


Effects of Plant Biostimulation Time Span and Soil Electrical Conductivity on Greenhouse Tomato ‘Miniplum’ Yield and Quality in Diverse Crop Seasons

 ,
, 

 ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
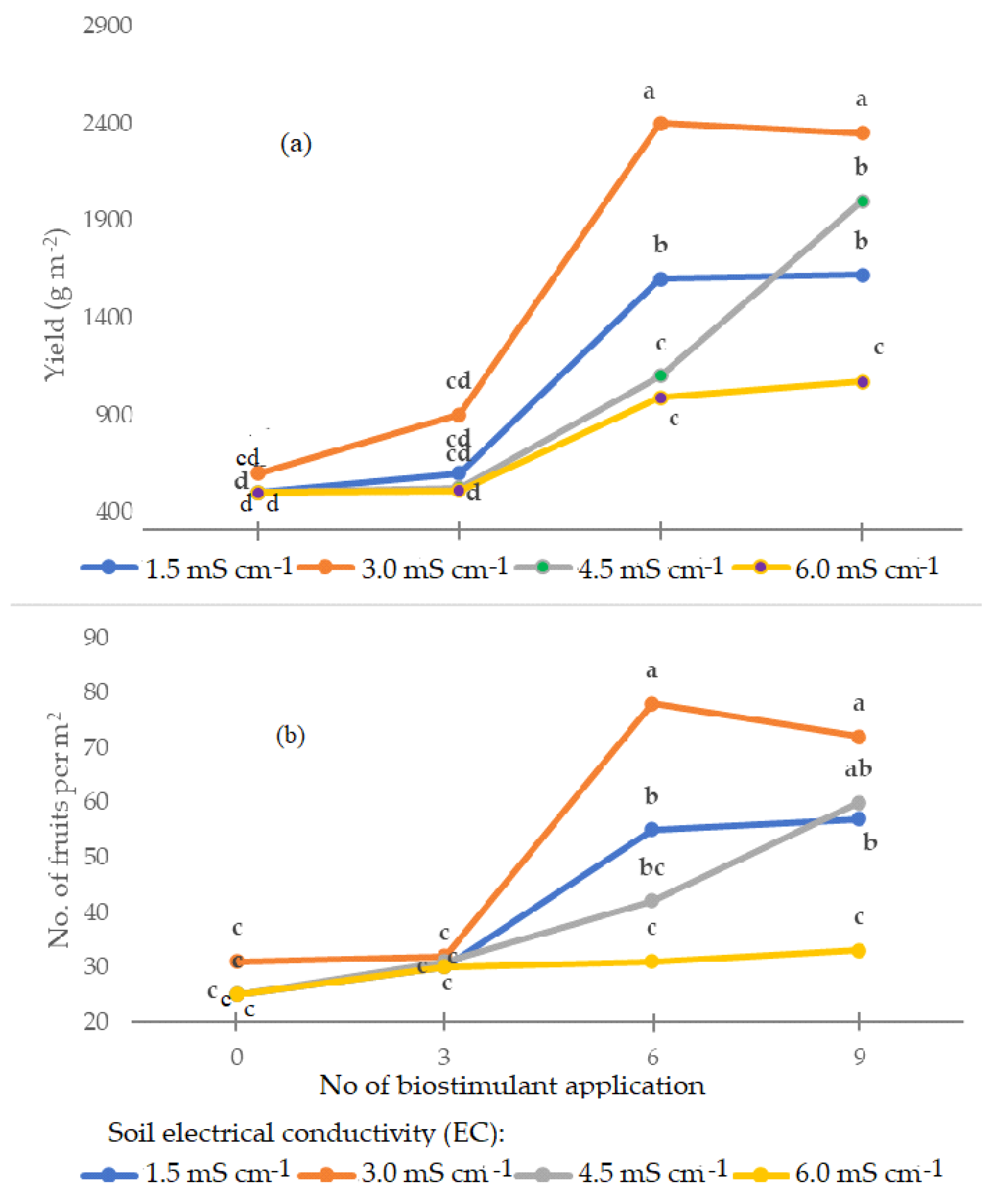
2.1. Yield Parameters
2.2. Quality and Phytochemical Parameters
2.3. Elemental Composition
3. Materials and Methods
3.1. Experimental Protocol and Growing Conditions
3.2. Determinations of Yield, Fruit Quality, Colour, Antioxidant Compounds and Activity, and Elemental Composition
3.3. Data Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kole, C. Genome Mapping and Molecular Breeding in Plants; Volume 5: Vegetables; Springer: Berlin/Heidelberg, Germany, 2007; p. 375. [Google Scholar]
- De Sio, F.; Rapacciuolo, M.; De Giorgi, A.; Sandei, L.; Giuliano, B.; Tallarita, A.; Golubkina, N.; Sekara, A.; Stoleru, V.; Cuciniello, A.; et al. Industrial processing affects product yield and quality of diced tomato. Agriculture 2021, 11, 230. [Google Scholar] [CrossRef]
- Ippolito, A.; Nigro, F.; Schenna, L. Control of postharvest diseases of fresh vegetable by application of antagonistic micro-organism. In Crop Management and Postharvest Handling of Horticultural Products; Dris, R., Niskanen, R., Jai, S.M., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2003; pp. 1–30. [Google Scholar]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Oliveira, S.M.; Brandao, T.R.S.; Silva, C.L.M. Influence of drying processes and pretreatments on nutritional and bioactive characteristics of dried vegetables: A review. Food Eng. Rev. 2016, 8, 134–163. [Google Scholar] [CrossRef]
- Causse, M.; Friguet, C.; Coiret, C.; Lépicier, M.; Navez, B.; Lee, M.; Holthuysen, L.; Sinesio, F.; Moneta, E.; Grandillo, S. Consumer preferences for fresh tomato at the European scale: A common segmentation on taste and firmness. J. Food Sci. 2010, 75, S531–S541. [Google Scholar] [CrossRef]
- Siddiqui, M.W.; Ayala-Zavala, J.F.; Dhua, R.S. Genotypic variation intomatoes affecting processing and antioxidant properties. Crit. Rev. Food Sci. Nutr. 2015, 55, 1819–1835. [Google Scholar] [CrossRef]
- Ghorbanli, M.; Gafarabad, M.; Amirkian, T.; Mamaghani, B. Investigation of proline, total protein, chlorophyll, ascorbate and dehydroascorbate changes under drought stress in Akria and Mobil tomato cultivars. Iran. J. Plant Phys. 2013, 3, 651–659. [Google Scholar]
- Sibomana, I.C.; Aguyoh, J.N.; Opiyo, A.M. Water stress affects growth and yield of container grown tomato (Lycopersicon esculentum M.). Glob. J. Biosci. Biotechnol. 2013, 2, 461–466. [Google Scholar]
