
Enhanced Stomatal Conductance Supports Photosynthesis in Wheat to Improved NH4+ Tolerance


Abstract
:1. Introduction
2. Results
2.1. Plant Growth and Nitrogen Concentration
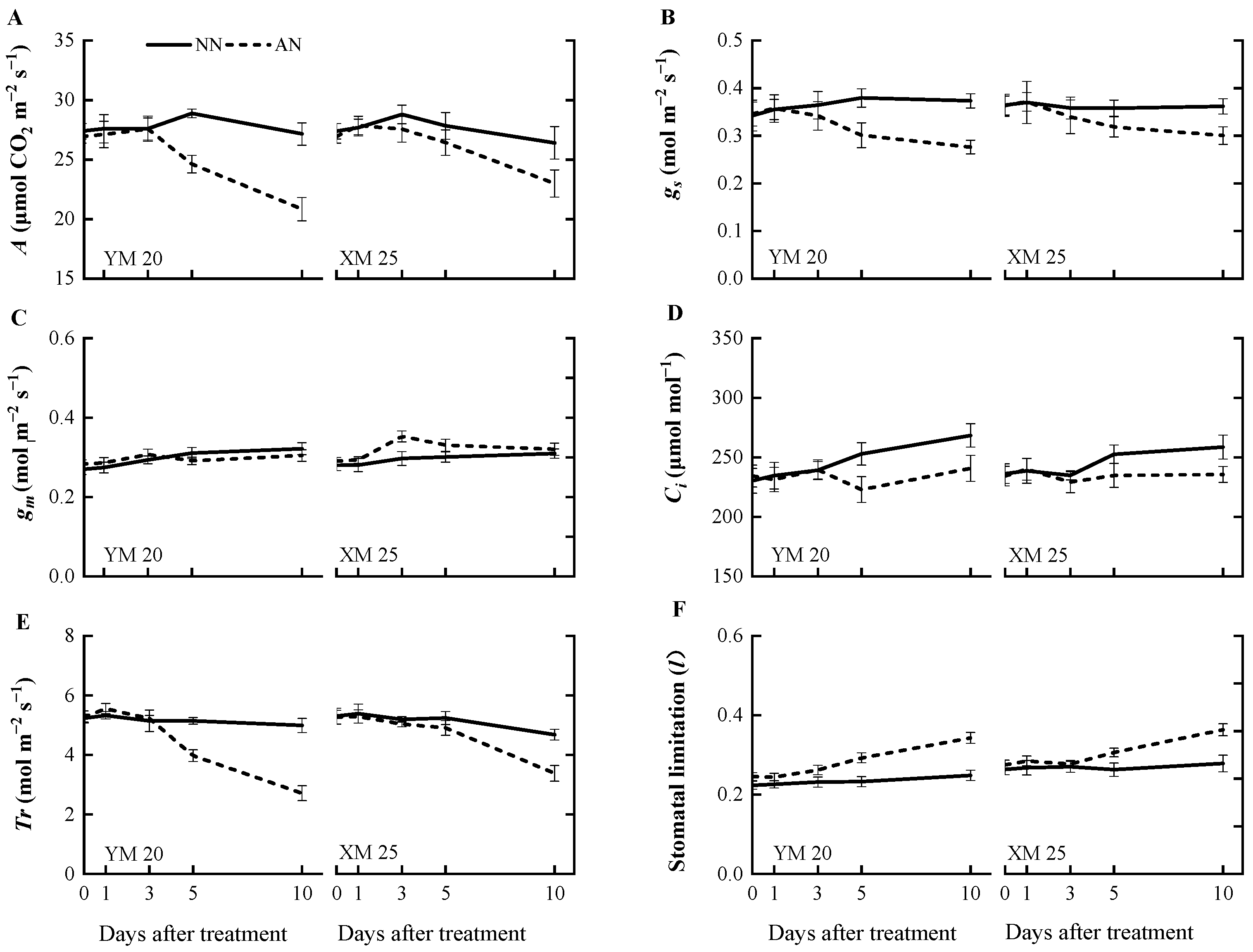
2.2. Photosynthesis and Related Attributes
2.3. Chlorophyll Content and Chlorophyll Fluorescence Parameters
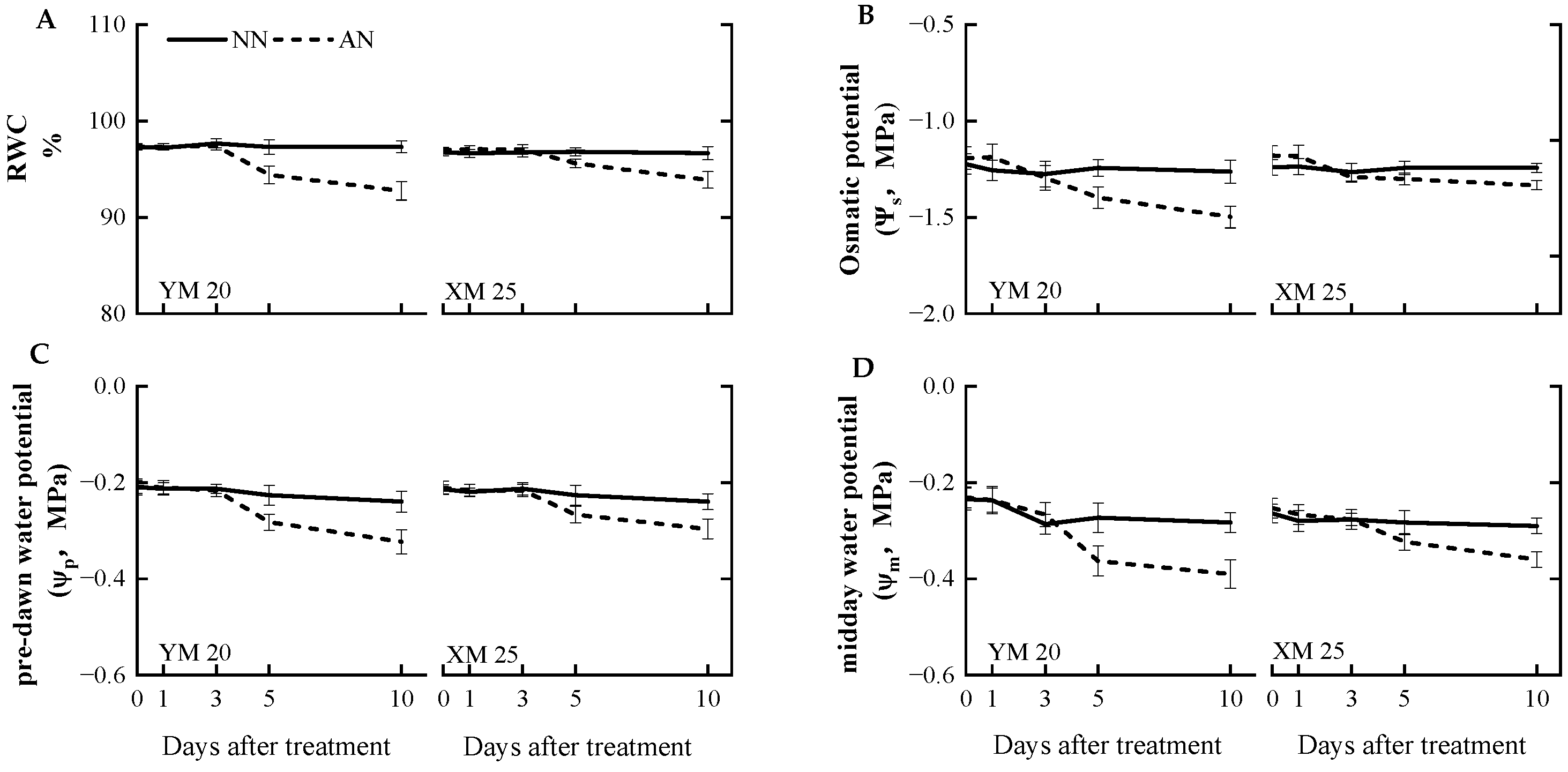
2.4. Leaf Moisture Status
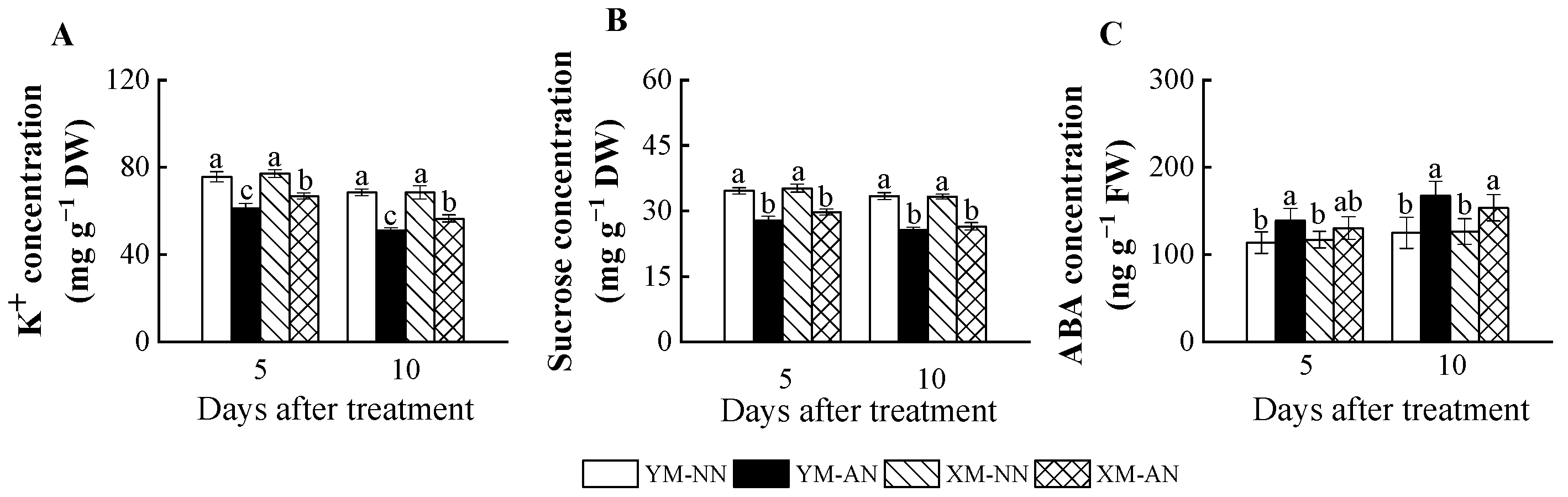
2.5. Concentration of K+, Sucrose and ABA
2.6. Relative Gene Expression
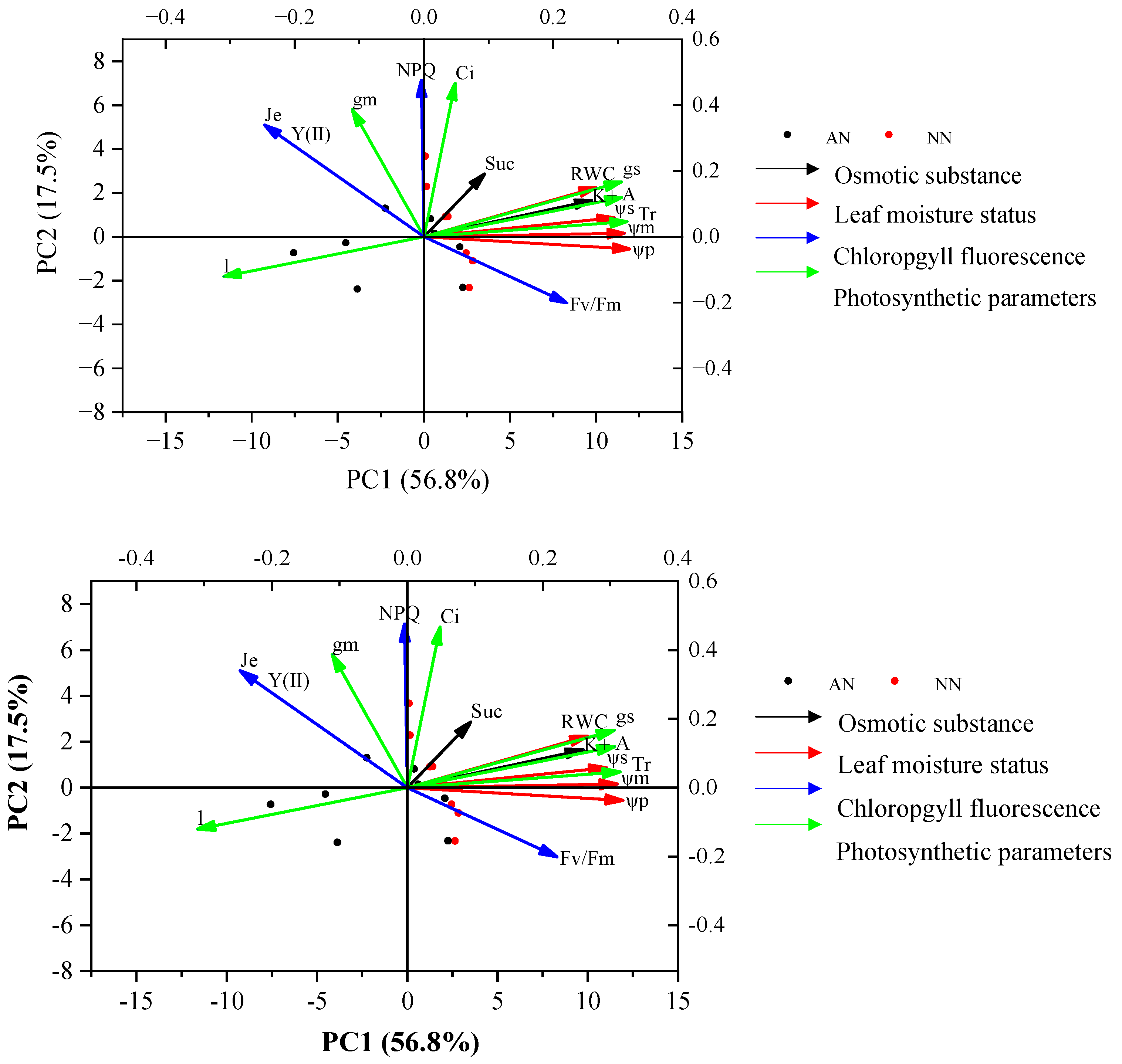
2.7. The Relationships between Photosynthetic Parameters and Physiological Traits with Nitrogen Forms
3. Discussion
3.1. Variation in Photosynthetic Response and Stomatal Limitation
3.2. Leaf Mesophyll Conductance (gm) under NH4+ Stress
3.3. Chlorophyll Concentration and Fluorescence under NH4+ Stress
3.4. Decline in Stomatal Conductance and Osmotic Potential under NH4+ Stress
3.5. Water Uptake under NH4+ Stress
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Gas Exchange Measurements
4.3. Leaf Area and Chlorophyll Fluorescence Measurements
4.4. Leaf Water Status
4.5. Determination of N, K+ and Sucrose Concentration
4.6. Determination of ABA Concentration
4.7. RT-PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; von Wirén, N. Ammonium as a signal for physiological and morphological responses in plants. J. Exp. Bot. 2017, 68, 2581. [Google Scholar] [CrossRef] [PubMed]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming ammonium toxicity. Plant Sci. 2015, 231, 184. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Moran, J.F. Can Ammonium Stress Be Positive for Plant Performance? Front. Plant Sci. 2019, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, I.; Marino, D.; Coleto, I. Arabidopsis MYB28 and MYB29 transcription factors are involved in ammonium-mediated alterations of root-system architecture. Plant Signal Behav. 2021, 16, 1879532. [Google Scholar] [CrossRef] [PubMed]
