
Iron Plaque: A Shield against Soil Contamination and Key to Sustainable Agriculture

 and
and
Abstract
Highlights
- Description of the formation process of iron plaque.
- The factors affecting the formation of iron plaque were summarized.
- Understanding the role of iron plaque in environmental processes.
- Insights into the interactions between iron plaque, plants, and microbes for environmental remediation.
Abstract
1. Introduction
2. Characteristics of IP
2.1. Discovery of IP
2.2. Mineral Composition of IP
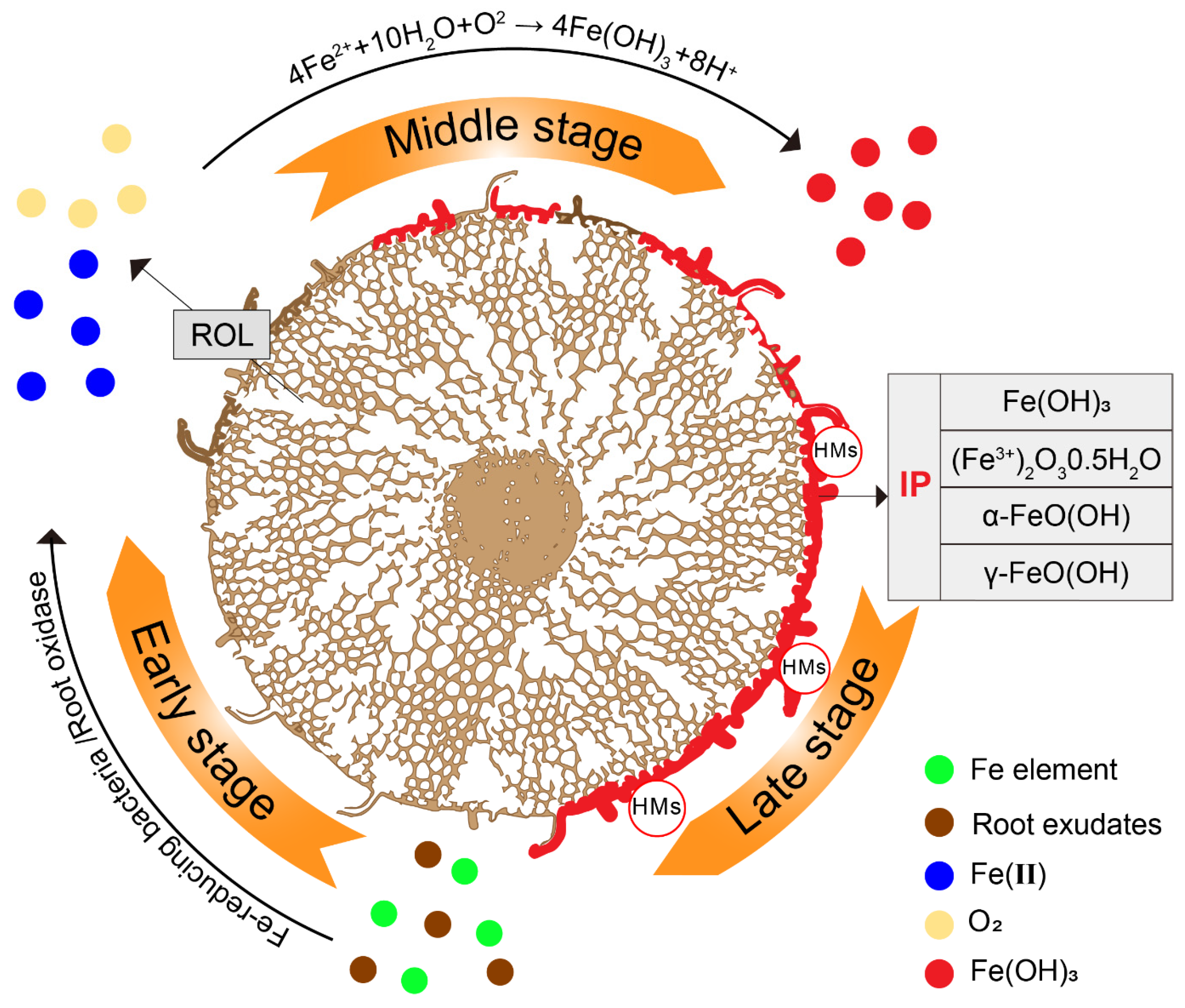
2.3. Formation of IP
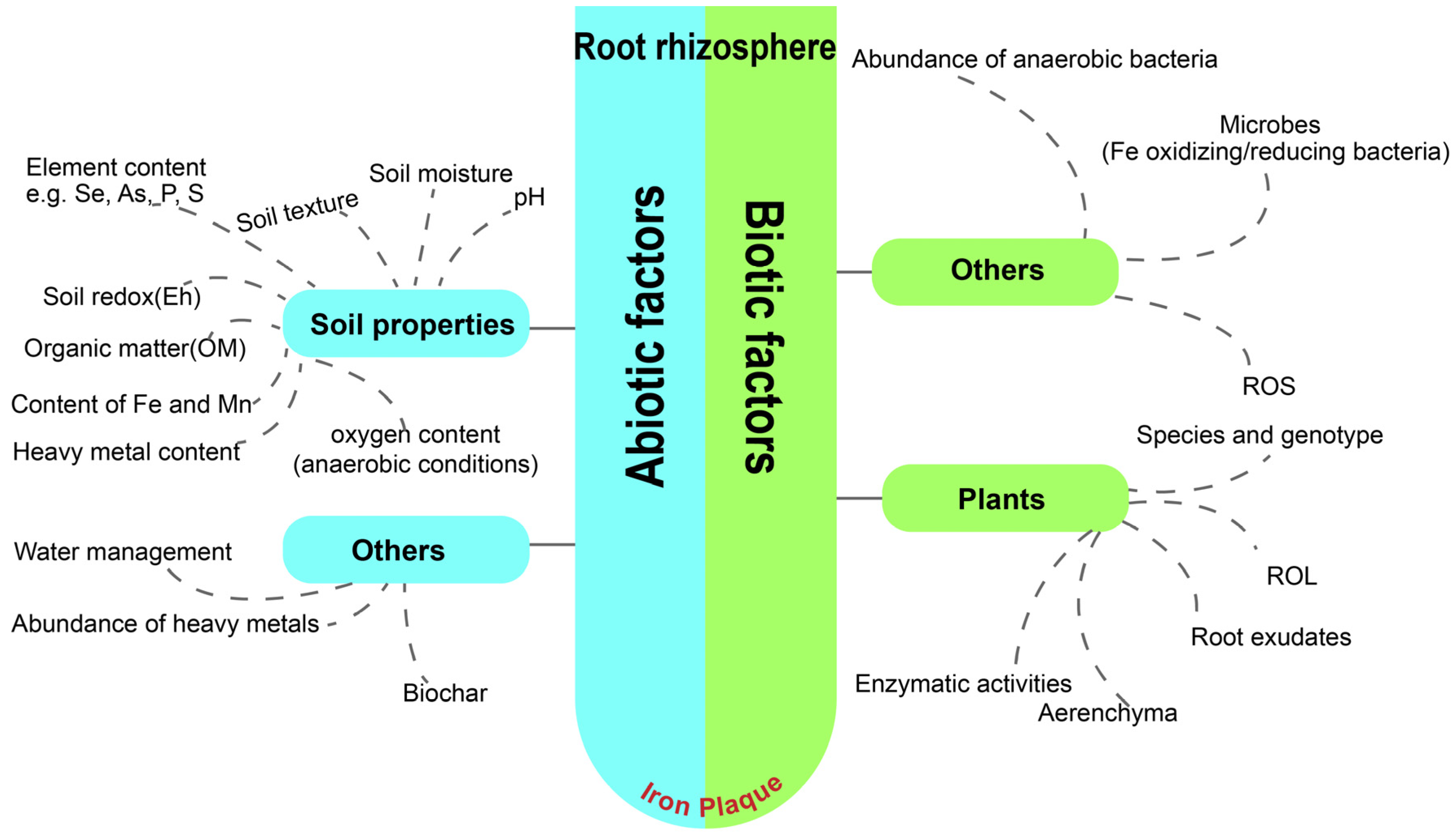
2.3.1. Effect of Abiotic Factors on the Formation of IP
Soil Properties
Irrigation Regime
Other Factors
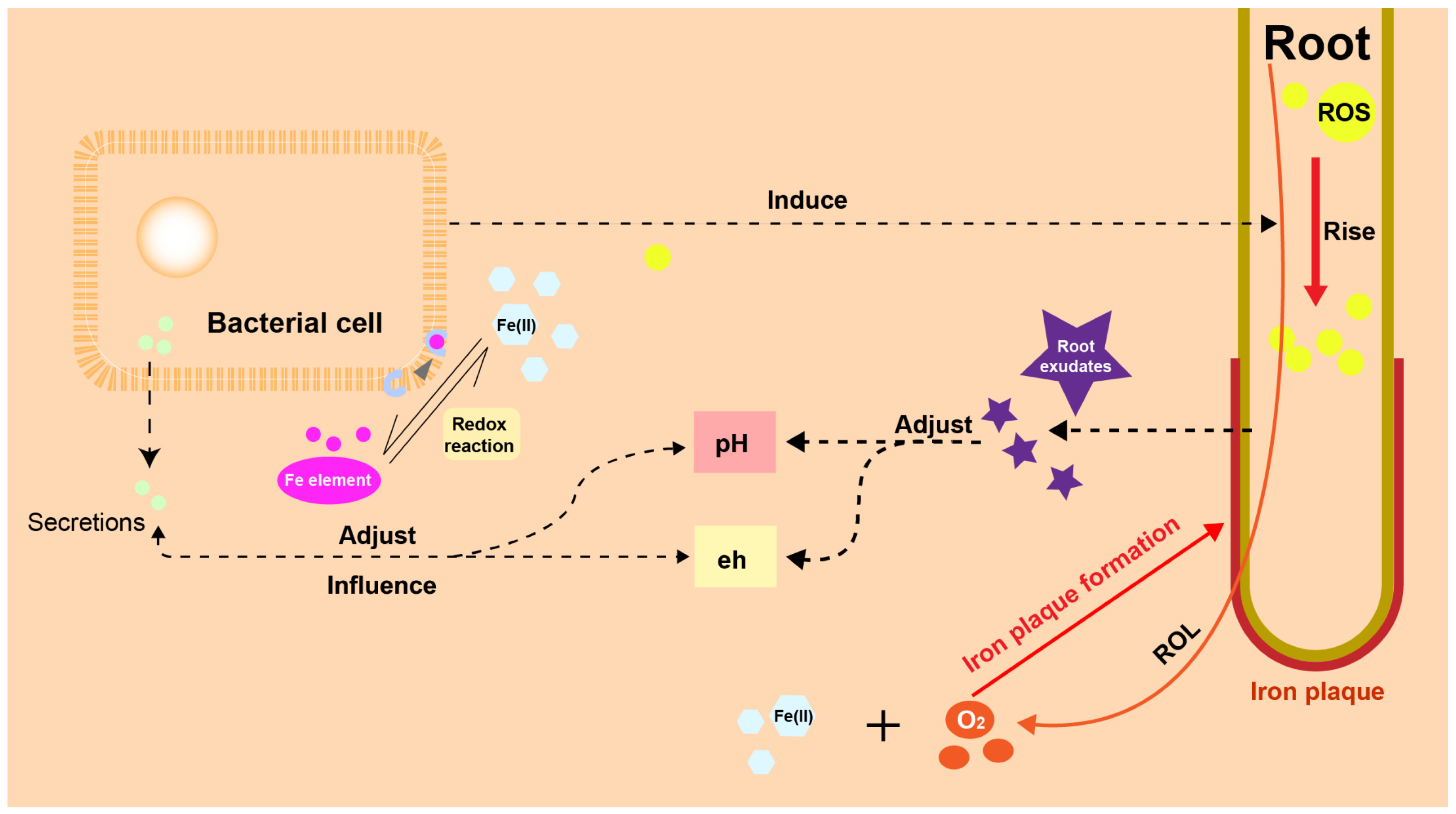
2.3.2. Biotic Factors Effect on IP Formation
Radical Oxygen Loss (ROL) Facilitated by Aeration Tissues (Aerenchyma)
Hydrophyte Oxidative Systems
Fe-Reducing and Fe-Oxidizing Bacteria
2.4. IP as an Armor for Metal Transfer in Plants
2.5. Native Plants in Phytoremediation: Interactions and Ecological Effects on Soils and Plants in Heavy Metal Contaminated Environments
2.5.1. Effect on Plants and Soils
2.5.2. Ecological Role in Environmental Remediation
2.6. Perspectives and Conclusion
2.6.1. Exploring the Cultivation of High-Yielding IP Plant–Microbe Combinations to Address Environmental Pollution
2.6.2. Policy and Sustainability
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Huang, Y.; Chen, Q.; Deng, M.; Japenga, J.; Li, T.; Yang, X.; He, Z. Heavy metal pollution and health risk assessment of agricultural soils in a typical peri-urban area in southeast China. J. Environ. Manag. 2018, 207, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, L.; Wang, W.; Li, T.; He, Z.; Yang, X. Current status of agricultural soil pollution by heavy metals in China: A meta-analysis. Sci. Total Environ. 2019, 651, 3034–3042. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.A.; Zarei, A.; Majidi, S.; Ghaderpoury, A.; Hashempour, Y.; Saghi, M.H.; Alinejad, A.; Yousefi, M.; Hosseingholizadeh, N.; Ghaderpoori, M. Carcinogenic and non-carcinogenic health risk assessment of heavy metals in drinking water of Khorramabad, Iran. MethodsX 2019, 6, 1642–1651. [Google Scholar] [CrossRef] [PubMed]
- Samiee, F.; Vahidinia, A.; Javad, M.T.; Leili, M. Exposure to heavy metals released to the environment through breastfeeding: A probabilistic risk estimation. Sci. Total Environ. 2019, 650, 3075–3083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Jiang, S.; Zhou, M.; Li, F.; Bi, X.; Xie, S.; Liu, J. Heavy metal pollution caused by cyanide gold leaching: A case study of gold tailings in central China. Environ. Sci. Pollut. Res. 2021, 28, 29231–29240. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, E.; Zhang, B.; Bai, X.; Lei, P.; Qiao, X.; Li, Y.-F.; Li, B.; Wu, G.; Gao, Y. Pollution characteristics and ecological risks associated with heavy metals in the Fuyang river system in North China. Environ. Pollut. 2021, 281, 116994. [Google Scholar] [CrossRef]
- da Silva, E.B.; Gao, P.; Xu, M.; Guan, D.; Tang, X.; Ma, L.Q. Background concentrations of trace metals As, Ba, Cd, Co, Cu, Ni, Pb, Se, and Zn in 214 Florida urban soils: Different cities and land uses. Environ. Pollut. 2020, 264, 114737. [Google Scholar] [CrossRef]
- Choppala, G.; Saifullah; Bolan, N.; Bibi, S.; Iqbal, M.; Rengel, Z.; Kunhikrishnan, A.; Ashwath, N.; Ok, Y.S. Cellular mechanisms in higher plants governing tolerance to cadmium toxicity. Crit. Rev. Plant Sci. 2014, 33, 374–391. [Google Scholar] [CrossRef]
- Sun, R.; Yang, J.; Xia, P.; Wu, S.; Lin, T.; Yi, Y. Contamination features and ecological risks of heavy metals in the farmland along shoreline of Caohai plateau wetland, China. Chemosphere 2020, 254, 126828. [Google Scholar] [CrossRef]
- Albornoz, C.B.; Larsen, K.; Landa, R.; Quiroga, M.A.; Najle, R.; Marcovecchio, J. Lead and zinc determinations in Festuca arundinacea and Cynodon dactylon collected from contaminated soils in Tandil (Buenos Aires Province, Argentina). Environ. Earth Sci. 2016, 75, 742. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Pourrut, B.; Lopareva-Pohu, A.; Pruvot, C.; Garçon, G.; Verdin, A.; Waterlot, C.; Bidar, G.; Shirali, P.; Douay, F. Assessment of fly ash-aided phytostabilisation of highly contaminated soils after an 8-year field trial: Part 2. Influence on plants. Sci. Total Environ. 2011, 409, 4504–4510. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Niu, Z.; Yu, J.; Li, Z.; Ma, J.; Xiang, P. Soil heavy metal pollution and food safety in China: Effects, sources and removing technology. Chemosphere 2021, 267, 129205. [Google Scholar] [CrossRef]
- Singh, N.; Rai, U.; Tewari, A.; Singh, M. Metal accumulation and growth response in Vigna radiata L. inoculated with chromate tolerant rhizobacteria and grown on tannery sludge amended soil. Bull. Environ. Contam. Toxicol. 2010, 84, 118–124. [Google Scholar] [CrossRef]
- Jamla, M.; Khare, T.; Joshi, S.; Patil, S.; Penna, S.; Kumar, V. Omics approaches for understanding heavy metal responses and tolerance in plants. Curr. Plant Biol. 2021, 27, 100213. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, X.; Shen, H. Root iron plaque alleviates cadmium toxicity to rice (Oryza sativa) seedlings. Ecotoxicol. Environ. Saf. 2018, 161, 534–541. [Google Scholar] [CrossRef]
- Ma, H.; Gao, F.; Zhang, X.; Cui, B.; Liu, Y.; Li, Z. Formation of iron plaque on roots of Iris pseudacorus and its consequence for cadmium immobilization is impacted by zinc concentration. Ecotoxicol. Environ. Saf. 2020, 193, 110306. [Google Scholar] [CrossRef]
- Wang, T.; Peverly, J.H. Iron oxidation states on root surfaces of a wetland plant (Phragmites australis). Soil Sci. Soc. Am. J. 1999, 63, 247–252. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Ohkura, T.; Takahashi, Y.; Maejima, Y.; Arao, T. Arsenic distribution and speciation near rice roots influenced by iron plaques and redox conditions of the soil matrix. Environ. Sci. Technol. 2014, 48, 1549–1556. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, H.; Xu, C.; Zhu, H.; Zhu, Q. Heavy metal uptake in rice is regulated by pH-dependent iron plaque formation and the expression of the metal transporter genes. Environ. Exp. Bot. 2019, 162, 392–398. [Google Scholar] [CrossRef]
- Fu, Y.-Q.; Yang, X.-J.; Ye, Z.-H.; Shen, H. Identification, separation and component analysis of reddish brown and non-reddish brown iron plaque on rice (Oryza sativa) root surface. Plant Soil 2016, 402, 277–290. [Google Scholar] [CrossRef]
- Chen, R.F.; Shen, R.F.; Gu, P.; Dong, X.Y.; Du, C.W.; Ma, J.F. Response of rice (Oryza sativa) with root surface iron plaque under aluminium stress. Ann. Bot. 2006, 98, 389–395. [Google Scholar] [CrossRef]
