
Interspecific Differences in Carbon and Nitrogen Metabolism and Leaf Epiphytic Bacteria among Three Submerged Macrophytes in Response to Elevated Ammonia Nitrogen Concentrations

Abstract
1. Introduction
2. Materials and Methods
2.1. Precultivation of Three Submerged Macrophytes
2.2. Experimental Design
2.3. Measurements of Aquatic Parameters, FAAs and SCs
2.4. Analysis of Epiphytic Bacteria
2.5. Statistical Analysis
3. Results
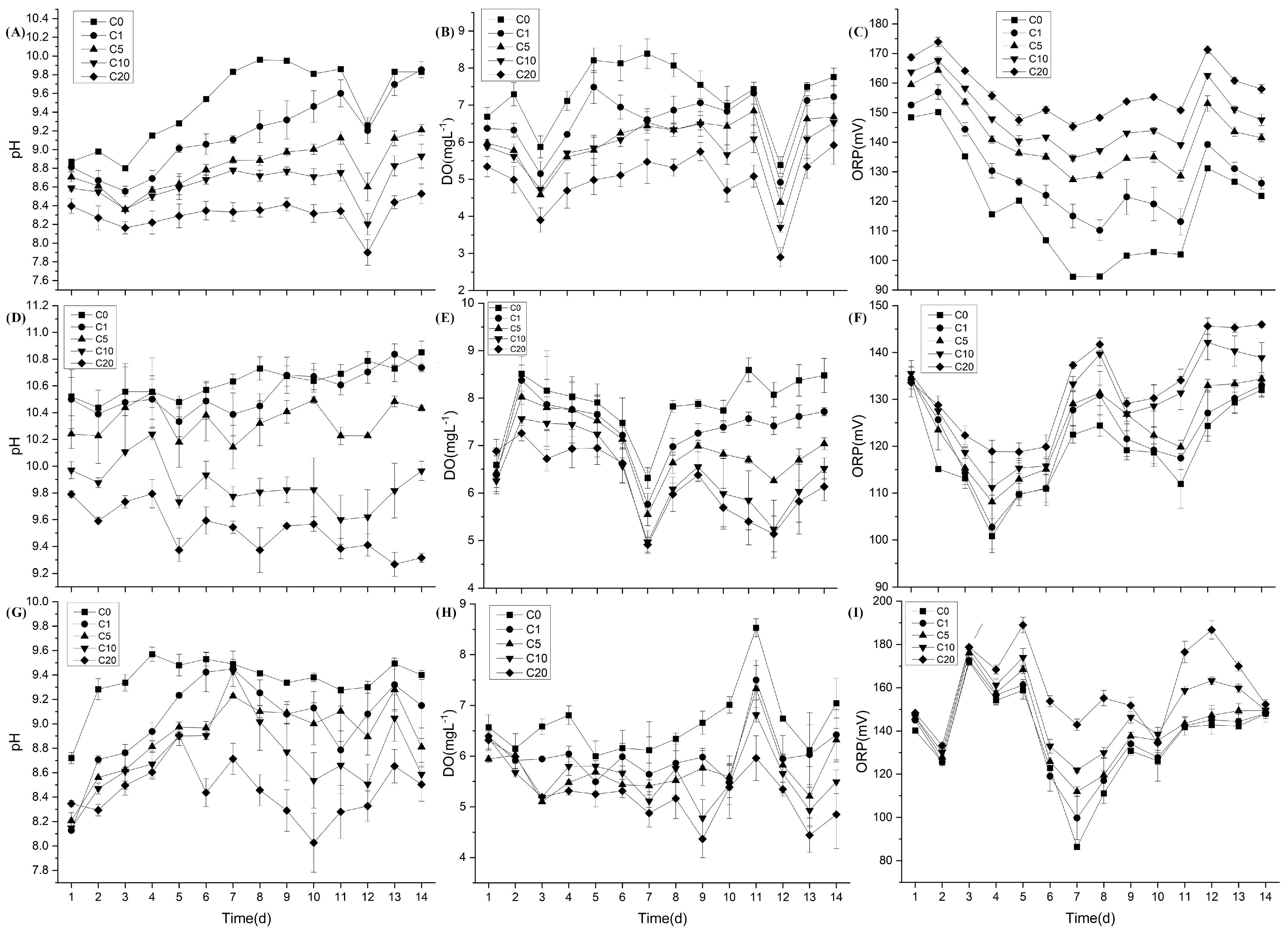
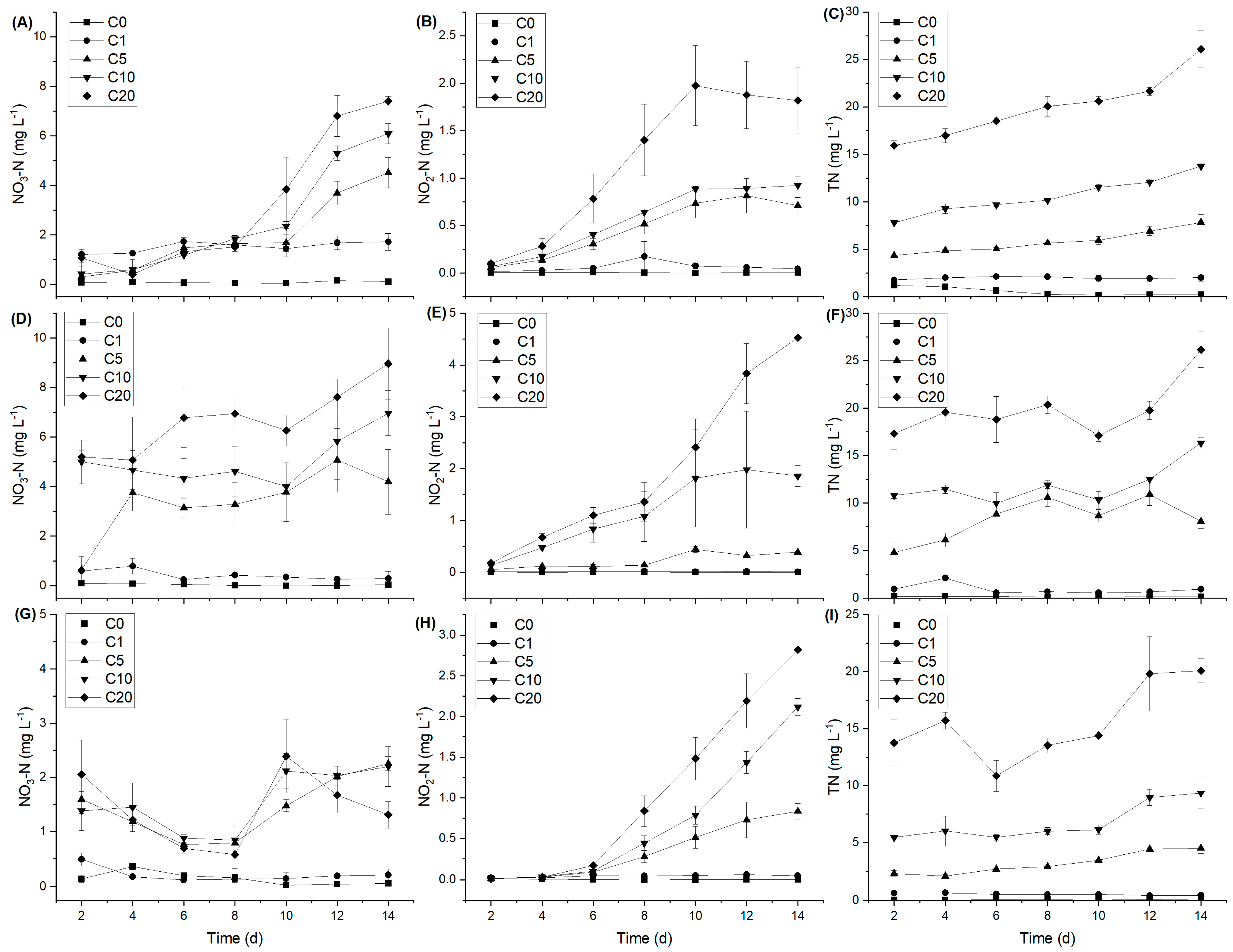
3.1. Changes in Aquatic Environmental Factors
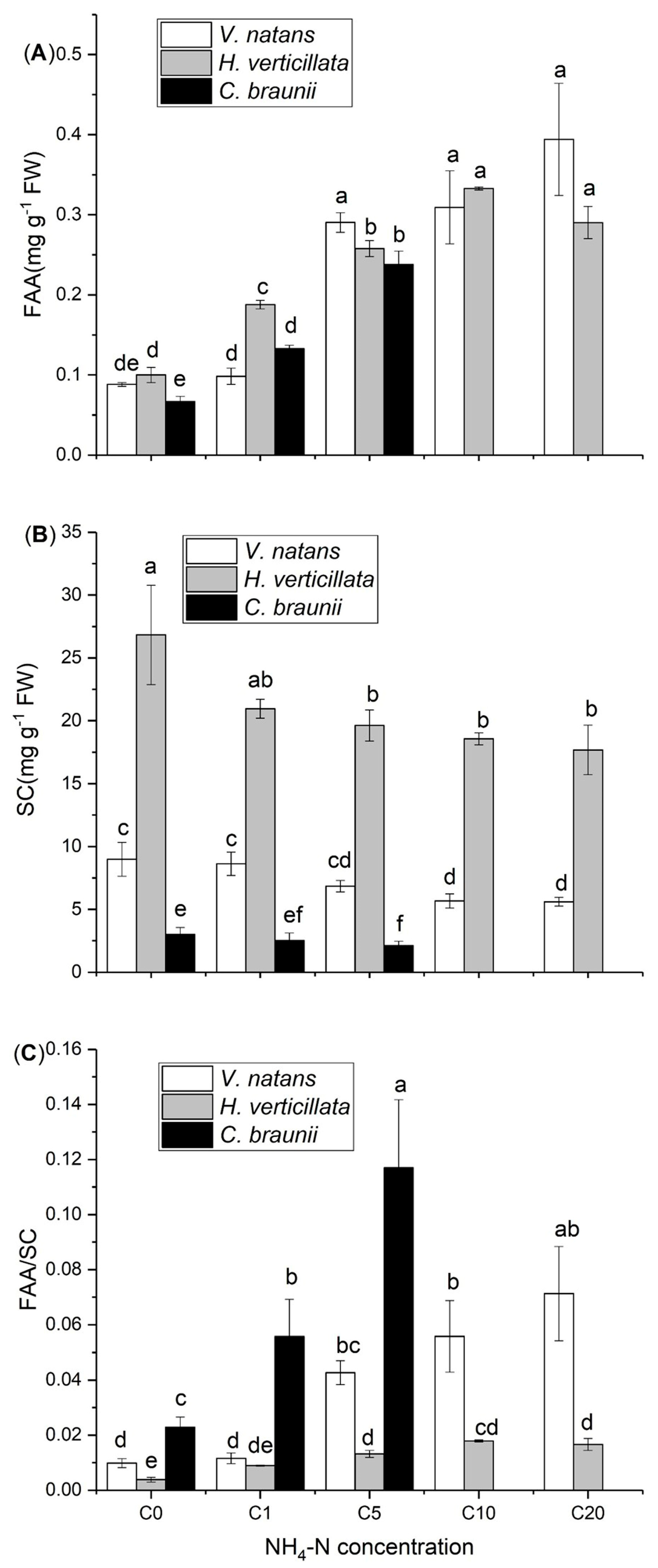
3.2. Interspecific Differences between FAAs and SCs
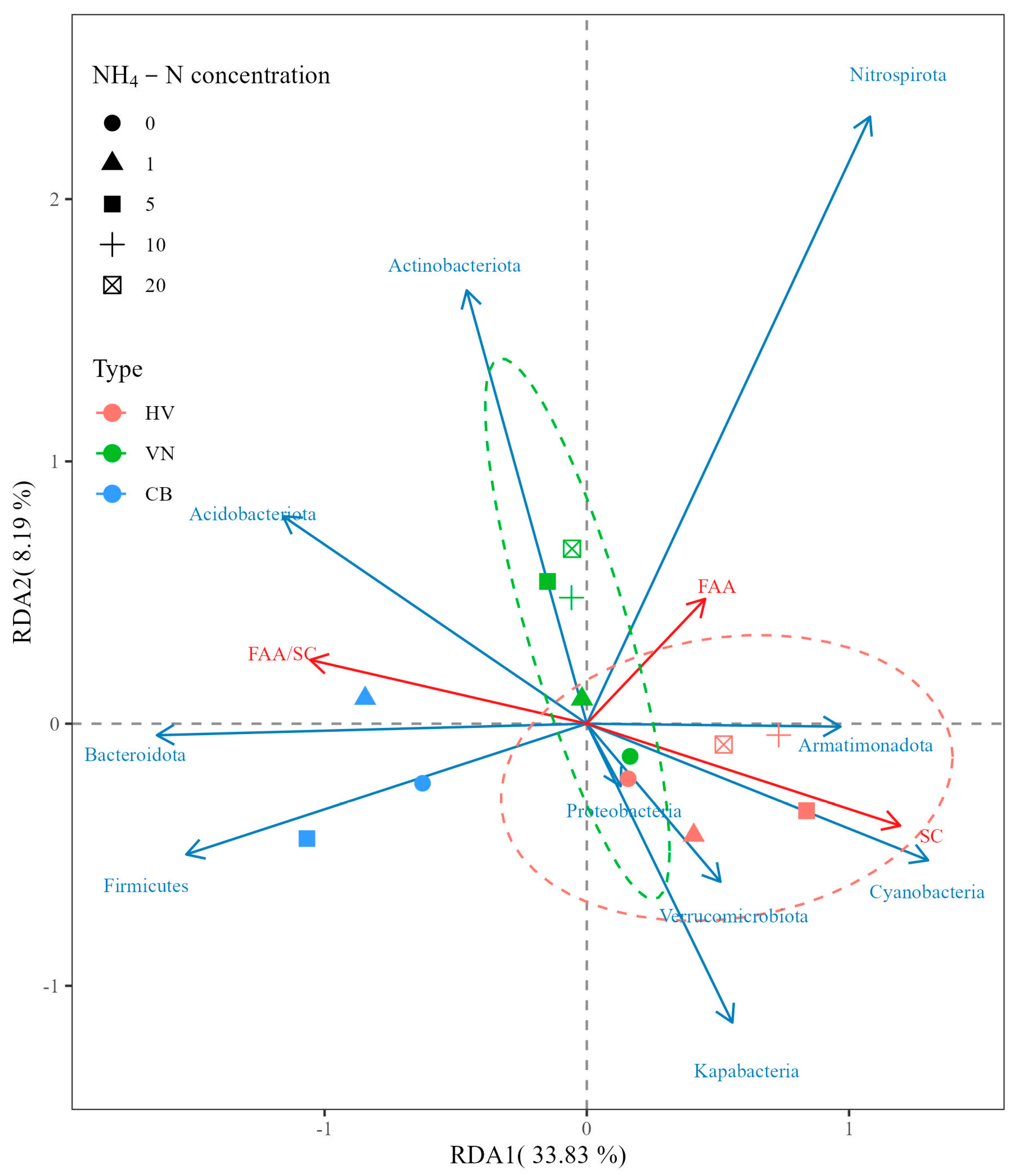
3.3. Correlations between Various Physical and Chemical Factors in Water and Leaf Epiphytic Bacterial Phyla
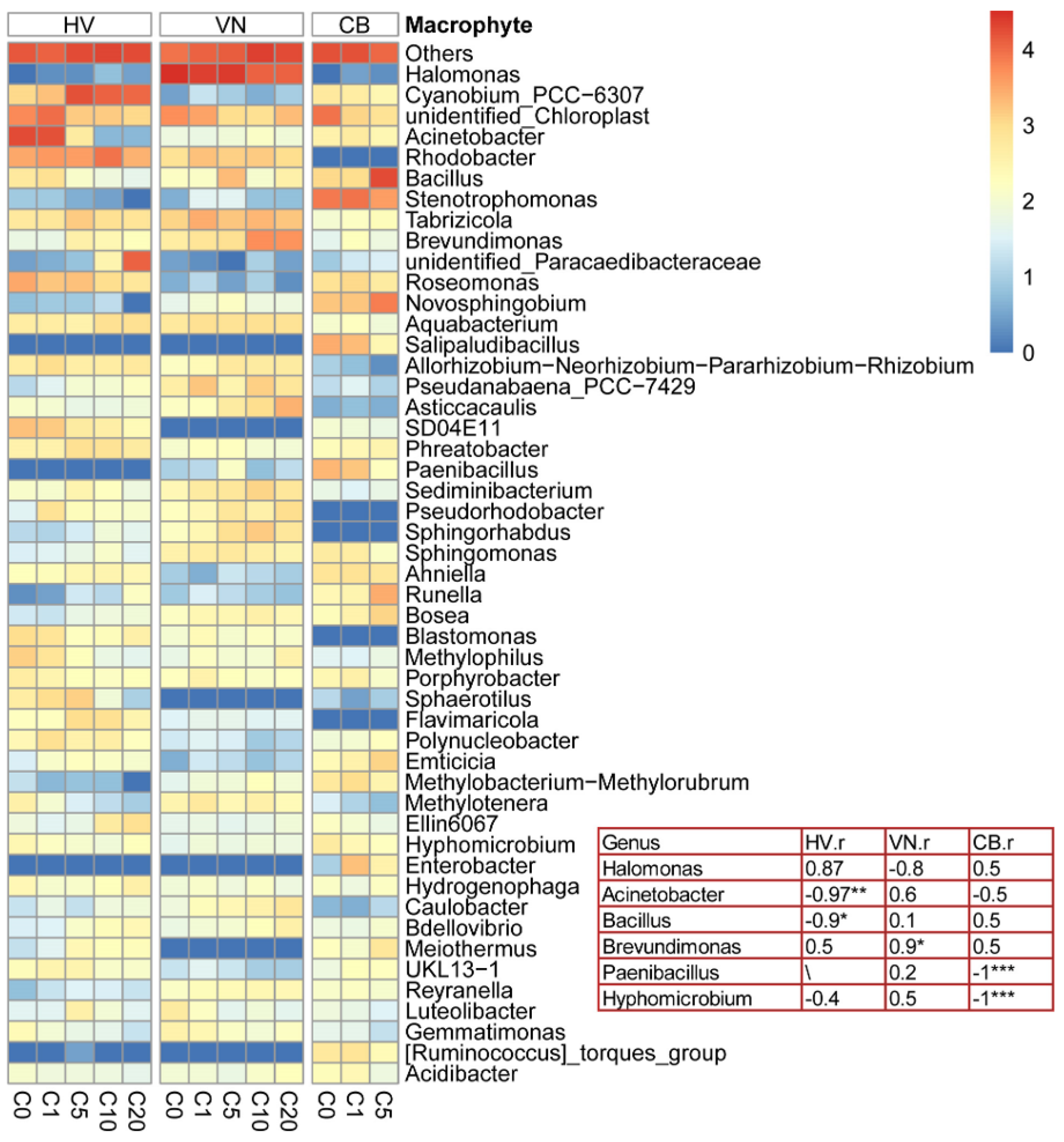
3.4. Interspecific Differences in the Response of Leaf Epiphytic Bacteria to Elevated NH4-N Concentrations
4. Discussion
4.1. Interspecific Differences in the FAA and SC Contents in Submerged Macrophytes in Response to Elevated NH4-N Concentrations
4.2. Interspecific Differences in the Response of Leaf Epiphytic Bacteria to Elevated NH4-N Concentrations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, S.M.; Wang, X.L.; Qiao, B.; Li, J.S.; Tu, J.M. First flush characteristics of rainfall runoff from a paddy field in the Taihu Lake watershed, China. Environ. Sci. Pollut. Res. 2017, 24, 8336–8351. [Google Scholar] [CrossRef]
- Li, C.; Zwiers, F.; Zhang, X.B.; Chen, G.; Lu, J.; Li, G.L.; Norris, J.; Tan, Y.H.; Sun, Y.; Liu, M. Larger increases in more extreme local precipitation events as climate warms. Geophys. Res. Lett. 2019, 46, 6885–6891. [Google Scholar] [CrossRef]
- Fong, C.R.; Gaynus, C.J.; Carpenter, R.C. Extreme rainfall events pulse substantial nutrients and sediments from terrestrial to nearshore coastal communities: A case study from French Polynesia. Sci. Rep. 2020, 10, 2955. [Google Scholar] [CrossRef]
- Brown, R.F.; Sala, O.E.; Sinsabaugh, R.L.; Collins, S.L. Temporal effects of monsoon rainfall pulses on plant available nitrogen in a Chihuahuan Desert grassland. J. Geophys. Res.-Biogeosci. 2022, 127, e2022JG006938. [Google Scholar] [CrossRef]
