



A Set of Molecular Markers to Accelerate Breeding and Determine Seed Purity of CMS Three-Line Hybrids in Brassica napus

Abstract
1. Introduction
2. Results
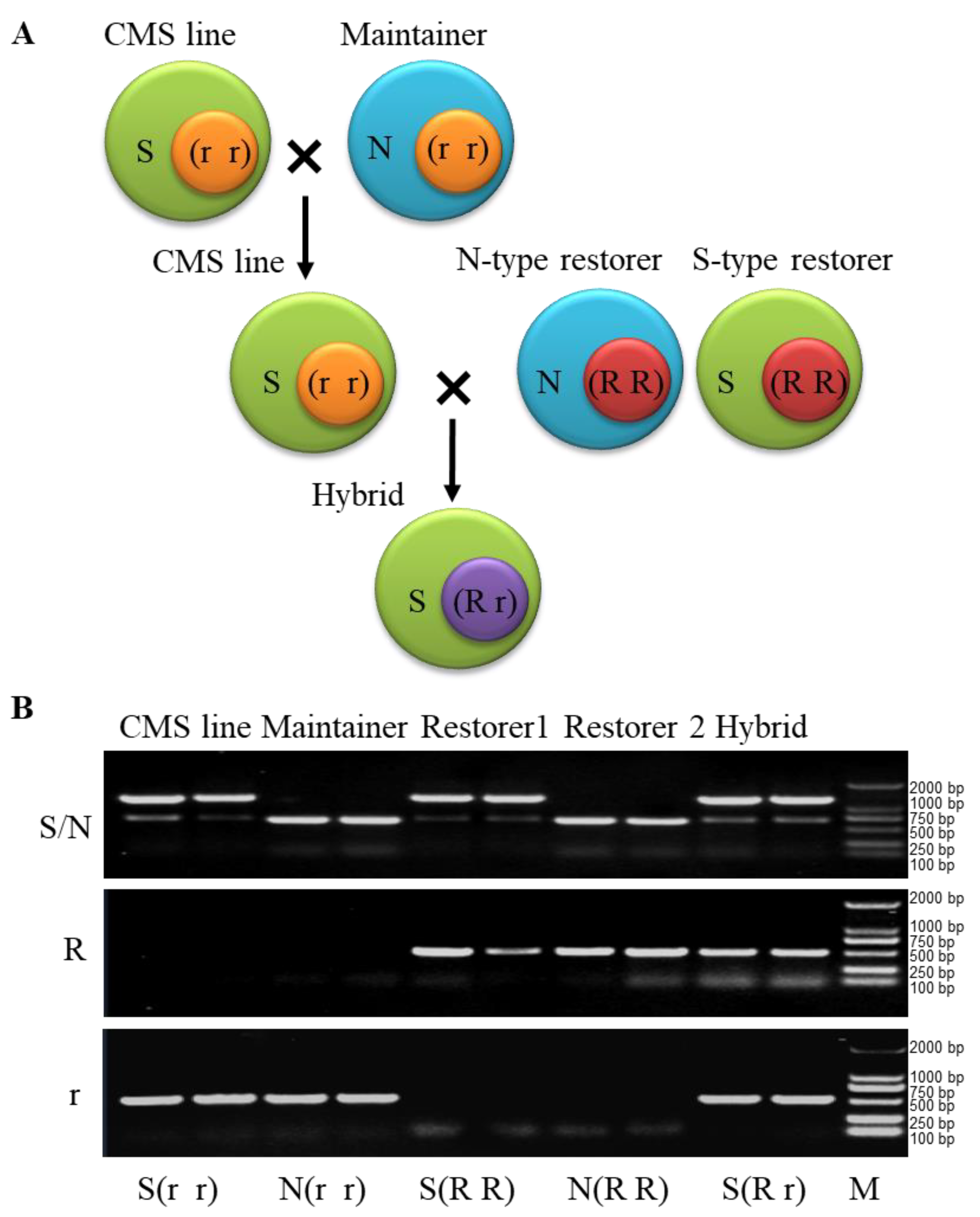
2.1. Developing Cytoplasmic Molecular Markers for N and S Cytoplasm Identification of the Pol and Shaan CMS Three-Line Hybrids
2.2. Developing Cytoplasmic Molecular Markers for N and S Cytoplasm Identification of the Pol and Shaan CMS Three-Line Hybrids
2.3. The Molecular Markers Are Suitable for the Identification of the Three Lines in the Pol- and Shaan-Type CMS Systems of B. Napus
2.4. Identification of Rapeseed Germplasm with the CMS Three-Line Molecular Markers
2.5. Identification of Rapeseed Germplasm with the CMS Three-Line Molecular Markers
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Protocol for High Throughput Screening and Rapid DNA Extraction for Rapeseed
4.3. Primer Design
4.4. PCR Reaction System and Procedure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Yang, Z.; Wang, X.; Li, K.; An, H.; Liu, J.; Yang, G.S.; Fu, T.D.; Yi, B.; Hong, D.F. A Mitochondria-Targeted PPR Protein Restores pol Cytoplasmic Male Sterility by Reducing orf224 Transcript Levels in Oilseed Rape. Mol. Plant 2016, 9, 1082–1084. [Google Scholar] [CrossRef] [PubMed]
- Havlíčková, L.; Čurn, V.; Jozová, E.; Kučera, V.; Klíma, M. Sequence analysis of the mtDNA region correlated with Shaan 2A cytoplasmic male sterility in rapeseed (Brassica napus L.). Czech J. Genet. Plant Breed. 2012, 48, 139–142. [Google Scholar] [CrossRef]
- Wang, Y.F.; Ma, S.M.; Wang, M.; Zheng, X.Q.; Gu, M.; Hu, S.W. Sequence analysis of the gene correlated with cytoplasmic male sterility (CMS) in rape-seed (Brassica napus) Polima and Shaan 2A. Chin. Sci. Bull. 2002, 47, 122–126. [Google Scholar]
- Yu, H.L.; Li, Z.Y.; Yang, L.M.; Liu, Y.M.; Zhuang, M.; Zhang, L.G.; Lv, H.H.; Li, Z.S.; Han, F.Q.; Liu, X.P.; et al. Morphological and molecular characterization of the second backcross progenies of Ogu-CMS Chinese kale and rapeseed. Euphytica 2017, 213, 55. [Google Scholar] [CrossRef]
- Wang, C.D.; Lezhneva, L.; Arnal, N.; Quadrado, M.; Mireau, H. The radish Ogura fertility restorer impedes translation elongation along its cognate CMS-causing mRNA. Proc. Natl. Acad. Sci. USA 2021, 118, e2105274118. [Google Scholar] [CrossRef] [PubMed]
- Engelke, T.; Hirsche, J.; Roitsch, T. Metabolically engineered male sterility in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2011, 122, 163–174. [Google Scholar] [CrossRef]