- Armstrong, W. The oxidising activity of roots in waterlogged soils. Physiol. Plant. 1967, 20, 920–926. [Google Scholar] [CrossRef]
- Bacha, R.; Hossner, L. Characteristics of coatings formed on rice roots as affected by iron and manganese additions. Soil Sci. Soc. Am. J. 1977, 41, 931–935. [Google Scholar] [CrossRef]
- Batty, L.C.; Baker, A.J.; Wheeler, B.D. Aluminium and phosphate uptake by Phragmites australis: The role of Fe, Mn and Al root plaques. Ann. Bot. 2002, 89, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Chabbi, A.; Hines, M.E.; Rumpel, C. The role of organic carbon excretion by bulbous rush roots and its turnover and utilization by bacteria under iron plaques in extremely acid sediments. Environ. Exp. Bot. 2001, 46, 237–245. [Google Scholar] [CrossRef]
- Khan, N.; Seshadri, B.; Bolan, N.; Saint, C.; Kirkham, M.; Chowdhury, S.; Yamaguchi, N.; Lee, D.; Li, G.; Kunhikrishnan, A. Root iron plaque on wetland plants as a dynamic pool of nutrients and contaminants. Adv. Agron. 2016, 138, 1–96. [Google Scholar]
- Zhang, X.; Zhang, F.; Mao, D. Effect of iron plaque outside roots on nutrient uptake by rice (Oryza sativa L.). Zinc uptake by Fe-deficient rice. Plant Soil 1998, 202, 33–39. [Google Scholar] [CrossRef]
- Zhong, S.; Yuping, W.; Jianming, X. Phosphorus utilization and microbial community in response to lead/iron addition to a waterlogged soil. J. Environ. Sci. 2009, 21, 1415–1423. [Google Scholar] [CrossRef]
- Hansel, C.M.; La Force, M.J.; Fendorf, S.; Sutton, S. Spatial and temporal association of As and Fe species on aquatic plant roots. Environ. Sci. Technol. 2002, 36, 1988–1994. [Google Scholar] [CrossRef] [PubMed]
- Hansel, C.M.; Fendorf, S.; Sutton, S.; Newville, M. Characterization of Fe plaque and associated metals on the roots of mine-waste impacted aquatic plants. Environ. Sci. Technol. 2001, 35, 3863–3868. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Baker, A.J.; Wong, M.-H.; Willis, A.J. Zinc, lead and cadmium accumulation and tolerance in Typha latifolia as affected by iron plaque on the root surface. Aquat. Bot. 1998, 61, 55–67. [Google Scholar] [CrossRef]
- Deng, D.; Wu, S.-C.; Wu, F.-Y.; Deng, H.; Wong, M.-H. Effects of root anatomy and Fe plaque on arsenic uptake by rice seedlings grown in solution culture. Environ. Pollut. 2010, 158, 2589–2595. [Google Scholar] [CrossRef]
- Lee, C.-H.; Hsieh, Y.-C.; Lin, T.-H.; Lee, D.-Y. Iron plaque formation and its effect on arsenic uptake by different genotypes of paddy rice. Plant Soil 2013, 363, 231–241. [Google Scholar] [CrossRef]
- Xiao, A.; Li, W.C.; Ye, Z. Effects of Fe-oxidizing bacteria (FeOB) on iron plaque formation, As concentrations and speciation in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2020, 190, 110136. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-J.; Zhang, J.-L.; Zhang, F.-S. Role of iron plaque in Cd uptake by and translocation within rice (Oryza sativa L.) seedlings grown in solution culture. Environ. Exp. Bot. 2007, 59, 314–320. [Google Scholar] [CrossRef]
- Greipsson, S.; Crowder, A. Amelioration of copper and nickel toxicity by iron plaque on roots of rice (Oryza sativa). Can. J. Bot. 1992, 70, 824–830. [Google Scholar] [CrossRef]
- Zandi, P.; Yang, J.; Xia, X.; Tian, Y.; Li, Q.; Możdżeń, K.; Barabasz-Krasny, B.; Wang, Y. Do sulfur addition and rhizoplane iron plaque affect chromium uptake by rice (Oryza sativa L.) seedlings in solution culture? J. Hazard. Mater. 2020, 388, 121803. [Google Scholar] [CrossRef]
- Xu, B.; Wang, F.; Zhang, Q.; Lan, Q.; Liu, C.; Guo, X.; Cai, Q.; Chen, Y.; Wang, G.; Ding, J. Influence of iron plaque on the uptake and accumulation of chromium by rice (Oryza sativa L.) seedlings: Insights from hydroponic and soil cultivation. Ecotoxicol. Environ. Saf. 2018, 162, 51–58. [Google Scholar] [CrossRef]
- Xiao, W.; Ye, X.; Zhu, Z.; Zhang, Q.; Zhao, S.; Chen, D.; Gao, N.; Hu, J. Continuous flooding stimulates root iron plaque formation and reduces chromium accumulation in rice (Oryza sativa L.). Sci. Total Environ. 2021, 788, 147786. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Yu, S. Root iron plaque formation and characteristics under N2 flushing and its effects on translocation of Zn and Cd in paddy rice seedlings (Oryza sativa). Ann. Bot. 2013, 111, 1189–1195. [Google Scholar] [CrossRef] [PubMed]
- Singha, K.T.; Sebastian, A.; Prasad, M.N.V. Iron plaque formation in the roots of Pistia stratiotes L.: Importance in phytoremediation of cadmium. Int. J. Phytoremediat. 2019, 21, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yan, Z.; Li, X. Ferrous iron facilitates the formation of iron plaque and enhances the tolerance of Spartina alterniflora to artificial sewage stress. Mar. Pollut. Bull. 2020, 157, 111379. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sun, X.; Zhang, Q.; Li, X.; Yan, Z. Iron plaque formation and heavy metal uptake in Spartina alterniflora at different tidal levels and waterlogging conditions. Ecotoxicol. Environ. Saf. 2018, 153, 91–100. [Google Scholar] [CrossRef]
- Chen, C.; Dixon, J.; Turner, F. Iron coatings on rice roots: Morphology and models of development. Soil Sci. Soc. Am. J. 1980, 44, 1113–1119. [Google Scholar] [CrossRef]
- Zhang, Q.; Yan, Z.; Li, X. Iron plaque formation and rhizosphere iron bacteria in Spartina alterniflora and Phragmites australis on the redoxcline of tidal flat in the Yangtze River Estuary. Geoderma 2021, 392, 115000. [Google Scholar] [CrossRef]
- Zhang, J.-Y.; Zhou, H.; Gu, J.-F.; Huang, F.; Yang, W.-J.; Wang, S.-L.; Yuan, T.-Y.; Liao, B.-H. Effects of nano-Fe3O4-modified biochar on iron plaque formation and Cd accumulation in rice (Oryza sativa L.). Environ. Pollut. 2020, 260, 113970. [Google Scholar] [CrossRef]
- Cornell, R.M.; Schwertmann, U. The Iron Oxides: Structure, Properties, Reactions, Occurrences, and Uses; Wiley-VCH: Weinheim, Germany, 2003; Volume 664. [Google Scholar]
- St-Cyr, L.; Fortin, D.; Campbell, P. Microscopic observations of the iron plaque of a submerged aquatic plant (Vallisneria americana Michx). Aquat. Bot. 1993, 46, 155–167. [Google Scholar] [CrossRef]
- Childs, C.W. Ferrihydrite: A review of structure, properties and occurrence in relation to soils. Z. Pflanzenernährung Bodenkd. 1992, 155, 441–448. [Google Scholar] [CrossRef]
- Cornell, R.; Schwertmann, U. The Iron Oxides; VCH Publishers: New York, NY, USA, 1996. [Google Scholar]
- Manceau, A.; Combes, J. Structure of Mn and Fe oxides and oxyhydroxides: A topological approach by EXAFS. Phys. Chem. Miner. 1988, 15, 283–295. [Google Scholar] [CrossRef]
