
The Role of the Ascorbic Acid–Glutathione Cycle in Young Wheat Ears’ Response to Spring Freezing Stress


 ,
,
Abstract
:1. Introduction
2. Results
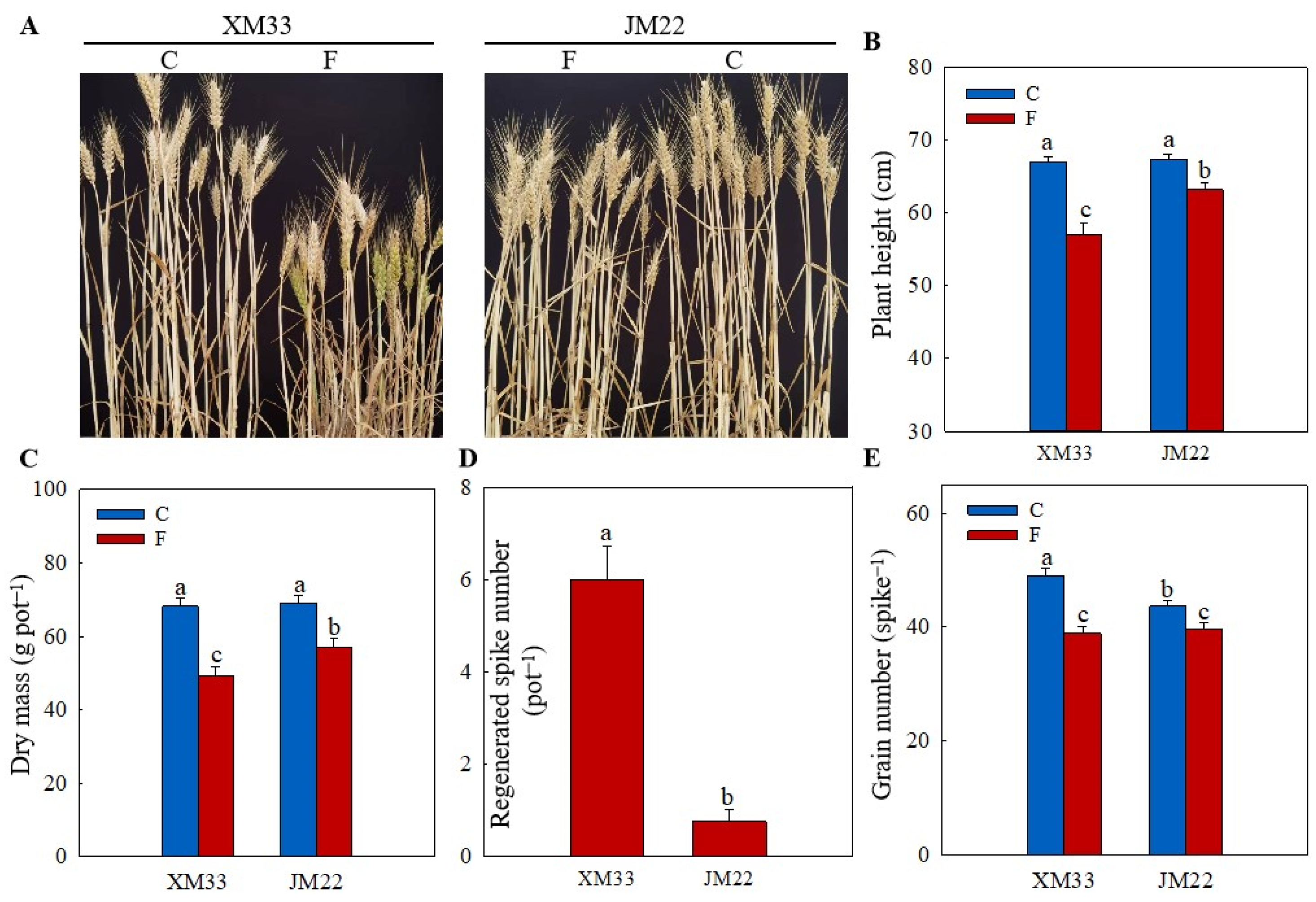
2.1. Growth Parameters
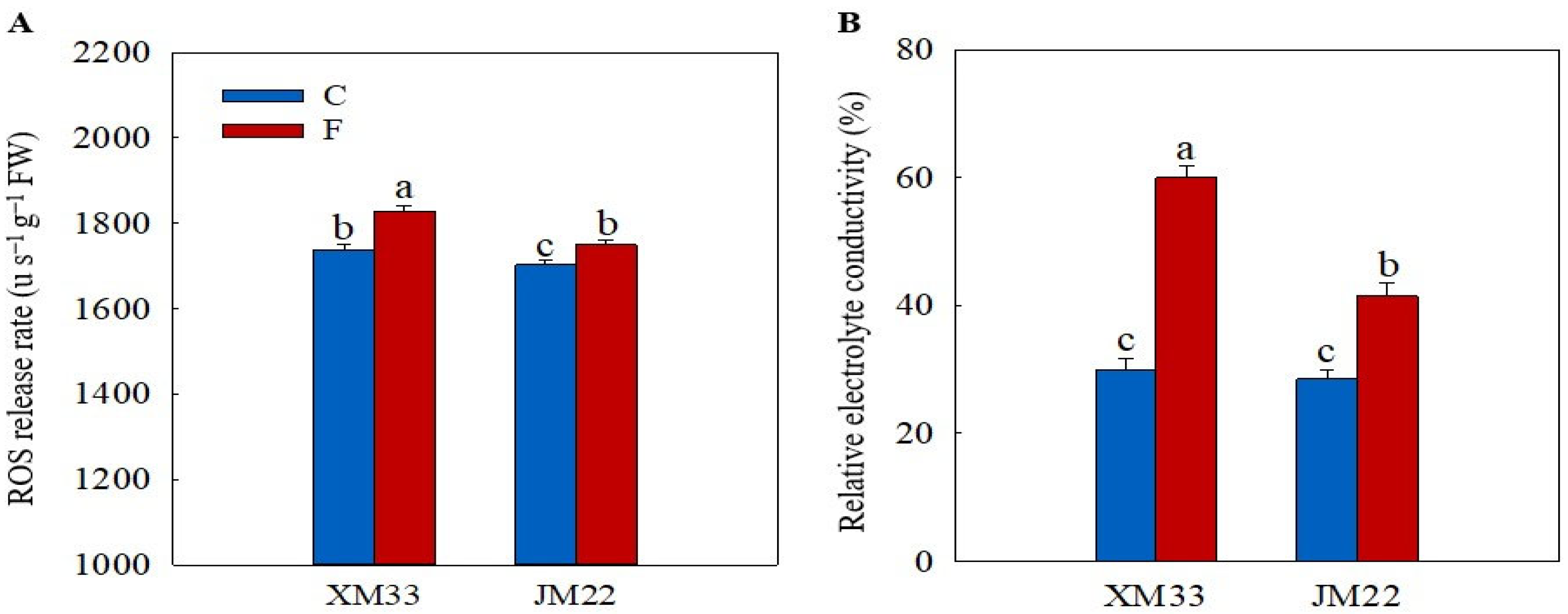
2.2. ROS Level and Electrolyte Leakage
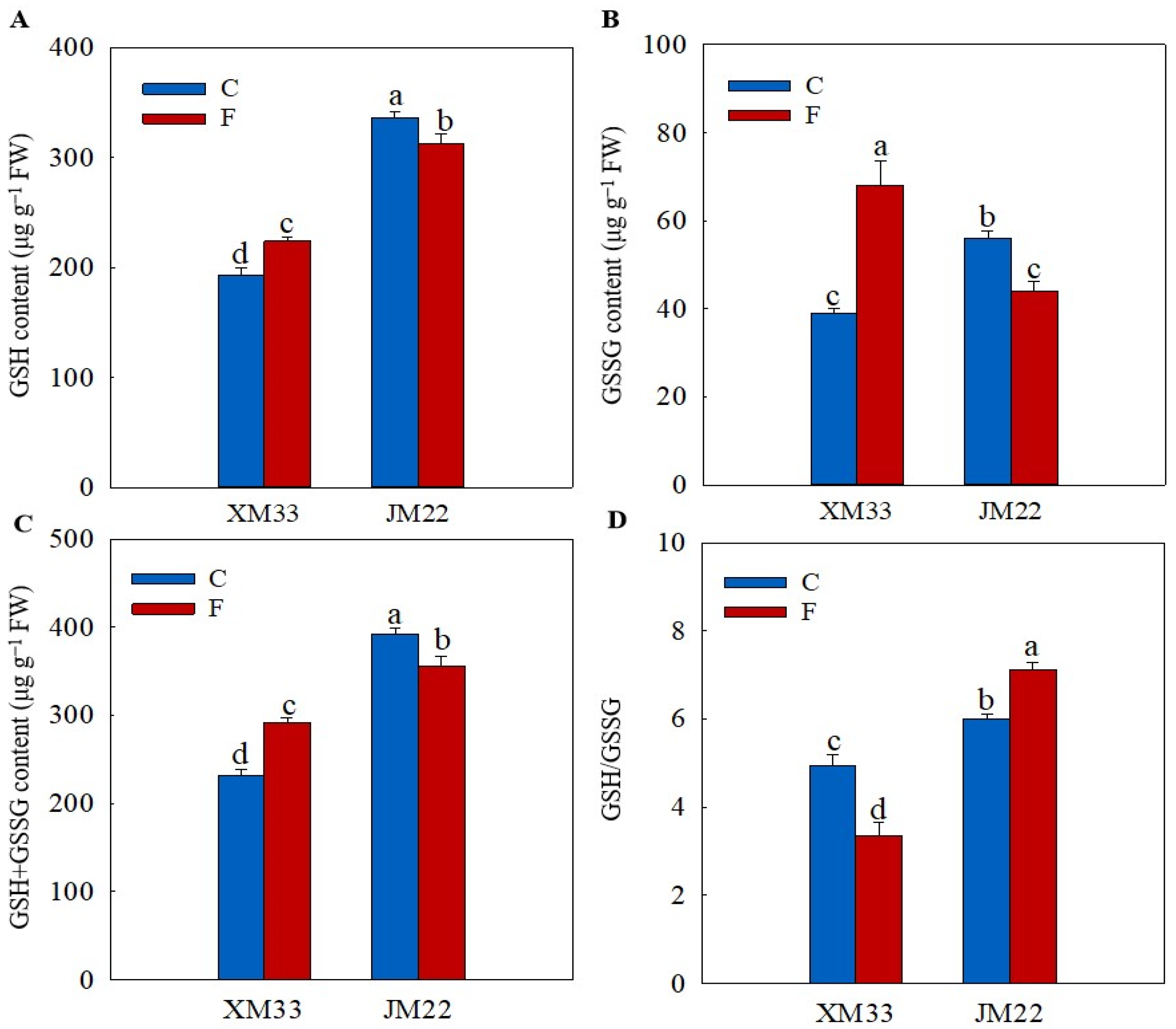
2.3. Ascorbate and Glutathione Content
2.4. AsA–GSH Metabolism Associated Enzymes Activities
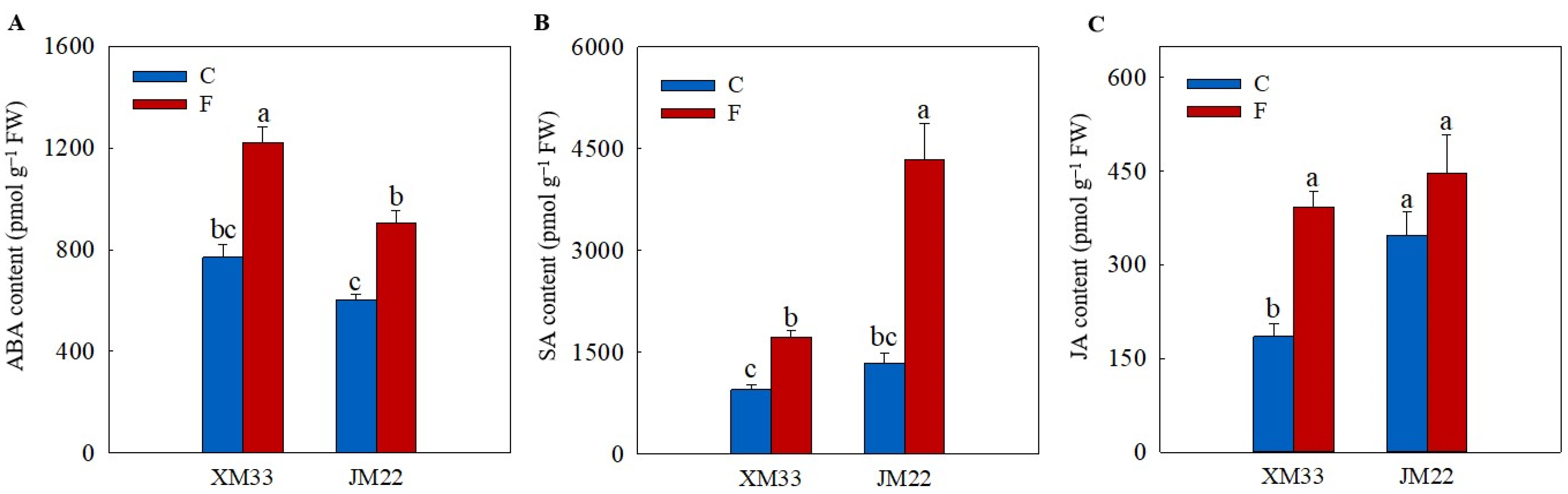
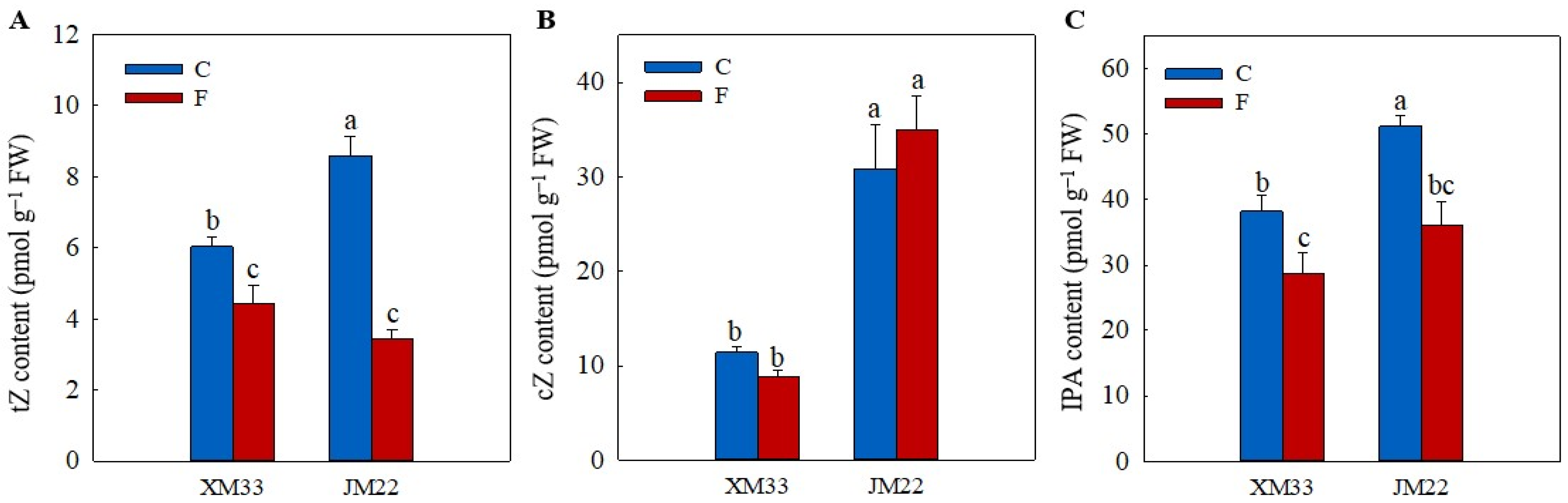
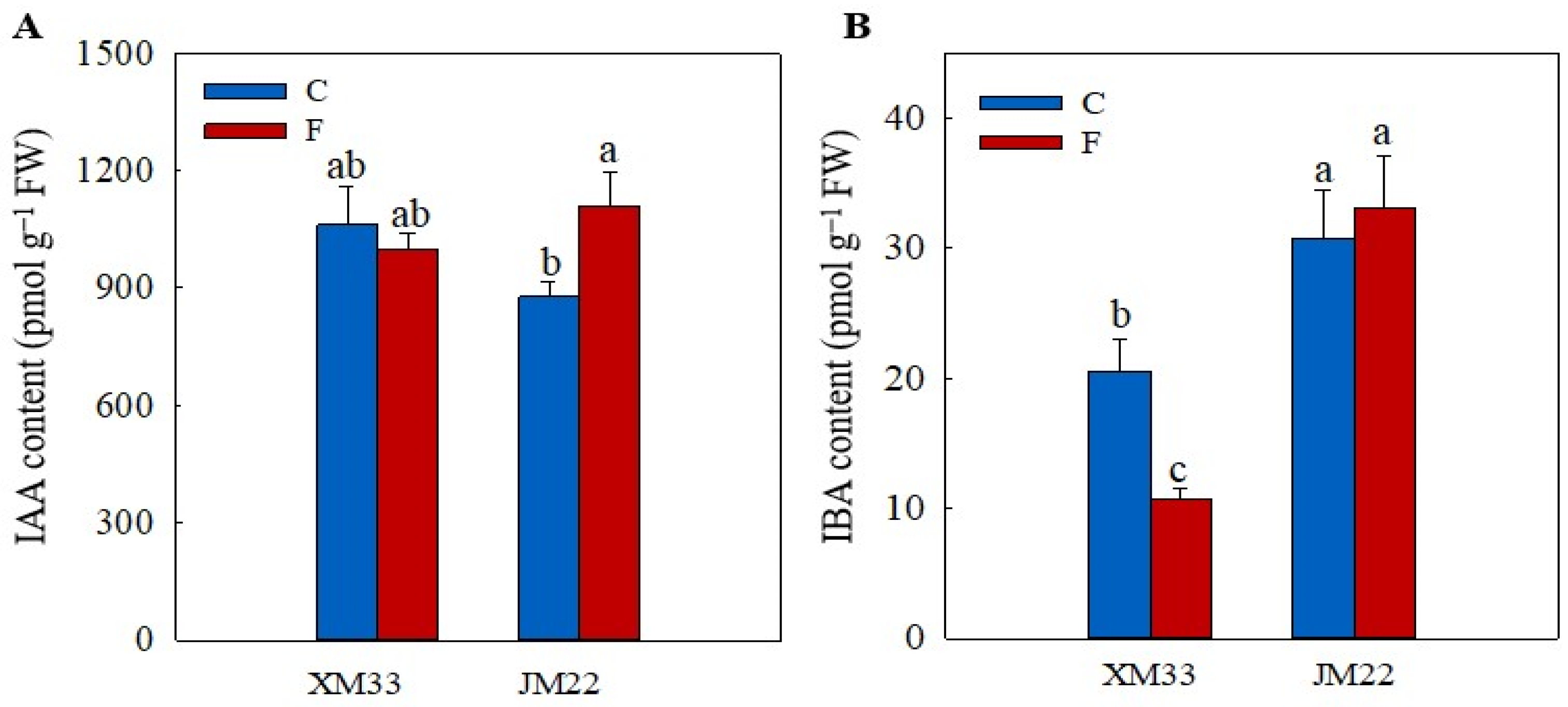
2.5. Hormones Levels
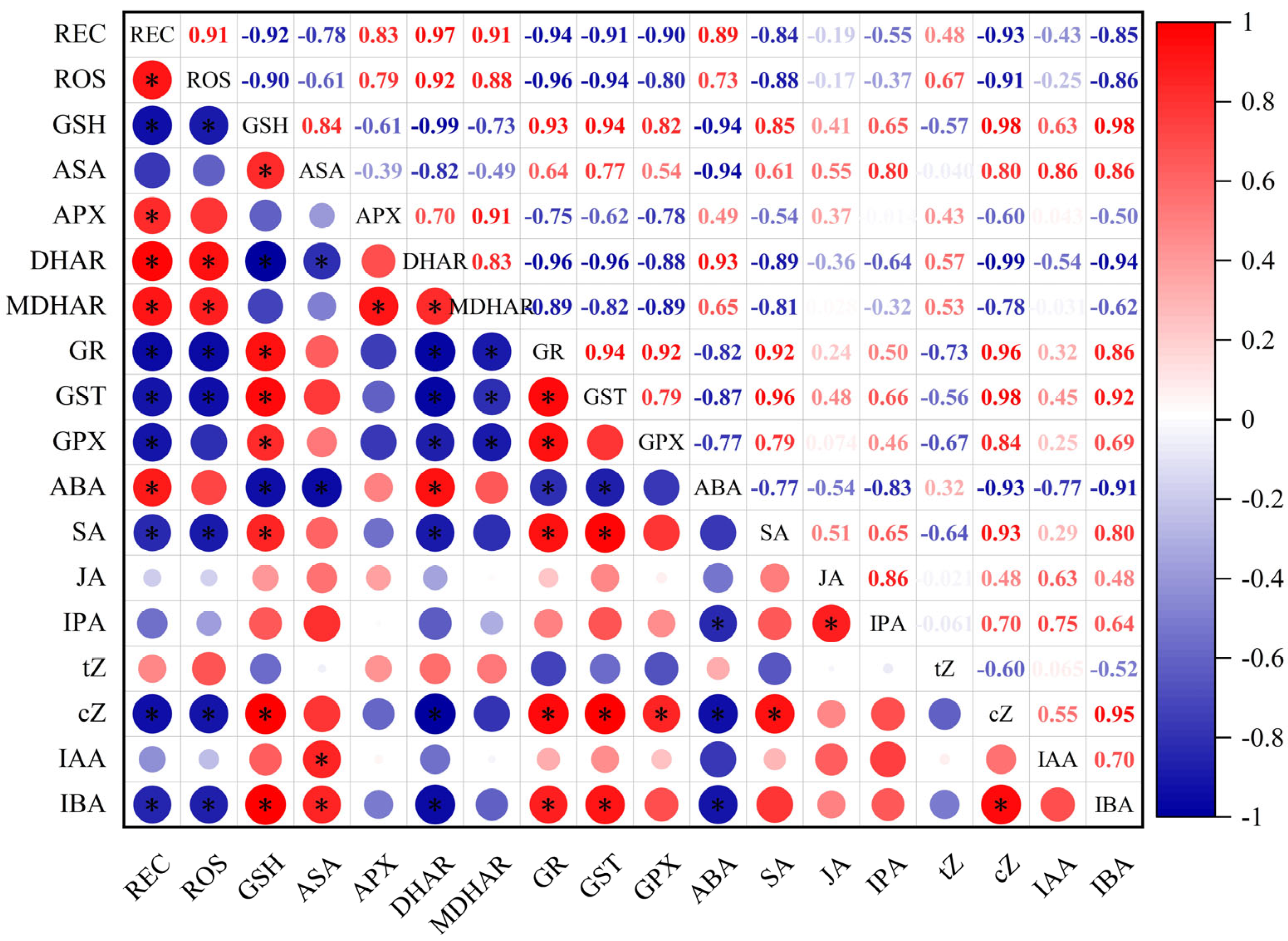
2.6. Correlation Analysis
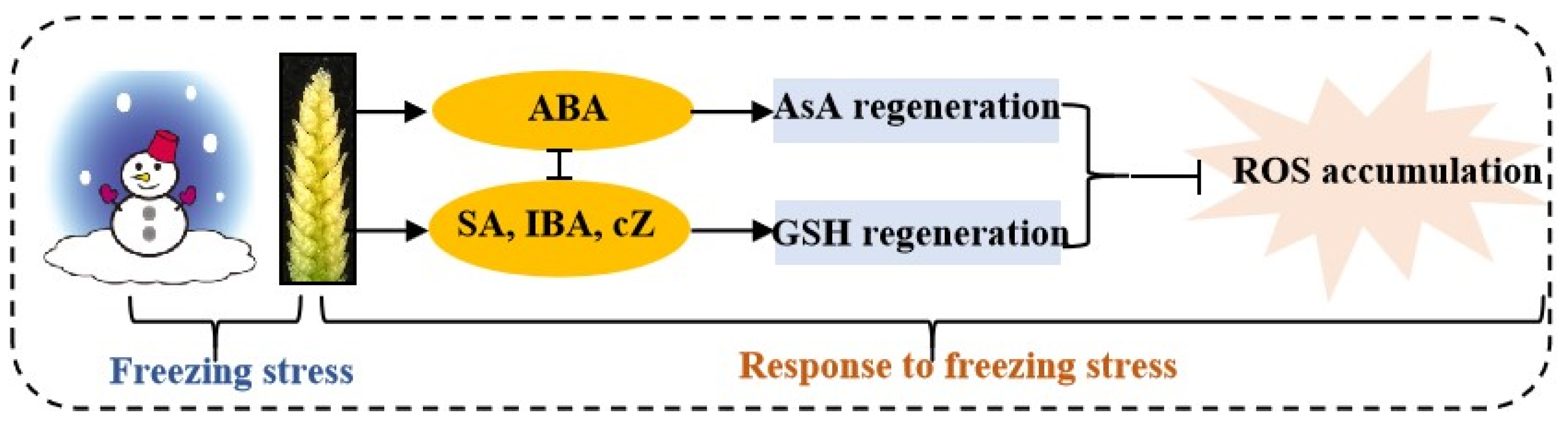
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Plant Material
4.2. Measurements
4.2.1. Plant Height and Dry Mass
4.2.2. Regenerated Tiller and Grain Number
4.2.3. ROS Release Rate and REC
4.2.4. AsA–GSH Cycle
4.2.5. Endogenous Hormone Contents
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, K.; Gong, Q.; Ye, X. Recent developments and applications of genetic transformation and genome editing technologies in wheat. Theor. Appl. Genet. 2020, 133, 1603–1622. [Google Scholar] [CrossRef]
- Zhong, X.; Wang, D.; Zhao, P.; Yan, X.; Su, C. Occurrence of frost temperature in Huanghuai wheat production zone after winter wheat elongation. Chin. J. Eco-Agri. 2007, 15, 17–20. [Google Scholar]
- Jiang, G.; Hassan, M.A.; Muhammad, N.; Arshad, M.; Chen, X.; Xu, Y.H.; Xu, H.; Ni, Q.Q.; Liu, B.B.; Yang, W.K.; et al. Comparative Physiology and Transcriptome Analysis of Young Spikes in Response to Late Spring Coldness in Wheat (Triticum aestivum L.). Front. Plant Sci. 2022, 13, 811–884. [Google Scholar] [CrossRef]