- Robert, G.; Yagyu, M.; Koizumi, T.; Naya, L.; Masclaux Daubresse, C.; Yoshimoto, K. Ammonium stress increases microautophagic activity while impairing macroautophagic flux in Arabidopsis roots. Plant J. 2021, 105, 1083. [Google Scholar] [CrossRef]
- Emily, L.; Harrison, L.A.C.J. The influence of stomatal morphology and distribution on photosynthetic gas exchange. Plant J. 2020, 101, 768. [Google Scholar]
- Cochrane, T.T.; Cochrane, T.A. Differences in the way potassium chloride and sucrose solutions effect osmotic potential of significance to stomata aperture modulation. Plant Physiol. Biochem. 2009, 47, 205. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, J.; Zhang, G.; Lu, J.; Ren, T.; Cong, R.; Lu, Z.; Zhang, Y.; Liao, S.; Li, X. Potassium deficiency limits water deficit tolerance of rice by reducing leaf water potential and stomatal area. Agric. Water Manag. 2022, 271, 107744. [Google Scholar] [CrossRef]
- Li, B.; Li, G.; Kronzucker, H.J.; Baluška, F.; Shi, W. Ammonium stress in Arabidopsis: Signaling, genetic loci, and physiological targets. Trends Plant Sci. 2014, 19, 107. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Lu, Z.; Ding, L.; Guo, J.; Wang, M.; Ling, N.; Guo, S.; Shen, Q. Role of Aquaporins in Determining Carbon and Nitrogen Status in Higher Plants. Int. J. Mol. Sci. 2018, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Naku, M.; Kambizi, L.; Matimati, I. Functional roles of ammonium (NH4+) and nitrate (NO3–) in regulation of day- and night-time transpiration in Phaseolus vulgaris. Funct. Plant Biol. 2019, 46, 806. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.D.; Hawkins, H.; Verboom, G.A. The importance of nutritional regulation of plant water flux. Oecologia 2009, 161, 15. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Gao, C.; Li, Y.; Li, Y.; Zhu, Y.; Xu, G.; Shen, Q.; Kaldenhoff, R.; Kai, L.; Guo, S. The enhanced drought tolerance of rice plants under ammonium is related to aquaporin (AQP). Plant Sci. 2015, 234, 14. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, L.; Wang, Q.; Jiao, L.; Hua, W.; Zhou, Q.; Huang, X. Photosynthesis, chlorophyll fluorescence characteristics, and chlorophyll content of soybean seedlings under combined stress of bisphenol A and cadmium. Environ. Toxicol. Chem. 2014, 33, 2455. [Google Scholar] [CrossRef] [PubMed]
- Rah, M.; Helali, S.; Nebli, H.; Kaddour, R.; Mahmoudi, H.; Lachaâl, M.; Ouerghi, Z. Influence of nitrate—Ammonium ratio on growth and nutrition of Arabidopsis thaliana. Plant Soil 2010, 336, 65. [Google Scholar]
- Sanchez-Zabala, J.; Gonzalez-Murua, C.; Marino, D. Mild ammonium stress increases chlorophyll content in Arabidopsis thaliana. Plant Signal Behav. 2015, 10, e991596. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.D.; Rasmusson, A.G.; Escobar, M.A. Distinct signalling pathways and transcriptome response signatures differentiate ammonium- and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef]
- Podgórska, A.; Gieczewska, K.; Łukawska-Kuźma, K.; Rasmusson, A.G.; Gardeström, P.; Szal, B. Long-term ammonium nutrition of Arabidopsis increases the extra chloroplastic NAD(P)H/NAD(P)+ ratio and mitochondrial reactive oxygen species level in leaves but does not impair photosynthetic capacity. Plant Cell Environ. 2013, 36, 2034. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Li, Y.; Tian, Z.; Hu, J.; Adkins, S.; Dai, T. Changes of oxidative metabolism in the roots of wheat (Triticum aestivum L.) seedlings in response to elevated ammonium concentrations. J. Integr. Agric. 2021, 20, 1216. [Google Scholar] [CrossRef]
