


Effects of Climate Conditions before Harvest Date on Edamame Metabolome

 ,
,
Abstract
1. Introduction
2. Results
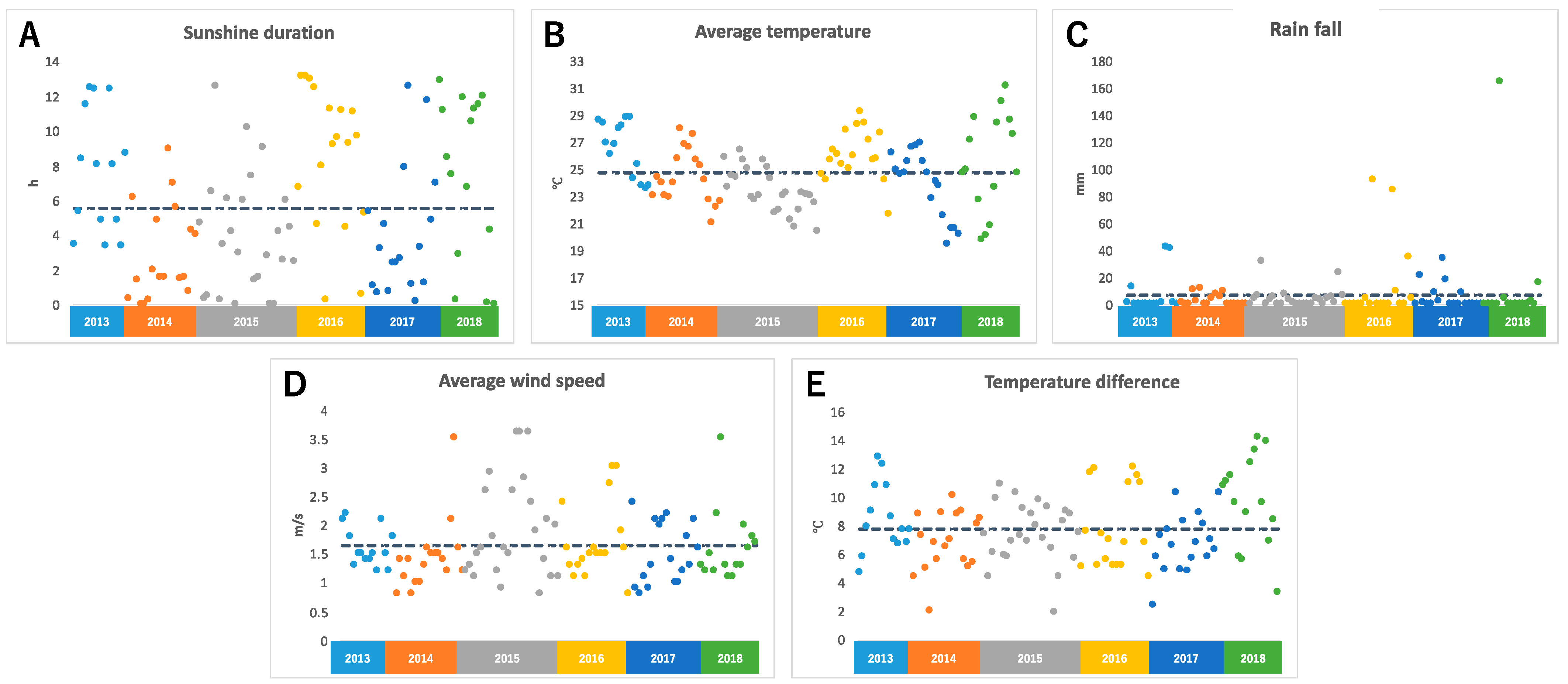
2.1. Climate Conditions before Harvest Days
2.1.1. Sunshine Duration, Average Temperature, Rainfall, Average Wind Speed, and Temperature Difference on the Day before Harvest
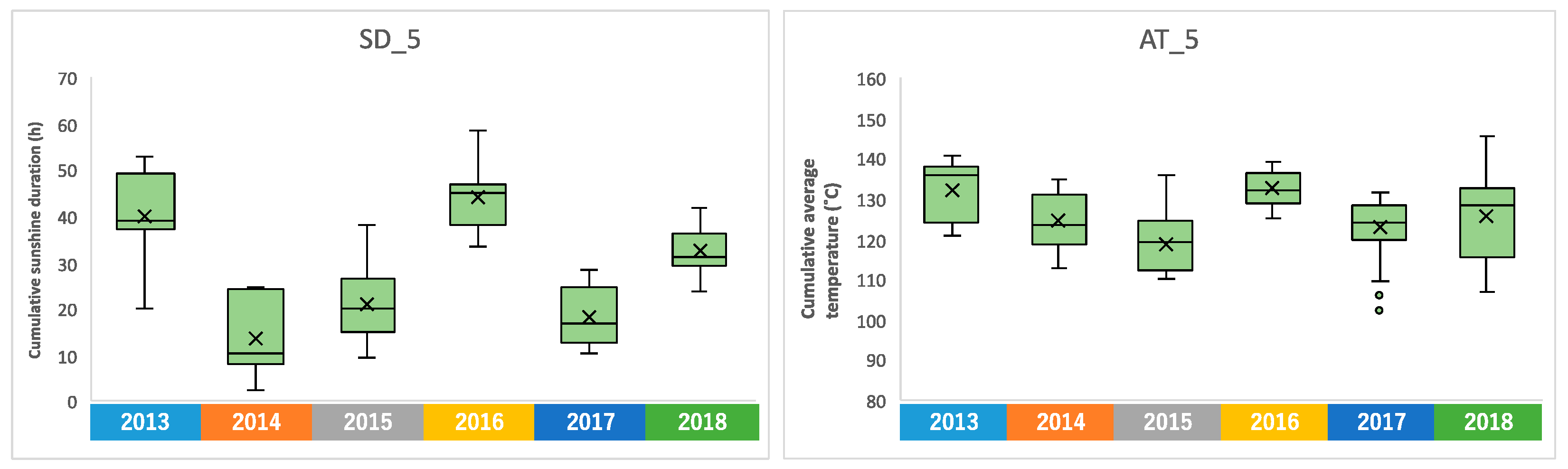
2.1.2. Cumulative Sunshine Duration, Average Temperature, and Rainfall in the Days before Harvest
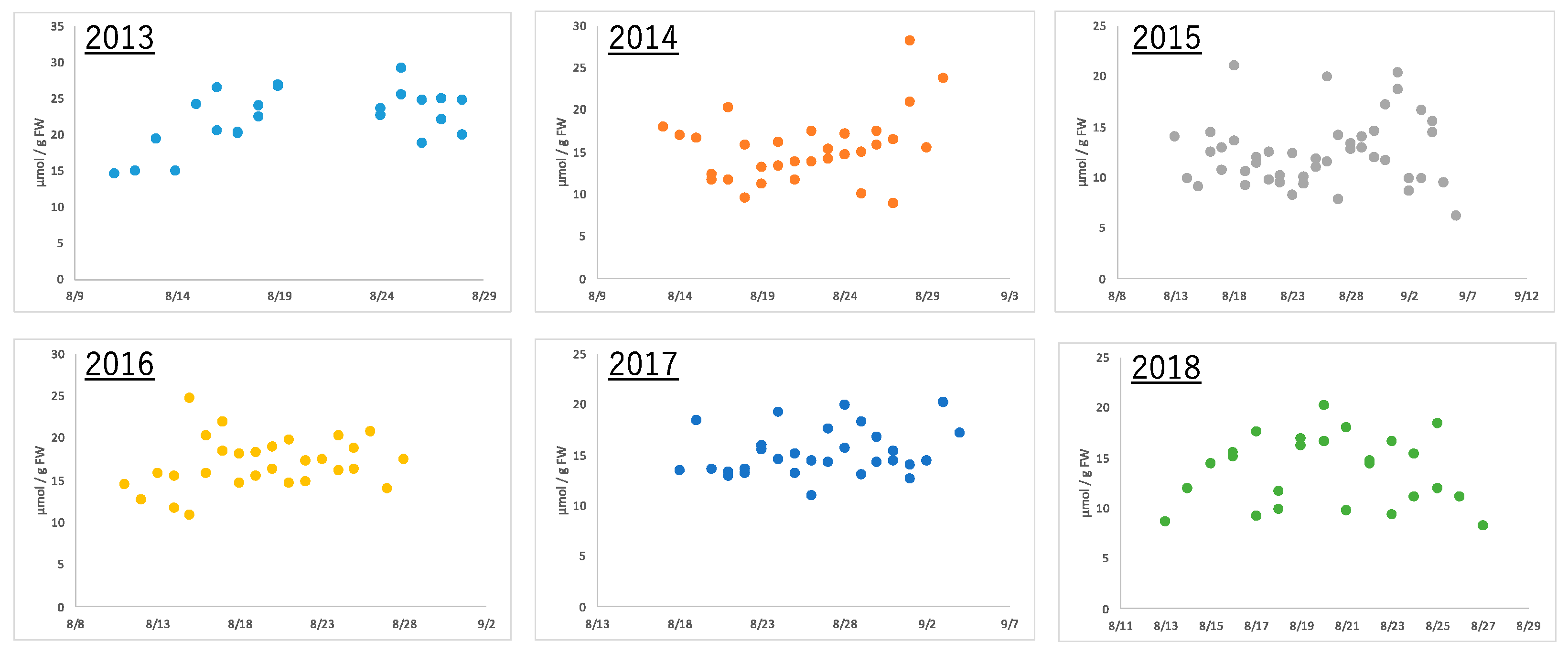
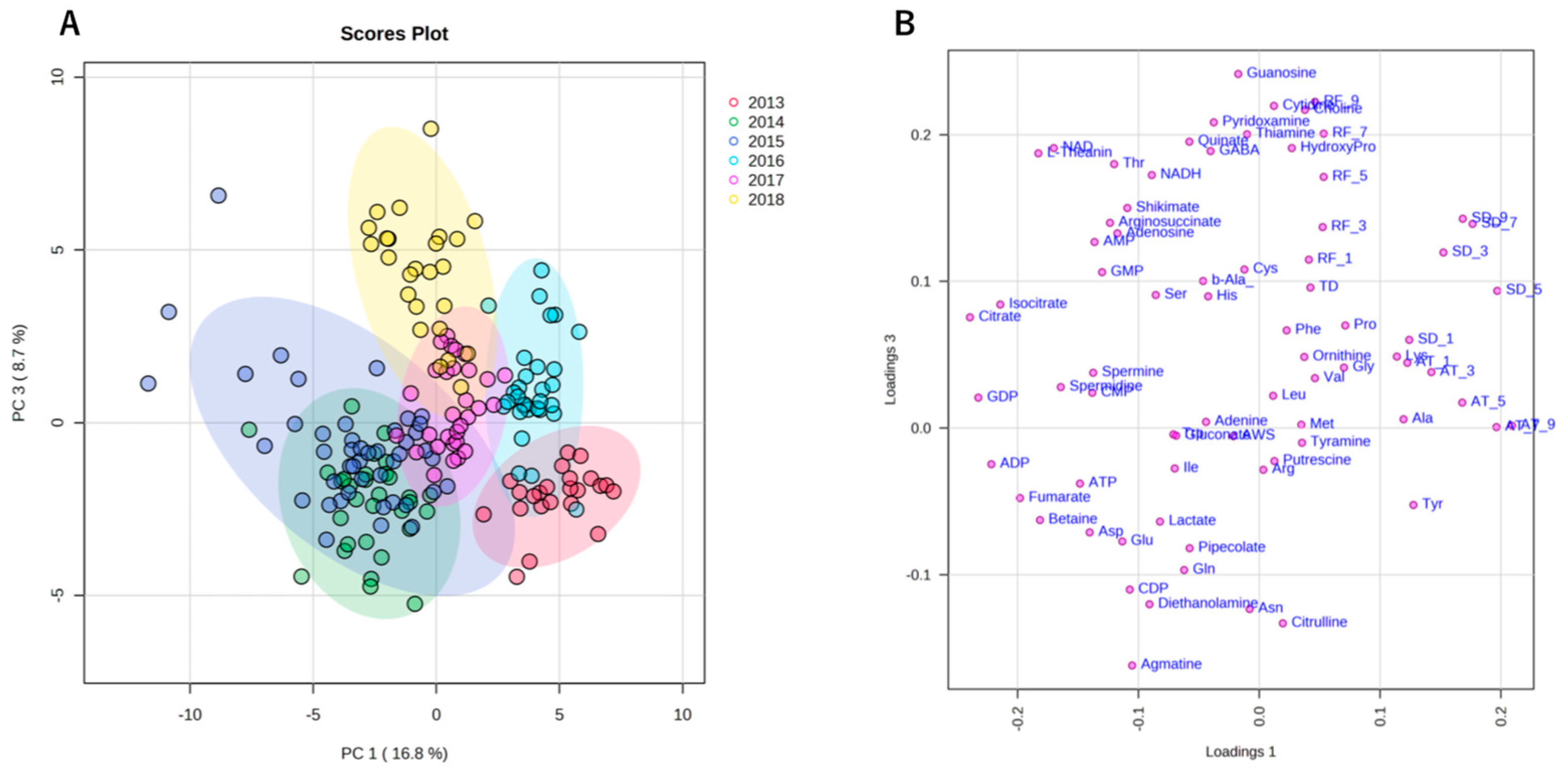
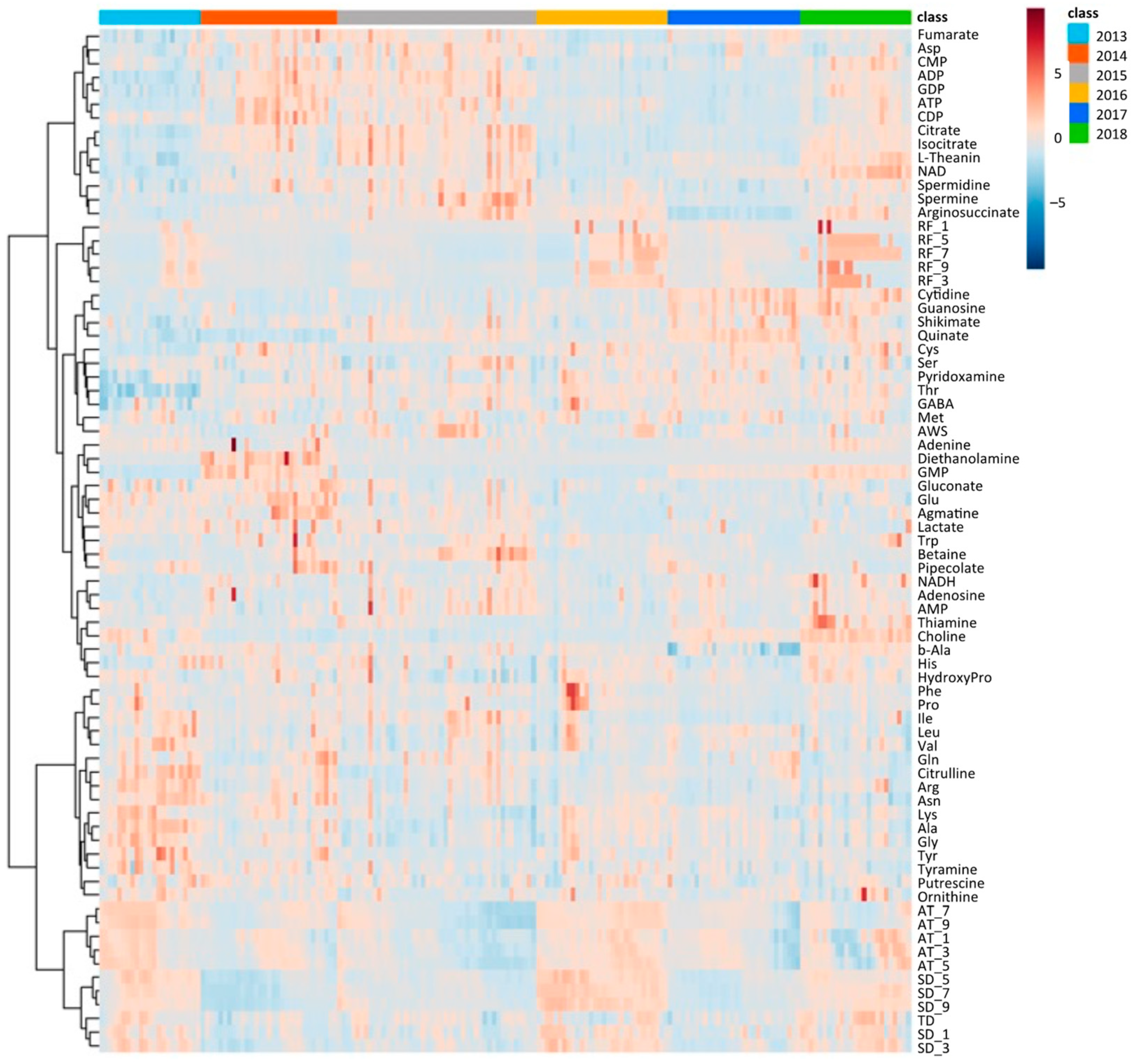
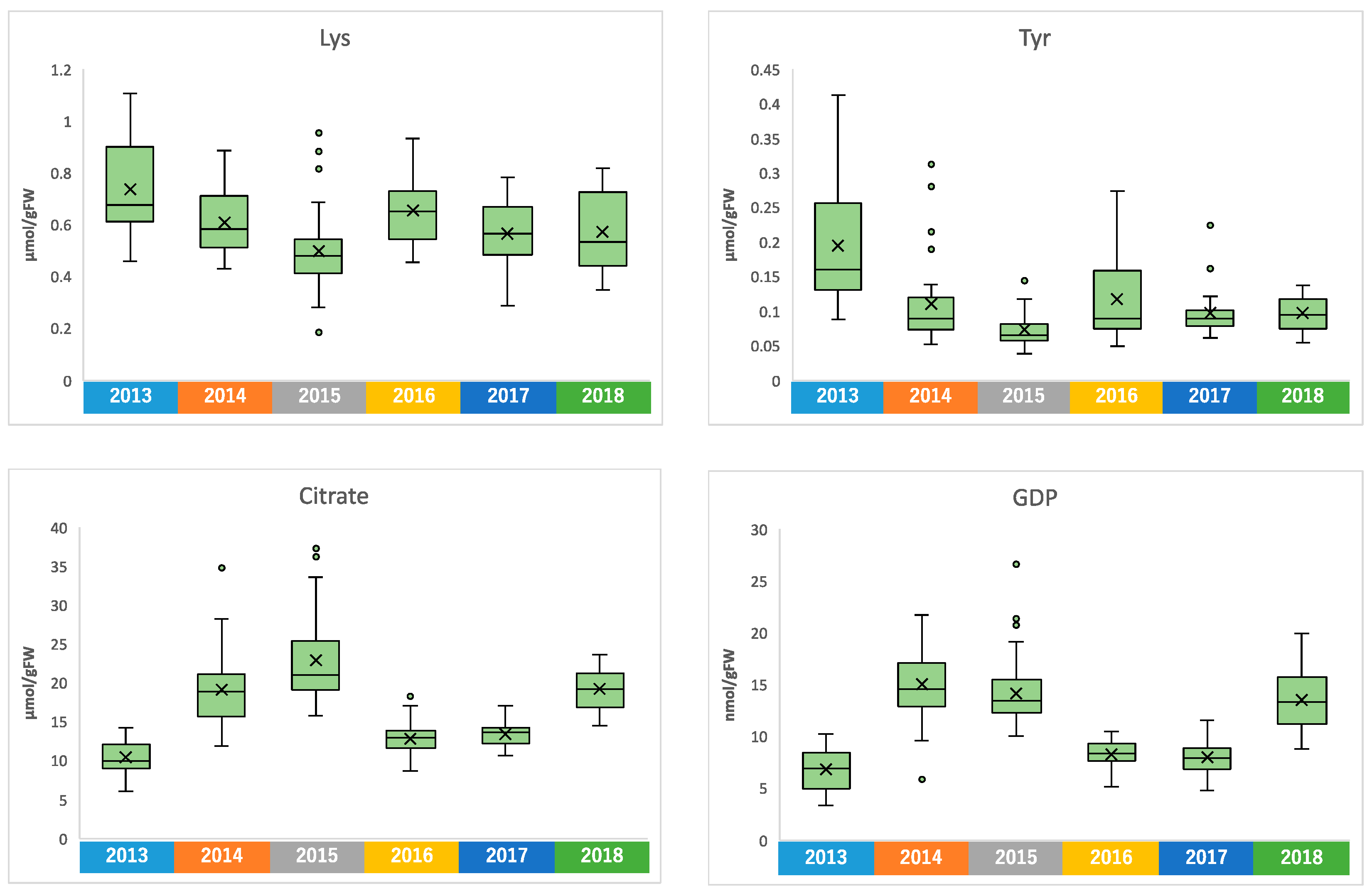
2.2. Metabolome of Harvested Edamame
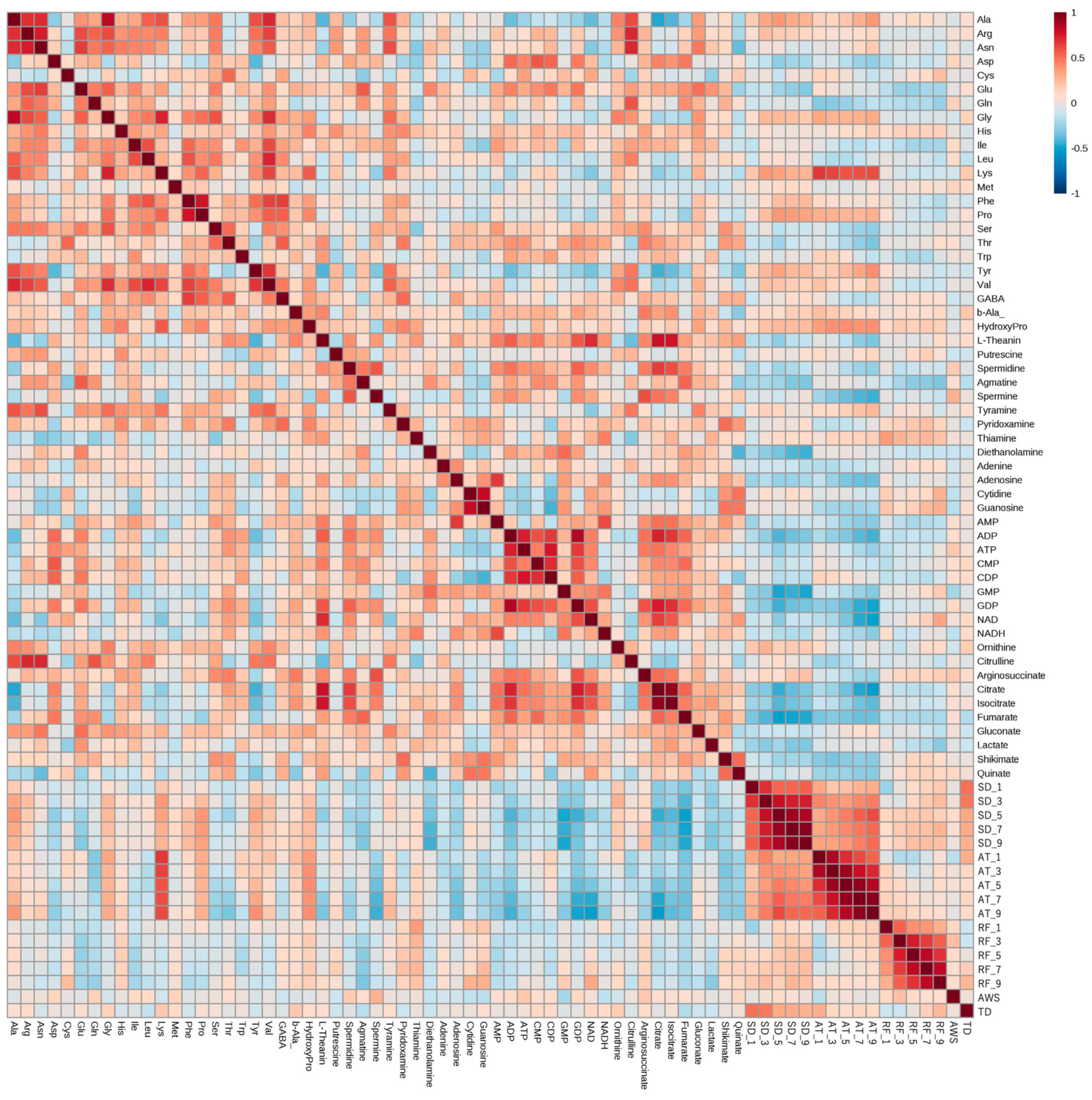
2.3. Correlation Analysis between Climate Conditions up to the Day before Harvest and Compound Concentrations in Harvested Edamame
2.3.1. Correlation Analysis between Climate Conditions and Edamame Metabolome
2.3.2. Compounds That Were Correlated with Sunshine Duration and Average Temperature
2.4. Annual Fluctuation between Meteorological Conditions and Edamame Metabolome
2.4.1. Annual Fluctuation of Climate Conditions
2.4.2. Annual Fluctuation of Edamame Metabolome
3. Discussion
4. Materials and Methods
4.1. Harvesting and Sampling of Edamame
4.2. Collection of Climate Conditions
4.3. Sample Extraction
4.4. CE-MS Conditions
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nair, R.M.; Boddepalli, V.N.; Yan, M.R.; Kumar, V.; Gill, B.; Pan, R.S.; Wang, C.; Hartman, G.L.; Silva e Souza, R.; Somta, P. Global status of vegetable soybean. Plants 2023, 12, 609. [Google Scholar] [CrossRef] [PubMed]
- Moseley, D.; Paulo da Silva, M.; Mozzoni, L.; Orazaly, M.; Florez-Palacios, L.; Acuña, A.; Wu, C.; Chen, P. Effect of planting date and cultivar maturity in edamame quality and harvest window. Front. Plant Sci. 2021, 11, 585856. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, K.; Zhu, Q.; Zhang, B.; Li, M.; Li, S. Analysis of shoot architecture traits in edamame reveals potential strategies to improve harvest efficiency. Front. Plant Sci. 2021, 12, 614926. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Lord, N.; Polk, J.; Dhakal, K.; Li, S.; Yin, Y.; Duncan, S.E.; Wang, H.; Zhang, B.; Huang, H. Physical and chemical properties of edamame during bean development and application of spectroscopy-based machine learning methods to predict optimal harvest time. Food Chem. 2022, 368, 130799. [Google Scholar] [CrossRef] [PubMed]
- Koshika, N.; Shioya, N.; Fujimura, T.; Oguchi, R.; Ota, C.; Kato, E.; Takahashi, R.; Kimura, S.; Furuno, S.; Saito, K.; et al. Development of ethyl methanesulfonate mutant edamame soybean (Glycine max (L.) Merr.) populations and forward and reverse genetic screening for early-flowering mutants. Plants 2022, 11, 1839. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, D.; Morota, G.; Dhakal, K.; Singer, W.; Lord, N.; Huang, H.; Chen, P.; Mozzoni, L.; Li, S.; et al. Genome-wide association analysis of sucrose and alanine contents in edamame beans. Front. Plant Sci. 2022, 13, 1086007. [Google Scholar] [CrossRef]
- Zang, Y.; Sato, H.; Igarashi, K. Anti-diabetic effects of a kaempferol glycoside-rich fraction from unripe soybean (edamame, Glycine max L. Merrill. ‘Jindai’) leaves on KK-Ay mice. Biosci. Biotechnol. Biochem. 2011, 75, 1677–1684. [Google Scholar] [CrossRef]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef]
- Sugimoto, M.; Goto, H.; Otomo, K.; Ito, M.; Onuma, H.; Suzuki, A.; Sugawara, M.; Abe, S.; Tomita, M.; Soga, T. Metabolomic profiles and sensory attributes of edamame under various storage duration and temperature conditions. J. Agric. Food Chem. 2010, 58, 8418–8425. [Google Scholar] [CrossRef]
- Roullier-Gall, C.; Lucio, M.; Noret, L.; Schmitt-Kopplin, P.; Gougeon, R.D. How subtle is the “terroir” effect? Chemistry-related signatures of two “climats de Bourgogne”. PLoS ONE 2014, 9, e97615. [Google Scholar] [CrossRef]
- Reshef, N.; Agam, N.; Fait, A. Grape berry acclimation to excessive solar irradiance leads to repartitioning between major flavonoid groups. J. Agric. Food Chem. 2018, 66, 3624–3636. [Google Scholar] [CrossRef] [PubMed]