- Lin, F.F.; Cheng, L.; Xu, B.; Chen, J.; Chen, A.H.; Hassan, M.A.; Liu, B.B.; Xu, H.; Chen, X.; Sun, J.Q.; et al. Late spring cold reduces grain number at various spike positions by regulating spike growth and assimilate distribution in winter wheat. Crop J. 2023, 11, 1272–1278. [Google Scholar] [CrossRef]
- Xiao, L.J.; Liu, L.L.; Asseng, S.; Xia, Y.M.; Tang, L.; Liu, B.; Cao, W.X.; Zhu, Y. Estimating spring frost and its impact on yield across winter wheat in China. Agric. For. Meteorol. 2018, 260–261, 154–164. [Google Scholar] [CrossRef]
- Crimp, S.; Zheng, B.; Khimashia, N.; Gobbett, D.; Chapman, S.; Howden, M.; Nicholls, N. Recent changes in Southern Australian frost occurrence: Implications for wheat production risk. Crop Pasture Sci. 2016, 8, 801–811. [Google Scholar] [CrossRef]
- Zhong, X.; Mei, X.; Li, Y.; Yoshida, H.; Zhao, P.; Wang, X.; Han, L.; Hu, X.; Huang, S.; Huang, J. Changes in frost resistance of wheat young ears with development during jointing stage. J. Agron. Crop Sci. 2008, 194, 343–349. [Google Scholar] [CrossRef]
- Saeidi, M.; Eliasi, P.; Abdoly, M.; Sasani, S. Freezing tolerance of wheat cultivars at the early growing season after winter. Afr. J. Biotechnol. 2012, 11, 4045–4052. [Google Scholar] [CrossRef]
- Shimono, H. Earlier rice phenology as a result of climate change can increase the risk of cold damage during reproductive growth in northern Japan. Agric. Ecosyst. Environ. 2011, 144, 201–207. [Google Scholar] [CrossRef]
- Li, L.; Han, C.L.; Yang, J.W.; Tian, Z.Q.; Jiang, R.Y.; Yang, F.; Jiao, K.M.; Qi, M.L.; Liu, L.L.; Zhang, B.Z.; et al. Comprehensive Transcriptome Analysis of Responses during Cold Stress in Wheat (Triticum aestivum L.). Genes 2023, 14, 844. [Google Scholar] [CrossRef] [PubMed]
