang, from the Qinghai Wildlife Conservation Association, for the sampling permit and field assistance, and to the curators and staff of BAU, IFP, LBG, GACP, PE, XJU, GNUG, HTC, and KUN for making specimens available for study through loans.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brinda, J.C.; Atwood, J.J. The Bryophyte Nomenclator. Available online: https://www.bryonames.org/ (accessed on 6 April 2024).
- Hedenäs, L.; Bisang, I. Key to European Dicranum species. Herzogia 2004, 17, 179–197. [Google Scholar]
- Ireland, R.R., Jr. Dicranum (Family Dicranaceae). In Flora of North America; Zander, R.H., Ed.; Oxford University Press: New York, NY, USA, 2007; Volume 27, pp. 397–420. [Google Scholar]
- Lang, A.S.; Stech, M. What’s in a name? Disentangling the Dicranum scoparium species complex (Dicranaceae, Bryophyta). Syst. Bot. 2014, 39, 369–379. [Google Scholar] [CrossRef]
- Ignatova, E.A.; Tubanova, D.Y.; Tumurova, O.D.; Goryunvo, D.V.; Kuznetsova, O.I. When the plant size matters: A new semi-cryptic species of Dicranum from Russia. Arctoa 2015, 24, 471–488. [Google Scholar] [CrossRef][Green Version]
- Ignatova, E.A.; Fedosov, V.E. Species of Dicranum (Dicranaceae, Bryophyta) with fragile leaves in Russia. Arctoa 2008, 17, 63–83. [Google Scholar] [CrossRef]
- Huang, W.Z.; Xu, H.; Ma, X.Y.; Zhu, R.L. Dicranum hengduanensis (Dicranaceae, Bryophyta), a new species with fragile leaves from the Hengduan Mountains in China. Bryologist 2023, 126, 226–235. [Google Scholar] [CrossRef]
- Tubanova, D.Y.; Goryunov, D.V.; Ignatova, E.A.; Ignatov, M.S. On the taxonomy of Dicranum acutifolium and D. fuscescens complexes (Dicranaceae, Bryophyta) in Russia. Arctoa 2010, 19, 151–164. [Google Scholar] [CrossRef]
- Tubanova, D.; Ignatova, E.A. A new species of Dicranum (Dicranaceae, Bryophytha) from Asiatic Russia. Arctoa 2011, 20, 183–190. [Google Scholar] [CrossRef]
- Tubanova, D.; Dugarova, O.D. Dicranum baicalense (Dicranaceae, Bryophyta), a new species from Russia. Arctoa 2022, 31, 145–154. [Google Scholar] [CrossRef]
- Lang, A.S.; Kruijer, J.D.; Stech, M. DNA barcoding of Arctic bryophytes: An example from the moss genus Dicranum (Dicranaceae, Bryophyta). Polar Biol. 2014, 37, 1157–1169. [Google Scholar] [CrossRef]
- Lang, A.S.; Tubanova, D.; Stech, M. Species delimitations in the Dicranum acutifolium complex (Dicranaceae, Bryophyta) using molecular markers. J. Bryol. 2014, 36, 279–290. [Google Scholar] [CrossRef]
- Kiebacher, T.; Szövényi, P. Morphological, genetic and ecological divergence in near-cryptic bryophyte species widespread in the Holarctic: The Dicranum acutifolium complex (Dicranales) revisited in the Alps. J. Plant Res. 2024, 137, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.S.; Bocksberger, G.; Stech, M. Phylogeny and species delimitations in European Dicranum (Dicranaceae, Bryophyta) inferred from nuclear and plastid DNA. Mol. Phylogenet. Evol. 2015, 92, 217–225. [Google Scholar] [CrossRef]
- Müller, C. Bryologia provinciae Schen-si Sinensis I. Nuovo Giorn. Bot. Ital. N.S. 1896, 3, 89–129. [Google Scholar]