- Bhatia, R.; Dey, S.S.; Sharma, K.; Singh, S.; Kumar, S.; Pramanik, A.; Parkash, C.; Kumar, R. Back-cross introgression of ‘Tour’ cytoplasm from Brassica napus through in vitro embryo rescue reveals partial restoration of sterility in B. oleracea. Sci. Hortic. 2021, 282, 110014. [Google Scholar] [CrossRef]
- Wan, Z.J.; Bing, J.; Tu, J.X.; Ma, C.Z.; Shen, J.X.; Yi, B.; Wen, J.; Huang, T.; Wang, X.J.; Fu, T.D. Genetic characterization of a new cytoplasmic male sterility system (hau) in Brassica juncea and its transfer to B. napus. Theor. Appl. Genet. 2008, 116, 355–362. [Google Scholar] [CrossRef]
- Liu, J.M.; Li, M.T.; Wang, H.; Yu, L.J.; Li, D.R. Sequence analysis and expression of orf224 gene associated with two types of cytoplasmic male sterility in Brassica napus L. Z. Nat. C J. Biosci. 2010, 65, 395–402. [Google Scholar] [CrossRef]
- Fu, T.; Yang, G.S.; Yang, X.N. Studies on “Three Line” Polima Cytoplasmic Male Sterility Developed in Brassica napus L. Plant Breed. 2010, 104, 115–120. [Google Scholar]
- Ning, L.Y.; Wang, H.; Li, D.R.; Lin, Z.W.; Li, Y.H.; Zhao, W.G.; Chao, H.B.; Miao, L.Y.; Li, M.T. Transcriptomic and Proteomic Analysis of Shaan2A Cytoplasmic Male Sterility and Its Maintainer Line in Brassica napus. Front. Plant Sci. 2019, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef]
- Bohra, A.; Jha, U.C.; Adhimoolam, P.; Bisht, D.; Singh, N.P. Cytoplasmic male sterility (CMS) in hybrid breeding in field crops. Plant Cell Rep. 2016, 35, 967–993. [Google Scholar] [CrossRef]
- Liu, Z.W.; Fu, T.D.; Tu, J.X.; Chen, B.Y. Inheritance of seed colour and identification of RAPD and AFLP markers linked to the seed colour gene in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2005, 110, 303–310. [Google Scholar]
- Li, Y.; Liu, Z.; Cai, Q.; Yang, G.S.; He, Q.B.; Liu, P.W. Identification of a SSR marker linked to the fertility-restoring gene for polima cytoplasmic male sterile line in Brassica napus. Afr. J. Biotechnol. 2011, 10, 9563–9569. [Google Scholar]
- Zeng, F.Q.; Yi, B.; Tu, J.X.; Fu, T.D. Identification of AFLP and SCAR markers linked to the male fertility restorer gene of pol CMS (Brassica napus L.). Euphytica 2009, 165, 363–369. [Google Scholar] [CrossRef]
