
Water and Nutrient Recovery for Cucumber Hydroponic Cultivation in Simultaneous Biological Treatment of Urine and Grey Water

 , ,
, ,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Cucumber Response to NUG Solution
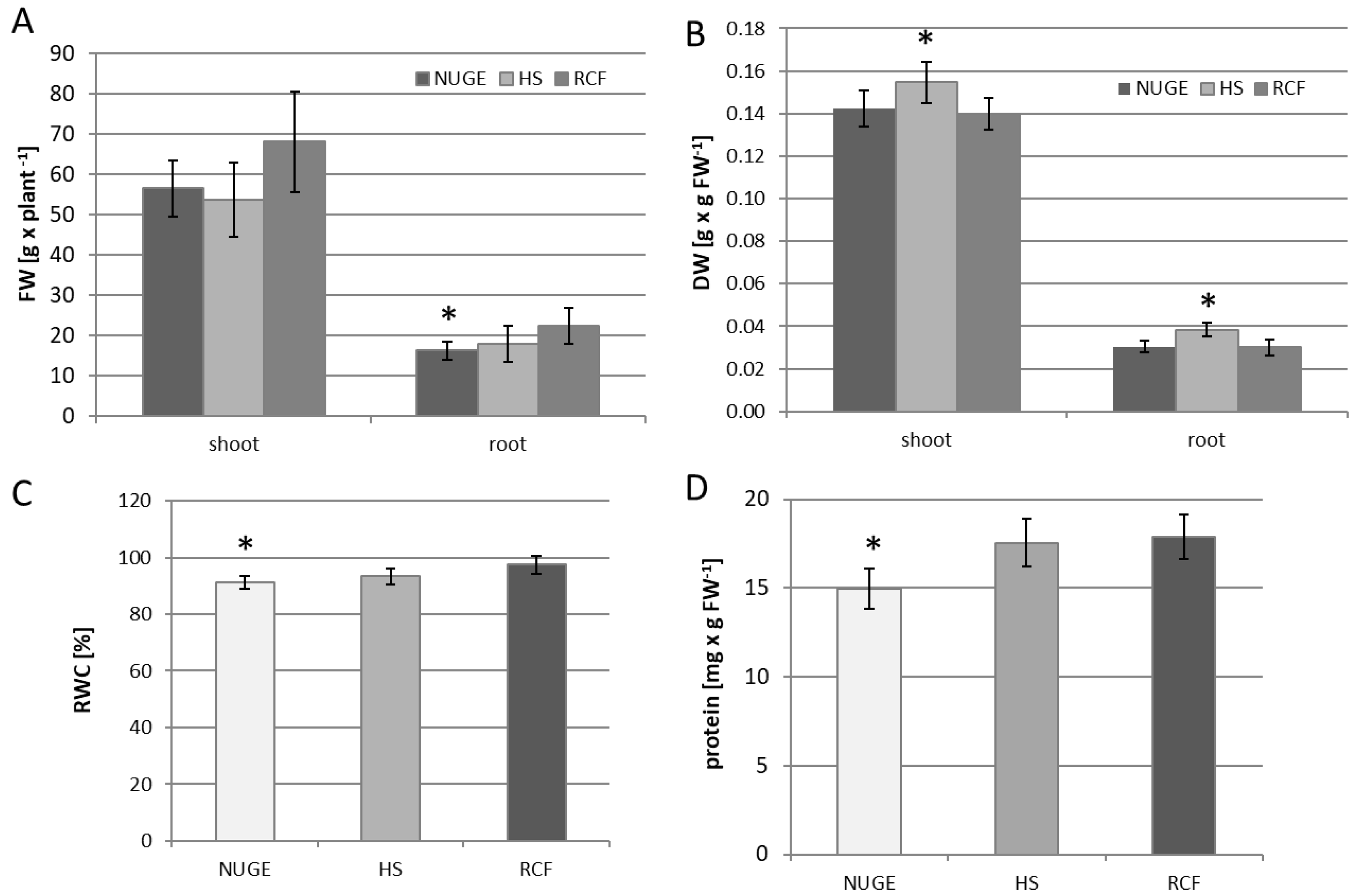
2.2. Yield, Water Status, and Protein Content of Cucumber Plants
2.3. Mineral Composition of Cucumber
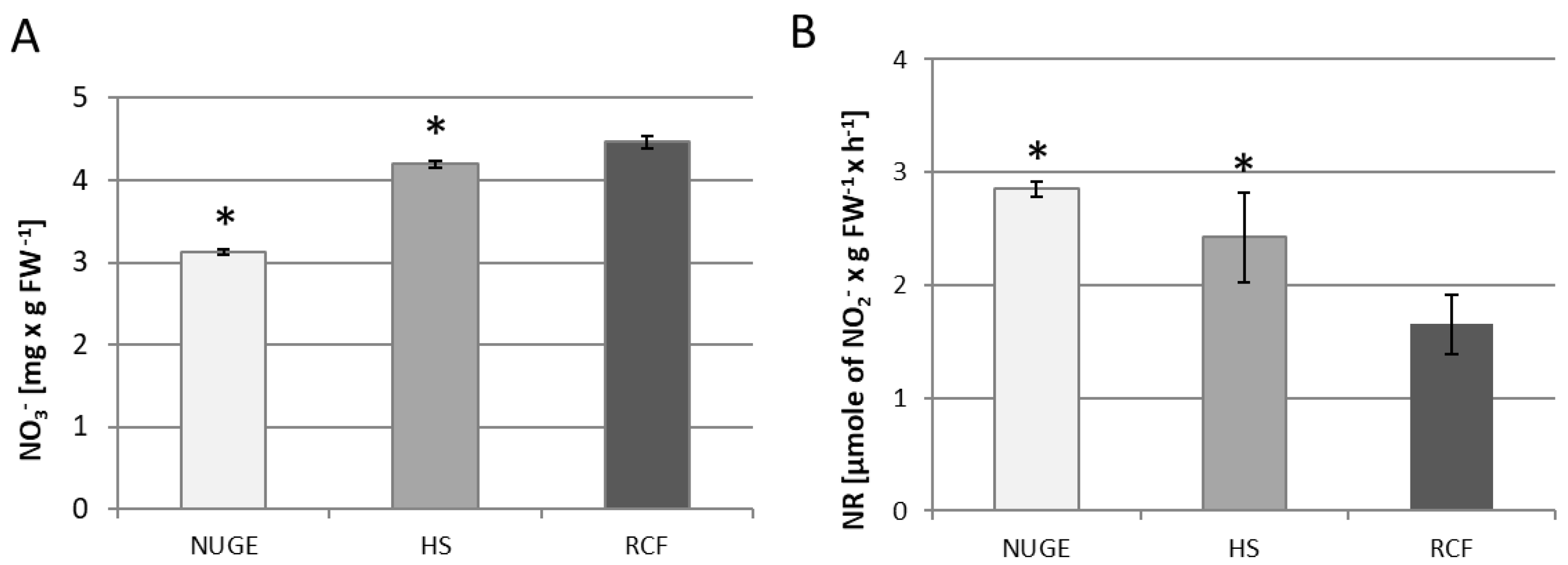
2.4. NR Activity and NO3− Content
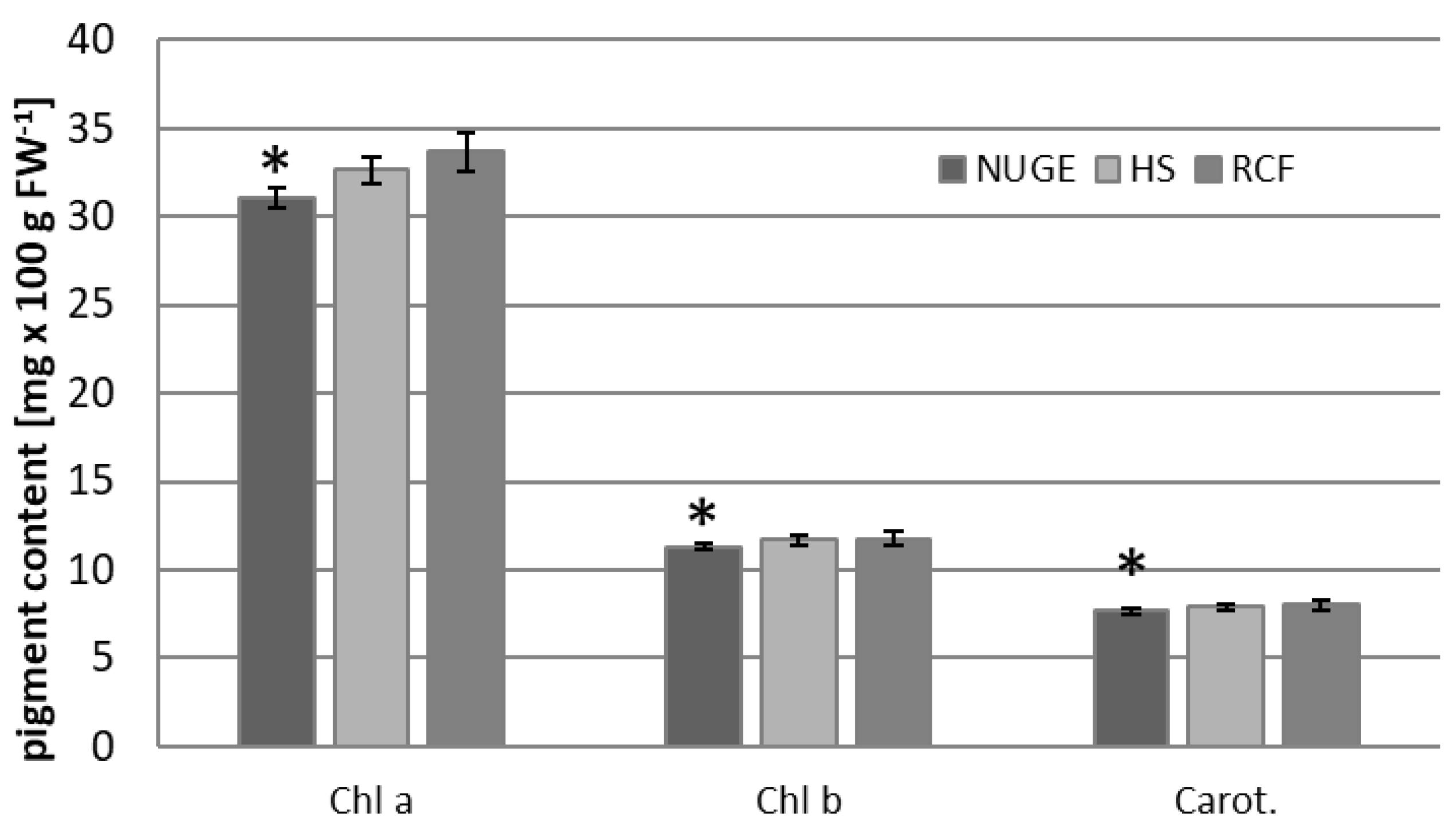
2.5. Photosynthetic Pigments and Photosynthetic Activity
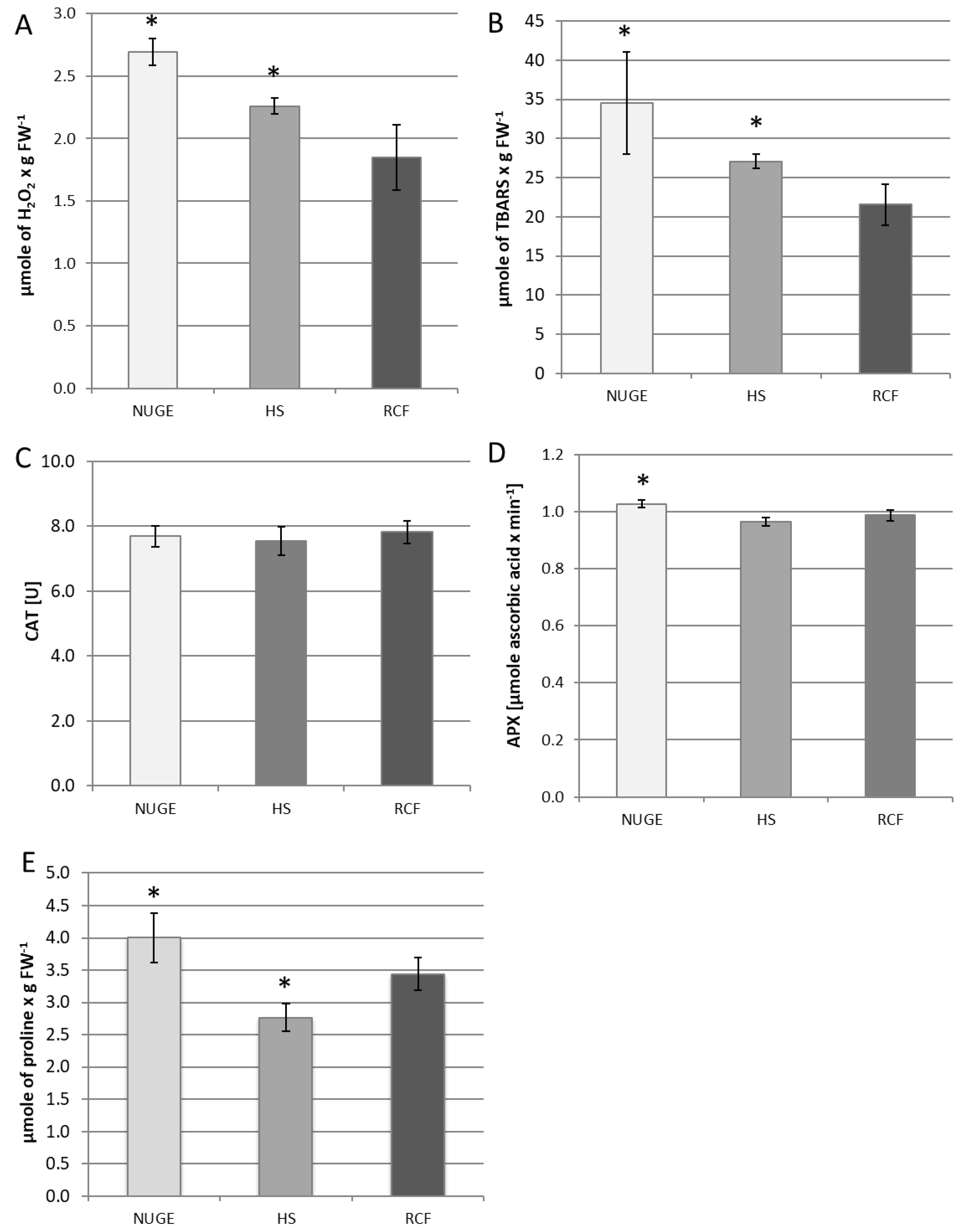
2.6. Analysis of Oxidative Stress Parameters
3. Discussion
4. Materials and Methods
4.1. Cultivation of Plants
4.2. Analytical Methods
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jaramillo, M.F.; Restrepo, I. Wastewater Reuse in Agriculture: A Review about Its Limitations and Benefits. Sustainability 2017, 9, 1734. [Google Scholar] [CrossRef]
- Mainardis, M.; Cecconet, D.; Moretti, A.; Callegari, A.; Goi, D.; Freguia, S.; Capodaglio, A.G. Wastewater Fertigation in Agriculture: Issues and Opportunities for Improved Water Management and Circular Economy. Environ. Pollut. 2022, 296, 118755. [Google Scholar] [CrossRef]
- Jurga, A.; Janiak, K.; Wizimirska, A.; Chochura, P.; Miodoński, S.; Muszyński-Huhajło, M.; Ratkiewicz, K.; Zięba, B.; Czaplicka-Pędzich, M.; Pilawka, T.; et al. Resource Recovery from Synthetic Nitrified Urine in the Hydroponic Cultivation of Lettuce (Lactuca Sativa Var. Capitata L.). Agronomy 2021, 11, 2242. [Google Scholar] [CrossRef]
- Karak, T.; Bhattacharyya, P. Human Urine as a Source of Alternative Natural Fertilizer in Agriculture: A Flight of Fancy or an Achievable Reality. Resour. Conserv. Recycl. 2011, 55, 400–408. [Google Scholar] [CrossRef]
- Martin, T.M.P.; Esculier, F.; Levavasseur, F.; Houot, S. Human Urine-Based Fertilizers: A Review. Crit. Rev. Env. Sci. Technol. 2020, 52, 1–47. [Google Scholar] [CrossRef]
- Shingiro, C.; Nyagatare, G.; Hirwa, H.; Solange, U. Effect of Human Urine and Compost Tea on Soil Physical and Chemical Properties Case Study Rubirizi Marshland. J. Agric. Sci. Eng. 2019, 5, 61–66. [Google Scholar]
- Jurga, A.; Ratkiewicz, K.; Wdowikowska, A.; Reda, M.; Janicka, M.; Chohura, P.; Janiak, K. Urine and Grey Water Based Liquid Fertilizer—Production and the Response of Plants. J. Env. Manag. 2023, 331, 117248. [Google Scholar] [CrossRef]
- Shafran, A.W.; Gross, A.; Ronen, Z.; Weisbrod, N.; Adar, E. Effects of Surfactants Originating from Reuse of Greywater on Capillary Rise in the Soil. Water Sci. Technol. 2005, 52, 157–166. [Google Scholar] [CrossRef]
