Abstract
The worldwide production of eggplant is estimated at about 58 Mt, with China, India and Egypt being the major producing countries. Breeding efforts in the species have mainly focused on increasing productivity, abiotic and biotic tolerance/resistance, shelf-life, the content of health-promoting metabolites in the fruit rather than decreasing the content of anti-nutritional compounds in the fruit. From the literature, we collected information on mapping quantitative trait loci (QTLs) affecting eggplant’s traits following a biparental or multi-parent approach as well as genome-wide association (GWA) studies. The positions of QTLs were lifted according to the eggplant reference line (v4.1) and more than 700 QTLs were identified, here organized into 180 quantitative genomic regions (QGRs). Our findings thus provide a tool to: (i) determine the best donor genotypes for specific traits; (ii) narrow down QTL regions affecting a trait by combining information from different populations; (iii) pinpoint potential candidate genes.
1. Introduction
Eggplant (Solanum melongena L., 2n = 2x = 24) is the fourth most important crop economically and nutritionally, belonging to Solanaceae, a large plant family including important crops such as tomato, potato, pepper, and tobacco. According to the latest FAOSTAT report [1], eggplant is cultivated worldwide, with a global production of 58 Mt in 2021. China and India are the main producing countries, accounting for about 86% of total production, while Egypt, Turkey, and Italy represent the main producers of the Mediterranean region. Contrary to most other solanaceous crops originating in the New World [2,3,4,5,6,7], eggplant has a phylogenetic uniqueness due to its exclusive Asian origin. The species has been reported to be the result of two or three independent domestication events, though recent studies have suggested a unique one [8,9]. Within the genus, eggplant and its relatives belong to the subgenus Leptostemonum, collectively known as the ‘spiny solanum’ group [10]. The most closely related species from the eggplant clade have been reported to be the direct wild ancestor S. insanum L. and the sister species S. incanum L. [11,12], while two other eggplant crops belonging to the Anguivi clade, the Ethiopian/scarlet eggplant (S. aethiopicum L.) and the African/Gboma eggplant (S. macrocarpon L.), have a locally important production, with the fruits and leaves of both species used for food and medicine [10,13,14]. Compared with cultivated eggplants, their wild relatives present a broader adaptation to the environment and climate, carry abundant genetic diversity, and have higher potential in crop improvement [15,16].
A necessary condition to exploit the introgression of traits of interest from crop wild relatives (CWRs) into cultivated plants is the knowledge of the associated or responsible genes/quantitative trait loci (QTLs) controlling the traits [17,18,19,20,21]. To dissect the genetic basis of complex traits, genomic studies using bi-parental QTL mapping (linkage mapping) and genome-wide association (GWA) mapping can be conducted, based on the significant association between markers and a phenotype of interest. Traditional biparental mapping approach is highly dependent on the genetic diversity of the two parental lines, and the effects of the detected QTLs vary depending on the chosen population [22,23]. Therefore, the number of genetic recombination events that occur during the construction of the mapping population affects both genetic mapping resolution and allele richness. The construction of a genetic linkage map requires a mapping population to analyze the recombination of specific molecular markers defining the position and relative genetic distance of the markers along the chromosomes. In the past few decades, several first-generation genetics’ maps (based on pre-NGS techniques) were developed from interspecific hybridizations between cultivated S. melongena and S. linnaeanum or S. incanum and applied for QTL analyses of domestication and morphological traits [24,25,26], as well as to locate genes involved in polyphenol biosynthesis [27]. Intra-specific maps were also constructed using both F2 and DH populations [28,29,30,31,32,33]. In parallel with the advances in the genetic linkage maps, the identification of QTL regions associated with agronomic traits has been considerably promoted in eggplant. The first NGS-based eggplant genetic map was developed on an intra-specific F2 population using RAD-tag derived markers [34] and genotyped via Illumina GoldenGate© assay [35,36]. Afterward, several genetic linkage maps were constructed for mapping disease resistance, parthenocarpy, and plant morphological-related traits [37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52]. Recently, a multiparent advanced generation intercross (MAGIC) population was developed by Mangino et al. [53], allowing the identification of putative regions and candidate genes for anthocyanin pigmentation.
In contrast, GWA studies are performed on a population of unrelated individuals in a heterogeneous collection, in which historical recombinations have accumulated over generations. As a result, the association mapping shows a higher map resolution and greater number of investigated alleles compared with the QTL mapping approach [54]. The detection power of the GWA approach can be affected by many factors including the population structure and dimension, allele frequency, as well as the phenotypic variation [55]. Furthermore, population structure (i.e., genetic relatedness between individuals in a population) may lead to false-positive associations between genotypes and the investigated traits if not taken into account [56,57,58]. For these reasons, an integrated approach may be crucial for the understanding of the architecture of complex quantitative traits. Nowadays, the availability of large germplasm collections, together with relatively low genotyping prices, provide robustness to GWA studies, making it possible to understand the architecture of complex traits [59]. However, only a few association mapping studies have been reported in eggplant. A first attempt was conducted by Ge et al. [60] to identify functional genes and QTLs related to fruit-related traits using a panel of 141 eggplant accessions. Afterwards, a larger panel (191 accessions) was employed to analyze the marker/trait associations for key breeding fruit and plant traits [61,62,63]. The described association mapping studies not only highlighted numerous previously identified genes/QTLs, but also allowed the discovery of novel loci and candidate genes, providing a valuable resource for the development of a marker-assisted selection breeding strategy.
A next step in the genomic era is provided by the concept of pangenome, the nonredundant set of genomic sequences within a species which include the core genes present in all individuals and dispensable genes only found in a subset [64]. Furthermore, pangenome approaches allow the identification of selective sweeps, presence/absence variations (PAVs), and structural variations linked to key agronomic traits. Compared to tomato and pepper, for which pangenome studies have been carried out using hundreds of accessions [65,66,67], the first eggplant pangenome has been recently established from 24 accessions of S. melongena, one accession of S. insanum, and one accession of S. incanum [68].