- Ijato, T.; Porras Murillo, R.; Ganz, P.; Ludewig, U.; Neuhäuser, B. Concentration-dependent physiological and transcriptional adaptations of wheat seedlings to ammonium. Physiol. Plant. 2021, 171, 328. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Bio, A.; Martins-Loução, M.A.; Moran, J.F. Intra-specific variation in pea responses to ammonium nutrition leads to different degrees of tolerance. Environ. Exp. Bot. 2011, 70, 233. [Google Scholar] [CrossRef]
- Vega-Mas, I.; Rossi, M.T.; Gupta, K.J.; González-Murua, C.; Ratcliffe, R.G.; Estavillo, J.M.; González-Moro, M.B. Tomato roots exhibit in vivo glutamate dehydrogenase aminating capacity in response to excess ammonium supply. J. Plant Physiol. 2019, 239, 83. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, T.; Inaba, J.; Wakazaki, M.; Sato, M.; Toyooka, K.; Miyagi, A.; Kawai-Yamada, M.; Sugiura, D.; Nakagawa, T.; Kiba, T.; et al. Excessive ammonium assimilation by plastidic glutamine synthetase causes ammonium toxicity in Arabidopsis thaliana. Nat. Commun. 2021, 12, 4944. [Google Scholar] [CrossRef] [PubMed]
- Vega-Mas, I.; Cukier, C.; Coleto, I.; González-Murua, C.; Limami, A.M.; González-Moro, M.B.; Marino, D. Isotopic labelling reveals the efficient adaptation of wheat root TCA cycle flux modes to match carbon demand under ammonium nutrition. Sci. Rep. 2019, 9, 8925. [Google Scholar] [CrossRef]
- Chen, H.; Hu, W.; Wang, Y.; Zhang, P.; Zhou, Y.; Yang, L.; Li, Y.; Chen, L.; Guo, J. Declined photosynthetic nitrogen use efficiency under ammonium nutrition is related to photosynthetic electron transport chain disruption in citrus plants. Sci. Hortic. 2023, 308, 111594. [Google Scholar] [CrossRef]
- Vega-Mas, I.; Marino, D.; Sánchez-Zabala, J.; González-Murua, C.; Estavillo, J.M.; González-Moro, M.B. CO2 enrichment modulates ammonium nutrition in tomato adjusting carbon and nitrogen metabolism to stomatal conductance. Plant Sci. 2015, 241, 32. [Google Scholar] [CrossRef]