- Janiak, K.; Jurga, A.; Kuźma, J.; Breś, W.; Muszyński-Huhajło, M. Surfactants Effect on Aeroponics and Important Mass Balances of Regenerative Life Support System—Lettuce Case Study. Sci. Total Environ. 2020, 718, 137324. [Google Scholar] [CrossRef]
- Sawadogo, B.; Sou, M.; Hijikata, N.; Sangare, D.; Hama MAIGA, A.; Funamizu, N. Effect of Detergents from Greywater on Irrigated Plants: Case of Okra (Abelmoschus Esculentus) and Lettuce (Lactuca Sativa). J. Arid Land Stud. 2014, 24, 117–120. [Google Scholar]
- Chai, L.; Yang, L.; Zhang, Y.; Zhou, Y.; Wang, F.; Wu, Z. Antagonism or Synergism? Responses of Hydrocharis Dubia (Bl.) Backer to Linear Alkylbenzene Sulfonate, Naphthalene and Their Joint Exposure. Ecotoxicol. Env. Saf. 2020, 200, 110747. [Google Scholar] [CrossRef]
- Fussy, A.; Papenbrock, J. An Overview of Soil and Soilless Cultivation Techniques—Chances, Challenges and the Neglected Question of Sustainability. Plants 2022, 11, 1153. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Acharya, S.; Kumar, K.; Singh, N.; Chaurasia, O.P. Hydroponics as an Advanced Technique for Vegetable Production: An Overview. J. Soil Water Conserv. 2018, 17, 364. [Google Scholar] [CrossRef]
- Mohsin, M.; Salam, M.M.A.; Nawrot, N.; Kaipiainen, E.; Lane, D.J.; Wojciechowska, E.; Kinnunen, N.; Heimonen, M.; Tervahauta, A.; Peräniemi, S.; et al. Phytoextraction and Recovery of Rare Earth Elements Using Willow (Salix Spp.). Sci. Total Environ. 2022, 809, 152209. [Google Scholar] [CrossRef]
- Mohsin, M.; Nawrot, N.; Wojciechowska, E.; Kuittinen, S.; Szczepańska, K.; Dembska, G.; Pappinen, A. Cadmium Accumulation by Phragmites Australis and Iris Pseudacorus from Stormwater in Floating Treatment Wetlands Microcosms: Insights into Plant Tolerance and Utility for Phytoremediation. J. Env. Manag. 2023, 331, 117339. [Google Scholar] [CrossRef] [PubMed]
- El-Nakhel, C.; Geelen, D.; de Paepe, J.; Clauwaert, P.; de Pascale, S.; Rouphael, Y. An Appraisal of Urine Derivatives Integrated in the Nitrogen and Phosphorus Inputs of a Lettuce Soilless Cultivation System. Sustainability 2021, 13, 4218. [Google Scholar] [CrossRef]
- Sibanda, M.; Mutanga, O.; Magwaza, L.S.; Dube, T.; Magwaza, S.T.; Odindo, A.O.; Mditshwa, A.; Mafongoya, P.L. Discrimination of Tomato Plants (Solanum Lycopersicum) Grown under Anaerobic Baffled Reactor Effluent, Nitrified Urine Concentrates and Commercial Hydroponic Fertilizer Regimes Using Simulated Sensor Spectral Settings. Agronomy 2019, 9, 373. [Google Scholar] [CrossRef]
- Zabel, P.; Bornemann, G.; Tajmar, M.; Schubert, D. Yield of Dwarf Tomatoes Grown with a Nutrient Solution Based on Recycled Synthetic Urine. Life Sci. Space Res. 2019, 20, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Adejumo, M.; Igwe, C.; Sridhar, M.K.C. Utilization of Urine for Cultivation of Selected Vegetable Crops in Backyard “Peeponics”. J. Env. Prot. 2019, 10, 22–38. [Google Scholar] [CrossRef]