- Szytuła, A.; Burewicz, A.; Dimitrijević, Ž.; Kraśnicki, S.; Rżany, H.; Todorović, J.; Wanic, A.; Wolski, W. Neutron diffraction studies of α-FeOOH. Phys. Status Solidi (b) 1968, 26, 429–434. [Google Scholar] [CrossRef]
- Singh, B.; Sherman, D.; Gilkes, R.; Wells, M.; Mosselmans, J.F. Incorporation of Cr, Mn and Ni into goethite (α-FeOOH): Mechanism from extended X-ray absorption fine structure spectroscopy. Clay Miner. 2002, 37, 639–649. [Google Scholar] [CrossRef]
- Schwertmann, U.; Cornell, R.M. Iron Oxides in the Laboratory: Preparation and Characterization; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Post, J.E. Manganese oxide minerals: Crystal structures and economic and environmental significance. Proc. Natl. Acad. Sci. USA 1999, 96, 3447–3454. [Google Scholar] [CrossRef]
- Ahmad, A.; Nye, P. Coupled diffusion and oxidation of ferrous iron in soils. I. Kinetics of oxygenation of ferrous iron in soil suspension. J. Soil Sci. 1990, 41, 395–409. [Google Scholar] [CrossRef]
- Povidisa, K.; Delefosse, M.; Holmer, M. The formation of iron plaques on roots and rhizomes of the seagrass Cymodocea serrulata (R. Brown) Ascherson with implications for sulphide intrusion. Aquat. Bot. 2009, 90, 303–308. [Google Scholar] [CrossRef]
- Shimamura, S.; Mochizuki, T.; Nada, Y.; Fukuyama, M. Formation and function of secondary aerenchyma in hypocotyl, roots and nodules of soybean (Glycine max) under flooded conditions. Plant Soil 2003, 251, 351–359. [Google Scholar] [CrossRef]
- Holzschuh, M.J.; Carlos, F.S.; Carmona, F.d.C.; Bohnen, H.; Anghinoni, I. Iron oxidation on the surface of adventitious roots and its relation to aerenchyma formation in rice genotypes. Rev. Bras. Ciência Solo 2014, 38, 185–192. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M. Iron plaque decreases cadmium accumulation in Oryza sativa L. and serves as a source of iron. Plant Biol. 2016, 18, 1008–1015. [Google Scholar] [CrossRef]
- Evans, D.E. Aerenchyma formation. New Phytol. 2004, 161, 35–49. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, H.; Chen, L.; Li, Y.; Wan, X. Efficient iron plaque formation on tea (Camellia sinensis) roots contributes to acidic stress tolerance. J. Integr. Plant Biol. 2019, 61, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-C.; Syu, C.-H.; Wang, P.-J.; Lee, D.-Y.; Fan, C.; Juang, K.-W. Field experiment for determining lead accumulation in rice grains of different genotypes and correlation with iron oxides deposited on rhizosphere soil. Sci. Total Environ. 2018, 610, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-L.; Yang, C.-M.; Syu, C.-H.; Lee, D.-Y.; Tsuang, B.-J.; Juang, K.-W. Combined effect of rice genotypes and soil characteristics on iron plaque formation related to Pb uptake by rice in paddy soils. J. Soils Sediments 2016, 16, 150–158. [Google Scholar] [CrossRef]
- Darma, A.; Yang, J.; Bloem, E.; Możdżen, K.; Zandi, P. Arsenic biotransformation and mobilization: The role of bacterial strains and other environmental variables. Environ. Sci. Pollut. Res. 2021, 29, 1763–1787. [Google Scholar] [CrossRef] [PubMed]
- Guerinot, M.L. It’s elementary: Enhancing Fe3+ reduction improves rice yields. Proc. Natl. Acad. Sci. USA 2007, 104, 7311–7312. [Google Scholar] [CrossRef]
- Tai, Y.; Tam, N.F.-Y.; Wang, R.; Yang, Y.; Lin, J.; Wang, J.; Yang, Y.; Li, L.; Sun, Y. Iron plaque formation on wetland-plant roots accelerates removal of water-borne antibiotics. Plant Soil 2018, 433, 323–338. [Google Scholar] [CrossRef]
- Zhang, C.; Ge, Y.; Yao, H.; Chen, X.; Hu, M. Iron oxidation-reduction and its impacts on cadmium bioavailability in paddy soils: A review. Front. Environ. Sci. Eng. 2012, 6, 509–517. [Google Scholar] [CrossRef]
- Christensen, K.K.; Jensen, H.S.; Andersen, F.Ø.; Holmer, M.; Wigand, C. Interferences between root plaque formation and phosphorus availability for isoetids in sediments of oligotrophic lakes. Biogeochemistry 1998, 43, 107–128. [Google Scholar] [CrossRef]
- Jun-Xing, Y.; Yong, L.; Zhi-Hong, Y. Root-induced changes of pH, Eh, Fe (II) and fractions of Pb and Zn in rhizosphere soils of four wetland plants with different radial oxygen losses. Pedosphere 2012, 22, 518–527. [Google Scholar]
- Masscheleyn, P.H.; Delaune, R.D.; Patrick, W.H., Jr. Effect of redox potential and pH on arsenic speciation and solubility in a contaminated soil. Environ. Sci. Technol. 1991, 25, 1414–1419. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Soil–plant transfer of trace elements—An environmental issue. Geoderma 2004, 122, 143–149. [Google Scholar] [CrossRef]
- Mishra, S.; Jha, A.; Dubey, R. Arsenite treatment induces oxidative stress, upregulates antioxidant system, and causes phytochelatin synthesis in rice seedlings. Protoplasma 2011, 248, 565–577. [Google Scholar] [CrossRef]
- Huang, G.; Ding, C.; Li, Y.; Zhang, T.; Wang, X. Selenium enhances iron plaque formation by elevating the radial oxygen loss of roots to reduce cadmium accumulation in rice (Oryza sativa L.). J. Hazard. Mater. 2020, 398, 122860. [Google Scholar] [CrossRef] [PubMed]
- Jiaofeng, G.; Yang, H.; Peng, Z.; Bohan, L.; Hang, Z. Increasing phosphorus inhibits the retention and prevention of cadmium by iron plaque and promotes cadmium accumulation in rice plants. Chemosphere 2022, 307, 135642. [Google Scholar] [CrossRef]
- Liu, W.J.; Zhu, Y.G.; Smith, F.; Smith, S. Do phosphorus nutrition and iron plaque alter arsenate (As) uptake by rice seedlings in hydroponic culture? New Phytol. 2004, 162, 481–488. [Google Scholar] [CrossRef]
- Yang, J.; Tam, N.F.-Y.; Ye, Z. Root porosity, radial oxygen loss and iron plaque on roots of wetland plants in relation to zinc tolerance and accumulation. Plant Soil 2014, 374, 815–828. [Google Scholar] [CrossRef]
- Lambers, H.; Hayes, P.E.; Laliberte, E.; Oliveira, R.S.; Turner, B.L. Leaf manganese accumulation and phosphorus-acquisition efficiency. Trends Plant Sci. 2015, 20, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Liu, J.; Liu, W.; Lu, H.; Jia, H.; Hong, H.; Yan, C. Phosphorus effects on radial oxygen loss, root porosity and iron plaque in two mangrove seedlings under cadmium stress. Mar. Pollut. Bull. 2017, 119, 262–269. [Google Scholar] [CrossRef]
- Yang, Y.-P.; Zhang, H.-M.; Yuan, H.-Y.; Duan, G.-L.; Jin, D.-C.; Zhao, F.-J.; Zhu, Y.-G. Microbe mediated arsenic release from iron minerals and arsenic methylation in rhizosphere controls arsenic fate in soil-rice system after straw incorporation. Environ. Pollut. 2018, 236, 598–608. [Google Scholar] [CrossRef]