All the above-mentioned approaches have allowed the identification of a wide number of eggplant QTL regions for many agronomic and quality-related traits. To provide a comprehensive overview of the current genetic knowledge, 28 scientific papers and their supplemental data were collected here, integrated, and summarized. Combined information represents a valuable tool for marker-assisted selection breeding schemes, since it may be employed to find potential donors for a particular trait, to highlight key QTL regions as well as potential candidate genes for clarifying the genetic architecture of the eggplant agronomic traits.
2. Construction of a Unified Eggplant QTL Map and Identification of Candidate Genes
A total of 28 scientific articles reporting both QTL mapping and GWA studies were analyzed (Table S1). The studies were selected based on the feasibility of retrieving the markers’ position on the genome. This is an issue for the research carried out before a reference genome came out, where just the genetic linkage positions (cM) of markers were reported, or dominant markers as AFLP were used. To overcome this constraint, the physical position of the markers (when possible) was retrieved by aligning their sequences on the eggplant reference genome (line ‘67/3′-version 4.1) [68] using BLASTn [69]. If markers were instead mapped to another genome [14,38,70,71], minimap2 [72] was employed to determine their position on the eggplant reference genome v4.1. All the collected QTLs data were organized in a single database (Table S2) including information on QTL name, QTLs, related marker ID, chromosome position (in cM and Mb), significance of the associations with the examined trait (p-value; LOD score; percentage of variation explained by the QTL and effect), and the mapping population (type, cross, or association panel). Eggplant traits included in this review were classified into seven classes: (i) morphological traits—including plant (PL), leaf (LF), and flower (FL) traits; (ii) prickles (PK); (iii) parthenocarpy (PT) and male sterility (MS); (iv) fruit-related traits—including shape (SH), productivity (PR), quality (QL), and metabolites (MT); (v) anthocyanins (AN); (vi) biotic resistance (pathogens and pests’ resistance) (RS); (vii) abiotic resistance. Furthermore, to suggest genomic regions harboring potential candidate genes, quantitative trait regions (QGRs) were retrieved from overlapping QTLs for each eggplant chromosome, and associated candidate genes were identified in literature. To overcome the absence of a reported genetic confidence intervals or (average) linkage disequilibrium (LD) decay for the collected QTLs, potential positional errors were standardized by setting an empirical defined window of ±2.5 Mb around the QTLs, as suggested by Martina et al. [73]. QGRs were named based on the trait classes defined above. Because no QTLs were available for male sterility and pests/abiotic resistances, the literature was investigated to identify potential candidate genes (Table S3).
3. Morphological Traits
Eggplant is a bushy and vigorous plant, with large leaves and woody stems, and several morphological traits can be targeted during varietal development to obtain cultivars with higher agronomical characteristics. For instance, height impacts on plant habit and on the agronomic management of the crop, while leaf shapes, length, and width are crucial for photosynthetic efficiency. Flower shape, together with its sexual organs’ development, is directly linked to flower fertility, and thus to plant productivity. Several QTLs have been reported for the main morphological traits in eggplant. To ease the search for regions of interest, QTLs were split into three categories: (i) plant (PL); (ii) leaf (LF); and (iii) flower (FL). Data collection revealed a total of 84 morphology-related QTLs. By comparing these regions, 45 unique QGRs were defined (Table S2; Figure 1). On the whole, 38 QTLs were not included in any QGRs due to the lack of their genomic position.
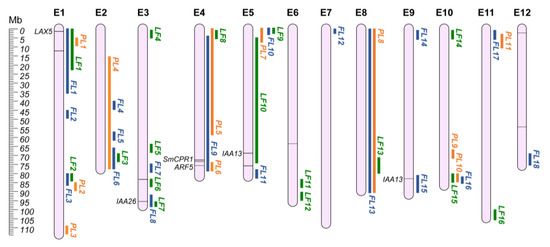
Figure 1.
Morphological QGRs chromosome map (E1–E12). Orange: Plant QGRs (PL); Green: Leaf QGRs (LF); Blue: Flower QGRs (FL). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) are shown in Table S3.
3.1. Plant (PL)
Plant-related traits can influence agronomic strategies for crop production. For example, by decreasing the plant height, the growing habitus of the plant can be changed, allowing different cultural methods and potentially improving the field productivity. No clear candidate genes have been reported in the literature for these traits, except for SmCPR1 (SMEL4.1_04g017430.1), a cytochrome P450 reductase putatively associated with plant dwarfism [74,75] and located on QGR PL6 (chr. E4).
3.2. Leaf (LF)
Understanding the mechanism of leaf development is essential to improve crop management, influencing plant productivity and stress tolerance. Small leaf mutants (slf) have been recently generated by ethyl methane sulfonate (EMS) mutagenesis [76]. Transcriptomic analysis indicated a dominance of the auxin signal during leaf development in mutated plants, allowing the identification of AUX1 (annotated as LAX5 in v4.1-SMEL4.1_01g003480.1), ARF5 (SMEL4.1_04g022210.1), and three Aux/IAA (SMEL4.1_05g020420.1, SMEL4.1_09g022160.1, and SMEL4.1_03g032430.1-QGR LF7) genes as potential candidates for the observed phenotype. The latter were proposed to be the main genes responsible for leaf growth and morphogenesis in the obtained mutants.
3.3. Flower (FL)
Sexual organs’ characteristics, such as ovary length, diameter, and hairiness, impact on the possibility of the flower being pollinated. Two QTLs hotspots were identified in QGR FL1 (chr. E1) and FL6 (chr. E2), associated with ovary length, ovary diameter, ovary hairs, flower shape, and peduncle length. A comparative proteomic analysis allowed the identification of differentially expressed proteins in heterostylous pistil development [77], highlighting the potential role of nine genes (Table S3) during flower development. Additionally, some proteins associated with programmed cell death were associated with S-morph pistils, belonging to flowers generally possessing a small and highly reduced gynoecium and lower productivity.