- Niu, G.; Rodriguez, D.; Dever, J.; Zhang, J. Growth and physiological responses of five cotton genotypes to sodium chloride and sodium sulfate saline water irrigation. J. Cotton. Sci. 2013, 17, 233–244. [Google Scholar]
- Ghonaim, M.M.; Mohamed, H.I.; Omran, A.A. Evaluation of wheat (Triticum aestivum L.) salt stress tolerance using physiological parameters and retrotransposon-based markers. Genet. Resour. Crop. Evol. 2021, 68, 227–242. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Akladious, S.A.; El-Beltagi, H.S. Mitigation the harmful effect of salt stress on physiological, biochemical and anatomical traits by foliar spray with trehalose on wheat cultivars. Fresenius Environ. Bull. 2018, 27, 7054–7065. [Google Scholar]
- Tuna, A.L.; Kaya, C.; Ashraf, M.; Altunlu, H.; Yokas, I.; Yagmono, B. The effects of calcium sulphate on growth, membrane stability and nutrient uptake of tomato plants grown under salt stress. Environ. Exp. Bot. 2007, 59, 173–178. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. An alternative use of horticultural crops: Stressed plants as biofactories of bioactive phenolic compounds. Agriculture 2012, 2, 259–271. [Google Scholar] [CrossRef]
- Rozema, J.; Schat, H. Salt tolerance of halophytes, research questions reviewed in the perspective of saline agriculture. Environ. Exp. Bot. 2013, 92, 83–95. [Google Scholar] [CrossRef]
- Huygens, D.; Saveyn, H.G.M.; Tonini, D.; Eder, P.; Delgado Sancho, L. Technical Proposals for Selected New Fertilising Materials under the Fertilising Products Regulation (Regulation (EU) 2019/1009)—Process and Quality Criteria, and Assessment of Environmental and Market Impacts for Precipitated Phosphate Salts & Derivate; EUR 29841 EN; Publications Office of the European Union: Luxembourg, 2019; ISBN 978-92-76-09888-1. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Mannino, G.; Beekwilder, J.; Contartese, V.; Karlova, R.; Bertea, C.M. The application of a biostimulant based on tannins affects root architecture and improves tolerance to salinity in tomato plants. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Domingo, G.; Marsoni, M.; Álvarez-Viñas, M.; Torres, M.D.; Domínguez, H.; Vannini, C. The Role of Protein-Rich Extracts from Chondrus crispus as Biostimulant and in Enhancing Tolerance to Drought Stress in Tomato Plants. Plants 2023, 12, 845. [Google Scholar] [CrossRef]
- Francesca, S.; Cirillo, V.; Raimondi, G.; Maggio, A.; Barone, A.; Rigano, M.M. A Novel Protein Hydrolysate-Based Biostimulant Improves Tomato Performances under Drought Stress. Plants 2021, 10, 783. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Bonini, P.; Colla, G.; Rouphael, Y. Yield and nutritional quality of Vesuvian Piennolo Tomato PDO as affected by farming system and biostimulant application. Agronomy 2019, 9, 505. [Google Scholar] [CrossRef]