- Wang, B.Q.; Farooq, Z.; Chu, L.; Liu, J.; Wang, H.D.; Guo, J.; Tu, J.X.; Ma, C.Z.; Dai, C.; Wen, J.; et al. High-generation near-isogenic lines combined with multi-omics to study the mechanism of polima cytoplasmic male sterility. BMC Plant Biol. 2021, 21, 130. [Google Scholar] [CrossRef]
- L’Homme, Y.; Stahl, R.J.; Li, X.Q.; Hameed, A.; Brown, G.G. Brassica nap cytoplasmic male sterility is associated with expression of a mtDNA region containing a chimeric gene similar to the pol CMS-associated orf224 gene. Curr. Genet. 1997, 31, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Brown, G.G. Suppression of Cytoplasmic Male Sterility by Nuclear Genes Alters Expression of a Novel Mitochondrial Gene Region. Plant Cell 1991, 3, 1349–1362. [Google Scholar]
- An, H.; Yang, Z.H.; Yi, B.; Wen, J.; Shen, J.X.; Tu, J.X.; Ma, C.Z.; Fu, T.D. Comparative transcript profiling of the fertile and sterile flower buds of pol CMS in B. napus. BMC Genom. 2014, 15, 258. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Yang, G.S.; Fu, T.D.; Li, Y. Transcriptional control of orf224/atp6 by the pol CMS restorer Rfp gene in Brassica napus L. Yi Chuan Xue Bao 2003, 30, 469–473. [Google Scholar]
- Barkan, A.; Small, I. Pentatricopeptide Repeat Proteins in Plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H. Map-Based Cloning and Function Analysis of the Restorer Gene of POLIMA Cytoplasmic Male Sterility in Brassica napus; Huazhong Agricultural University: Wuhan, China, 2016. [Google Scholar]
- Liao, Z.Q.; Kuang, C.G.; Xu, L.F.; Zhou, C.Y. The Main Styles of Cytoplasmic Male Sterility in Brassica napus L. and Their Applications in China. Chin. Agric Sci. Bull. 2010, 26, 105–110. [Google Scholar]
- Havlickova, L.; Jozová, E.; Rychlá, A.; Klíma, M.; Kučera, V.; Čurn, V. Genetic Diversity Assessment in Winter Oilseed Rape (Brassica napus L.) Collection Using AFLP, ISSR and SSR Markers. Czech J. Genet. Plant Breed. 2014, 20, 216–225. [Google Scholar] [CrossRef]
- Koutu, G.; Shrivastava, A.; Samaiya, R.; Mishra, D. Morphological characterization and assessment of genetic purity of rice hybrids using SSR markers. Oryza 2016, 53, 21–26. [Google Scholar]
- Zhang, W.; Shi, H.R.; Zhou, Y.; Liang, X.Y.; Luo, X.; Xiao, C.; Li, Y.; Xu, P.; Wang, J.S.; Gong, W.Z.; et al. Rapid and Synchronous Breeding of Cytoplasmic Male Sterile and Maintainer Line Through Mitochondrial DNA Rearrangement Using Doubled Haploid Inducer in Brassica napus. Front. Plant Sci. 2022, 13, 871006. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Guan, C.Y.; Guan, M. Comparative Cytological and Transcriptome Analyses of Anther Development in Nsa Cytoplasmic Male Sterile (1258A) and Maintainer Lines in Brassica napus Produced by Distant Hybridization. Int. J. Mol. Sci. 2022, 23, 2004. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Andersen, S.B.; Dixelius, C.; Hansen, L.N. Production of fertile intergeneric somatic hybrids between B. napus and S. arvensis for the enrichment of rapeseed gene pool. Plant Cell Rep. 2002, 21, 147–152. [Google Scholar]
- Yamagishi, H.; Bhat, S.R. Cytoplasmic male sterility in Brassicaceae crops. Breed. Sci. 2014, 64, 38–47. [Google Scholar] [CrossRef]
- Ning, L.Y.; Wang, H.; Li, D.R.; Li, Y.H.; Chen, K.; Chao, H.B.; Li, H.X.; He, J.J.; Li, M.T. Genome-wide identification of the restorer-of-fertility-like (RFL) gene family in Brassica napus and expression analysis in shaan2A cytoplasmic male sterility. BMC Genom. 2020, 21, 765. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) | Base Number |
|---|---|---|
| SN-F | TTCATACGGCGAGAGTCATTG | 22 bp |
| N-R | CAAGACCATAGAATAGGAGAACCAC | 22 bp |
| S-R | GCTCGTTCGGCTACTTATCTTG | 22 bp |
| R-F | GGGATGCGATCCTGATATTTG | 21 bp |
| r-F | GGGATGCGATCCTGATATCG | 20 bp |
| Rr-R | CTCCAAAAGGACCAGAAAGCA | 21 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; An, R.; Song, M.; Xie, C.; Wei, S.; Wang, D.; Dong, Y.; Jia, Q.; Huang, S.; Mu, J. A Set of Molecular Markers to Accelerate Breeding and Determine Seed Purity of CMS Three-Line Hybrids in Brassica napus. Plants 2023, 12, 1514. https://doi.org/10.3390/plants12071514
Zhang Y, An R, Song M, Xie C, Wei S, Wang D, Dong Y, Jia Q, Huang S, Mu J. A Set of Molecular Markers to Accelerate Breeding and Determine Seed Purity of CMS Three-Line Hybrids in Brassica napus. Plants. 2023; 12(7):1514. https://doi.org/10.3390/plants12071514
Chicago/Turabian StyleZhang, Yanfeng, Ran An, Min Song, Changgen Xie, Shihao Wei, Daojie Wang, Yuhong Dong, Qingli Jia, Shuhua Huang, and Jianxin Mu. 2023. "A Set of Molecular Markers to Accelerate Breeding and Determine Seed Purity of CMS Three-Line Hybrids in Brassica napus" Plants 12, no. 7: 1514. https://doi.org/10.3390/plants12071514
APA StyleZhang, Y., An, R., Song, M., Xie, C., Wei, S., Wang, D., Dong, Y., Jia, Q., Huang, S., & Mu, J. (2023). A Set of Molecular Markers to Accelerate Breeding and Determine Seed Purity of CMS Three-Line Hybrids in Brassica napus. Plants, 12(7), 1514. https://doi.org/10.3390/plants12071514