4. Prickles (PK)
Eggplant is the only solanaceous crop possessing a prickly phenotype. Prickles can be found on eggplant leaves, stems, and fruit calyxes and are modified glandular trichomes and cortical cells used as a defensive strategy against herbivore attacks, generally perceived as an undesirable commercial trait [78]. Many eggplant cultivars present prickles on the fruit calyx, since in certain world regions they are perceived of superior organoleptic quality, while the prickles on the vegetative tissues are generally absent, as the result of the positive selection in breeding programs [79]. Despite the several mapping studies that reported QTLs for this trait [24,27,42,62,63,80], the genetic basis of prickle formation in eggplant remains unclear. Data collection revealed a total of 115 QTLs and, on comparing these regions, 20 unique QGRs were defined (Table S2; Figure 2). The remaining nine QTLs were not included in the QGRs due to the lack of genomic position.
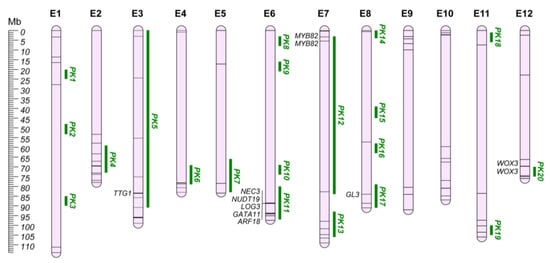
Figure 2.
Prickles’ QGRs (PK; Green) chromosome map (E1–E12). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) are shown in Table S3.
The QGR PK11, located on chr. E6, has been considered the main region responsible for eggplant prickles, and several mapping studies have located prickly related QTL in this region [24,42,79]. In addition, Portis et al. [63] confirmed such associations through GWA analysis. By fine-mapping the region (named PI locus), Miyatake et al. [80] delineated candidate genes encoding for: (i) carbonic anhydrases (SMEL4.1_06g026670.1, SMEL4.1_06g026680.1); (ii) nudix family hydrolases (SMEL4.1_06g026690.1); (iii) GATA transcription factor (SMEL4.1_06g026740.1); and (iv) Auxin Response Factor (SMEL4.1_06g026750.1). Furthermore, the PI locus-ascribed 0.5-kb deletion in the ‘Togenashi-senryo-nigo’ genotype was proposed as affecting the gene expression level of neighboring genes, particularly the downstream GATA11 (SMEL4_06g026740.1). Furthermore, comparing prickled and non-prickled genotypes, a selective sweep (SS) harboring genes encoding NUDT19 (SMEL4_06g026690.1), GATA11 (SMEL4_06g026740.1), and ARF18 (SMEL4_06g026750.1) was identified in the eggplant pangenome by Barchi et al. [68]. The SS on chr. E6 is close to the previously reported QTLs and QTNs, as well as the morphological marker PRICKLINESS [27] and the PI locus, suggesting a possible target for eggplant breeding improvement. In addition, the transcriptome analysis performed by Zhang et al. [78] identified the ARF18 gene (SMEL4.1_06g026750.1), which was located in the PI locus, as the key responsible for the formation of prickles, providing new insights into the regulatory molecular processes driving prickles’ morphogenesis in eggplant.
Another major QTL affecting the prickles’ development in the plant was recently identified on chr. E12 (QGR PK20) [79]. This genomic region has been thoroughly investigated underlying seven putative candidate genes involved in the prickle’s formation. Among these, SMEL4.1_12g013270.1 and SMEL4.1_12g013280.1, encoding a WUSCHEL-related homeobox 3B protein (WOX3), were proposed as candidates influencing calyx prickle formation. Indeed, higher expression levels of SMEL4.1_12g013280.1 in prickly individuals, and a 22-bp deletion affecting the second exon of the same gene in prickleless individuals suggest that WOX3 genes are likely involved in the development of calyx prickles in eggplant.
On chr. E7, two genomic loci controlling prickles’ traits have been identified (QGR PK12 and PK13). In particular, QGR PK12 was associated with an SS comprising two adjacent MYB82 (SMEL4.1_07g003480.1 and SMEL4.1_07g003490.1), which are homologous of the Arabidopsis GLABRA1 gene [68]. GLABRA1 plays a pivotal role in the formation of a trimeric complex with GLABRA3/ENHANCER of GL3 (GL3/EGL3) and TRANSPARENT TESTA GLABRA1 (TTG1), which is essential for the positive regulation of A. thaliana trichome initiation [78,81,82,83,84]. Both the BHLH family protein GL3 (SMEL4.1_08g024700.1) and the WD repeat family protein TTG1 (SMEL4.1_03g019420.1) are also located in QGRs PK17 (chr. E8) and PK5 (chr. E3), respectively [78].
5. Parthenocarpy and Male-Sterility (PT and MS)
Crop reproduction is tightly connected to plant productivity and fruit quality. If sexual behaviors, such as male sterility (MS) and self-incompatibility (SI), can be employed for hybrids’ production, seedlessness, as a result of parthenocarpy, is particularly appreciated by consumers [85,86].
Male sterility (MS) consists in the failure of plants to produce functional anthers, pollen, or male gametes. Male sterile mutants are classified into (i) structural, (ii) sporogenous, and (iii) functional types, based on anthers’ development and phenotype [87]. Male sterility is a useful trait in breeding programs, facilitating the production of hybrid seeds and avoiding emasculation, and genic male sterility (GMS) and cytoplasmic male sterility (CMS) have been proposed as causative mechanisms of this trait [88]. A number of genes responsible for both CMS and GMS are known for many plant species [88,89,90,91,92], and in eggplant, several mutants manifesting GMS caused by recessive nuclear genes were reported [93], as well as genes involved in CMS [94,95,96]. In the last decade, different biotechnological strategies have been tested [97,98], differentially expressed genes (DEGs) were identified (Table S3) [99,100,101,102], and the protein–protein interactions of SmCOI1 (SMEL4.1_05g001020.1) with SmOPR3 (SMEL4.1_07g003350.1) and SmJAZ1 (annotated as TIFY10A in v4.1-SMEL4.1_12g001970.1) were investigated [103,104], providing valuable information for the dissection of the genetic basis of male sterility.