- Rustioni, L.; Altomare, A.; Shanshiashivili, G.; Greco, F.; Buccolieri, R.; Blanco, I.; Cola, G.; Fracassetti, D. Microclimate of grape bunch and sunburn of white grape berries: Effect on wine quality. Foods 2023, 12, 621. [Google Scholar] [CrossRef] [PubMed]
- Reshef, N.; Walbaum, N.; Agam, N.; Fait, A. Sunlight modulates fruit metabolic profile and shapes the spatial pattern of compound accumulation within the grape cluster. Front. Plant Sci. 2017, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- Garrido, A.; Engel, J.; Mumm, R.; Conde, A.; Cunha, A.; De Vos, R.C.H. Metabolomics of photosynthetically active tissues in white grapes: Effects of light microclimate and stress mitigation strategies. Metabolites 2021, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, M.; Chen, K.; Zhang, J.; Xue, T.; Cheng, Z.; Zhang, B.; Zhang, K.; Fang, Y. The roles of different photoselective nets in the targeted regulation of metabolite accumulation, wine aroma and sensory profiles in warm viticulture regions. Food Chem. 2022, 396, 133629. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.B.; Liu, Y.; Lu, H.C.; Hu, L.; Wang, Y.; Cheng, C.F.; Chen, W.; Li, S.D.; He, F.; Duan, C.Q.; et al. Volatomics of ‘Cabernet Sauvignon’ grapes and wines under the fan training system revealed the nexus of microclimate and volatile compounds. Food Chem. 2023, 403, 134421. [Google Scholar] [CrossRef] [PubMed]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How does tomato quality (sugar, acid, and nutritional quality) vary with ripening stage, temperature, and irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef]
- Leyva, R.; Constán-Aguilar, C.; Blasco, B.; Sánchez-Rodríguez, E.; Romero, L.; Soriano, T.; Ruíz, J.M. Effects of climatic control on tomato yield and nutritional quality in Mediterranean screenhouse. J. Sci. Food Agric. 2014, 94, 63–70. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hakata, M. Atlas of rice grain filling-related metabolism under high temperature: Joint analysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of amino acid accumulation. Plant Cell Physiol. 2010, 51, 795–809. [Google Scholar] [CrossRef]