- Yuan, G.X.; Tan, X.Y.; Guo, P.Q.; Xing, K.; Chen, Z.L.; Li, D.B.; Yu, S.Z.; Peng, H.; Li, W.; Fu, H. Linking trait network to growth performance of submerged macrophytes in response to ammonium pulse. Water Res. 2023, 229, 119403. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Barker, A.V.; Maynard, D.N.; Mioduchowska, B.; Buch, A. Ammonium and salt inhibition of some physiological processes associated with seed germination. Physiol. Plant. 1970, 23, 898–907. [Google Scholar] [CrossRef]
- Westwood, J.H.; Foy, C.L. Influence of nitrogen on germination and early development of broomrape (Orobanche spp.). Weed Sci. 1999, 47, 2–7. [Google Scholar] [CrossRef]
- Cao, T.; Xie, P.; Ni, L.Y.; Wu, A.P.; Zhang, M.; Wu, S.K.; Smolders, A.J.P. The role of NH4+ toxicity in the decline of the submersed macrophyte Vallisneria natans in lakes of the Yangtze River basin, China. Mar. Freshw. Res. 2007, 58, 581–587. [Google Scholar] [CrossRef]
- Smolders, A.J.P.; Den Hartog, C.; Van Gestel, C.B.L.; Roelofs, J.G.M. The effects of ammonium on growth, accumulation of free amino acids and nutritional status of young phosphorus deficient Stratiotes aloides plants. Aquat. Bot. 1996, 53, 85–96. [Google Scholar] [CrossRef]
- Cao, T.; Ni, L.Y.; Xie, P. Acute biochemical responses of a submersed macrophyte, Potamogeton crispus L., to high ammonium in an aquarium experiment. J. Freshw. Ecol. 2004, 19, 279–284. [Google Scholar] [CrossRef]
- Cao, T.; Xie, P.; Li, Z.Q.; Ni, L.Y.; Zhang, M.; Xu, J. Physiological Stress of High NH4+ Concentration in Water Column on the Submersed Macrophyte Vallisneria natans L. Bull. Environ. Contam. Tox. 2009, 82, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Xie, P.; Ni, L.Y.; Zhang, M.; Xu, J. Carbon and nitrogen metabolism of an eutrophication tolerative macrophyte, Potamogeton crispus, under NH4+ stress and low light availability. Environ. Exp. Bot. 2009, 66, 74–78. [Google Scholar] [CrossRef]
- Wang, H.Y.; Ni, L.Y.; Xie, P. The mitigating effect of calcification-dependent of utilization of inorganic carbon of Chara vulgaris Linn on NH4-N toxicity. Chemosphere 2013, 93, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Chen, K.; Chen, Q.; Wang, H.Y. Distinguished responses of curled pondweed (Potamogeton crispus L.) and hornwort (Ceratophyllum demersum L.) to NH4-N stress under elevated HCO3- conditions. Appl. Ecol. Environ. Res. 2022, 20, 4737–4748. [Google Scholar] [CrossRef]