- Cao, Z.-Z.; Qin, M.-L.; Lin, X.-Y.; Zhu, Z.-W.; Chen, M.-X. Sulfur supply reduces cadmium uptake and translocation in rice grains (Oryza sativa L.) by enhancing iron plaque formation, cadmium chelation and vacuolar sequestration. Environ. Pollut. 2018, 238, 76–84. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Wan, X.; Zheng, G.; Yang, J.; Zhang, H.; Guo, L.; Wang, X.; Zhou, X.; Guo, Q. Interaction between sulfur and lead in toxicity, iron plaque formation and lead accumulation in rice plant. Ecotoxicol. Environ. Saf. 2016, 128, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E. Reactive iron in marine sediments. Geochim. Cosmochim. Acta 1989, 53, 619–632. [Google Scholar] [CrossRef] [PubMed]
- D de Wit, R.; Stal, L.J.; Lomstein, B.A.; Herbert, R.A.; Van Gemerden, H.; Viaroli, P.; Cecherelli, V.-U.; Rodríguez-Valera, F.; Bartoli, M.; Giordani, G. ROBUST: The ROle of BUffering capacities in STabilising coastal lagoon ecosystems. Cont. Shelf Res. 2001, 21, 2021–2041. [Google Scholar] [CrossRef]
- Liu, J.; Cao, C.; Wong, M.; Zhang, Z.; Chai, Y. Variations between rice cultivars in iron and manganese plaque on roots and the relation with plant cadmium uptake. J. Environ. Sci. 2010, 22, 1067–1072. [Google Scholar] [CrossRef]
- Gschwend, F.; Aregger, K.; Gramlich, A.; Walter, T.; Widmer, F. Periodic waterlogging consistently shapes agricultural soil microbiomes by promoting specific taxa. Appl. Soil Ecol. 2020, 155, 103623. [Google Scholar] [CrossRef]
- Sun, W.; Sun, X.; Li, B.; Häggblom, M.M.; Han, F.; Xiao, E.; Zhang, M.; Wang, Q.; Li, F. Bacterial response to antimony and arsenic contamination in rice paddies during different flooding conditions. Sci. Total Environ. 2019, 675, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wei, Q.; Yu, Y.; Peng, S.; Yang, S. Influence of water management on the mobility and fate of copper in rice field soil. J. Soils Sediments 2013, 13, 1180–1188. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, W.; Liu, J.; Li, H.; Cai, H.; Hu, H.; Ren, D.; Zhang, Y.; Shen, Y.; Wang, J. Crucial role of iron plaque on thallium uptake by rice plant. Waste Dispos. Sustain. Energy 2023, 5, 89–96. [Google Scholar] [CrossRef]
- Weiss, J.V.; Emerson, D.; Megonigal, J.P. Geochemical control of microbial Fe (III) reduction potential in wetlands: Comparison of the rhizosphere to non-rhizosphere soil. FEMS Microbiol. Ecol. 2004, 48, 89–100. [Google Scholar] [CrossRef]
- Wissing, L.; Kölbl, A.; Schad, P.; Bräuer, T.; Cao, Z.-H.; Kögel-Knabner, I. Organic carbon accumulation on soil mineral surfaces in paddy soils derived from tidal wetlands. Geoderma 2014, 228, 90–103. [Google Scholar] [CrossRef]
- Yan, D.; Ma, W.; Song, X.; Bao, Y. The effect of iron plaque on uptake and translocation of norfloxacin in rice seedlings grown in paddy soil. Environ. Sci. Pollut. Res. 2017, 24, 7544–7554. [Google Scholar] [CrossRef]
- Yang, X.-J.; Xu, Z.; Shen, H. Drying–submergence alternation enhanced crystalline ratio and varied surface properties of iron plaque on rice (Oryza sativa) roots. Environ. Sci. Pollut. Res. 2018, 25, 3571–3587. [Google Scholar] [CrossRef]
- Tripathi, R.D.; Tripathi, P.; Dwivedi, S.; Kumar, A.; Mishra, A.; Chauhan, P.S.; Norton, G.J.; Nautiyal, C.S. Roles for root iron plaque in sequestration and uptake of heavy metals and metalloids in aquatic and wetland plants. Metallomics 2014, 6, 1789–1800. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Li, R.; Chai, M.; Cheng, S.; Tam, N.F.Y.; Han, J. Does combined heavy metal stress enhance iron plaque formation and heavy metal bioaccumulation in Kandelia obovata? Environ. Exp. Bot. 2021, 186, 104463. [Google Scholar] [CrossRef]
- Cheng, H.; Jiang, Z.-Y.; Liu, Y.; Ye, Z.-H.; Wu, M.-L.; Sun, C.-C.; Sun, F.-L.; Fei, J.; Wang, Y.-S. Metal (Pb, Zn and Cu) uptake and tolerance by mangroves in relation to root anatomy and lignification/suberization. Tree Physiol. 2014, 34, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Wang, M.; Wong, M.H.; Ye, Z. Does radial oxygen loss and iron plaque formation on roots alter Cd and Pb uptake and distribution in rice plant tissues? Plant Soil 2014, 375, 137–148. [Google Scholar] [CrossRef]
- Pi, N.; Tam, N.; Wong, M.H. Formation of iron plaque on mangrove roots receiving wastewater and its role in immobilization of wastewater-borne pollutants. Mar. Pollut. Bull. 2011, 63, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Basir, A.; Fahad, S.; Adnan, M.; Saleem, M.H.; Iqbal, A.; Amanullah; Al-Huqail, A.A.; Alosaimi, A.A.; Saud, S. Biochar optimizes wheat quality, yield, and nitrogen acquisition in low fertile calcareous soil treated with organic and mineral nitrogen fertilizers. Front. Plant Sci. 2022, 13, 879788. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zheng, G.; Yang, J.; Wan, X.; Song, B.; Cai, W.; Guo, J. Phytoaccumulation of heavy metals (Pb, Zn, and Cd) by 10 wetland plant species under different hydrological regimes. Ecol. Eng. 2017, 107, 56–64. [Google Scholar] [CrossRef]
- Zheng, R.-L.; Cai, C.; Liang, J.-H.; Huang, Q.; Chen, Z.; Huang, Y.-Z.; Arp, H.P.H.; Sun, G.-X. The effects of biochars from rice residue on the formation of iron plaque and the accumulation of Cd, Zn, Pb, As in rice (Oryza sativa L.) seedlings. Chemosphere 2012, 89, 856–862. [Google Scholar] [CrossRef]
- Beesley, L.; Moreno-Jiménez, E.; Gomez-Eyles, J.L. Effects of biochar and greenwaste compost amendments on mobility, bioavailability and toxicity of inorganic and organic contaminants in a multi-element polluted soil. Environ. Pollut. 2010, 158, 2282–2287. [Google Scholar] [CrossRef] [PubMed]
- Emerson, D.; Weiss, J.V.; Megonigal, J.P. Iron-oxidizing bacteria are associated with ferric hydroxide precipitates (Fe-plaque) on the roots of wetland plants. Appl. Environ. Microbiol. 1999, 65, 2758–2761. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhu, Y.; Chen, Z.; Yin, X.; Sun, G. Arsenic mobilization and speciation during iron plaque decomposition in a paddy soil. J. Soils Sediments 2012, 12, 402–410. [Google Scholar] [CrossRef]
- Neubauer, S.C.; Toledo-Durán, G.E.; Emerson, D.; Megonigal, J.P. Returning to their roots: Iron-oxidizing bacteria enhance short-term plaque formation in the wetland-plant rhizosphere. Geomicrobiol. J. 2007, 24, 65–73. [Google Scholar] [CrossRef]
- Becker, M.; Asch, F. Iron toxicity in rice—Conditions and management concepts. J. Plant Nutr. Soil Sci. 2005, 168, 558–573. [Google Scholar] [CrossRef]
- Syu, C.-H.; Lee, C.-H.; Jiang, P.-Y.; Chen, M.-K.; Lee, D.-Y. Comparison of As sequestration in iron plaque and uptake by different genotypes of rice plants grown in As-contaminated paddy soils. Plant Soil 2014, 374, 411–422. [Google Scholar] [CrossRef]