- Fuller, M.P.; Fuller, A.M.; Kaniouras, S.; Christophers, J.; Fredericks, T. The freezing characteristics of wheat at ear emergence. Eur. J. Agron. 2007, 26, 435–441. [Google Scholar] [CrossRef]
- Gabriela, P.; Eugen, P.; Cătălin, D.S.; Cătălin, M. Basal sterility of wheat ears—Index of sensitivity to delayed spring frozen and its relationship with yield and productivity elements, in the climatic conditions of 2020 year at ards caracal. Ann. Univ. Craiova Agri. Montan. Cad. Ser. 2020, 50, 180–184. [Google Scholar]
- Zhang, L.L.; Chen, X.; Ke, Y.Y.; Liu, B.B.; Muhammad, A.H.; Zhang, Y.; Xu, H.; Liu, J.C. An Inclusive Comparison of Identification Methods and Indices of Winter Wheat Tolerance against Late Spring Coldness. Chin. J. Agrom. 2021, 42, 146–157. [Google Scholar]
- Yue, Y.J.; Zhou, Y.; Wang, J.A.; Ye, X.Y. Assessing Wheat Frost Risk with the Support of GIS: An Approach Coupling a Growing Season Meteorological Index and a Hybrid Fuzzy Neural Network Model. Sustainability 2016, 8, 1308. [Google Scholar] [CrossRef]
- Choudhury, S.; Panda, P.; Sahoo, L.; Panda, S.K. Reactive oxygen species signaling in plants under abiotic stress. Plant Signal. Behav. 2013, 8, e23681. [Google Scholar] [CrossRef]
- Li, X.N.; Cai, J.; Liu, F.L.; Dai, T.B.; Cao, W.X.J.D. Cold priming drives the sub-cellular antioxidant systems to protect photosynthetic electron transport against subsequent low temperature stress in winter wheat. Plant Physiol. Biochem. 2014, 82, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.L.; Zhang, Y.T.; Liu, C.; Dong, Y.W.; Jiang, X.; Zhao, C.; Li, G.H.; Xu, K.; Huo, Z.Y. Label-Free Quantitative Proteomics Reveal the Mechanisms of Young Wheat (Triticum aestivum L.) Ears’ Response to Spring Freezing. Int. J. Mol. Sci. 2023, 24, 15892. [Google Scholar] [CrossRef]