- Cao, T.; Ni, L.; Xie, P.; Xu, J.; Zhang, M. Effects of moderate ammonium enrichment on three submersed macrophytes under contrasting light availability. Freshw. Biol. 2011, 56, 1620–1629. [Google Scholar] [CrossRef]
- Gao, J.Q.; Ren, P.; Zhou, Q.Y.; Zhang, J.S. Comparative studies of the response of sensitive and tolerant submerged macrophytes to high ammonium concentration stress. Aquat. Toxicol. 2019, 211, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.X.; Cao, T.; Fu, H.; Ni, L.Y.; Zhang, X.L.; Li, W.; Song, X.; Xie, P.; Jeppesen, E. Linking carbon and nitrogen metabolism to depth distribution of submersed macrophytes using high ammonium dosing tests and a lake survey. Freshw. Biol. 2013, 58, 2532–2540. [Google Scholar] [CrossRef]
- Yuan, G.X.; Fu, H.; Zhong, J.Y.; Cao, T.; Ni, L.Y.; Zhu, T.S.; Li, W.; Song, X. Nitrogen/carbon metabolism in response to NH4+ pulse for two submersed macrophytes. Aquat. Bot. 2015, 121, 76–82. [Google Scholar] [CrossRef]
- Castiblanco, L.F.; Sundin, G.W. New insights on molecular regulation of biofilm formation in plant-associated bacteria. J. Integr. Plant Biol. 2016, 58, 362–372. [Google Scholar] [CrossRef]
- Xia, P.H.; Yan, D.B.; Sun, R.G.; Song, X.; Lin, T.; Yi, Y. Community composition and correlations between bacteria and algae within epiphytic biofilms on submerged macrophytes in a plateau lake, southwest China. Sci. Total Environ. 2020, 727, 138398. [Google Scholar] [CrossRef] [PubMed]
- Wijewardene, L.; Wu, N.C.; Fohrer, N.; Riis, T. Epiphytic biofilms in freshwater and interactions with macrophytes: Current understanding and future directions. Aquat. Bot. 2022, 176, 103467. [Google Scholar] [CrossRef]
- Jones, J.I.; Young, J.O.; Eaton, J.W.; Moss, B. The influence of nutrient loading, dissolved inorganic carbon and higher trophic levels on the interaction between submerged plants and periphyton. J. Ecol. 2002, 90, 12–24. [Google Scholar] [CrossRef]