- Chen, G.; Zheng, Z.; Bai, M.; Li, Q. Chronic effects of microcystin-LR at environmental relevant concentrations on photosynthesis of Typha angustifolia Linn. Ecotoxicology 2020, 29, 514. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Caemmerer, S.V.; Berry, J.A. biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78. [Google Scholar] [CrossRef]
- Alencar, V.T.C.B.; Lobo, A.K.M.; Carvalho, F.E.L.; Silveira, J.A.G. High ammonium supply impairs photosynthetic efficiency in rice exposed to excess light. Photosynth. Res. 2019, 140, 321. [Google Scholar] [CrossRef] [PubMed]
- Torralbo, F.; González-Moro, M.B.; Baroja-Fernández, E.; Aranjuelo, I.; González-Murua, C. Differential Regulation of Stomatal Conductance as a Strategy to Cope with Ammonium Fertilizer under Ambient versus Elevated CO2. Front. Plant Sci. 2019, 10, 597. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Y.; Xiao, J.; Xu, K. Reduced photosynthetic dark reaction triggered by ABA application increases intercellular CO2 concentration, generates H2O2 and promotes closure of stomata in ginger leaves. Environ. Exp. Bot. 2015, 113, 11. [Google Scholar] [CrossRef]
- Liu, M.; Liu, X.; Du, X.; Korpelainen, H.; Niinemets, Ü.; Li, C. Anatomical variation of mesophyll conductance due to salt stress in Populus cathayana females and males growing under different inorganic nitrogen sources. Tree Physiol. 2021, 41, 1462. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ren, B.; Yang, X.; Xu, G.; Shen, Q.; Guo, S. Chloroplast Downsizing under Nitrate Nutrition Restrained Mesophyll Conductance and Photosynthesis in Rice (Oryza sativa L.) under Drought Conditions. Plant Cell Physiol. 2012, 53, 892. [Google Scholar] [CrossRef] [PubMed]