- Apostolova1, P.; Yordanova1, R.; Popova, L. Response of antioxidative defence system to low temperature stress in two wheat cultivars. Gen. Appl. Plant Physiol. 2008, 34, 281–294. [Google Scholar]
- Ruelland, E.; Vaultier, M.N.; Zachowski, A.; Hurry, V. Cold signalling and cold acclimation in plants. Adv. Bot. Res. 2009, 49, 35–150. [Google Scholar]
- Wang, W.L.; Wang, X.; Lv, Z.S.; Khanzada, A.; Huang, M.; Cai, J.; Zhou, Q.; Huo, Z.Y.; Jiang, D. Effects of cold and salicylic acid priming on free proline and sucrose accumulation in winter wheat under freezing stress. J. Plant Growth Regul. 2022, 41, 2171–2184. [Google Scholar] [CrossRef]
- Tiwari, Y.K.; Yadav, S.K. Effect of High-Temperature Stress on Ascorbate–Glutathione Cycle in Maize. Agric. Res. 2020, 9, 179–187. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, D.Y.; Zhu, M.; Li, F.H. Exogenous 6-benzyladenine improves waterlogging tolerance in maize seedlings by mitigating oxidative stress and upregulating the ascorbate-glutathione cycle. Front. Plant Sci. 2021, 12, 680376. [Google Scholar] [CrossRef] [PubMed]
- Iwaniuk, P.; Kaczyński, P.; Pietkun, M.; Łozowicka, B. Evaluation of titanium and silicon role in mitigation of fungicides toxicity in wheat expressed at the level of biochemical and antioxidant profile. Chemosphere 2022, 308, 136284. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.L.; Wang, X.; Zhang, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.B.; Jiang, D. Salicylic acid and cold priming induce late-spring freezing tolerance by maintaining cellular redox homeostasis and protecting photosynthetic apparatus in wheat. Plant Growth Regul. 2021, 90, 109–121. [Google Scholar] [CrossRef]