- Zuluaga, M.Y.A.; Monterisi, S.; Rouphael, Y.; Colla, G.; Lucini, L.; Cesco, S.; Pii, Y. Different vegetal protein hydrolysates distinctively alleviate salinity stress in vegetable crops: A case study on tomato and lettuce. Front. Plant Sci. 2023, 14, 1077140. [Google Scholar] [CrossRef]
- Patwary, M.; Rahman, M.; Ahmad, S.; Miah, M.; Rahman, M. Performance of some tomato (Solanum lycopersicon L.) genotypes in summer and winter seasons. Agriculturists 2014, 12, 64–73. [Google Scholar] [CrossRef]
- Sen, S.; Mukherji, S. Season-controlled changes in biochemical constituents and oxidase enzyme activities in tomato (Lycopersicon esculentum Mill.). J. Environ. Biol. 2009, 30, 479–483. [Google Scholar]
- Rajametov, S.N.; Lee, K.; Jeong, H.B.; Cho, M.C.; Nam, C.W.; Yang, E.Y. Physiological traits of thirty-five tomato accessions in response to low temperature. Agriculture 2021, 11, 792. [Google Scholar] [CrossRef]
- Guo, X.; Liu, D.; Chong, K. Cold signaling in plants: Insights into mechanisms and regulation. J. Integr. Plant Biol. 2018, 60, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Geraldine, A.; Junior, A.; Gastmann, R.; Adamski, J.M.; Sperotto, R.A. Root responses of contrasting rice genotypes to low temperature stress. J. Plant Physiol. 2020, 255, 153307. [Google Scholar]
- Sun, S.J.; Li, J.M.; Zong, J.W.; Yao, Y.Z.; Chen, K.L. Effects of sub-low temperature and drought stress on root morphology and leaf structure of tomato seedlings. Chin. J. Appl. Ecol. 2012, 23, 3027. [Google Scholar]
- Niu, C.; Wang, G.; Sui, J.; Liu, G.; Ma, F.; Bao, Z. Biostimulants alleviate temperature stress in tomato seedlings. Sci. Hort. 2022, 293, 110712. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M.; et al. A combined phenotypic and metabolomic approach for elucidating the biostimulant action of a plant-derived protein hydrolysate on tomato grown under limited water availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The effect of a plant-derived biostimulant on metbolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Francesca, S.; Arena, C.; Hay Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The use of a plant-based biostimulant improves plant performances and fruit quality in tomato plants grown at elevated temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef]