- El-kereamy, A.; Bi, Y.M.; Ranathunge, K.; Beatty, P.H.; Good, A.G.; Rothstein, S.J. The rice R2R3-MYB transcription factor OsMYB55 is involved in the tolerance to high temperature and modulates amino acid metabolism. PLoS ONE 2012, 7, e52030. [Google Scholar] [CrossRef]
- Hung, K.A.; Hsu, Y.W.; Chen, Y.C.; Lin, T.P. Influence of microclimate control on the growth of asparagus under greenhouse in tropical climates. Int. J. Biometeorol. 2023, 67, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Bosmali, I.; Giannenas, I.; Christophoridou, S.; Ganos, C.G.; Papadopoulos, A.; Papathanasiou, F.; Kolonas, A.; Gortzi, O. Microclimate and genotype impact on nutritional and antinutritional quality of locally adapted landraces of common bean (Phaseolus vulgaris L.). Foods 2023, 12, 1119. [Google Scholar] [CrossRef] [PubMed]
- Stone, C.H.; Close, D.C.; Bound, S.A.; Corkrey, R. Orchard microclimate, tree water uptake and sweet cherry fruit quality under protected cropping. Front. Plant Sci. 2022, 13, 993817. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Schmilovitch, Z.; Ginzberg, I. Pomegranate fruit growth and skin characteristics in hot and dry climate. Front. Plant Sci. 2021, 12, 725479. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.R.; Paul, N.D. Seduced by the dark side: Integrating molecular and ecological perspectives on the influence of light on plant defence against pests and pathogens. New Phytol. 2006, 170, 677–699. [Google Scholar] [CrossRef] [PubMed]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Jorge, T.F.; Rodrigues, J.A.; Caldana, C.; Schmidt, R.; van Dongen, J.T.; Thomas-Oates, J.; António, C. Mass spectrometry-based plant metabolomics: Metabolite responses to abiotic stress. Mass Spec. Rev. 2016, 35, 620–649. [Google Scholar] [CrossRef]
- Zheng, J.; Huang, C.; Yang, B.; Kallio, H.; Liu, P.; Ou, S. Regulation of phytochemicals in fruits and berries by environmental variation–sugars and organic acids. J. Food Biochem. 2019, 43, e12642. [Google Scholar] [CrossRef]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The influence of environmental conditions on secondary metabolites in medicinal plants: A literature review. Chem. Biodivers. 2021, 18, e2100345. [Google Scholar] [CrossRef]