Parthenocarpy (PT) is defined as the growth of the ovary into a fruit without pollination and/or fertilization, and results in the acquisition of seedless commercial varieties with a high fruit yield [105]. In eggplant berries, the presence of seeds causes a more intense and faster fruit pulp browning, due to oxidation of chlorogenic acid by polyphenol oxidases, and the biosynthesis of bitterness-related and flesh hardness-related compounds such as saponin and solasonin [106,107]. Furthermore, as sub-optimal environmental conditions negatively influence fruit yield and impact on reproductive processes (i.e., pollen formation, dispersal, germination, and fruit fertilization), parthenocarpic varieties represent a cost-effective solution to improve fruit set and growth in different environments [108]. In 2012, Miyatake et al. [109] investigated the genetic basis of parthenocarpy, reporting the trait as polygenic and identifying a major QTL on chr. E8 (~30% explained variability; Cop8.1; QGR PT2; Figure 3), and one on chr. E3 (Cop3.1; QGR PT1). Furthermore, several DEGs were reported (Table S3) [110], and SmARF8 (SMEL4.1_02g004290.1) [111] and Pad-1 (SMEL4.1_03g031670.1, annotated as ISS1) [112], an aminotransferase involved in auxin homeostasis, were recently highlighted as inducing parthenocarpy. The latter was reported to be mainly responsible for the Pad-1 locus identified on chr. E3, 10Mb upstream QGR PT1.
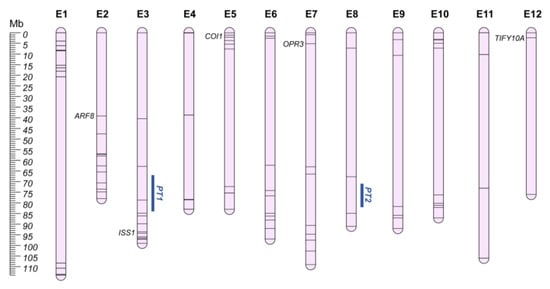
Figure 3.
Parthenocarpic QGRs (PT; Orange) and Male Sterility chromosome map (E1–E12). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) are shown in Table S3.
6. Fruit-Related Traits
Plant productivity and fruit quality have been the main focus of plant breeding for decades. Indeed, production directly impacts on producers’ acceptance of novel lines, with traits such as fruit weight and number of fruits affecting total yield [113]. On the other hand, fruit quality perception by the consumer is no more merely related to a morphological focus (fruit shape, glossiness, presence of seeds in the berry, pericarp firmness, and chlorophyll pigmentation) but also to a nutritional one [114]. Most of the nutrient properties of the eggplant fruit are related to vitamins, phenolic compounds, especially chlorogenic and hydroxycinnamic acids and their conjugates, and other phenylpropanoids [115]. Anti-nutritional compounds, such as steroidal glycoalkaloids and polyamine conjugates, are accumulated both in the flesh and peel as a toxic defense mechanism against herbivores, providing a bitter taste to the fruits [116]. As several traits are involved in fruit quality, to ease the search for regions of interest, QTLs were split into four categories: (i) shape; (ii) productivity; (iii) quality; and (iv) metabolites. The comprehensive list of the traits included in each category can be found in Table S2. A total of 304 QTLs were used for the identification of 79 distinct QGRs (Table S2; Figure 4), while 41 QTLs had insufficient information to retrieve their physical position.
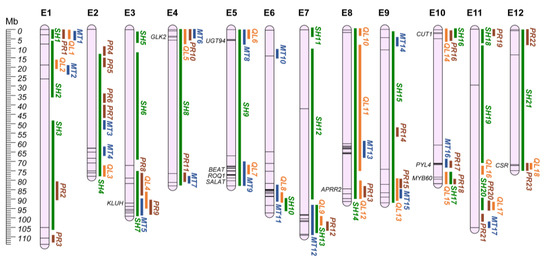
Figure 4.
Fruit-related QGRs chromosome map (E1–E12). Green: Shape QGRs (SH); Brown: Productivity (PR); Orange: Quality (QL); Blue: Metabolites (MT). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) are shown in Table S3.
6.1. Shape (SH)
Twenty-one QGRs, identified by 162 QTLs, were associated with shape-related traits. In these regions, 40 candidate genes for fruit shape were identified (Table S3) [53,68,117,118], including the SUN, OVATE and YABBY gene families. In tomato and pepper, SUN and OVATE have been associated with fruit elongation [119,120,121], while a YABBY transcription factor has been reported to be involved in fruit size determination associated with the fas locus in tomato [122]. The SUN-associated protein is a positive regulator of growth and has been proposed to be involved in fruit elongation and hormones or secondary metabolite levels [123], while Ovate family proteins (OFPs) have been identified as encoders of a negative regulator of fruit growth [119].
6.2. Productivity (PR)
Fifty-four QTLs were identified, defining 23 QGRs related to productivity traits. The genetic basis of fruit weight is poorly understood in the Solanaceae family, and in tomato few genomic regions have been strongly associated with the trait, with almost no candidate genes identified [124]. Mu et al. [125] identified a mutant allele in cell-size regulator (CSR-Solyc11g071940-fw11.3) genes associated with the domestication of tomato fruit, assessing its expansion in the Solanaceae family. In eggplant, we identified SMEL4.1_12g014140.1 through ortholog-driven gene mining, associated with QGR PR23, on chr. E12 (annotated as At5g22090 in V4.1). Recently, Li et al. [126] identified SlKLUH (Solyc03g114940), a CUP78A that positively regulates fruit weight by increasing the number of cell layers in the pericarp [127], as being mainly responsible for QTL fw.3.2 in tomato. Furthermore, pangenome analysis [128] revealed a positive association between SlKLUH copy number and fruit weight in tomato. CYP78A5 (SMEL4.1_03g027710.1) is its orthologous gene and is located in QGR PR9 (chr. E3). These two genes are, to our knowledge, the first candidates reported for fruit weight in eggplant.