- Onoda, Y.; Wright, I.J.; Evans, J.R.; Hikosaka, K.; Kitajima, K.; Niinemets, Ü.; Poorter, H.; Tosens, T.; Westoby, M. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytol. 2017, 214, 1447. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Clemente-Moreno, M.J.; Bota, J.; Brodribb, T.J.; Gago, J.; Mizokami, Y.; Nadal, M.; Perera-Castro, A.V.; Roig-Oliver, M.; Sugiura, D.; et al. Cell wall thickness and composition are involved in photosynthetic limitation. J. Exp. Bot. 2021, 72, 3971. [Google Scholar] [CrossRef]
- Jian, S.; Liao, Q.; Song, H.; Liu, Q.; Lepo, J.E.; Guan, C.; Zhang, J.; Ismail, A.M.; Zhang, Z. NRT1.1-Related NH4+ Toxicity Is Associated with a Disturbed Balance between NH4+ Uptake and Assimilation. Plant Physiol. 2018, 178, 1473. [Google Scholar] [CrossRef]
- Mao, L.Z.; Lu, H.F.; Wang, Q.; Cai, M.M. Comparative photosynthesis characteristics of Calycanthus chinensis and Chimonanthus praecox. Photosynthetica 2007, 45, 601. [Google Scholar] [CrossRef]
- Brownlee, C. Stomatal Physiology: Cereal Successes. Curr. Biol. 2018, 28, R551. [Google Scholar] [CrossRef]
- Andrés, Z.; Pérez-Hormaeche, J.; Leidi, E.O.; Schlücking, K.; Steinhorst, L.; McLachlan, D.H.; Schumacher, K.; Hetherington, A.M.; Kudla, J.; Cubero, B.; et al. Control of vacuolar dynamics and regulation of stomatal aperture by tonoplast potassium uptake. Proc. Natl. Acad. Sci. USA 2014, 11, E1806–E1814. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A.; Szarejko, I. Open or Close the Gate—Stomata Action under the Control of Phytohormones in Drought Stress Conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, Y.; Yang, X.; Li, H.; Shen, Q.; Guo, S. Ammonium nutrition increases water absorption in rice seedlings (Oryza sativa L.) under water stress. Plant Soil 2010, 331, 193. [Google Scholar] [CrossRef]