- Tardieu, F.; Simonneau, T.; Muller, B. The physiological basis of drought tolerance in crop plants: A scenario-dependent probabilistic approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H. Adverse high temperature effects on pollen viability, seed-set, seed yield and harvest index of grain-sorghum [Sorghum bicolor (L.) Moench] are more severe at elevated carbon dioxide due to higher tissue temperatures. Agric. For. Meteorol. 2006, 139, 237–251. [Google Scholar] [CrossRef]
- Dong, B.; Zheng, X.; Liu, H.; Able, J.A.; Yang, H.; Zhao, H.; Zhang, M.; Qiao, Y.; Wang, Y.; Liu, M. Effects of drought stress on pollen sterility, grain yield, abscisic acid and protective enzymes in two winter wheat cultivars. Front. Plant Sci. 2017, 8, 1008. [Google Scholar] [CrossRef] [PubMed]
- Paupière, M.J.; van Haperen, P.; Rieu, I.; Visser, R.G.F.; Tikunov, Y.M.; Bovy, A.G. Screening for pollen tolerance to high temperatures in tomato. Euphytica 2017, 213, 1–8. [Google Scholar] [CrossRef]
- Sonneveld, C.; Vanderburg, A.M.M. Sodium-chloride salinity in fruit vegetable crops in soilless culture. Neth. J. Agr. Sci. 1991, 39, 115–122. [Google Scholar] [CrossRef]
- Urrea-López, R.; Díaz de la Garza, R.I.; Valiente-Banuet, J.I. Effects of substrate salinity and nutrient levels on physiological response, yield, and fruit quality of habanero pepper. Hort. Sci. 2014, 49, 812–818. [Google Scholar] [CrossRef]
- Zaki, H.E.M.; Yokoi, S. A comparative in vitro study of salt tolerance in cultivated tomato and related wild species. Plant Biotechnol. 2016, 33, 1–12. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Akram, N.A.; Saleem, M.H.; Shafiq, S.; Naz, H.; Farid-ul-Haq, M.; Ali, B.; Shafiq, F.; Iqbal, M.; Jaremko, M.; Qureshi, K.A. Phytoextracts as Crop Biostimulants and Natural Protective Agents—A Critical Review. Sustainability 2022, 14, 14498. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the Regulation of Reactive Oxygen Species Metabolism in Plants under Abiotic Stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Sarwar, M.I. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar]
- Shelford, V.E. Some concepts of bioecology. Ecology 1931, 12, 455–467. [Google Scholar] [CrossRef]
- Adams, P.; Ho, L.C. Effects of constant and fluctuating salinity on the yield, quality and calcium status of tomatoes. J. Hortic. Sci. 1989, 64, 725–732. [Google Scholar] [CrossRef]
- Savvas, D.; Stamati, E.; Tsirogiannis, I.L.; Mantzos, N.; Barouchas, P.E.; Katsoulas, N.; Kittas, C. Interactions between salinity and irrigation frequency in greenhouse pepper grown in closedcycle hydroponic systems. Agric. Water Manag. 2007, 91, 102–111. [Google Scholar] [CrossRef]
- Tadesse, T.; Nichols, M.A.; Fisher, K.J. Nutrient conductivity effects on sweet pepper plants grown using a nutrient film technique 1. Yield and fruit quality. N. Z. J. Crop. Hortic. Sci. 1999, 27, 141–148. [Google Scholar]