- Palmieri, L.; Masuero, D.; Martinatti, P.; Baratto, G.; Martens, S.; Vrhovsek, U. Genotype-by-environment effect on bioactive compounds in strawberry (Fragaria x ananassa Duch.). J. Sci. Food Agric. 2017, 97, 4180–4189. [Google Scholar] [CrossRef]
- Casati, P.; Campi, M.; Morrow, D.J.; Fernandes, J.; Walbot, V. Transcriptomic, proteomic and metabolomic analysis of maize responses to UV-B. Plant Signal. Behav. 2011, 6, 1146–1153. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wheeler, G.R.; Brassil, C.E.; Knops, J.M.H. Functional traits’ annual variation exceeds nitrogen-driven variation in grassland plant species. Ecology 2023, 104, e3886. [Google Scholar] [CrossRef] [PubMed]
- Westerman, L.; Roddick, J.G. Annual variation in sterol levels in leaves of Taraxacum officinale Weber. Plant Physiol. 1981, 68, 872–875. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Ujihara, T.; Tanaka, E.; Kishi, Y.; Ogawa, H.; Matsuo, H. Annual variation of natural 15N abundance in tea leaves and its practicality as an organic tea indicator. J. Agric. Food Chem. 2011, 59, 10317–10321. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.L.; Katuuramu, D.N. Comparison of seed fatty and amino acids in edamame dried using two oven-drying methods and mature soybean. J. Sci. Food Agric. 2021, 101, 1515–1522. [Google Scholar] [CrossRef]
- Iman, M.N.; Irdiani, R.; Rahmawati, D.; Fukusaki, E.; Putri, S.P. Improvement of the functional value of green soybean (edamame) using germination and tempe fermentation: A comparative metabolomics study. J. Biosci. Bioeng. 2023, 136, 205–212. [Google Scholar] [CrossRef]
- Ugarte, R.M.; Escudero, A.; Gavilán, R.G. Metabolic and physiological responses of Mediterranean high-mountain and alpine plants to combined abiotic stresses. Physiol. Plant. 2019, 165, 403–412. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Suravajhala, P.; Rathnagiri, P.; Sreenivasulu, N. Intriguing role of proline in redox potential conferring high temperature stress tolerance. Front. Plant Sci. 2022, 13, 867531. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Javid, M.G.; Soltani, E.; Wojtyla, L.; Garnczarska, M. Contirbution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M.U.; Saeed, F.; Haider, S.; Sharif, R.; Anand, A.; Corpas, F.J.; Jin, W.; et al. Assessment of proline function in higher plants under extreme temperature. Plant Biol. 2023, 25, 379–395. [Google Scholar] [CrossRef]
- Du, H.; Wang, Z.; Yu, W.; Liu, Y.; Huang, B. Differential metabolic responses of perennial grass Cynodon transvaalensis x Cynodon dactylon (C4) and Poa Pratensis (C3) to heat stress. Physiol. Plant. 2011, 141, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Castander-Olarieta, A.; Montalbán, I.A.; Oliveira, E.M.; Dell’Aversana, E.; D’Amelia, L.; Carillo, P.; Steiner, N.; Fraga, H.P.F.; Guerra, M.P.; Goicoa, T.; et al. Effect of thermal stress on tissue ultrastructure and metabolite profiles during initiation of radiata pine somatic embryogenesis. Front. Plant Sci. 2019, 9, 2004. [Google Scholar] [CrossRef] [PubMed]
- Jancewicz, A.L.; Gibbs, N.M.; Masson, P.H. Cadaverine’s functional role in plant development and environmental response. Front. Plant Sci. 2016, 7, 870. [Google Scholar] [CrossRef] [PubMed]
- García-Calderon, M.; Pérez-Delgado, C.M.; Palove-Balang, P.; Betti, M.; Márquez, A.J. Flavonoids and isoflavonoids biosynthesis in the model legume Lotus japonicus; connections to nitrogen metabolism and photorespiration. Plants 2020, 9, 774. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, Y.; Yun, S.K.; Kim, S.S.; Joa, J.; Moon, Y.E.; Do, G.R. The anatomical differences and physiological responses of sunburned satsuma mandarin (Citrus unshiu Marc.) fruits. Plants 2022, 11, 1801. [Google Scholar] [CrossRef] [PubMed]
- Sicher, R.C. Temperature shift experiments suggest that metabolic impairment and enhanced rates of photorespiration decrease organic acid levels in soybean leaflets exposed to supra-optimal growth temperatures. Metabolites 2015, 5, 443–454. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, Z.; Luo, J.; Zhang, Y.; Jiang, N.; Khattak, W.A. Transcriptome analysis of sugar and acid metabolism in young tomato fruits under high temperature and nitrogen fertilizer influence. Front. Plant Sci. 2023, 14, 1197553. [Google Scholar] [CrossRef]