- Yu, J.; Cang, J.; Lu, Q.W.; Fan, B.; Xu, Q.H.; Li, W.N.; Wang, X.T. ABA enhanced cold tolerance of wheat ‘dn1′via increasing ROS scavenging system. Plant Signal. Behav. 2020, 15, 1780403. [Google Scholar] [CrossRef]
- Zhang, L.R.; Zhang, N.; Wang, S.S.; Tian, H.Y.; Liu, L.; Pei, D.; Yu, X.D.; Zhao, L.; Chen, F. A TaSnRK1 α Modulates TaPAP6L-Mediated Wheat Cold Tolerance through Regulating Endogenous Jasmonic Acid. Adv. Sci. 2023, 10, 2303478. [Google Scholar] [CrossRef]
- Liu, L.L.; Xia, Y.M.; Liu, B.; Chang, C.Y.; Xiao, L.J.; Shen, J.; Tang, L.; Cao, W.X. Individual and combined effects of jointing and booting low-temperature stress on wheat yield. Eur. J. Agron. 2020, 113, 125989. [Google Scholar] [CrossRef]
- Roxas, V.P.; Lodhi, S.A.; Garrett, D.K.; Mahan, J.R.; Allen, R.D. Stress tolerance in transgenic tobacco seedlings that overexpress glutathione S-transferase/glutathione peroxidase. Plant Cell Physiol. 2000, 41, 1229–1234. [Google Scholar] [CrossRef]
- Han, Q.X.; Kang, G.Z.; Guo, T.C. Proteomic analysis of spring freeze-stress responsive proteins in leaves of bread wheat (Triticum aestivum L.). Plant Physiol Biochem. 2013, 63, 236–244. [Google Scholar]
- Cheong, B.E.; Onyemaobi, O.; Wing Ho Ho, W.W.; Biddulph, T.B.; Rupasinghe, T.W.; Roessner, U.; Dolferus, R. Phenotyping the Chilling and Freezing Responses of Young Microspore Stage Wheat Spikes Using Targeted Metabolome and Lipidome Profiling. Cells 2020, 9, 1309. [Google Scholar] [CrossRef]