- Müller, C. Bryologia provinciae Schen-si Sinensis II. Nuovo Giorn. Bot. Ital. N.S. 1897, 4, 245–276. [Google Scholar]
- Müller, C. Bryologia provinciae Schen-si Sinensis III. Nuovo Giorn. Bot. Ital. N.S. 1898, 5, 158–209. [Google Scholar]
- Salmon, E.S. On some mosses from China and Japan. J. Linn. Soc. Bot. 1900, 34, 449–474. [Google Scholar] [CrossRef]
- Paris, J.É.G.N. Muscinées de I’Asie Orientale (7e article). Rev. Bryol. 1908, 35, 40–55. [Google Scholar]
- Brotherus, V.F. Musci novi sinenses collecti a Dr. Henr. Handel-Mazzetti. II. Sitzungsber. Akad. Wiss. Wien Math.-Naturwiss. Kl. Abt. 1 1924, 133, 559–584. [Google Scholar]
- Brotherus, V.F. Musci. Symb. Sin. 1929, 4, 1–147. [Google Scholar]
- Brotherus, V.F. Musci novi Asiatici. Rev. Bryol. N.S. 1929, 2, 1–16. [Google Scholar]
- Reimers, H. Beiträge zur Moosflora Chinas. I. Hedwigia 1931, 71, 1–77. [Google Scholar]
- Bartram, E.B. Additions to the moss flora of China. Ann. Bryol. 1935, 8, 6–21. [Google Scholar]
- Raven, P.H. Greeting from the Missouri Botanical Garden at the International bryological symposium on the occasion of prof. Pan-chieh Chen’s centennial birthday. Chenia 2007, 9, 5–6. [Google Scholar]
- Chen, P.C. Genera Muscorum Sinicorum, I; Science Publisher: Beijing, China, 1963; pp. 1–304. (In Chinese) [Google Scholar]
- Li, X.J. The bryophytes of the alpine forest region in western Sichuan. In Report on the Comprehensive Investigation of Alpine Forest Vegetation of SW China; Li, X.J., Ed.; Science Publisher: Beijing, China, 1963; pp. 361–410, 417–426. [Google Scholar]
- Gao, C. Flora Muscorum China Boreali-Orientalis; Science Press: Beijing, China, 1977; pp. 1–404. (In Chinese) [Google Scholar]
- Gao, C.; Chang, K.C. Plantae novae bryophytarum Tibeticarum. Acta Phytotax. Sin. 1979, 17, 115–120. [Google Scholar]
- Li, X.J. Bryoflora of Xizang; Science Press: Beijing, China, 1985; pp. 1–581. (In Chinese) [Google Scholar]
- Gao, C.; Cao, T. A synopsis of Chinese Dicranum (Dicranaceae, Musci). Bryobrothera 1992, 1, 215–220. [Google Scholar]
- Wu, P.C. The moss flora of Xishuangbanna, southern Yunnan, China. Trop. Bryol. 1992, 5, 27–33. [Google Scholar]
- Gao, Q. Flora Bryophytarum Sinicorum, Vol. 1.; Science Press: Beijing, China, 1994; pp. 1–368. (In Chinese) [Google Scholar]
- Gao, C.; Crosby, M.R.; He, S. Moss Flora of China (English Version), Vol. 1.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1999; pp. 1–273. [Google Scholar]
- Jia, Y.; He, S. Species Catalogue of China, Vol. 1. Plants: Bryophytes; Science Press: Beijing, China, 2013; pp. 1–525. (In Chinese) [Google Scholar]
- Wang, L.S.; Jia, Y.; Zhang, X.C.; Qin, H.N. Species Catalogue of China, Vol. 1. Plants: A Synoptic Checklist (I); Science Press: Beijing, China, 2018; pp. 1–141. (In Chinese) [Google Scholar]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Huang, W.Z.; Tong, S.G.; Zhu, R.L. Dicranum shennongjiaense WZ Huang & RL Zhu (Dicranaceae, Bryophyta), a new species from Central China supported by morphological and molecular evidence. J. Bryol. 2024, in press. [CrossRef]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 7 April 2024).