- Heinonen-Tanski, H.; Sjöblom, A.; Fabritius, H.; Karinen, P. Pure Human Urine Is a Good Fertiliser for Cucumbers. Bioresour. Technol. 2007, 98, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Lin, L.; Luo, H.; Zhou, S.; Zhu, Y.; Wang, X.; Miao, L.; Wang, H.; Zhang, P. Recent Progress in the Regeneration and Genetic Transformation System of Cucumber. Appl. Sci. 2022, 12, 7180. [Google Scholar] [CrossRef]
- WANG, S.; Ku, S.S.; YE, X.; HE, C.; Kwon, S.Y.; Choi, P.S. Current Status of Genetic Transformation Technology Developed in Cucumber (Cucumis Sativus L.). J. Integr. Agric. 2015, 14, 469–482. [Google Scholar] [CrossRef]
- Jakubowska, D.; Janicka-Russak, M.; Kabała, K.; Migocka, M.; Reda, M. Modification of Plasma Membrane NADPH Oxidase Activity in Cucumber Seedling Roots in Response to Cadmium Stress. Plant Sci. 2015, 234, 50–59. [Google Scholar] [CrossRef]
- Janicka, M.; Reda, M.; Czyżewska, K.; Kabała, K. Involvement of Signalling Molecules NO, H2O2 and H2S in Modification of Plasma Membrane Proton Pump in Cucumber Roots Subjected to Salt or Low Temperature Stress. Funct. Plant Biol. 2018, 45, 428. [Google Scholar] [CrossRef] [PubMed]
- Janicka, M.; Reda, M.; Napieraj, N.; Michalak, A.; Jakubowska, D.; Kabała, K. Involvement of Diamine Oxidase in Modification of Plasma Membrane Proton Pump Activity in Cucumis Sativus L. Seedlings under Cadmium Stress. Int. J. Mol. Sci. 2023, 24, 262. [Google Scholar] [CrossRef] [PubMed]
- Janicka-Russak, M.; Kabała, K.; Wdowikowska, A.; Kłobus, G. Modification of Plasma Membrane Proton Pumps in Cucumber Roots as an Adaptation Mechanism to Salt Stress. J. Plant Physiol. 2013, 170, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Janicka-Russak, M.; KabaŁa, K.; MŁodzińska, E.; KŁobus, G. The Role of Polyamines in the Regulation of the Plasma Membrane and the Tonoplast Proton Pumps under Salt Stress. J. Plant Physiol. 2010, 167, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Janicka-Russak, M.; Kabała, K.; Wdowikowska, A.; Kłobus, G. Response of Plasma Membrane H+-ATPase to Low Temperature in Cucumber Roots. J. Plant Res. 2012, 125, 291–300. [Google Scholar] [CrossRef]
- Kabała, K.; Janicka-Russak, M.; Reda, M.; Migocka, M. Transcriptional Regulation of the V-ATPase Subunit c and V-PPase Isoforms in Cucumis Sativus under Heavy Metal Stress. Physiol. Plant. 2014, 150, 32–45. [Google Scholar] [CrossRef]
- Kabała, K.; Reda, M.; Wdowikowska, A.; Janicka, M. Role of Plasma Membrane NADPH Oxidase in Response to Salt Stress in Cucumber Seedlings. Antioxidants 2022, 11, 1534. [Google Scholar] [CrossRef]