- Zhang, W.J.; Wang, J.Q.; Huang, Z.L.; Mi, L.; Xu, K.; Wu, J.J.; Fan, Y.H.; Ma, S.Y.; Jiang, D.G. Effects of low temperature at booting stage on sucrose metabolism and endogenous hormone contents in winter wheat spikelet. Front. Plant Sci. 2019, 10, 498. [Google Scholar] [CrossRef] [PubMed]
- Li, X.N.; Pu, H.C.; Liu, F.L.; Zhou, Q.; Cai, J.; Dai, T.B.; Cao, W.X.; Jiang, D. Winter wheat photosynthesis and grain yield responses to spring freeze. Agron. J. 2015, 107, 1002–1010. [Google Scholar] [CrossRef]
- Liang, Z.; Luo, J.; Wei, B.; Liao, Y.; Liu, Y. Trehalose can alleviate decreases in grain number per spike caused by low-temperature stress at the booting stage by promoting floret fertility in wheat. J. Agron. Crop Sci. 2021, 207, 717–732. [Google Scholar] [CrossRef]
- Wang, W.L.; Wang, X.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.B.; Cao, W.X.; Jiang, D. Hydrogen peroxide and abscisic acid mediate salicylic acid-induced freezing tolerance in wheat. Front. Plant Sci. 2018, 9, 1137. [Google Scholar] [CrossRef]
- Si, T.; Wang, X.; Wu, L.; Zhao, C.Z.; Zhang, L.N.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.B.; Zhu, J.K.; et al. Nitric oxide and hydrogen peroxide mediate wounding-induced freezing tolerance through modifications in photosystem and antioxidant system in wheat. Front. Plant Sci. 2017, 8, 1284. [Google Scholar] [CrossRef]
- Kosová, K.; Prášil, I.T.; Vítámvás, P.; Dobrev, P.; Motyka, V.; Floková, K.; Vanková, R. Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter Samanta and spring Sandra. J. Plant Physiol. 2012, 169, 567–576. [Google Scholar] [CrossRef]