6.3. Quality (QL)
Eighteen QGRs, identified by 42 QTLs, were associated with fruit quality-related traits. Six QGRs contained nine QTLs associated with chlorophyll pigmentation of the berry, generally corresponding with the presence of a flesh green ring after fruit cut and chlorophyll accumulation in the peel. Among the reported genes (Table S3), two were identified as the main genes responsible for chlorophyll-controlling regions on chrs. E4 and E8 [129]. Indeed, GLK2 (SMEL4.1_04g003340.1-chr. E4-QGR QL5) and APRR2 (SMEL4.1_08g020990.1-chr. E8-QGR QL12) have been reported to be promoters of chloroplast development in several solanaceous and cucurbitaceous crops regulating pigment accumulation [130,131,132,133,134]. For glossiness (i.e., accumulation of waxes in the berry epidermis) nine QTLs were identified in five QGRs (QL4, QL8, QL11, QL13, and QL15). In QGR QL15 (chr. E10), a MYB60 (SMEL4.1_10g024240.1-formerly annotated as MYB30), a regulator of cutin and wax biosynthesis and cuticle development [135,136,137,138,139], was reported to be involved in the expression of a 3-ketoacyl-CoA synthase 6 (CUT1, formerly reported as KCS6; SMEL4.1_10g001780.1) [140], falling within the QGR QL14. KCSs are a family of synthases involved in the biosynthesis of very long chain fatty acids, playing a key role in wax biosynthesis [141]. KCSs mutants have been reported as producing lower amounts of cuticular wax in Medicago truncatula [142], Arabidopsis spp. [143,144,145], Populus trichocarpa [146], and Brassica rapa [147].
6.4. Metabolites (MT)
Seventeen QGRs, identified by 46 QTLs, were associated with metabolites’ accumulation in the berry. In these regions, six candidate genes were reported as potentially involved in the metabolism of steroidal glycoalkaloids (SGAs) and polyamine conjugates. On chr. E10, the abscisic acid receptor PYL4 (QGR MT16; SMEL4.1_10g018120.1) was associated with pseudoprodioscin biosynthesis, while a UDP-glycosyltransferases (QGR MT8; UGT94; SMEL4.1_05g004840.1) was identified in a QTL for demissine on chr. E5. Four more genes were identified on chr. E5, potentially involved in polyamine conjugates accumulation: (i) TMV resistance protein N (QGR MT9; ROQ1; SMEL4.1_05g020480.1); (ii) acetyl-CoA-benzyl alcohol acetyltransferase (QGR MT9; BEAT; SMEL4.1_05g018170.1); (iii) an acyl-lipid (9-3)-desaturase (QGR MT13; SMEL4.1_08g012960.1); and (iv) a salutaridinol 7-O-acetyltransferase (QGR MT9; SALAT; SMEL4.1_05g020260.1), involved in alkaloid biosynthesis in Papaver somniferum [46].
7. Anthocyanins (AN)
Anthocyanins are an important class of flavonoids, glycosylated polyphenolic compounds that represent a vast class of plant pigments, with a range of color from orange to blue [148]. These plant secondary metabolites with high antioxidant capabilities play an important role in plant reproduction by attracting pollinators, protecting plants from several biotic and abiotic stresses. Anthocyanins’ accumulation avoids lipid peroxidation and maintains membrane integrity, lowering cell senescence, and improving vegetables’ postharvest performance [149]. In plants, the most common anthocyanins are derived from the metabolism of six anthocyanidins, namely pelargonidin, cyanidin, delphinidin, peonidin, petunidin, and malvidin [150]. In violet/black eggplant, as well as in pepper, the only anthocyanins reported to be accumulated are derived from delphinidin, which can also be present in the vegetative organs of the plants. In the fruits, the delphinidin level is higher at the unripe stage and decreases upon ripening to complete disappearance [115]. Delphinidin-3-(p-coumaroyl-rutinoside)-5-glucoside, commonly known as nasunin, is the most frequent anthocyanin structure in pepper and eggplant fruits [151]. In addition, some eggplant accessions have been observed accumulating a non-acylated anthocyanin, delphinidin-3-rutinoside [44]. The genetic control of anthocyanin biosynthesis, its distribution, and accumulation in Solanaceae species, including eggplant, has been extensively studied [152,153,154,155,156,157,158,159], and candidate genes included in the defined QGRs were retrieved from the literature (Table S3). The primary level of regulation for anthocyanin biosynthesis is the expression of regulatory and structural biosynthetic genes. Structural genes are classified as early (EBG; chalcone–flavonone synthase-CHS; chalcone–flavonone isomerase-CHI; flavanone 3-hydroxylase-F3H) and late (LBG; flavonoid 3′-hydroxylase-F3′H; flavonoid 3′,5′-hydroxylase-F3′5′H; dihydroflavonol 4-reductase-DFR; anthocyanidin synthase-ANS; flavonoid 3-O-glucosyltransferase-UFGT; flavonol synthase-FLS) biosynthetic genes [160,161,162]. Data collection indicated 153 QTLs as associated with anthocyanin levels in various organs, grouped in 20 QGRs (Table S2; Figure 5). Four papers additionally identified 17 QTLs that lacked information on their chromosomal position.

Figure 5.
Anthocyanins QGRs (AN; Blue) chromosome map (E1–E12). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) are shown in Table S3.