- Sharaf, A.R.; Hobson, G.E. Effect of salinity on the yield and quality of normal and non-ripening mutant tomatoes. Acta Hortic. 1986, 190, 175–182. [Google Scholar] [CrossRef]
- Meza, S.L.; Egea, I.; Massaretto, I.L.; Morales, B.; Purgatto, E.; Egea-Fernández, J.M.L. Traditional tomato varieties improve fruit quality without affecting fruit yield under moderate salt stress. Front. Plant Sci. 2020, 11, 1717. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Patanè, C. Effect of application of biostimulants on the biomass, nitrate, pigments, and antioxidants content in radish and turnip microgreens. Agronomy 2023, 13, 145. [Google Scholar] [CrossRef]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Luts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef]
- Riggi, E.; Patané, C.; Ruberto, G. Content of carotenoids at different ripening stages in processing tomato in relation to soil water availability. Aust. J. Agric. Res. 2008, 59, 348–353. [Google Scholar] [CrossRef]
- Ali, M.M.; Jeddi, K.; Attia, M.S.; Elsayed, S.M.; Yusuf, M.; Osman, M.S.; Soliman, M.-H.; Hessini, K. Wuxal amino (Bio stimulant) improved growth and physiological performance of tomato plants under salinity stress through adaptive mechanisms and antioxidant potential. Saudi J. Biol. Sci. 2021, 28, 3204–3213. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid—A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 2018. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Abedin, M.M.; Singh, A.K.; Das, S. Role of phenolic compounds in plant-defensive mechanisms. In Plant Phenolics in Sustainable Agricuoture; Chapter 22; Lone, R., Ed.; Springer: Singapore, 2020; pp. 517–532. [Google Scholar] [CrossRef]
- Ciudad-Mulero, M.; Pinela, J.; Carvalho, A.M.; Barros, L.; Fernández-Ruiz, V.; Ferreira, I.C.F.R.; Sánchez-Mata, M.d.C.; Morales, P. Bioaccessibility of Macrominerals and Trace Elements from Tomato (Solanum lycopersicum L.) Farmers’ Varieties. Foods 2022, 11, 1968. [Google Scholar] [CrossRef] [PubMed]
- Hernández Suárez, M.; Rodríguez Rodríguez, E.M.; Díaz Romero, C. Mineral and trace element concentrations in cultivars of tomatoes. Food Chem. 2007, 104, 489–499. [Google Scholar] [CrossRef]
- Guo, M.; Wang, X.-S.; Guo, H.-D.; Bai, S.-Y.; Khan, A.; Wang, X.-M.; Gao, Y.-M.; Li, J.-S. Tomato salt tolerance mechanisms and their potential applications for fighting salinity: A review. Front. Plant Sci. 2022, 13, 949541. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, R.L.; Page, A.L.; Haghnia, G.H. Effect of salinity and its composition on the accumulation of selenium by alfalfa. Plant Soil 1988, 107, 63–67. [Google Scholar] [CrossRef]