- Kirscht, A.; Kaptan, S.S.; Bienert, G.P.; Chaumont, F.; Nissen, P.; de Groot, B.L.; Kjellbom, P.; Gourdon, P.; Johanson, U. Crystal Structure of an Ammonia-Permeable Aquaporin. PLoS Biol. 2016, 14, e1002411. [Google Scholar] [CrossRef] [PubMed]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, Y.; Xu, X.; Shen, Q.; Guo, S. Light-saturated photosynthetic rate in high-nitrogen rice (Oryza sativa L.) leaves is related to chloroplastic CO2 concentration. J. Exp. Bot. 2009, 60, 2351. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393. [Google Scholar] [CrossRef]
- Harley, P.C.; LFMG. Theoretical Considerations when Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2. Plant Physiol. 1992, 98, 1429. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Nadal, M.; Brodribb, T.J.; Fernández Marín, B.; García Plazaola, J.I.; Arzac, M.I.; López Pozo, M.; Perera Castro, A.V.; Gulías, J.; Flexas, J.; Farrant, J.M. Differences in biochemical, gas exchange and hydraulic response to water stress in desiccation tolerant and sensitive fronds of the fernAnemia caffrorum. N. Phytol. 2021, 231, 1415. [Google Scholar] [CrossRef]
- Zeng, Y.; Yu, J.; Cang, J.; Liu, L.; Mu, Y.; Wang, J.; Zhang, D. Detection of Sugar Accumulation and Expression Levels of Correlative Key Enzymes in Winter Wheat (Triticum aestivum) at Low Temperatures. Biosci. Biotechnol. Biochem. 2014, 75, 681. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Chiappetta, A.; Bruno, L.; Bitonti, M.B. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. J. Exp. Bot. 2012, 63, 695. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Sun, J.; Tian, Z.; Hu, H.; Michel, E.J.S.; Gao, J.; Jiang, D.; Cao, W.; Dai, T. Root extension and nitrate transporter up-regulation induced by nitrogen deficiency improves nitrogen status and plant growth at the seedling stage of winter wheat (Triticum aestivum L.). Environ. Exp. Bot. 2017, 141, 28. [Google Scholar] [CrossRef]