- Kabała, K.; Zboińska, M.; Głowiak, D.; Reda, M.; Jakubowska, D.; Janicka, M. Interaction between the Signaling Molecules Hydrogen Sulfide and Hydrogen Peroxide and Their Role in Vacuolar H+-ATPase Regulation in Cadmium-Stressed Cucumber Roots. Physiol. Plant. 2019, 166, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Reda, M.; Golicka, A.; Kabała, K.; Janicka, M. Involvement of NR and PM-NR in NO Biosynthesis in Cucumber Plants Subjected to Salt Stress. Plant Sci. 2018, 267, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Wdowikowska, A.; Klobus, G. The Plasma Membrane Proton Pump Gene Family in Cucumber. Acta Physiol. Plant. 2016, 38, 1–14. [Google Scholar] [CrossRef]
- Kwon, T.; Kim, K.; Yoon, H.-J.; Lee, S.; Kim, B.; Siddiqui, Z.S. Phenotyping of Plants for Drought and Salt Tolerance Using Infra-Red Thermography. Plant Breed. Biotechnol. 2015, 3, 299–307. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Larré, C.F.; Fernando, J.A.; Marini, P.; Bacarin, M.A.; Peters, J.A. Growth and Chlorophyll a Fluorescence in Erythrina Crista-Galli L. Plants under Flooding Conditions. Acta Physiol. Plant. 2013, 35, 1463–1471. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of Nutrient Deficiency in Maize and Tomato Plants by in Vivo Chlorophyll a Fluorescence Measurements. Plant Physiol. Biochem. 2014, 81, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Bąba, W.; Gediga, K.; Goltsev, V.; Samborska, I.A.; Cetner, M.D.; Dimitrova, S.; Piszcz, U.; Bielecki, K.; Karmowska, K.; et al. Chlorophyll Fluorescence as a Tool for Nutrient Status Identification in Rapeseed Plants. Photosynth. Res. 2018, 136, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Janiak, K.; Jurga, A.; Wizimirska, A.; Miodoński, S.; Muszyński-Huhajło, M.; Ratkiewicz, K.; Zięba, B. Urine Nitrification Robustness for Application in Space: Effect of High Salinity and the Response to Extreme Free Ammonia Concentrations. J. Env. Manag. 2021, 279, 111610. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus Uptake by Plants: From Soil to Cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef]
- Wu, H. Plant Salt Tolerance and Na+ Sensing and Transport. Crop J. 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Yao, X.; Horie, T.; Xue, S.; Leung, H.-Y.; Katsuhara, M.; Brodsky, D.E.; Wu, Y.; Schroeder, J.I. Differential Sodium and Potassium Transport Selectivities of the Rice OsHKT2;1 and OsHKT2;2 Transporters in Plant Cells. Plant Physiol. 2009, 152, 341–355. [Google Scholar] [CrossRef]
- Kabała, K.; Janicka-Russak, M. Antiport Activity in Plasma Membrane and Tonoplast Vesicles Isolated from NaCl-Treated Cucumber Roots. Biol. Plant. 2012, 56, 377–382. [Google Scholar] [CrossRef]