- Schiavon, M.; Nardi, S.; Dalla Vecchia, F.; Ertani, A. Selenium biofortification in the 21st century: Status and challenges for healthy human nutrition. Plant Soil 2020, 453, 245–270. [Google Scholar] [CrossRef]
- Francesco De Sio, F.; Servillo, L.; Loiudice, R.; Castaldo, D. A chromatographic procedure for the determination of carotenoids and chlorophylls in vegetable products. Acta Aliment. 2001, 30, 395–405. [Google Scholar] [CrossRef]
- Golubkina, N.A.; Kosheleva, O.V.; Krivenkov, L.V.; Dobrutskaya, H.G.; Nadezhkin, S.; Caruso, G. Intersexual differences in plant growth, yield, mineral composition and antioxidants of spinach (Spinacia oleracea L.) as affected by selenium form. Sci. Hortic. 2017, 225, 350–358. [Google Scholar] [CrossRef]
- AOAC Association Official Analytical Chemists. The Official Methods of Analysis of AOAC International; 22 Vitamin C; AOAC: Rockville, MD, USA, 2012. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. LWT–Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hort. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Alfthan, G.V. A micromethod for the determination of selenium in tissues and biological fluids by single-test-tube fluorimetry. Anal. Chim. Acta 1984, 165, 187–194. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Autumn-Winter | |||
|---|---|---|---|
| Experimental Treatment | Yield (kg·m−2) | No. Fruits per Plant | Mean Weight (g) |
| Electrical conductivity (EC, mS·cm−1) | |||
| 1.5 | 1.72 c | 18.0 b | 24.1 c |
| 3.0 | 2.18 b | 19.9 b | 27.5 b |
| 4.5 | 2.64 a | 23.1 a | 28.6 a |
| 6.0 | 2.11 b | 19.3 b | 27.3 b |
| Biostimulant applications (B) | |||
| 0 | 1.85 b | 17.1 c | 26.9 b |
| 3 | 2.57 a | 24.5 a | 26.3 b |
| 6 | 2.49 a | 22.1 b | 28.0 a |
| 9 | 1.75 b | 16.7 c | 26.3 b |
| Spring–Summer | |||
|---|---|---|---|
| Experimental Treatment | Dry Residue (%) | Soluble Solids (°Brix) | Firmness (Kg·cm−1) |
| Electrical conductivity (EC, mS·cm−1) | |||
| 1.5 | 10.6 c | 9.0 c | 0.61 a |
| 3.0 | 11.0 bc | 9.5 bc | 0.63 a |
| 4.5 | 11.7 b | 10.2 ab | 0.64 a |
| 6.0 | 12.6 a | 10.5 a | 0.65 a |
| Biostimulant applications (B) | |||
| 0 | 10.6 c | 9.3 b | 0.61 a |
| 3 | 11.5 ab | 9.7 ab | 0.64 a |
| 6 | 12.1 a | 10.0 a | 0.65 a |
| 9 | 11.0 bc | 9.6 ab | 0.63 a |
| Autumn–Winter | |||
| Electrical conductivity (EC, mS·cm−1) | |||
| 1.5 | 6.4 b | 6.38 c | 0.75 c |
| 3.0 | 6.8 b | 6.77 b | 0.81 b |
| 4.5 | 7.2 a | 7.15 a | 0.85 ab |
| 6.0 | 7.3 a | 7.27 a | 0.87 a |
| Biostimulant applications (B) | |||
| 0 | 6.1 b | 6.11 c | 0.73 c |
| 3 | 7.1 a | 6.93 b | 0.82 b |