- Gilbert, B.; Frenzel, P. Rice roots and CH4 oxidation: The activity of bacteria, their distribution and the microenvironment. Soil Biol. Biochem. 1998, 30, 1903–1916. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.-H.; Zhu, Y.-G.; Huang, Y.-Z.; Hu, H.-Q.; Christie, P. Sequestration of As by iron plaque on the roots of three rice (Oryza sativa L.) cultivars in a low-P soil with or without P fertilizer. Environ. Geochem. Health 2005, 27, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Ye, Z.; Li, H.; Wu, S.; Deng, D.; Zhu, Y.; Wong, M. Do radial oxygen loss and external aeration affect iron plaque formation and arsenic accumulation and speciation in rice? J. Exp. Bot. 2012, 63, 2961–2970. [Google Scholar] [CrossRef]
- Zhou, H.; Zeng, M.; Zhou, X.; Liao, B.-H.; Peng, P.-Q.; Hu, M.; Zhu, W.; Wu, Y.-J.; Zou, Z.-J. Heavy metal translocation and accumulation in iron plaques and plant tissues for 32 hybrid rice (Oryza sativa L.) cultivars. Plant Soil 2015, 386, 317–329. [Google Scholar] [CrossRef]
- Jackson, M.; Armstrong, W. Formation of aerenchyma and the processes of plant ventilation in relation to soil flooding and submergence. Plant Biol. 1999, 1, 274–287. [Google Scholar] [CrossRef]
- Jackson, M.B.; Fenning, T.M.; Jenkins, W. Aerenchyma (gas-space) formation in adventitious roots of rice (Oryza sativa L.) is not controlled by ethylene or small partial pressures of oxygen. J. Exp. Bot. 1985, 36, 1566–1572. [Google Scholar] [CrossRef]
- Drew, M.C.; Jackson, M.B.; Giffard, S.C.; Campbell, R. Inhibition by silver ions of gas space (aerenchyma) formation in adventitious roots of Zea mays L. subjected to exogenous ethylene or to oxygen deficiency. Planta 1981, 153, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Trought, M.; Drew, M. The development of waterlogging damage in young wheat plants in anaerobic solution cultures. J. Exp. Bot. 1980, 31, 1573–1585. [Google Scholar] [CrossRef]
- Arikado, H. Anatomical and ecological responses of barley and some forage crops to the flooding treatment. Bull. Fac. Agric. Mie Univ. 1955, 11, 1–29. [Google Scholar]
- Colmer, T. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef]
- Zandi, P.; Yang, J.; Xia, X.; Barabasz-Krasny, B.; Możdżeń, K.; Puła, J.; Bloem, E.; Wang, Y.; Hussain, S.; Hashemi, S.M. Sulphur nutrition and iron plaque formation on roots of rice seedlings and their consequences for immobilisation and uptake of chromium in solution culture. Plant Soil 2021, 462, 365–388. [Google Scholar] [CrossRef]
- Grosse, W.; Büchel, H.B.; Tiebel, H. Pressurized ventilation in wetland plants. Aquat. Bot. 1991, 39, 89–98. [Google Scholar] [CrossRef]
- Deng, H.; Ye, Z.; Wong, M.H. Lead, zinc and iron (Fe2+) tolerances in wetland plants and relation to root anatomy and spatial pattern of ROL. Environ. Exp. Bot. 2009, 65, 353–362. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, Y.; Smith, F.; Smith, S. Do iron plaque and genotypes affect arsenate uptake and translocation by rice seedlings (Oryza sativa L.) grown in solution culture? J. Exp. Bot. 2004, 55, 1707–1713. [Google Scholar] [CrossRef]
- Crowder, A. Iron oxide plaque on wetland roots. Trends Soil Sci. 1991, 1, 315–329. [Google Scholar]
- Kirk, G. Changes in rice root architecture, porosity, and oxygen and proton release under phosphorus deficiency. New Phytol. 1997, 135, 191–200. [Google Scholar] [CrossRef]
- Kirk, G.; Greenway, H.; Atwell, B.; Ismail, A.; Colmer, T.D. Adaptation of rice to flooded soils. Prog. Bot. 2014, 75, 215–253. [Google Scholar]
- Mei, X.; Wong, M.H.; Yang, Y.; Dong, H.; Qiu, R.; Ye, Z. The effects of radial oxygen loss on arsenic tolerance and uptake in rice and on its rhizosphere. Environ. Pollut. 2012, 165, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.Y.; Chen, X.; Luo, A.C. Iron plaque formation on wetland plants and its influence on phosphorus, calcium and metal uptake. Aquat. Ecol. 2009, 43, 879–890. [Google Scholar] [CrossRef]
- Karimian, N.; Johnston, S.G.; Burton, E.D. Iron and sulfur cycling in acid sulfate soil wetlands under dynamic redox conditions: A review. Chemosphere 2018, 197, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Maisch, M.; Lueder, U.; Kappler, A.; Schmidt, C. From plant to paddy—How rice root iron plaque can affect the paddy field iron cycling. Soil Syst. 2020, 4, 28. [Google Scholar] [CrossRef]
- Yu, C.; Xie, S.; Song, Z.; Xia, S.; Åström, M.E. Biogeochemical cycling of iron (hydr-) oxides and its impact on organic carbon turnover in coastal wetlands: A global synthesis and perspective. Earth-Sci. Rev. 2021, 218, 103658. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Q.; Zhang, Z. Comparative transcriptome combined with metabolomic and physiological analyses revealed ROS-mediated redox signaling affecting rice growth and cellular iron homeostasis under varying pH conditions. Plant Soil 2019, 434, 343–361. [Google Scholar] [CrossRef]
- Bravin, M.N.; Travassac, F.; Le Floch, M.; Hinsinger, P.; Garnier, J.-M. Oxygen input controls the spatial and temporal dynamics of arsenic at the surface of a flooded paddy soil and in the rhizosphere of lowland rice (Oryza sativa L.): A microcosm study. Plant Soil 2008, 312, 207–218. [Google Scholar] [CrossRef]
- Zhang, X.; Dippold, M.A.; Kuzyakov, Y.; Razavi, B.S. Spatial pattern of enzyme activities depends on root exudate composition. Soil Biol. Biochem. 2019, 133, 83–93. [Google Scholar] [CrossRef]
- Yao, J.; Qin, S.; Liu, T.; Clough, T.J.; Wrage-Mönnig, N.; Luo, J.; Hu, C.; Ge, T.; Zhou, S. Rice root Fe plaque enhances oxidation of microbially available organic carbon via Fe (III) reduction-coupled microbial respiration. Soil Biol. Biochem. 2022, 167, 108568. [Google Scholar] [CrossRef]
- Awad, F.; Römheld, V.; Marschner, H. Effect of root exudates on mobilization in the rhizosphere and uptake of iron by wheat plants. Plant Soil 1994, 165, 213–218. [Google Scholar] [CrossRef]
- Ansari, A.A.; Naeem, M.; Gill, S.S.; AlZuaibr, F.M. Phytoremediation of contaminated waters: An eco-friendly technology based on aquatic macrophytes application. Egypt. J. Aquat. Res. 2020, 46, 371–376. [Google Scholar] [CrossRef]
- Delhaize, E.; Ryan, P.R.; Randall, P.J. Aluminum tolerance in wheat (Triticum aestivum L.) (II. Aluminum-stimulated excretion of malic acid from root apices). Plant Physiol. 1993, 103, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Hue, N.; Craddock, G.; Adams, F. Effect of organic acids on aluminum toxicity in subsoils. Soil Sci. Soc. Am. J. 1986, 50, 28–34. [Google Scholar] [CrossRef]
- Ma, J.F.; Zheng, S.J.; Matsumoto, H. Specific secretion of citric acid induced by Al stress in Cassia tora L. Plant Cell Physiol. 1997, 38, 1019–1025. [Google Scholar] [CrossRef]
- Pellet, D.M.; Grunes, D.L.; Kochian, L.V. Organic acid exudation as an aluminum-tolerance mechanism in maize (Zea mays L.). Planta 1995, 196, 788–795. [Google Scholar] [CrossRef]