By focusing on highly QTL-dense QGRs (AN9, AN16, AN18), four structural genes, two CHS (SMEL4.1_05g000250.1 and SMEL4.1_09g023150.1) and two CHI (SMEL4.1_05g001480.1 and SMEL4.1_10g016630.1), were identified by several studies as closely associated with anthocyanin accumulation under different conditions. Moreover, the MYB-bHLH-WD40 (MBW) complex, which is composed of the MYB, basic helix-loop-helix (bHLH), and WD40 repeat families, has been proposed as the main regulatory element for anthocyanin accumulation in Solanaceae. Among the reported regulatory candidates, three MYB1s (SMEL4.1_10g019180.1, SMEL4.1_05g015570.1, SMEL4.1_01g009630.1), biosynthetic activators formerly named MYB113 [163,164,165,166,167], SmelMYBL1 (SMEL4.1_10g000420.1, anthocyanin repressor) commonly known as MYB4, and a BHLH42 (SMEL4.1_09g014720.1) [168,169], also called TRANSPARENT TESTA 8 (TT8), were included in the defined QGRs (AN1, AN9, AN15, AN17, AN18). Environmental variables, such as light [170,171] and temperature [172], can also affect anthocyanin metabolism. CRY1 (SMEL4.1_05g017270.1), COP1 (SMEL4.1_10g002450.1), SPA3 (SMEL4.1_10g004500.1), the main genes reported to be responsible for light-dependent anthocyanin pigmentation, were associated with QGR AN9 and AN17 [173,174,175,176]. Finally, anthocyanin pigmentation is associated not only with biosynthetic elements, but enzymatic and non-enzymatic factors can interact in the degradation of the pigments, leading to the regulation of anthocyanin discoloration mechanisms [151]. A number of studies have reported potential candidate genes involved in these mechanisms, and the one included in the defined QGRs can be found in Table S3.
8. Biotic Resistances (RS)
8.1. Pathogens’ Resistance
World-wide, plant pathogens and pests are among the major effectors in crop production, affecting yield and strongly impacting on social, environmental, and economic costs [177]. As the climate changes, enlarging the geographical area suitable for their establishment and growth, these organisms can spread more easily, requiring new strategies for their control. Eggplant production can be drastically affected by pathogens, with fungal and bacterial wilts representing the main hazards in many parts of the globe. Bacterial wilt is caused by the Ralstonia solanacearum species complex, a soil-borne pathogen well adapted to tropical/subtropical regions [178], while fungal wilts are generally caused by Verticillium dahliae and Fusarium oxysporum f. sp. melongenae, mostly causing more than 50% of yield loss [179]. As has occurred in different crops, human selection has caused an erosion of the genetic variability of the cultivated germplasm, leading to a reduction in the number of resistant/tolerant genotypes that have been conventionally applied in breeding programs [180]. For this reason, wild and allied relatives have been employed for the introgression of resistance traits in cultivated eggplants. Data collection revealed a total of 66 QTLs for resistance/tolerance to fungal and bacterial wilt, identifying 14 QGRs (Table S2; Figure 6). Thirty-seven QTLs were not included in the QGRs due to the lack of sufficient information to establish their chromosomal position.
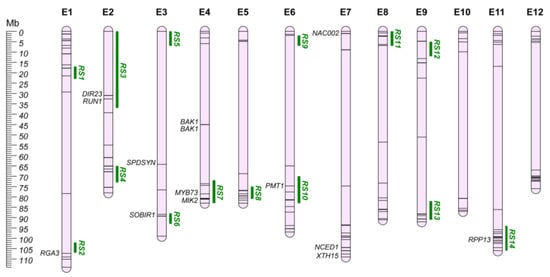
Figure 6.
Pathogen resistance QGRs (RS; Green) and pest resistance chromosome map (E1–E12). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) are shown in Table S3.
DIR proteins are involved in the biosynthesis of cell wall lignins and lignans, playing a key role in abiotic and biotic stress tolerance [181,182,183,184,185]. A recent genome-wide identification of the eggplant DIR gene family identified potential candidates for biotic resistance, reported to be involved in ROS accumulation and callose deposition in the infection sites (Table S3) [186]. Among them, seven DIRs were included in the defined QGRs, in QTLs associated both with fungal and bacterial wilt.
Four candidate genes for bacterial wilt resistance aligned with the defined QGR regions, both originating from synthenic and transcriptomic approaches. On chr. E4, MIK2 (QGR RS7; SMEL4.1_04g016080.1) has been reported as maintaining cell wall integrity in resistant genotypes, while SOBIR1 (QGR RS6; SMEL4.1_03g024090.1) seems to be involved in plant cell death [187]; these two genes activated the pathogen-resistance responses in Arabidopsis [188,189]. Salgon et al. [39] identified tomato synthenic regions for eggplant resistance to R. solanacearum (Table S3). By mining them, we identified a putative disease resistance protein RGA from S. bulbocastanum (RGA3; SMEL4.1_01g035130.1) on chr. E1 (200kb upstream QGR RS2) as potentially responsible for the reported RE-bw resistance locus [190]. Finally, SmMYB44 (SMEL4.1_04g019540.1; annotated as MYB73 in v4.1; QGR RS7) has been reported as regulator of the spermidine synthases SmSPDS (SMEL4.1_03g012150.1) and SmSPDS-like (SME4L.1_06g012830.1, annotated as PMT1 in v4.1; QGR RS10), leading to spermidine accumulation and resistance to R. solanacearum in eggplants [191].
For fungal wilt resistance, two QGRs were identified on chr. E2 (QGR RS3) and E11 (QGR RS14). In QGR RS3, dirigent protein 23 (DIR23; SMEL4.1_02g003080.1) was proposed by Barchi et al. [37] as associated with the FomE02.01 resistance locus. This locus was recently investigated by Tassone et al. [192] through the BSAseq approach, assessing the introgression of the resistance locus from S. aethiopicum. Thanks to the availability of the eggplant pangenome [68], ten potential candidate genes were identified on the S. aethiopicum genome. Among them, RES1 was described as a putative TMV resistance protein N-like (Solyc02g032200.2) [43]. This gene was annotated as a disease resistance protein RUN1 (SMEL4.1_02g003050.1) in the 4.1 version of the eggplant genome. RUN1 proteins have been reported to be involved in ROS accumulation and callose deposition in the infection sites after pathogens’ inoculum, providing resistance to fungal penetration in the tissue, and further absence of hypha proliferation [193]. On chr. E11 (QGR RS14), three candidates, including a putative late-blight resistance protein (R1C-3; SMEL4.1_00g001090.1) and two homologs of RPP13, were selected by Tassone et al. [192] as mainly responsible for the FomCH11 locus. The latter were annotated as proteins of unknown function in the last version of the reference genome, while the annotation of the other six candidates (Table S3) in version v4.1 was consistent with the one reported in the literature.