- Rienth, M.; Torregrosa, L.; Sarah, G.; Ardisson, M.; Brillouet, J.M.; Romieu, C. Temperature desynchronizes sugar and organic acid metabolism in ripening grapevine fruits and remodels their transcriptome. BMC Plant Biol. 2016, 16, 164. [Google Scholar] [CrossRef]
- Raymond, P.; Pradet, A. Stabilization of adenine nucleotide ratio at various values by an oxygen limitation of respiration in germinating lettuce (Lactuca sativa) seeds. Biochem. J. 1980, 190, 39–44. [Google Scholar] [CrossRef]
- Drew, M.C.; Saglio, P.H.; Pradet, A. Larger adenylate energy charge and ATP/ADP ratios in aerenchymatous roots of Zea mays in anaerobic media as a consequence of improved internal oxygen transport. Planta 1985, 165, 51–58. [Google Scholar] [CrossRef]
- Duff, S.M.G.; Moorhead, G.B.G.; Lefebvre, D.D.; Plaxton, W.C. Phosphate starvation inducible ‘bypasses’ of adenylate and phosphate dependent glycolytic enzymes in Brassica nigra suspension cells. Plant Physiol. 1989, 90, 1275–1278. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; Tran, H.T. Metabolic adaptations of phosphate-starved plants. Plant Physiol. 2011, 156, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C. The organization and regulation of plant glycolysis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 185–214. [Google Scholar] [CrossRef] [PubMed]
- Bolwell, G.P. Cyclic AMP, the reluctant messenger in plants. Trend Biochem. Sci. 1995, 20, 492–495. [Google Scholar] [CrossRef]
- Maekawa, M.; Honoki, R.; Ihara, Y.; Sato, R.; Oikawa, A.; Kanno, Y.; Ohta, H.; Seo, M.; Saito, K.; Masuda, S. Impact of the plastidial stringent response in plant growth and stress responses. Nat. Plants 2015, 1, 15167. [Google Scholar] [CrossRef]
- Blanco, E.; Fortunato, S.; Viggiano, L.; de Pinto, M.C. Cyclic AMP: A polyhedral signalling molecule in plants. Int. J. Mol. Sci. 2020, 21, 4862. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | SD_1 | SD_3 | SD_5 | SD_7 | SD_9 |