- PR, R. Malate efflux from root apices and tolerance to aluminum are highly correlated in wheat. Aust. J. Plant Physiol. 1995, 22, 531–536. [Google Scholar]
- Yang, Y.-Y.; Jung, J.-Y.; Song, W.-Y.; Suh, H.-S.; Lee, Y. Identification of rice varieties with high tolerance or sensitivity to lead and characterization of the mechanism of tolerance. Plant Physiol. 2000, 124, 1019–1026. [Google Scholar] [CrossRef]
- Ando, T.; Yoshida, S.; Nishiyama, I. Nature of oxidizing power of rice roots. Plant Soil 1983, 72, 57–71. [Google Scholar] [CrossRef]
- Hancock, J.; Desikan, R.; Neill, S. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 29, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Sairam, R.; Srivastava, G. Oxidative stress and antioxidative system in plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Seppänen, M.; Turakainen, M.; Hartikainen, H. Selenium effects on oxidative stress in potato. Plant Sci. 2003, 165, 311–319. [Google Scholar] [CrossRef]
- Nelidov, S. Microbiology of the flooded soils of rice paddies. Eurasian Soil Sci. 1994, 26, 41–56. [Google Scholar]
- Rentz, J.A.; Kraiya, C.; Luther, G.W.; Emerson, D. Control of ferrous iron oxidation within circumneutral microbial iron mats by cellular activity and autocatalysis. Environ. Sci. Technol. 2007, 41, 6084–6089. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Dong, H.; Kukkadapu, R.; Agrawal, A.; Liu, D.; Zhang, J.; Edelmann, R.E. Biological oxidation of Fe (II) in reduced nontronite coupled with nitrate reduction by Pseudogulbenkiania sp. strain 2002. Geochim. Cosmochim. Acta 2013, 119, 231–247. [Google Scholar] [CrossRef]
- Melton, E.D.; Swanner, E.D.; Behrens, S.; Schmidt, C.; Kappler, A. The interplay of microbially mediated and abiotic reactions in the biogeochemical Fe cycle. Nat. Rev. Microbiol. 2014, 12, 797–808. [Google Scholar] [CrossRef]
- Hedrich, S.; Schlömann, M.; Johnson, D.B. The iron-oxidizing proteobacteria. Microbiology 2011, 157, 1551–1564. [Google Scholar] [CrossRef]
- Field, E.K.; Kato, S.; Findlay, A.J.; MacDonald, D.J.; Chiu, B.K.; Luther III, G.W.; Chan, C.S. Planktonic marine iron oxidizers drive iron mineralization under low-oxygen conditions. Geobiology 2016, 14, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Lueder, U.; Druschel, G.; Emerson, D.; Kappler, A.; Schmidt, C. Quantitative analysis of O2 and Fe2+ profiles in gradient tubes for cultivation of microaerophilic Iron (II)-oxidizing bacteria. FEMS Microbiol. Ecol. 2018, 94, fix177. [Google Scholar] [CrossRef] [PubMed]
- Engel, H.; Hanert, H. Ispleirung von Gallionella ferruginea Ehrenberg. Naturwissenschaften 1967, 54, 147. [Google Scholar] [CrossRef] [PubMed]
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583. [Google Scholar] [CrossRef] [PubMed]
- Kostka, J.E.; Dalton, D.D.; Skelton, H.; Dollhopf, S.; Stucki, J.W. Growth of iron (III)-reducing bacteria on clay minerals as the sole electron acceptor and comparison of growth yields on a variety of oxidized iron forms. Appl. Environ. Microbiol. 2002, 68, 6256–6262. [Google Scholar] [CrossRef] [PubMed]
- Colmer, A.R.; Temple, K.L.; Hinkle, M.E. An iron-oxidizing bacterium from the acid drainage of some bituminous coal mines. J. Bacteriol. 1950, 59, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Hedrich, S.; Schippers, A. Distribution of acidophilic microorganisms in natural and man-made acidic environments. Curr. Issues Mol. Biol. 2021, 40, 25–48. [Google Scholar] [CrossRef] [PubMed]
- Zegeye, A.; Ona-Nguema, G.; Carteret, C.; Huguet, L.; Abdelmoula, M.; Jorand, F. Formation of hydroxysulphate green rust 2 as a single iron (II-III) mineral in microbial culture. Geomicrobiol. J. 2005, 22, 389–399. [Google Scholar] [CrossRef]
- Guo, L.; Cutright, T.J. Effect of citric acid and bacteria on metal uptake in reeds grown in a synthetic acid mine drainage solution. J. Environ. Manag. 2015, 150, 235–242. [Google Scholar] [CrossRef]
- Weiss, J.V.; Emerson, D.; Backer, S.M.; Megonigal, J.P. Enumeration of Fe (II)-oxidizing and Fe (III)-reducing bacteria in the root zone of wetland plants: Implications for a rhizosphere iron cycle. Biogeochemistry 2003, 64, 77–96. [Google Scholar] [CrossRef]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.-E. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [PubMed]
- Begg, C.; Kirk, G.; Mackenzie, A.; Neue, H.U. Root-induced iron oxidation and pH changes in the lowland rice rhizosphere. New Phytol. 1994, 128, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yan, Z.; Bi, Y.; Lei, Y.; Gao, X.; Li, X.; Brix, H. Iron plaque crystallinity, heavy metal toxicity, and metal translocation in Kandelia obovata seedlings as altered by an iron-reducing bacterium under different flooding regimes. Plant Soil 2023, 487, 267–282. [Google Scholar] [CrossRef]
- Chandra, R.; Kumar, V. Mechanism of wetland plant rhizosphere bacteria for bioremediation of pollutants in an aquatic ecosystem. In Advances in Biodegradation and Bioremediation of Industrial Waste; CRC Press: Boca Raton, FL, USA, 2015; p. 329. [Google Scholar]
- Emerson, D. Biogeochemistry and microbiology of microaerobic Fe (II) oxidation. Biochem. Soc. Trans. 2012, 40, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Green, P.; Crowder, A. Iron oxide deposition on axenic and non-axenic roots of rice seedlings (Oryza sativa L.). J. Plant Nutr. 1991, 14, 375–386. [Google Scholar] [CrossRef]
- Greipsson, S. Effect of iron plaque on roots of rice on growth of plants in excess zinc and accumulation of phosphorus in plants in excess copper or nickel. J. Plant Nutr. 1995, 18, 1659–1665. [Google Scholar] [CrossRef]
- Zakaria, Z.; Zulkafflee, N.S.; Mohd Redzuan, N.A.; Selamat, J.; Ismail, M.R.; Praveena, S.M.; Tóth, G.; Abdull Razis, A.F. Understanding potential heavy metal contamination, absorption, translocation and accumulation in rice and human health risks. Plants 2021, 10, 1070. [Google Scholar] [CrossRef]
- Yu, X.-Z.; Lu, M.-R.; Zhang, X.-H. The role of iron plaque in transport and distribution of chromium by rice seedlings. Cereal Res. Commun. 2017, 45, 598–609. [Google Scholar] [CrossRef]
- Wu, J.; Jiao, Y.; Ran, M.; Li, J. The role of an Sb-oxidizing bacterium in modulating antimony speciation and iron plaque formation to reduce the accumulation and toxicity of Sb in rice (Oryza sativa L.). J. Hazard. Mater. 2024, 469, 133897. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhu, Y.-G.; Liu, W.-J.; Meharg, A.A. Direct evidence showing the effect of root surface iron plaque on arsenite and arsenate uptake into rice (Oryza sativa) roots. New Phytol. 2005, 165, 91–97. [Google Scholar] [CrossRef]