| 6 | 7.1 a | 7.29 a | 0.90 a |
| 9 | 5.5 c | 7.24 a | 0.83 b |
| Spring–Summer | |||
|---|---|---|---|
| Experimental Treatment | L* | A* | B* |
| Electrical conductivity (EC, mS·cm−1) | |||
| 1.5 | 40.2 a | 32.1 a | 19.6 a |
| 3.0 | 39.1 a | 31.2 a | 19.3 a |
| 4.5 | 39.1 a | 32.1 a | 20.4 a |
| 6.0 | 39.5 a | 31.2 a | 19.1 a |
| Biostimulant applications (B) | |||
| 0 | 40.0 a | 34.5 a | 20.2 a |
| 3 | 39.8 a | 32.6 a | 21.0 a |
| 6 | 39.2 a | 31.7 ab | 18.3 b |
| 9 | 38.6 a | 28.9 b | 17.7 b |
| Autumn–Winter | |||
| Electrical conductivity (EC, mS·cm−1) | |||
| 1.5 | 39.1 a | 31.6 a | 19.5 a |
| 3.0 | 39.3 a | 31.8 a | 19.6 a |
| 4.5 | 39.4 a | 32.1 a | 19.8 a |
| 6.0 | 39.7 a | 32.2 a | 20.1 a |
| Biostimulant applications (B) | |||
| 0 | 38.0 a | 32.6 a | 22.5 a |
| 3 | 37.5 a | 30.5 a | 22.1 a |
| 6 | 38.8 a | 27.8 ab | 21.5 ab |
| 9 | 39.4 a | 24.2 b | 18.5 b |
| Spring–Summer | |||||
|---|---|---|---|---|---|
| Experimental Treatment | Lycopene | Total Polyphenols | Ascorbic Acid | LAA | HAA |
| (mg·100 g−1 f.w.) | (mg gallic Acid eq 100 g−1 f.w.) | (mg·100 g−1 f.w.) | (mM Trolox eq 100 g−1 d.w.) | (mM Ascorbic Acid eq 100 g−1 d.w.) | |
| Electrical conductivity (EC, mS·cm−1) | |||||
| 1.5 | 2044.7 b | 2.2 b | 24.2 c | 24.12 c | 6.5 a |
| 3.0 | 2159.3 ab | 2.4 ab | 27.9 bc | 27.35 bc | 7.18 a |
| 4.5 | 2363.3 ab | 2.6 ab | 32.4 ab | 31.81 ab | 7.43 a |
| 6.0 | 2580.7 a | 2.7 a | 35.0 a | 35.86 a | 7.64 a |
| Biostimulant applications (B) | |||||
| 0 | 2003.3 b | 2.3 b | 26.9 a | 22.51 b | 5.0 b |
| 3 | 2455.3 a | 2.5 ab | 30.2 a | 30.34 a | 7.56 a |
| 6 | 2294.7 ab | 2.5 ab | 30.8 a | 32.85 a | 7.92 a |
| 9 | 2394.7 a | 2.6 a | 31.6 a | 33.92 a | 8.26 a |
| Autumn–Winter | |||||
| Electrical conductivity (EC, mS·cm−1) | |||||
| 1.5 | 1063.1 b | 1.68 c | 42.4 b | 14.54 a | 8.68 a |
| 3.0 | 1290.2 a | 1.89 c | 45.5 ab | 16.70 a | 9.19 a |
| 4.5 | 1271.2 a | 2.40 b | 48.0 a | 16.95 a | 9.28 a |
| 6.0 | 1211.1 a | 2.82 a | 48.9 a | 17.35 a | 9.31 a |
| Biostimulant applications (B) | |||||
| 0 | 1050.6 b | 1.66 c | 35.4 b | 7.45 c | 6.15 b |
| 3 | 1112.0 b | 2.12 b | 49.1 a | 8.03 c | 6.17 b |
| 6 | 1253.9 ab | 2.39 ab | 49.9 a | 23.15 b | 12.14 a |
| 9 | 1419.1 a | 2.62 a | 50.5 a | 26.90 a | 12.01 a |
| Spring–Winter | |||||
|---|---|---|---|---|---|
| Experimental Treatment | P | Ca | K | Mg | Na |
| (mg·kg−1 f.w.) | (mg·kg−1 f.w.) | (mg·kg−1 f.w.) | (mg·kg−1 f.w.) | (mg·kg−1 f.w.) | |
| Electrical conductivity (EC, mS·cm−1) | |||||
| 1.5 | 449.2 ab | 110.4 a | 2638 a | 163.0 a | 104.6 a |
| 3.0 | 468.8 a | 109.2 a | 2740 a | 172.3 a | 105.5 a |
| 4.5 | 488.3 a | 107.0 a | 2889 a | 184.5 a | 108.5 a |