- Pardo, J.M.; Rubio, F. Na+ and K+ Transporters in Plant Signaling. Transp. Pumps Plant Signal. 2011, 7, 65–98. [Google Scholar]
- Olías, R.; Eljakaoui, Z.; Li, J.; de Morales, P.A.; Marín-Manzano, M.C.; Pardo, J.M.; Belver, A. The Plasma Membrane Na+/H+ Antiporter SOS1 Is Essential for Salt Tolerance in Tomato and Affects the Partitioning of Na+/between Plant Organs. Plant Cell Environ. 2009, 32, 904–916. [Google Scholar] [CrossRef] [PubMed]
- Sudhir, P.; Murthy, S.D.S. Effects of Salt Stress on Basic Processes of Photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.-L.; Liu, L.-N.; Xie, Q.; Sui, N. Photosynthetic Regulation Under Salt Stress and Salt-Tolerance Mechanism of Sweet Sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef] [PubMed]
- Hannachi, S.; Steppe, K.; Eloudi, M.; Mechi, L.; Bahrini, I.; van Labeke, M.-C. Salt Stress Induced Changes in Photosynthesis and Metabolic Profiles of One Tolerant (‘Bonica’) and One Sensitive (‘Black Beauty’) Eggplant Cultivars (Solanum Melongena L.). Plants 2022, 11, 590. [Google Scholar] [CrossRef]
- Zhao, H.; Liang, H.; Chu, Y.; Sun, C.; Wei, N.; Yang, M.; Zheng, C. Effects of Salt Stress on Chlorophyll Fluorescence and the Antioxidant System in Ginkgo Biloba L. Seedlings. HortScience 2019, 54, 2125–2133. [Google Scholar] [CrossRef]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and Salinity. Crop Sci. 2004, 44, 806–811. [Google Scholar] [CrossRef]
- Wang, R.; Chen, S.; Deng, L.; Fritz, E.; Hüttermann, A.; Polle, A. Leaf Photosynthesis, Fluorescence Response to Salinity and the Relevance to Chloroplast Salt Compartmentation and Anti-Oxidative Stress in Two Poplars. Trees 2007, 21, 581–591. [Google Scholar] [CrossRef]
- Athar, H.-R.; Zulfiqar, F.; Moosa, A.; Ashraf, M.; Zafar, Z.U.; Zhang, L.; Ahmed, N.; Kalaji, H.M.; Nafees, M.; Hossain, M.A.; et al. Salt Stress Proteins in Plants: An Overview. Front. Plant Sci. 2022, 13, 999058. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Astorga, G.I.; Alcaraz-Meléndez, L. Salinity Effects on Protein Content, Lipid Peroxidation, Pigments, and Proline in Paulownia Imperialis (Siebold & Zuccarini) and Paulownia Fortunei (Seemann & Hemsley) Grown in Vitro. Electron. J. Biotechnol. 2010, 13, 1–15. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological Functions of Mineral Macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Pettigrew, W.T. Potassium Influences on Yield and Quality Production for Maize, Wheat, Soybean and Cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in Plant Morphology and Dry Matter Partitioning Caused by Potassium Deficiency in Gossypium Hirsutum (L.). Env. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Benlloch-González, M.; Arquero, O.; Fournier, J.M.; Barranco, D.; Benlloch, M. K+ Starvation Inhibits Water-Stress-Induced Stomatal Closure. J. Plant Physiol. 2008, 165, 623–630. [Google Scholar] [CrossRef]