- Zandi, P.; Yang, J.; Darma, A.; Bloem, E.; Xia, X.; Wang, Y.; Li, Q.; Schnug, E. Iron plaque formation, characteristics, and its role as a barrier and/or facilitator to heavy metal uptake in hydrophyte rice (Oryza sativa L.). Environ. Geochem. Health 2023, 45, 525–559. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, J.; Christie, P.; Zhang, F. Influence of iron plaque on uptake and accumulation of Cd by rice (Oryza sativa L.) seedlings grown in soil. Sci. Total Environ. 2008, 394, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Maity, S.; Biswas, C.; Banerjee, S.; Guchhait, R.; Adhikari, M.; Chatterjee, A.; Pramanick, K. Interaction of plastic particles with heavy metals and the resulting toxicological impacts: A review. Environ. Sci. Pollut. Res. 2021, 28, 60291–60307. [Google Scholar] [CrossRef]
- Greipsson, S. Effects of iron plaque on roots of rice on growth and metal concentration of seeds and plant tissues when cultivated in excess copper. Commun. Soil Sci. Plant Anal. 1994, 25, 2761–2769. [Google Scholar] [CrossRef]
- Liu, J.; Luo, L. Uptake and transport of Pb across the iron plaque of waterlogged dropwort (Oenanthe javanica DC.) based on micro-XRF and XANES. Plant Soil 2019, 441, 191–205. [Google Scholar] [CrossRef]
- Batty, L.; Baker, A.; Wheeler, B.; Curtis, C. The effect of pH and plaque on the uptake of Cu and Mn in Phragmites australis (Cav.) Trin ex. Steudel. Ann. Bot. 2000, 86, 647–653. [Google Scholar] [CrossRef]
- Wang, H.; Jia, Y.; Wang, S.; Zhu, H.; Wu, X. Bioavailability of cadmium adsorbed on various oxides minerals to wetland plant species Phragmites australis. J. Hazard. Mater. 2009, 167, 641–646. [Google Scholar] [CrossRef]
- Hochella, M.F.; Eggleston, C.M.; Elings, V.B.; Parks, G.A.; Brown, G.E.; Wu, C.M.; Kjoller, K. Mineralogy in two dimensions: Scanning tunneling microscopy of semiconducting minerals with implications for geochemical reactivity. Am. Mineral. 1989, 74, 1233–1246. [Google Scholar]
- Rea, B.A.; Davis, J.A.; Waychunas, G.A. Studies of the reactivity of the ferrihydrite surface by iron isotopic exchange and Mössbauer spectroscopy. Clays Clay Miner. 1994, 42, 23–34. [Google Scholar] [CrossRef]
- Waychunas, G.; Rea, B.; Fuller, C.; Davis, J. Surface chemistry of ferrihydrite: Part 1. EXAFS studies of the geometry of coprecipitated and adsorbed arsenate. Geochim. Cosmochim. Acta 1993, 57, 2251–2269. [Google Scholar] [CrossRef]
- Fuller, C.C.; Davis, J.A.; Waychunas, G.A. Surface chemistry of ferrihydrite: Part 2. Kinetics of arsenate adsorption and coprecipitation. Geochim. Cosmochim. Acta 1993, 57, 2271–2282. [Google Scholar] [CrossRef]
- Eggleston, C.M.; Hug, S.; Stumm, W.; Sulzberger, B.; Afonso, M.D.S. Surface complexation of sulfate by hematite surfaces: FTIR and STM observations. Geochim. Cosmochim. Acta 1998, 62, 585–593. [Google Scholar] [CrossRef]
- Davis, J.A.; Leckie, J.O. Surface ionization and complexation at the oxide/water interface. 3. Adsorption of anions. J. Colloid Interface Sci. 1980, 74, 32–43. [Google Scholar] [CrossRef]
- Robertson, A.; Leckie, J. Acid/base, copper binding, and Cu2+/H+ exchange properties of goethite, an experimental and modeling study. Environ. Sci. Technol. 1998, 32, 2519–2530. [Google Scholar] [CrossRef]
- Hiemstra, T.; Venema, P.; Van Riemsdijk, W.H. Intrinsic proton affinity of reactive surface groups of metal (hydr) oxides: The bond valence principle. J. Colloid Interface Sci. 1996, 184, 680–692. [Google Scholar] [CrossRef]
- Liesack, W.; Schnell, S.; Revsbech, N.P. Microbiology of flooded rice paddies. FEMS Microbiol. Rev. 2000, 24, 625–645. [Google Scholar] [CrossRef]
- Møller, C.L.; Sand-Jensen, K. Iron plaques improve the oxygen supply to root meristems of the freshwater plant, Lobelia dortmanna. New Phytol. 2008, 179, 848–856. [Google Scholar] [CrossRef]
- Møller, C.L.; Sand-Jensen, K. High sensitivity of Lobelia dortmanna to sediment oxygen depletion following organic enrichment. New Phytol. 2011, 190, 320–331. [Google Scholar] [CrossRef]
- Du, J.; Liu, J.; Lu, H.; Hansell, D.; Zhang, Q.; Wang, W.; Yan, C. Effect of external phosphate addition on solid-phase iron distribution and iron accumulation in Mangrove Kandelia obovata (SL). Environ. Sci. Pollut. Res. 2015, 22, 13506–13513. [Google Scholar] [CrossRef]
- Mei, X.-Q.; Yang, Y.; Tam, N.F.-Y.; Wang, Y.-W.; Li, L. Roles of root porosity, radial oxygen loss, Fe plaque formation on nutrient removal and tolerance of wetland plants to domestic wastewater. Water Res. 2014, 50, 147–159. [Google Scholar] [CrossRef]
- Wu, S.; Vymazal, J.; Brix, H. Critical review: Biogeochemical networking of iron in constructed wetlands for wastewater treatment. Environ. Sci. Technol. 2019, 53, 7930–7944. [Google Scholar] [CrossRef]
- Liang, Y.; Zhu, Y.G.; Xia, Y.; Li, Z.; Ma, Y. Iron plaque enhances phosphorus uptake by rice (Oryza sativa) growing under varying phosphorus and iron concentrations. Ann. Appl. Biol. 2006, 149, 305–312. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, F.; Mao, D. Effect of iron plaque outside roots on nutrient uptake by rice (Oryza sativa L.): Phosphorus uptake. Plant Soil 1999, 209, 187–192. [Google Scholar] [CrossRef]
- Kuo, S. Concurrent sorption of phosphate and zinc, cadmium, or calcium by a hydrous ferric oxide. Soil Sci. Soc. Am. J. 1986, 50, 1412–1419. [Google Scholar] [CrossRef]
- Parfitt, R.; Russell, J. Adsorption on hydrous oxides. IV. Mechanisms of adsorption of various ions on goethite. J. Soil Sci. 1977, 28, 297–305. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, H.; Huang, D.; Xu, C.; Zhu, H.; Zhu, Q. Water managements limit heavy metal accumulation in rice: Dual effects of iron-plaque formation and microbial communities. Sci. Total Environ. 2019, 687, 790–799. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, D. Pb-induced cellular defense system in the root meristematic cells of Allium sativum L. BMC Plant Biol. 2010, 10, 40. [Google Scholar] [CrossRef]
- Tong, H.; Hu, M.; Li, F.; Chen, M.; Lv, Y. Burkholderiales participating in pentachlorophenol biodegradation in iron-reducing paddy soil as identified by stable isotope probing. Environ. Sci. Process. Impacts 2015, 17, 1282–1289. [Google Scholar] [CrossRef]
- Yan, Z.; Meng, H.; Zhang, Q.; Bi, Y.; Gao, X.; Lei, Y. Effects of cadmium and flooding on the formation of iron plaques, the rhizosphere bacterial community structure, and root exudates in Kandelia obovata seedlings. Sci. Total Environ. 2022, 851, 158190. [Google Scholar] [CrossRef]
- Majumder, S.; Banik, P. Geographical variation of arsenic distribution in paddy soil, rice and rice-based products: A meta-analytic approach and implications to human health. J. Environ. Manag. 2019, 233, 184–199. [Google Scholar] [CrossRef]