- Song, Y.Z.; Wang, J.Q.; Gao, Y.X. Effects of epiphytic algae on biomass and physiology of Myriophyllum spicatum L. with the increase of nitrogen and phosphorus availability in the water body. Environ. Sci. Pollut. Res. 2017, 24, 9548–9555. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Li, T.L.; Yang, L.Z. Mechanisms of removing pollutants from aqueous solutions by microorganisms and their aggregates: A review. Bioresour. Technol. 2012, 107, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.G.; Fang, Y.; Jin, Y.L.; Huang, J.; Ma, X.R.; He, K.Z.; He, Z.M.; Wang, F.; Zhao, H. Microbial community and removal of nitrogen via the addition of a carrier in a pilot-scale duckweed-based wastewater treatment system. Bioresour. Technol. 2015, 179, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Wu, N.C.; Manolaki, P.; Baattrup-Pedersen, A.; Riis, T. Short-period hydrological regimes override physico-chemical variables in shaping stream diatom traits, biomass and biofilm community functions. Sci. Total Environ. 2020, 743, 140720. [Google Scholar] [CrossRef]
- O’Hare, M.T.; Baattrup-Pedersen, A.; Baumgarte, I.; Freeman, A.; Gunn, I.D.M.; Lázár, A.N.; Sinclair, R.; Wade, A.J.; Bowes, M.J. Responses of aquatic plants to eutrophication in rivers: A revised conceptual model. Front. Plant Sci. 2018, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Sultana, M.; Asaeda, T.; Ekram Azim, M.; Fujino, T. Morphological responses of a submerged macrophyte to epiphyton. Aquat. Ecol. 2010, 44, 73–81. [Google Scholar] [CrossRef]
- Gong, L.X.; Zhang, S.H.; Chen, D.Q.; Liu, K.H.; Lu, J. Response of biofilms-leaves of two submerged macrophytes to high ammonium. Chemosphere 2018, 192, 152–160. [Google Scholar] [CrossRef]
- Yan, L.L.; Mu, X.Y.; Han, B.; Zhang, S.H.; Qiu, C.H.; Ohore, O.E. Ammonium loading disturbed the microbial food webs in biofilms attached to submersed macrophyte Vallisneria natans. Sci. Total Environ. 2019, 659, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.Y.; Zhang, S.H.; Lin, D.; Guo, C.; Yan, L.L.; Wang, S.P.; He, Z.L. Nitrogen loading affects microbes, nitrifiers and denitrifiers attached to submerged macrophyte in constructed wetlands. Sci. Total Environ. 2018, 622, 121–126. [Google Scholar] [CrossRef]