- Benlloch-Gonzalez, M.; Romera, J.; Cristescu, S.; Harren, F.; Fournier, J.M.; Benlloch, M. K+ Starvation Inhibits Water-Stress-Induced Stomatal Closure via Ethylene Synthesis in Sunflower Plants. J. Exp. Bot. 2010, 61, 1139–1145. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sano, T.; Tamaoki, M.; Nakajima, N.; Kondo, N.; Hasezawa, S. Ethylene Inhibits Abscisic Acid-Induced Stomatal Closure in Arabidopsis. Plant Physiol. 2005, 138, 2337–2343. [Google Scholar] [CrossRef]
- Xu, X.; Du, X.; Wang, F.; Sha, J.; Chen, Q.; Tian, G.; Zhu, Z.; Ge, S.; Jiang, Y. Effects of Potassium Levels on Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock Seedlings. Front. Plant Sci. 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.M.; Romero, L. Relationship between Potassium Fertilisation and Nitrate Assimilation in Leaves and Fruits of Cucumber (Cucumis Sativus) Plants. Ann. Appl. Biol. 2002, 140, 241–245. [Google Scholar] [CrossRef]
- Thornburg, T.E.; Liu, J.; Li, Q.; Xue, H.; Wang, G.; Li, L.; Fontana, J.E.; Davis, K.E.; Liu, W.; Zhang, B.; et al. Potassium Deficiency Significantly Affected Plant Growth and Development as Well as MicroRNA-Mediated Mechanism in Wheat (Triticum Aestivum L.). Front. Plant Sci. 2020, 11, 1219. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, W.; Yang, J.; Oosterhuis, D.M.; Loka, D.A.; Zhou, Z. Relationship between Potassium Fertilization and Nitrogen Metabolism in the Leaf Subtending the Cotton ( Gossypium Hirsutum L.) Boll during the Boll Development Stage. Plant Physiol. Biochem. 2016, 101, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Reda, M.; Migocka, M.; Kłobus, G. Effect of Short-Term Salinity on the Nitrate Reductase Activity in Cucumber Roots. Plant Sci. 2011, 180, 783–788. [Google Scholar] [CrossRef]
- Rohilla, P.; Yadav, J.P. Acute Salt Stress Differentially Modulates Nitrate Reductase Expression in Contrasting Salt Responsive Rice Cultivars. Protoplasma 2019, 256, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Peng, Y.; Li, Z.; Guo, H.; Xia, X.; Li, B.; Yin, W. The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1202. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Mittler, R. Reactive Oxygen Species and Temperature Stresses: A Delicate Balance between Signaling and Destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Labudda, M. Lipid Peroxidation As A Biochemical Marker For Oxidative Stress During Drought. In An Effective Tool For Plant Breeding; E-wydawnictwo: Poland, 2013; Available online: http://wwww.e-wydawnictwo.eu/Document/DocumentPreview/3342 (accessed on 1 February 2023).