- Potter, P.M.; Navratilova, J.; Rogers, K.R.; Al-Abed, S.R. Transformation of silver nanoparticle consumer products during simulated usage and disposal. Environ. Sci. Nano 2019, 6, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guo, W.; Li, Q.; Wang, Z.; Liu, S. The effects and the potential mechanism of environmental transformation of metal nanoparticles on their toxicity in organisms. Environ. Sci. Nano 2018, 5, 2482–2499. [Google Scholar] [CrossRef]
- Doolette, C.L.; McLaughlin, M.J.; Kirby, J.K.; Navarro, D.A. Bioavailability of silver and silver sulfide nanoparticles to lettuce (Lactuca sativa): Effect of agricultural amendments on plant uptake. J. Hazard. Mater. 2015, 300, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Ramachandra, T.; Sudarshan, P.; Mahesh, M.; Vinay, S. Spatial patterns of heavy metal accumulation in sediments and macrophytes of Bellandur wetland, Bangalore. J. Environ. Manag. 2018, 206, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, J.K.; Chandra, H.; Kalra, S.J.; Mishra, P.; Khan, H.; Yadav, P. Plant–microbe interaction in aquatic system and their role in the management of water quality: A review. Appl. Water Sci. 2017, 7, 1079–1090. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Z.; Wang, S.; Yao, J.; Borthwick, A.G. Vanadium contamination and associated health risk of farmland soil near smelters throughout China. Environ. Pollut. 2020, 263, 114540. [Google Scholar]
- Negrin, V.L.; Botté, S.E.; La Colla, N.S.; Marcovecchio, J.E. Uptake and accumulation of metals in Spartina alterniflora salt marshes from a South American estuary. Sci. Total Environ. 2019, 649, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cao, L.; Song, X.; Wang, X.; Qian, Q.; Liu, W. Effect of Iron Plaque and Selenium on Cadmium Uptake and Translocation in Rice Seedlings (Oryza sativa) Grown in Solution Culture. Int. J. Agric. Biol. 2014, 16, 1159. [Google Scholar]
- Jia, X.; Otte, M.L.; Liu, Y.; Qin, L.; Tian, X.; Lu, X.; Jiang, M.; Zou, Y. Performance of iron plaque of wetland plants for regulating iron, manganese, and phosphorus from agricultural drainage water. Water 2018, 10, 42. [Google Scholar] [CrossRef]
- Hu, X.; Xie, H.; Zhuang, L.; Zhang, J.; Hu, Z.; Liang, S.; Feng, K. A review on the role of plant in pharmaceuticals and personal care products (PPCPs) removal in constructed wetlands. Sci. Total Environ. 2021, 780, 146637. [Google Scholar] [CrossRef]
- Luo, Y.; Shi, W.; You, M.; Zhang, R.; Li, S.; Xu, N.; Sun, W. Polybrominated diphenyl ethers (PBDEs) in the Danjiangkou Reservoir, China: Identification of priority PBDE congeners. Environ. Sci. Pollut. Res. 2021, 28, 12587–12596. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Sun, S.; He, S. Iron plaque formation and its effect on key elements cycling in constructed wetlands: Functions and outlooks. Water Res. 2023, 235, 119837. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-L.; Zhang, X.; Hu, Y.; Sheng, G.-P. New Barrier Role of Iron Plaque: Producing Interfacial Hydroxyl Radicals to Degrade Rhizosphere Pollutants. Environ. Sci. Technol. 2023, 58, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Chen, S.; Shen, C.; He, M.; Zhang, Y.; Ye, J.; Liu, J.; Shi, J. Iron plaque: A barrier layer to the uptake and translocation of copper oxide nanoparticles by rice plants. Environ. Sci. Technol. 2018, 52, 12244–12254. [Google Scholar] [CrossRef] [PubMed]
- Dixit, R.; Wasiullah, X.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.P.; Rai, J.P.; Sharma, P.K. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef]
- Pishchik, V.; Vorob’Ev, N.; Provorov, N.; Khomyakov, Y.V. Mechanisms of plant and microbial adaptation to heavy metals in plant–microbial systems. Microbiology 2016, 85, 257–271. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Plants. | HMs | IP Composition | Reference |
|---|---|---|---|
| Typha latifolia | As | Ferrihydrite (Fe2O3.nH2O) Lepidocrocite (γ-FeOOH) Goethite (α-FeOOH) Siderite (FeCO3) | Hansel et al. (2002) [31] |
| Phalaris arundinacea | As | Ferrihydrite (Fe2O3.nH2O) Goethite (α-FeOOH) Siderite (FeCO3) | Hansel et al. (2002) [31] |
| Phalaris arundinacea | Mn, Pb, Zn | Ferrihydrite (Fe2O3.nH2O) Goethite (α-FeOOH) Siderite (FeCO3) | Hansel et al. (2001) [32] |
| Typha latifolia | Zn, Pb, Cd | Not mentioned | Ye et al. (1998) [33] |
| Oryza sativa L. | As | Not mentioned | Deng et al. (2010) [34]; Lee et al. (2013) [35]; Xiao et al. (2020) [36]; |
| Oryza sativa L. | Cd | Not mentioned | Liu et al. (2007) [37] |
| Oryza sativa L. | As | Not mentioned | Deng et al. (2010) [34]; Lee et al. (2013) [35]; Xiao et al. (2020) [36]; |
| Oryza sativa L. | Cd | Not mentioned | Liu et al. (2007) [37] |
| Oryza sativa L. | Cu, Ni | Not mentioned | Greipsson and Crowder (1992) [38] |
| Oryza sativa L. | Cr | Not mentioned | Zandi et al. (2020) [39]; Xu et al. (2018) [40]; Xiao et al. (2021) [41] |
| Oryza sativa L. | Zn, Cd | Not mentioned | Xu and Yu, (2013) [42] |
| Pistia stratiotes L. | Cd | Not mentioned | Singha et al. (2019) [43] |
| Iris pseudacorus | Cd | Not mentioned | Ma et al. (2020) [18] |
| Spartina alterniflora | Cu, Zn, Pb, Cr | Not mentioned | Zhang et al. (2020) [44]; Xu et al. (2018) [45] |
| Oxides | Hydroxides and Oxide-Hydroxides |
|---|---|
| Hematite α-Fe2O3 | Ferrihydrite Fe2O3·nH2O |
| β-Fe2O3 | Goethite α-FeOOH |
| Maghemite γ-Fe2O3 | Lepidocrocite γ-FeOOH |
| ε-Fe2O3 | Bernalite Fe(OH)3 |
| Magnetite Fe3O4 (Fe2+O4) | Akaganéite β-FeOOH |
| Wüstite FeO | δ-FeOOH |
| Feroxyhyte δ′-FeOOH | |
| High pressure FeOOH (laboratory compound) | |
| Fe(OH)2 | |
| Schwertmannite Fe16O16(OH)y(SO4)z·nH2O | |
| Green rusts: (OH)3x+2y−z()z; = ;S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Chen, J.; Yuan, S.; Chen, S.; Hu, Y.; Zheng, Y.; Li, D. Iron Plaque: A Shield against Soil Contamination and Key to Sustainable Agriculture. Plants 2024, 13, 1476. https://doi.org/10.3390/plants13111476
He Z, Chen J, Yuan S, Chen S, Hu Y, Zheng Y, Li D. Iron Plaque: A Shield against Soil Contamination and Key to Sustainable Agriculture. Plants. 2024; 13(11):1476. https://doi.org/10.3390/plants13111476
Chicago/Turabian StyleHe, Zeping, Jinyuan Chen, Shilin Yuan, Sha Chen, Yuanyi Hu, Yi Zheng, and Ding Li. 2024. "Iron Plaque: A Shield against Soil Contamination and Key to Sustainable Agriculture" Plants 13, no. 11: 1476. https://doi.org/10.3390/plants13111476
APA StyleHe, Z., Chen, J., Yuan, S., Chen, S., Hu, Y., Zheng, Y., & Li, D. (2024). Iron Plaque: A Shield against Soil Contamination and Key to Sustainable Agriculture. Plants, 13(11), 1476. https://doi.org/10.3390/plants13111476