- Zhang, N.; Li, L.j.; Zhang, L.R.; Li, J.; Fang, Y.H.; Zhao, L.; Ren, Y.; Chen, F. Abscisic acid enhances tolerance to spring freeze stress and regulates the expression of ascorbate-glutathione biosynthesis-related genes and stress-responsive genes in common wheat. Mol. Breed. 2020, 40, 108. [Google Scholar] [CrossRef]
- Wang, W.L.; Wang, X.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.B.; Jiang, D. Alleviation of field low-temperature stress in winter wheat by exogenous application of salicylic acid. J. Plant Growth Regul. 2021, 40, 811–823. [Google Scholar] [CrossRef]
- Repkina, N.; Ignatenko, A.; Holoptseva, E.; MiszalskI, Z.; Kaszycki, P.; Talanova, V. Exogenous methyl jasmonate improves cold tolerance with parallel induction of two cold-regulated (COR) genes expression in Triticum aestivum L. Plants 2021, 10, 1421. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmed, S.S.; He, Y.M. Exogenous abscisic acid increases antioxidant enzymes and related gene expression in pepper (Capsicum annuum) leaves subjected to chilling stress. Genet. Mol. Res. 2012, 11, 4063–4080. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K.; Arora, K. Arsenic-induced root growth inhibition in mung bean (Phaseolus aureus Roxb.) is due to oxidative stress resulting from enhanced lipid peroxidation. Plant Growth Regul. 2007, 53, 65–73. [Google Scholar] [CrossRef]
- Xia, J.X.; Wang, Y.J.M.; Zhou, Y.H.; Tao, Y.; Mao, W.H.; Shi, K.; Asami, T.; Chen, Z.X.; Yu, J.Q. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Simura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant Hormonomics: Multiple Phytohormone Profiling by Targeted Metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | XM33 | JM22 | ||
|---|---|---|---|---|
| C | F | C | F | |