- Calheiros, C.S.C.; Teixeira, A.; Pires, C.; Franco, A.R.; Duque, A.F.; Crispim, L.F.C.; Moura, S.C.; Castro, P.M.L. Bacterial community dynamics in horizontal flow constructed wetlands with different plants for high salinity industrial wastewater polishing. Water Res. 2010, 44, 5032–5038. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Ren, L.J.; Wu, Q.L. Growing season drives the compositional changes and assembly processes of epiphytic bacterial communities of two submerged macrophytes in Taihu Lake. FEMS Microbiol. Ecol. 2020, 96, fiaa025. [Google Scholar] [CrossRef] [PubMed]
- Hempel, M.; Blume, M.; Blindow, I.; Gross, E.M. Epiphytic bacterial community composition on two common submerged macrophytes in brackish water and freshwater. BMC Microbiol. 2008, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Levi, P.S.; Starnawski, P.; Poulsen, B.; Baattrup-Pedersen, A.; Schramm, A.; Riis, T. Microbial community diversity and composition varies with habitat characteristics and biofilm function in macrophyte-rich streams. Oikos 2017, 126, 398–409. [Google Scholar] [CrossRef]
- Lu, S.J.; He, R.J.; Zhao, D.Y.; Zeng, J.; Huang, X.L.; Li, K.Y.; Yu, Z.B.; Wu, Q.L. Effects of shading levels on the composition and co-occurrence patterns of bacterioplankton and epibiotic bacterial communities of Cabomba caroliniana. Sci. Total Environ. 2021, 785, 147286. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.L.; Gao, G.; Tang, X.M.; Dong, B.L.; Dai, J.Y.; Chen, D.; Song, Y.Z. The response of epiphytic microbes to habitat and growth status of Potamogeton malaianus Miq. in Lake Taihu. J. Basic Microb. 2013, 53, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, N.; Giorgi, A.; Feijoó, C. Effects of macrophyte architecture and leaf shape complexity on structural parameters of the epiphytic algal community in a Pampean stream. Aquat. Ecol. 2013, 47, 389–401. [Google Scholar] [CrossRef]
- Han, B.; Zhang, S.H.; Zhang, L.S.; Liu, K.H.; Yan, L.Y.; Wang, P.F.; Wang, C.; Pang, S. Characterization of microbes and denitrifiers attached to two species of floating plants in the wetlands of Lake Taihu. PLoS ONE 2018, 13, e0207443. [Google Scholar] [CrossRef]
- He, D.; Zheng, J.W.; Ren, L.J.; Wu, Q.L. Substrate type and plant phenolics influence epiphytic bacterial assembly during short-term succession. Sci. Total Environ. 2021, 792, 148410. [Google Scholar] [CrossRef] [PubMed]