8.2. Pests Resistance
Insects and nematodes can drastically affect plant productivity, causing a wide range of damage both to the vegetative tissues and the berries [194,195]. The genetic basis of eggplant resistance/susceptibility to pests has been poorly investigated, producing only a few transcriptomic works on the topic. Recently, eggplant and tomato were compared for their biochemical and transcriptomic reaction to Tuta absoluta (Meyrick) attack [196]. This insect is a leaf miner whose invasion is seriously threatening the commercial tomato industry, easily spreading on other Solanaceae [197]. Multi-omics analysis have been performed in tomato, the main model host for the pest, identifying a signaling cascade mediated by the JA complex as first transcriptional changes upon infection, followed by the activation of genes involved in trichomes’ growth and the biosynthesis of terpene volatiles and phenylpropanoids [198]. In eggplant, the transcriptomic analysis suggested gene regulation in ER protein processing and phenylpropanoid biosynthesis as main responsible in the inhibition of T. absoluta infestation (Table S3).
Root-knot nematodes (RKNs, Meloidogyne spp.) are endoparasites that attack many cultivated plants, seriously threatening global food safety and production [199]. The southern root-knot nematode Meloidogyne incognita is one of the main eggplant nematode parasites [200]. Under invasion conditions, plants react with a wide range of defense mechanisms, including phytohormone biosynthesis (e.g., auxin, cytokinins, salicylic acid, jasmonate, gibberellin, abscisic acid, and brassinosteroids) and modifications in the cell wall composition [201,202]. Zhang et al. [203] investigated the transcriptomic changes in the gene expression of Solanum torvum (Sw.), reported to be less susceptible to the nematode, and eggplant under M. incognita infestation, and reported several DEGs potentially associated with the pathogen tolerance and susceptibility (Table S3). Among them, 13 nucleotide-binding site–leucine-rich repeat (NBS-LRR) resistance genes were upregulated in eggplant, suggesting their role in the plants’ reaction to pathogen-related damages. Interestingly, NCED1 (SMEL4.1_07g020880.1), the key enzyme in the defense response mediated by ABA biosynthesis, was upregulated in S. torvum, together with two BAK1s (SMEL4.1_04g008770.1 and SMEL4.1_04g008780.1), associated with brassinosteroids biosynthesis, mainly proposed to interact with RKN effectors during invasions [204]. Finally, two xyloglucan endotransglucosylases, associated with structural changes in cell wall expansion, essential for nematode feeding sites’ formation [205], were repressed in S. torvum (XTH15-SMEL4.1_07g020690.1-and NAC002-SMEL4.1_07g000510.1).
9. Abiotic Resistances
It is clear that the breeding focus in the next few years will be targeted to tolerance and resistance to the main biotic and abiotic stresses, especially in the climate change scenario. Modern eggplant varieties are generally susceptible to several abiotic stresses, including drought, salinity, low and high temperatures, and soil toxicity [206,207,208,209,210]. Thus, a deeper knowledge of the genetic mechanisms involved in the tolerance of such stresses is required to develop new breeding materials able to face and rapidly recover from suboptimal growing conditions. While classical mapping studies and GWA panels have poorly investigated the genetic elements providing tolerance to the main abiotic stresses, great efforts have been focused on transcriptome analysis of sensitive/tolerant accessions under different environmental conditions. Recently, Toppino et al. [211] reviewed in depth the available material for abiotic stresses in eggplant. Here, we provide a selection of the main candidate genes to be explored in the development of novel high-value eggplant cultivars (Table S3; Figure 7).
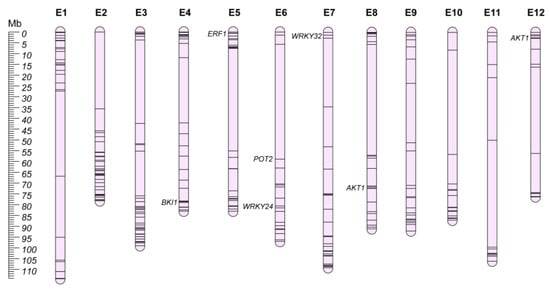
Figure 7.
Abiotic resistance chromosome map (E1–E12). Main candidate genes are reported in italics, while information about the other candidates (represented by bars on the chromosomes) is shown in Table S3.
9.1. Osmotic Stress
Osmotic stress represents one of the most important environmental aspects that can negatively impact crop growth and productivity [212], causing an increase in carotenoid and proline content [213]. Drought and water scarcity also negatively affects nitrogen, phosphorus, and potassium uptake, decreasing total soluble solids (TSS), increasing total phenols, superoxide dismutase (SOD), glutathione reductase (GR), electrolyte leakage, pH, and vitamin C [214]. Photosynthetic pigments’ reduction, together with proline, malondialdehyde, total phenolics, and total flavonoids’ accumulation have been reported to be the main effects associated with the water stress in eggplant [210]. Not only water scarcity, but also other environmental conditions (e.g., high salinity and temperatures, land flooding and soil contamination) can produce osmotic stress in plants [215,216,217,218]. For this reason, a wide range of common expression patterns have been observed in reaction to different abiotic stressors [219,220,221]. Stress-associated proteins (SAP), NAC transcription factors, apetala2/ethylene responsive factor (AP2/ERF), and DNA methyltransferases have been reported to be constantly upregulated in eggplant under abiotic stress conditions (Table S3) [222,223,224,225,226,227], while C-repeat binding factors (CBFs) have been proposed as early-stage effectors in the plant response to osmotic and cold stress [228]. For instance, the role of SmERF1 (SMEL4.1_05g001670.1) was validated under salinity stress by virus-induced gene silencing assay (VIGs), enhancing susceptibility to abiotic stress and downregulating expression levels of other stress defense-related genes [225].