| 6.0 | 381.0 b | 106.7 a | 2434 a | 116.7 b | 109.6 a |
| Biostimulant applications (B) | |||||
| 0 | 414.6 a | 110.7 a | 2702 a | 155.2 a | 105.1 a |
| 3 | 438.7 a | 108.9 a | 2684 a | 158.3 a | 106.5 a |
| 6 | 463.6 a | 107.0 a | 2667 a | 160.9 a | 108.1 a |
| 9 | 470.5 a | 106.7 a | 2648 a | 162.2 a | 108.5 a |
| Spring–Winter | |||||
|---|---|---|---|---|---|
| Experimental Treatment | Fe | Zn | Cu | Se | Mn |
| (mg·kg−1 f.w.) | (mg·kg−1 f.w.) | (mg·kg−1 f.w.) | (µg·kg−1 f.w.) | (mg·kg−1 f.w.) | |
| Electrical conductivity (EC, mS·cm−1) | |||||
| 1.5 | 3.89 a | 0.66 a | 233.7 a | 129 a | 157.9 a |
| 3.0 | 3.94 a | 0.68 a | 219.4 a | 121 a | 155.8 a |
| 4.5 | 4.00 a | 0.71 a | 194.0 ab | 111 ab | 151.8 a |
| 6.0 | 4.03 a | 0.72 a | 159.4 b | 97 b | 146.8 a |
| Biostimulant applications (B) | |||||
| 0 | 4.43 a | 0.76 a | 258.4 a | 104 a | 98.5 c |
| 3 | 4.17 ab | 0.74 a | 238.7 a | 108 a | 135.2 b |
| 6 | 3.83 ab | 0.68 ab | 190.5 b | 120 a | 181.3 a |
| 9 | 3.43 b | 0.60 b | 118.9 c | 125 a | 197.3 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tallarita, A.V.; Vecchietti, L.; Golubkina, N.A.; Sekara, A.; Cozzolino, E.; Mirabella, M.; Cuciniello, A.; Maiello, R.; Cenvinzo, V.; Lombardi, P.; et al. Effects of Plant Biostimulation Time Span and Soil Electrical Conductivity on Greenhouse Tomato ‘Miniplum’ Yield and Quality in Diverse Crop Seasons. Plants 2023, 12, 1423. https://doi.org/10.3390/plants12071423
Tallarita AV, Vecchietti L, Golubkina NA, Sekara A, Cozzolino E, Mirabella M, Cuciniello A, Maiello R, Cenvinzo V, Lombardi P, et al. Effects of Plant Biostimulation Time Span and Soil Electrical Conductivity on Greenhouse Tomato ‘Miniplum’ Yield and Quality in Diverse Crop Seasons. Plants. 2023; 12(7):1423. https://doi.org/10.3390/plants12071423
Chicago/Turabian StyleTallarita, Alessio V., Lorenzo Vecchietti, Nadezhda A. Golubkina, Agnieszka Sekara, Eugenio Cozzolino, Massimo Mirabella, Antonio Cuciniello, Roberto Maiello, Vincenzo Cenvinzo, Pasquale Lombardi, and et al. 2023. "Effects of Plant Biostimulation Time Span and Soil Electrical Conductivity on Greenhouse Tomato ‘Miniplum’ Yield and Quality in Diverse Crop Seasons" Plants 12, no. 7: 1423. https://doi.org/10.3390/plants12071423
APA StyleTallarita, A. V., Vecchietti, L., Golubkina, N. A., Sekara, A., Cozzolino, E., Mirabella, M., Cuciniello, A., Maiello, R., Cenvinzo, V., Lombardi, P., & Caruso, G. (2023). Effects of Plant Biostimulation Time Span and Soil Electrical Conductivity on Greenhouse Tomato ‘Miniplum’ Yield and Quality in Diverse Crop Seasons. Plants, 12(7), 1423. https://doi.org/10.3390/plants12071423