- Hodgetts, N.G. Checklist and Country Status of European Bryophytes—Towards a New Red List for Europe; Irish Wildlife Manuals, No. 84; National Parks and Wildlife Service: Mayo, Ireland, 2015; pp. 1–125. [Google Scholar]
- Afonina, O.M.; Breen, A. Dicranum dispersum (Dicranaceae) and Sciuro-hypnum ornellanum (Brachytheciaceae), new to North America. Bryologist 2009, 112, 268–272. [Google Scholar] [CrossRef]
- Valitova, L.A.; Baisheva, E.Z.; Kutueva, A.G. On the distribution of the rare moss species Dicranum dispersum Engelmark in the Southern Urals. Environ. Sci. 2020, 10, 35–40. [Google Scholar]
- Lacy, R.C. Considering threats to the viability of small populations using individual-based models. Ecol. Bull. 2000, 48, 39–51. [Google Scholar]
- Di Marco, M.; Collen, B.; Rondinini, C.; Mace, G.M. Historical drivers of extinction risk: Using past evidence to direct future monitoring. Proc. R. Soc. Lond. Ser. B 2015, 282, 20150928. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Tmelot, M.E. High altitude plants, chemistry of acclimation and adaptation. Stud. Nat. Prod. Chem. 2008, 34, 883–982. [Google Scholar]
- Ma, X.Y.; Xu, H.; Cao, Z.Y.; Shu, L.; Zhu, R.L. Will climate change cause the global peatland to expand or contract? Evidence from the habitat shift pattern of Sphagnum mosses. Glob. Change Biol. 2022, 28, 6419–6432. [Google Scholar] [CrossRef]
- Engelmark, T.B. Dicranum dispersum spec. nov. (Dicranaceae: Bryopsida: Bryophyta). Stuttgarter Beitr. Naturk. A 1999, 592, 1–8. [Google Scholar]
- Otnyukova, T.N. Dicranum orientale (Dicranaceae, Musci), a new species from South Siberia (Tuva Republic, Russia). Bot. Zhurn. 2000, 85, 82–87. [Google Scholar]
- Ignatov, M.S.; Ignatova, E.A. Moss Flora of the Middle European Russia. Vol. 1: Sphagnaceae-Hedwigiaceae; KMK Scientific Press Ltd.: Moscow, Russia, 2003; pp. 1–608. [Google Scholar]
- Lüth, M. Mosses of Europe—A Photographic Flora; Michael Lüth: Freiburg, Germany, 2019; Volume 1, pp. 1–1306. [Google Scholar]
- Otnyukova, T.N.; Ochyra, R. On Dicranum dispersum Engelmark and Dicranum orientale Otnyukova. Arctoa 2003, 12, 115–116. [Google Scholar] [CrossRef][Green Version]
- Hooker, W.J. Musci Exotici 2; Longman: London, UK, 1820; pp. 97–176+31. [Google Scholar]
- Brotherus, V.F. Musci (Laubmoose) 1. Hälfte. In Die Natürlichen Pflanzenfamilien, Zweite Auflage; Engler, H.G.A., Prantl, K., Eds.; Duncker & Humblot: Berlin, Germany, 1924; pp. 1–478. [Google Scholar]
- Klazenga, N. A revision of the Malesian species of Dicranoloma (Dicranaceae, Musci). J. Hattori Bot. Lab. 1999, 87, 1–130. [Google Scholar]
- Redfearn, P.L.J.; Tan, B.C.; He, S. A newly updated and annotated checklist of Chinese mosses. J. Hattori Bot. Lab. 1996, 97, 163–357. [Google Scholar]
- Li, X.J. Flora Yunnanica, Tomus 18, Bryophyta: Musci; Science Press: Beijing, China, 2002; pp. 1–525. [Google Scholar]
- Dixon, H.N. Mosses collected in Assam. J. Bombay Nat. Hist. Soc. 1937, 39, 769–795. [Google Scholar]
- Takaki, N. A revision of Japanese Dicranum. J. Hattori Bot. Lab. 1964, 27, 73–123. [Google Scholar]
- Bai, X.L. Flora Bryophytarum Intramongolicarum; Inner Mongolia University Press: Hohhot, China, 1997; pp. 1–541. (In Chinese) [Google Scholar]
- Chang, M.S. Bryophyta in Flora Tsinglingensis. Tom III. Bryophyta (Part l); Science Publisher: Beijing, China, 1978; pp. 1–329. (In Chinese) [Google Scholar]