- Kaur, N.; Dhawan, M.; Sharma, I.; Pati, P.K. Interdependency of Reactive Oxygen Species Generating and Scavenging System in Salt Sensitive and Salt Tolerant Cultivars of Rice. BMC Plant Biol. 2016, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Dans, P.D.; Coitiño, E.L.; Borsani, O.; Monza, J. Connecting Proline and γ-Aminobutyric Acid in Stressed Plants through Non-Enzymatic Reactions. PLoS ONE 2015, 10, e0115349. [Google Scholar] [CrossRef] [PubMed]
- el Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How Does Proline Treatment Promote Salt Stress Tolerance During Crop Plant Development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Kim, G.-B.; Hyun, S.-H.; Lee, S.-Y.; Lee, C.-Y.; Choi, H.-K.; Choi, H.-K.; Nam, Y.-W. Physiological and Metabolomic Analysis of a Knockout Mutant Suggests a Critical Role of MtP5CS3 Gene in Osmotic Stress Tolerance of Medicago Truncatula. Euphytica 2013, 193, 101–120. [Google Scholar] [CrossRef]
- Quan, L.-J.; Zhang, B.; Shi, W.-W.; Li, H.-Y. Hydrogen Peroxide in Plants: A Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- Jabeen, N.; Ahmad, R. The Activity of Antioxidant Enzymes in Response to Salt Stress in Safflower (Carthamus Tinctorius L.) and Sunflower (Helianthus Annuus L.) Seedlings Raised from Seed Treated with Chitosan. J. Sci. Food Agric. 2013, 93, 1699–1705. [Google Scholar] [CrossRef]
- Kibria, M.G.; Hossain, M.; Murata, Y.; Hoque, M.A. Antioxidant Defense Mechanisms of Salinity Tolerance in Rice Genotypes. Rice Sci. 2017, 24, 155–162. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- González, L.; González-Vilar, M. Determination of Relative Water Content. In Handbook of Plant Ecophysiology Techniques; Reigosa Roger, M.J., Ed.; Springer: Dordrecht, The Netherland, 2001; pp. 207–212. ISBN 978-0-306-48057-7. [Google Scholar]
- Sáez-Plaza, P.; Michałowski, T.; Navas, M.J.; Asuero, A.G.; Wybraniec, S. An Overview of the Kjeldahl Method of Nitrogen Determination. Part I. Early History, Chemistry of the Procedure, and Titrimetric Finish. Crit. Rev. Anal. Chem. 2013, 43, 178–223. [Google Scholar] [CrossRef]
- Reda, M. Response of Nitrate Reductase Activity and NIA Genes Expression in Roots of Arabidopsis Hxk1 Mutant Treated with Selected Carbon and Nitrogen Metabolites. Plant Sci. 2015, 230, 51–58. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- LICHTENTHALER, H.K.; WELLBURN, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient As a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosyntesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty , P., Eds.; CRC Press: Boca Raton, FL, USA, 2000; Chapter 25; pp. 445–483. [Google Scholar]
- Kabała, K.; Janicka-Russak, M.; Kłobus, G. Different Responses of Tonoplast Proton Pumps in Cucumber Roots to Cadmium and Copper. J. Plant Physiol. 2010, 167, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Gong-Xiang Chen; Kozi Asada Ascorbate Peroxidase in Tea Leaves: Occurrence of Two Isozymes and the Differences in Their Enzymatic and Molecular Properties. Plant Cell Physiol. 1989, 30, 987–998. [CrossRef]
- Hugo Aebi Catalase in Vitro. Methods Enzym. 1984, 105, 121–126.
- Kaiser, W.M.; Huber, S.C. Correlation between Apparent Activation State of Nitrate Reductase (NR), NR Hysteresis and Degradation of NR Protein. J. Exp. Bot. 1997, 48, 1367–1374. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Unit | NUG | NUGE | HS | RCF |
|---|---|---|---|---|---|
| pH | - | 7.21 | 6.5 | 6.5 | 6.5 |
| N-NH4+ | mg·L−1 | trace | trace | trace | trace |
| N-NO3− | mg·L−1 | 189 | 192 | 224 | 180 |
| P-PO4 | mg·L−1 | 20 | 41 | 62 | 60 |
| K | mg·L−1 | 73 | 265 | 235 | 250 |
| Ca | mg·L−1 | 31 | 150 | 160 | 150 |
| Mg | mg·L−1 | 7 | 25 | 24 | 50 |
| Na | mg·L−1 | 497 | 485 | 12.5 | 10.6 |
| Cl | mg·L−1 | 205 | 198 | 1.77 | 3.7 |
| S-SO42− | mg·L−1 | 166 | 208 | 32 | 30 |
| Fe | mg·L−1 | trace | 1.5 | 2.00 | 2.00 |
| Mn | mg·L−1 | trace | 0.75 | 0.80 | 0.75 |
| Cu | mg·L−1 | trace | 0.05 | 0.03 | 0.10 |
| Zn | mg·L−1 | trace | 0.35 | 0.13 | 0.50 |