| MDHAR (U g−1 FW) | 2.845 ± 0.013 c | 3.621 ± 0.031 a | 2.822 ± 0.109 c | 3.162 ± 0.145 b |
| DHAR (U g−1 FW) | 0.153 ± 0.005 b | 0.266 ± 0.008 a | 0.152 ± 0.005 b | 0.109 ± 0.005 c |
| APX (U g−1 FW) | 3.233 ± 0.109 c | 4.102 ± 0.023 a | 3.701 ± 0.085 b | 3.892 ± 0.149 ab |
| GR (U g−1 FW) | 0.302 ± 0.007 b | 0.189 ± 0.010 c | 0.382 ± 0.010 a | 0.304 ± 0.009 b |
| GST (U g−1 FW) | 0.436 ± 0.014 a | 0.240 ± 0.008 b | 0.437 ± 0.014 a | 0.417 ± 0.011 a |
| GPX (U g−1 FW) | 1.211 ± 0.0323 a | 1.042 ± 0.044 b | 0.957 ± 0.014 b | 1.204 ± 0.024 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ni, C.; Dong, Y.; Jiang, X.; Liu, C.; Wang, W.; Zhao, C.; Li, G.; Xu, K.; Huo, Z. The Role of the Ascorbic Acid–Glutathione Cycle in Young Wheat Ears’ Response to Spring Freezing Stress. Plants 2023, 12, 4170. https://doi.org/10.3390/plants12244170
Zhang Y, Ni C, Dong Y, Jiang X, Liu C, Wang W, Zhao C, Li G, Xu K, Huo Z. The Role of the Ascorbic Acid–Glutathione Cycle in Young Wheat Ears’ Response to Spring Freezing Stress. Plants. 2023; 12(24):4170. https://doi.org/10.3390/plants12244170
Chicago/Turabian StyleZhang, Yuting, Chunyang Ni, Yongwen Dong, Xue Jiang, Chang Liu, Weiling Wang, Can Zhao, Guohui Li, Ke Xu, and Zhongyang Huo. 2023. "The Role of the Ascorbic Acid–Glutathione Cycle in Young Wheat Ears’ Response to Spring Freezing Stress" Plants 12, no. 24: 4170. https://doi.org/10.3390/plants12244170
APA StyleZhang, Y., Ni, C., Dong, Y., Jiang, X., Liu, C., Wang, W., Zhao, C., Li, G., Xu, K., & Huo, Z. (2023). The Role of the Ascorbic Acid–Glutathione Cycle in Young Wheat Ears’ Response to Spring Freezing Stress. Plants, 12(24), 4170. https://doi.org/10.3390/plants12244170