|---|---|---|---|---|---|
| Lys | 0.20 | 0.37 | 0.34 | 0.28 | 0.27 |
| Ala | 0.18 | 0.32 | 0.38 | 0.31 | 0.27 |
| Pro | 0.05 | 0.23 | 0.38 | 0.36 | 0.34 |
| Tyr | 0.13 | 0.25 | 0.34 | 0.25 | 0.23 |
| Hydroxy Pro | 0.10 | 0.22 | 0.26 | 0.24 | 0.24 |
| Fumarate | −0.33 | −0.40 | −0.49 | −0.48 | −0.47 |
| GMP | −0.26 | −0.30 | −0.47 | −0.44 | −0.48 |
| Citrate | −0.28 | −0.30 | −0.44 | −0.36 | −0.31 |
| GDP | −0.24 | −0.29 | −0.43 | −0.35 | −0.33 |
| Diethanolamine | −0.20 | −0.28 | −0.33 | −0.40 | −0.43 |
| Compounds | AT_1 | AT_3 | AT_5 | AT_7 | AT_9 |
|---|---|---|---|---|---|
| Lys | 0.67 | 0.64 | 0.59 | 0.60 | 0.61 |
| Hydroxy Pro | 0.35 | 0.40 | 0.41 | 0.39 | 0.41 |
| Gly | 0.33 | 0.28 | 0.27 | 0.30 | 0.31 |
| Pro | 0.30 | 0.29 | 0.27 | 0.27 | 0.30 |
| Tyr | 0.16 | 0.17 | 0.25 | 0.33 | 0.41 |
| GDP | −0.27 | −0.30 | −0.36 | −0.45 | −0.48 |
| Citrate | −0.19 | −0.24 | −0.35 | −0.47 | −0.51 |
| Spermine | −0.23 | −0.29 | −0.36 | −0.41 | −0.43 |
| Fumarate | −0.33 | −0.30 | −0.33 | −0.36 | −0.35 |
| NAD | −0.16 | −0.20 | −0.32 | −0.46 | −0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oikawa, A.; Takeuchi, K.; Morita, K.; Horibe, Y.; Sasaki, R.; Murayama, H. Effects of Climate Conditions before Harvest Date on Edamame Metabolome. Plants 2024, 13, 87. https://doi.org/10.3390/plants13010087
Oikawa A, Takeuchi K, Morita K, Horibe Y, Sasaki R, Murayama H. Effects of Climate Conditions before Harvest Date on Edamame Metabolome. Plants. 2024; 13(1):87. https://doi.org/10.3390/plants13010087
Chicago/Turabian StyleOikawa, Akira, Katsutaka Takeuchi, Kei Morita, Yamato Horibe, Ryosuke Sasaki, and Hideki Murayama. 2024. "Effects of Climate Conditions before Harvest Date on Edamame Metabolome" Plants 13, no. 1: 87. https://doi.org/10.3390/plants13010087
APA StyleOikawa, A., Takeuchi, K., Morita, K., Horibe, Y., Sasaki, R., & Murayama, H. (2024). Effects of Climate Conditions before Harvest Date on Edamame Metabolome. Plants, 13(1), 87. https://doi.org/10.3390/plants13010087