- Hempel, M.; Grossart, H.-P.; Gross, E.M. Community composition of bacterial biofilms on two submerged macrophytes and an artificial substrate in a pre-alpine lake. Aquat. Microb. Ecol. 2009, 58, 79–94. [Google Scholar] [CrossRef]
- Wolters, J.W.; Reitsema, R.E.; Verdonschot, R.C.M.; Schoelynck, J.; Verdonschot, P.F.M.; Meire, P. Macrophyte-specific effects on epiphyton quality and quantity and resulting effects on grazing macroinvertebrates. Freshw. Biol. 2019, 64, 1131–1142. [Google Scholar] [CrossRef]
- He, D.; Ren, L.J.; Wu, Q.L. Epiphytic bacterial communities on two common submerged macrophytes in Taihu Lake: Diversity and host-specificity. Chin. J. Oceanol. Limnol. 2012, 30, 237–247. [Google Scholar] [CrossRef]
- Chambers, P.A. Light and nutrients in the control of aquatic plant community structure. II. In situ observations. J. Ecol. 1987, 75, 621–628. [Google Scholar] [CrossRef]
- Chambers, P.A.; Kalff, J. Light and nutrients in the control of aquatic plant community structure. I. In situ experiments. J. Ecol. 1987, 75, 611–619. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington DC, USA, 2005. [Google Scholar]
- Yemm, E.W.; Cocking, E.C.; Ricketts, R.E. The determination of amino-acids with ninhydrin. Analyst 1955, 80, 209–214. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. R Package, Version 2.6-4. Vegan: Community Ecology Package; The Comprehensive R Archive Network: Vienna, Austria, 2024. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Revelle, W. R Package, Version 2.4.3; Psych: Procedures for Personality and Psychological Research; The Comprehensive R Archive Network: Vienna, Austria, 2024. [Google Scholar]
- Kolde, R. R Package 2019, Version 1.0.12. Pheatmap: Pretty Heatmaps; The Comprehensive R Archive Network: Vienna, Austria, 2019. [Google Scholar]
- Gao, J.Q.; Liu, L.N.; Ma, N.; Yang, J.; Dong, Z.K.; Zhang, J.S.; Zhang, J.L.; Cai, M. Effect of ammonia stress on carbon metabolism in tolerant aquatic plant—Myriophyllum aquaticum. Environ. Pollut. 2020, 263, 114412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cao, T.; Ni, L.Y.; Xie, P.; Li, Z.Q. Carbon, nitrogen and antioxidant enzyme responses of Potamogeton crispus to both low light and high nutrient stresses. Environ. Exp. Bot. 2010, 68, 44–50. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Z.Q.; Xu, J.; Liu, Y.Q.; Ni, L.Y.; Cao, T.; Xie, P. Ammonium, microcystins, and hypoxia of blooms in eutrophic water cause oxidative stress and C–N imbalance in submersed and floating-leaved aquatic plants in Lake Taihu, China. Chemosphere 2011, 82, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Blindow, I. Decline of charophytes during eutrophication: Comparison with angiosperms. Freshw. Biol. 1992, 28, 9–14. [Google Scholar] [CrossRef]
- Van den Berg, M.S.; Scheffer, M.; Van Nes, E.; Coops, H. Dynamics and stability of Chara sp. and Potamogeton pectinatus in a shallow lake changing in eutrophication level. Hydrobiologia 1999, 408, 335–342. [Google Scholar] [CrossRef]
- He, D.; Ren, L.J.; Wu, Q.L. Contrasting diversity of epibiotic bacteria and surrounding bacterioplankton of a common submerged macrophyte, Potamogeton crispus, in freshwater lakes. FEMS Microbiol. Ecol. 2014, 90, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Zhang, S.H.; Lv, X.Y.; Han, B.; Liu, K.H.; Qiu, C.H.; Wang, C.; Wang, P.F.; Toland, H.; He, Z.L. Characterization of bacterial community in biofilm and sediments of wetlands dominated by aquatic macrophytes. Ecol. Eng. 2016, 97, 242–250. [Google Scholar] [CrossRef]
- Li, Q.; Gu, P.; Ji, X.Y.; Li, H.M.; Zhang, J.B.; Zheng, Z. Response of submerged macrophytes and periphyton biofilm to water flow in eutrophic environment: Plant structural, physicochemical and microbial properties. Ecotoxicol. Environ. Saf. 2020, 189, 109990. [Google Scholar] [CrossRef]
- Wang, H.Y.; Hu, R.; Zhou, Y.; Chen, K.; Chen, H.-Q.; Chang, F.Y. The responses of two submerged plants and their leaf epiphytic bacterial community to the addition of calcium and phosphorus. J. Hydroecol. 2024. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Kits, K.D.; Sedlacek, C.J.; Lebedeva, E.V.; Han, P.; Bulaev, A.; Pjevac, P.; Daebeler, A.; Romano, S.; Albertsen, M.; Stein, L.Y.; et al. Kinetic analysis of a complete nitrifier reveals an oligotrophic lifestyle. Nature 2017, 549, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, U.; Schumann, P.; Stieglmeier, M.; Pukall, R.; Augustin, J.; Spröer, C.; Schwendner, P.; Moissl-Eichinger, C.; Ulrich, A. Characterization of heterotrophic nitrifying bacteria with respiratory ammonification and denitrification activity–description of Paenibacillus uliginis sp. nov., an inhabitant of fen peat soil and Paenibacillus purispatii sp. nov., isolated from a spacecraft assembly clean room. Syst. Appl. Microbiol. 