- Xiong, Y.X. Bryophyte Flora of Guizhou China, Vol. 1; Guizhou Publishing Group, Guizhou Science and Technology Publishing House: Guiyang, China, 2014; pp. 1–509. (In Chinese) [Google Scholar]
- Luo, X.J.; Zhang, Y.M.; Chen, X.S. A preliminary study on bryophytes in Mount Laoshan. J. Shandong Agric. Univ. 1991, 22, 63–70. [Google Scholar]
- Sulayman, M. New checklist of Xinjiang Liverworts, hornworts and mosses. J. Xinjiang Univ. 2012, 29, 259–267. [Google Scholar]
- Hong, R.L.; Hu, R.L. A study of mosses of Mt. Jiulong in Zhejiang. Res. For. Ecosyst. 1984, 4, 207–241. [Google Scholar]
- Hu, X.Y.; Wu, P.C. Study on the moss flora of Mt. Jinfu, Sichuan Province. Acta Phytotax. Sin. 1991, 29, 315–334. [Google Scholar]
- Hu, R.L.; Wang, Y.F. A survey on the bryophytes from Tian Mu Mountain in Zhejiang Province. J. E. China Norm. Univ. Nat. Sci. 1981, 1, 86–104. (In Chinese) [Google Scholar]
- Ji, M.C.; Qiang, S. Bryophytes of the Matoshan Natural conservation region, Jiangxi Province, China. Chenia 2005, 8, 101–118. [Google Scholar]
- Liu, Y.; Cao, T.; Guo, S.L. The mosses of Zhejiang province, China: An annotated checklist. Aratoa 2005, 14, 95–133. [Google Scholar]
- Wang, S.Y.; Wang, D.Y.; Gao, G.W.; Wang, Z.Y.; Ma, X.F.; Yang, Z.L. The plant resources of Baotianman Natural Sanctuary in Neixiang County. In Scientific Survey of the Baotianman Mountain Reserve; Song, C.S., Ed.; Chinese Forestry Publisher: Beijing, China, 1994; pp. 76–84. [Google Scholar]
- Crosby, M.R.; Magill, R.E.; Allen, B.; He, S. A Checklist of the Mosses; Missouri Botanical Garden: St. Louis, MO, USA, 1999; pp. 1–308. [Google Scholar]
- Price, M.J.; Maier, E.; Ellis, L.T. Dicranum scottianum Turner (Dicranaceae): Synonyms, a type and the correct author. J. Bryol. 2019, 41, 149–156. [Google Scholar] [CrossRef]
- Sulayman, M.; Abdurehim, A.; Li, W.; Wu, P.C. Two new records of Dicranaceae (Bryophyta) to China from Altai mountains, Xinjiang. Acta Bot. Boreali.-Occident. Sin. 2012, 32, 1267–1273. [Google Scholar]
- Zheng, G.L.; Liu, S.X.; Chen, G.Y.; Wang, C.L.; Chen, N. Studies on the vertical distribution of mosses in Mt. Jiugongshan, Hubei, China. Wuhan Bot. Res. 2002, 20, 429–432. (In Chinese) [Google Scholar]
- Gong, Z.S.; Abliz, A.; Abla, G.; Sulayman, M. Study on the bryoflora in the Mushishan Mountain, Hotan, Xinjiang. Arid. Zone Res. 2010, 27, 578–584. (In Chinese) [Google Scholar]
- Stech, M. A reclassification of Dicranaceae (Bryopsida) based on non-coding cpDNA sequence data. J. Hattori Bot. Lab. 1999, 86, 137–159. [Google Scholar]
- Stech, M.; Pfeiffer, T.; Frey, W. Molecular relationships and divergence of palaeoaustral Dicranoloma species (Dicranaceae, Bryopsida). Studies in austral temperate rain forest bryophytes 31. J. Hattori Bot. Lab. 2006, 100, 451–464. [Google Scholar]
- Xiang, Y.L.; Jin, X.J.; Shen, C.; Cheng, X.F.; Shu, L.; Zhu, R.L. New insights into the phylogeny of the complex thalloid liverworts (Marchantiopsida) based on chloroplast genomes. Cladistics 2022, 38, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gbaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MRBAYES 3.2: Efficient Bayesian phylogenetic inference and model selection across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Meth. 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).