- Wu, S.; Sun, X.; Tan, Q.; Hu, C. Molybdenum improves water uptake via extensive root morphology, aquaporin expressions and increased ionic concentrations in wheat under drought stress. Environ. Exp. Bot. 2019, 157, 241. [Google Scholar] [CrossRef]
- Yang, M.; He, J.; Sun, Z.; Li, Q.; Cai, J.; Zhou, Q.; Wollenweber, B.; Jiang, D.; Wang, X. Drought priming mechanisms in wheat elucidated by in-situ determination of dynamic stomatal behavior. Front. Plant Sci. 2023, 14, 1138494. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatment | Plant Dry Weight (mg plant−1) | Plant Leaf Area (cm2 plant−1) | Leaf Nitrogen Concentration (mg g−1) | Specific Leaf Weight of 4th Leaf (mg cm−2 DW) |
|---|---|---|---|---|---|
| Yangmai20 | NN | 1266 ± 39.7 a | 100 ± 4.6 a | 69 ± 1.9 b | 0.30 ± 0.008 b |
| AN | 980 ± 31.2 c | 82 ± 5.4 c | 77 ± 2.5 a | 0.38 ± 0.017 a | |
| Xumai25 | NN | 1128 ± 50.3 b | 97 ± 4.7 ab | 65 ± 3.1 b | 0.29 ± 0.013 b |
| AN | 1033 ± 50.2 c | 86 ± 3.4 bc | 75 ± 2.2 a | 0.37 ± 0.006 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Zheng, Q.; Dong, C.; Liang, Z.; Tian, Z.; Dai, T. Enhanced Stomatal Conductance Supports Photosynthesis in Wheat to Improved NH4+ Tolerance. Plants 2024, 13, 86. https://doi.org/10.3390/plants13010086
Hu J, Zheng Q, Dong C, Liang Z, Tian Z, Dai T. Enhanced Stomatal Conductance Supports Photosynthesis in Wheat to Improved NH4+ Tolerance. Plants. 2024; 13(1):86. https://doi.org/10.3390/plants13010086
Chicago/Turabian StyleHu, Jinling, Qiaomei Zheng, Chaofeng Dong, Zhihui Liang, Zhongwei Tian, and Tingbo Dai. 2024. "Enhanced Stomatal Conductance Supports Photosynthesis in Wheat to Improved NH4+ Tolerance" Plants 13, no. 1: 86. https://doi.org/10.3390/plants13010086
APA StyleHu, J., Zheng, Q., Dong, C., Liang, Z., Tian, Z., & Dai, T. (2024). Enhanced Stomatal Conductance Supports Photosynthesis in Wheat to Improved NH4+ Tolerance. Plants, 13(1), 86. https://doi.org/10.3390/plants13010086