2010, 33, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Lindell, A.H.; Tuckfield, R.C.; McArthur, J.V. Differences in the effect of coal pile runoff (low pH, high metal concentrations) versus natural Carolina bay water (low pH, low metal concentrations) on plant condition and associated bacterial epiphytes of Salvinia minima. Bull. Environ. Contam. Toxicol. 2016, 96, 602–607. [Google Scholar] [CrossRef]
- Urakami, T.; Sasaki, J.; Suzuki, K.-I.; Komagata, K. Characterization and description of Hyphomicrobium denitrificans sp. nov. Int. J. Syst. Evol. Microbiol. 1995, 45, 528–532. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Species (S) | NH4-N (N) | S × N | Error |
|---|---|---|---|---|
| FAA | 0.47 ns | 85.71 *** | 7.96 *** | 5.85 |
| SC | 90.58 *** | 3.87 *** | 2.82 ** | 2.74 |
| FAA/SC | 42.55 *** | 31.91 *** | 17.02 *** | 8.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Chen, K.; Jin, H.; Hu, R. Interspecific Differences in Carbon and Nitrogen Metabolism and Leaf Epiphytic Bacteria among Three Submerged Macrophytes in Response to Elevated Ammonia Nitrogen Concentrations. Plants 2024, 13, 1427. https://doi.org/10.3390/plants13111427
Wang H, Chen K, Jin H, Hu R. Interspecific Differences in Carbon and Nitrogen Metabolism and Leaf Epiphytic Bacteria among Three Submerged Macrophytes in Response to Elevated Ammonia Nitrogen Concentrations. Plants. 2024; 13(11):1427. https://doi.org/10.3390/plants13111427
Chicago/Turabian StyleWang, Heyun, Kuang Chen, Hui Jin, and Rui Hu. 2024. "Interspecific Differences in Carbon and Nitrogen Metabolism and Leaf Epiphytic Bacteria among Three Submerged Macrophytes in Response to Elevated Ammonia Nitrogen Concentrations" Plants 13, no. 11: 1427. https://doi.org/10.3390/plants13111427
APA StyleWang, H., Chen, K., Jin, H., & Hu, R. (2024). Interspecific Differences in Carbon and Nitrogen Metabolism and Leaf Epiphytic Bacteria among Three Submerged Macrophytes in Response to Elevated Ammonia Nitrogen Concentrations. Plants, 13(11), 1427. https://doi.org/10.3390/plants13111427