| B | mg·L−1 | 0.07 | 0.35 | 0.27 | 0.40 |
| Mo | mg·L−1 | trace | 0.05 | 0.05 | 0.05 |
| Anionic surfactant SMCT | mg·L−1 | 31 | 26 | 0 | 0 |
| Nutrient solution | Cucumber organ | Macroelements in g·kg DM−1 | |||||
| N | P | K | Ca | Mg | S | ||
| NUGE | leaf | 30.5 ± 0.24 | 5.17 ± 0.12 | 26.5 ± 0.3 | 19.6 ± 0.36 | 3.2 ± 0.08 | 4.47 ± 0.25 |
| HS | 47.2 ± 0.25 | 7.83 ± 0.12 | 44.5 ± 0.2 | 30.2 ± 0.2 | 4.8 ± 0.1 | 6.2 ± 0.1 | |
| RCF | 43.1 ± 0.21 | 6.17 ± 0.26 | 41.2 ± 0.3 | 26.4 ± 0.2 | 4.4 ± 0.1 | 5.5 ± 0.08 | |
| NUGE | root | 14.5 ± 0.12 | 2.47 ± 0.05 | 15.5 ± 0.02 | 3.2 ± 0.05 | 3.4 ± 0.05 | 1.2 ± 0.1 |
| HS | 16.5 ± 0.12 | 4.23 ± 0.05 | 15.2 ± 0.2 | 4.1 ± 0.1 | 3.8 ± 0.08 | 1.4 ± 0.05 | |
| RCF | 15.2 ± 0.22 | 3.8 ± 0.08 | 16.5 ± 0.3 | 3.6 ± 0.1 | 4.1 ± 0.08 | 1.6 ± 0.09 | |
| NUGE | stem | 18.9 ± 0.12 | 4.53 ± 0.12 | 10.5 ± 0.2 | 9.2 ± 0.1 | 2.8 ± 0.1 | 4.8 ± 0.1 |
| HS | 21.2 ± 0.24 | 5.6 ± 0.16 | 11.2 ± 0.2 | 12.1 ± 0.2 | 4.23 ± 0.1 | 5.2 ± 0.1 | |
| RCF | 22.4 ± 0.17 | 6.17 ± 0.2 | 11.6 ± 0.12 | 11.2 ± 0.2 | 3.9 ± 0.1 | 4.3 ± 0.1 | |
| Nutrient solution | Cucumber organ | Microelements and sodium in mg·kg DM−1 | |||||
| Fe | Mn | Cu | Zn | B | Na | ||
| NUGE | leaf | 74.47 ± 0.74 | 75.60 ± 0.41 | 6.25 ± 0.05 | 48.57 ± 0.39 | 46.23 ± 0.45 | 845.3 ± 9.3 |
| HS | 121.47 ± 0.4 | 102.27 ± 0.3 | 9.54 ± 0.04 | 56.53 ± 0.4 | 72.13 ± 0.31 | 631 ± 7.1 | |
| RCF | 89.63 ± 0.34 | 96.5 ± 0.29 | 8.14 ± 0.01 | 64.23 ± 0.34 | 62.13 ± 0.25 | 598.3 ± 2.1 | |
| NUGE | root | 86.47 ± 0.26 | 55.57 ± 0.12 | 32.13 ± 0.21 | 86.63 ± 0.26 | 82.33 ± 0.17 | 1123.3 ± 4.5 |
| HS | 106.3 ± 0.36 | 98.63 ± 0.21 | 45.63 ± 0.37 | 102.33 ± 0.4 | 72.43 ± 0.12 | 367.7 ± 1.2 | |
| RCF | 98.63 ± 0.09 | 85.23 ± 0.12 | 49.80 ± 0.16 | 92.13 ± 0.12 | 80.23 ± 0.31 | 322 ± 2.4 | |
| NUGE | stem | 86.23 ± 0.66 | 62.5 ± 0.22 | 17.54 ± 0.12 | 72.40 ± 0.29 | 56.77 ± 0.37 | 1322 ± 14.5 |
| HS | 98.63 ± 0.42 | 85.57 ± 0.26 | 28.6 ± 0.37 | 87.46 ± 0.73 | 76.33 ± 0.56 | 421.3 ± 6.9 | |
| RCF | 92.3 ± 0.36 | 96.3 ± 0.41 | 25.57 ± 0.5 | 92.27 ± 0.48 | 71.17 ± 0.37 | 452.3 ± 3.3 | |
| NUGE ± SD | HS ± SD | RCF ± SD | |
|---|---|---|---|
| T for Fm | 343 ± 89 | 312 ± 44 | 314 ± 48 |
| Area | 55,364 ± 5741 | 61,749 ± 4346 | 58,984 ± 4536 |
| Fo | 425 ± 12 | 448 ±20 | 457 ± 21 |
| Fm | 2595 ± 131 * | 2679 ± 56 | 2676 ± 42 |
| Fv | 2169 ± 125 | 2238 ± 77 | 2229 ± 29 |
| Fo/Fm | 0.164 ± 0.007 | 0.165 ± 0.01 | 0.167 ± 0.009 |
| Fv/Fm | 0.836 ± 0.007 | 0.839 ± 0.009 | 0.833 ± 0.009 |
| ABS/RC | 2.064 ± 0.053 | 1.998 ± 0.089 | 2.065 ± 0.078 |
| Tro/RC | 1.725 ± 0.053 | 1.668 ± 0.058 | 1.720 ± 0.065 |
| Dio/CSo | 69.780 ± 3.870 | 75.377 ± 7.896 | 77.787 ± 6.59 |
| Eto/RC | 0.964 ± 0.066 | 0.982 ± 0.043 | 0.983 ± 0.03 |
| PIABS | 3.186 ± 0.598 | 3.772 ± 0.888 | 3.237 ± 0.264 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wdowikowska, A.; Reda, M.; Kabała, K.; Chohura, P.; Jurga, A.; Janiak, K.; Janicka, M. Water and Nutrient Recovery for Cucumber Hydroponic Cultivation in Simultaneous Biological Treatment of Urine and Grey Water. Plants 2023, 12, 1286. https://doi.org/10.3390/plants12061286
Wdowikowska A, Reda M, Kabała K, Chohura P, Jurga A, Janiak K, Janicka M. Water and Nutrient Recovery for Cucumber Hydroponic Cultivation in Simultaneous Biological Treatment of Urine and Grey Water. Plants. 2023; 12(6):1286. https://doi.org/10.3390/plants12061286
Chicago/Turabian StyleWdowikowska, Anna, Małgorzata Reda, Katarzyna Kabała, Piotr Chohura, Anna Jurga, Kamil Janiak, and Małgorzata Janicka. 2023. "Water and Nutrient Recovery for Cucumber Hydroponic Cultivation in Simultaneous Biological Treatment of Urine and Grey Water" Plants 12, no. 6: 1286. https://doi.org/10.3390/plants12061286
APA StyleWdowikowska, A., Reda, M., Kabała, K., Chohura, P., Jurga, A., Janiak, K., & Janicka, M. (2023). Water and Nutrient Recovery for Cucumber Hydroponic Cultivation in Simultaneous Biological Treatment of Urine and Grey Water. Plants, 12(6), 1286. https://doi.org/10.3390/plants12061286