9.2. Salt Toxicity Stress
Soil salinity has a negative impact on plant growth, fruit quality, and yield [229] and eggplant has been reported to be moderately susceptible to salinity when compared to other Solanaceae [230]. Different studies have reported an association between salt-induced growth reduction and high accumulation of Na+ and Cl− in both roots and shoots, causing stomata closure and increasing leaf turgor potential [231,232]. Furthermore, calcium (Ca2+) and potassium (K+) concentrations, water consumption, and the K+/Na+ ratio have been reported to decrease under salinity stress [232,233]. Overall, an excess of NaCl appears to reduce seed germination [234], roots and shoots’ growth, chlorophyll content, and the photosynthetic rates, ending in a reduction in fruit yield [235]. At present, the genetic control of plant reaction to salt accumulation has been poorly investigated, but salts transport mechanisms appear to play a key role in cell detoxification [236], and transcriptome analysis identified a series of transcription factors and structural genes associated with K+ and Na+ homeostasis (Table S3) [233]. Among them, the interaction of SmAKT1 (SMEL4.1_08g015230.1 and SMEL4.1_12g001280.1) and SmSOS1 (annotated as POT2; SMEL4.1_06g009410.1) was proposed to regulate Na+ transport and accumulation in leaves.
9.3. Heat Stress
As the climate is gradually becoming warmer, seedlings’ growth, flower development, fruit set and growth can be drastically compromised by high temperatures that can be scored during summer periods [237]. Furthermore, as high temperatures fasten up fruit ripening leading to significant decreases in total anthocyanin content, heat stress can also harm fruit quality [238,239,240]. Under heat stress, plant cells respond by inducing the expression of genes encoding heat shock proteins (Hsps), involved in preventing heat-related damage and conferring thermotolerance [241]. Generally, these proteins behave as molecular chaperones, preventing protein misfolding and aggregation, maintaining protein homeostasis in cells [242]. Furthermore, specific transcription factors (i.e., heat shock factors-Hsfs) have been reported to control and regulate Hsps’ expression and activation in the cell [243]. Recently, Gong et al. [244] performed a genome-wide identification of Hsps and Hsfs in eggplant, followed by transcriptomic analysis on two inbred lines, contrasting for heat tolerance. The results highlighted that Hsgs and Hsps, belonging to Hsp60, Hsp70, Hsp90, and Hsp100 protein families, were induced by heat stress treatment in the thermotolerant inbred line (Table S3). Hsp70 and Hsp100 families and Hsf class A and B were previously reported by Zhang et al. [239] and Wang et al. [245] to be differentially expressed under heat stress conditions, together with a number of transcription factors (e.g., MYB, ERF/DREB, NAC; Table S3), suggesting potential candidates for elucidating thermotolerance mechanisms in eggplant.
9.4. Cold Stress
Contrasting with summer high temperatures, low temperatures in the early stage of cultivation have been recorded in recent years. Cold stress limits plant growth, development, and production, and eggplant appears to be much more sensitive to it compared with other solanaceous crops [246]. Eggplant grows slowly when the temperature is below 17 °C, suffers rapid physiological disorders below 10 °C, and undergoes chilling injury near 7.2 °C [247]. Furthermore, chilling injuries can occur, causing rapid low-pollen viability, plant aging, fruit skin shrinkage, and calyx deterioration and browning [207,248]. Cold sensitivity has been reported to be enhanced by the effect of brassinosteroids (BR), and BKI1 (SMEL4.1_04g020080.1), under-expressed in sensitive genotypes, was reported to regulate the low temperature-induced BR signal in eggplant [249]. In addition, transcriptomic analysis revealed that a wide number of DEGs were represented by transcription factor families (e.g., AP2/ERF, C2H2, WRKY, bHLH, NAC, and MYB-related; Table S3), and the downregulation of two WRKY transcription factors-SmWRKY26 (annotated as WRKY24 in v.4.1; SMEL4.1_06g016680.1) and SmWRKY32 (SMEL4.1_07g001740.1) through VIGs increased eggplant sensitivity to cold stress, aggravating injuries caused by low temperature [250].
9.5. Heavy Metals Stress
The presence of high concentrations of heavy metals (e.g., cadmium, chromium, lead, and nickel) in the soil may have a toxic effect for eggplant [251,252], leading also to the accumulation of such elements in the fruits [253,254]. While soil toxicity effects in eggplant have not been investigated, S. torvum has been used as grafting material to improve Cd toxicity and plant resilience [252,255,256,257]. Recently, Cui et al. [258] investigated the methylation impact of S. torvum grafting on eggplant genes involved in sulfur metabolism, associated with a lower accumulation of Cd in aerial tissues, highlighting that grafting regulates S metabolism genes (e.g., STR, MGL, CGS, SULTR21, DCYD, and SUR; Table S3), enhancing S absorption and translocation in plants and modulating Cd accumulation.
9.6. Low Nitrate Stress
Nitrogen fertilization affects plant vigor, leaf chlorophyll content, fruit settings, dry matter production, and ascorbic acid content [259], as well as flower number, fruit pH and total solid content, fruit weight, and seed number [260]. The development of cultivars with higher N uptake, translocation, and use efficiency, i.e., nitrogen-use-efficiency (NUE) would lower production costs, and will be one of the main challenges to maintain high yields in a sustainable agriculture. However, limited information on genetic variation for this trait is available for eggplant, whose productivity is highly sensitive to N fertilization [261,262]. For example, transcriptomic analysis on four eggplant lines reported upregulation of light-harvesting complexes (LHCs) genes and ferredoxin–NADP reductases (FNRs) in the N-use genotypes, impacting on photosynthetic efficiency [263]. Furthermore, genes involved in responses to inorganic substances, abiotic stimuli, and chemicals were also differentially expressed between contrasting genotypes (Table S3) [264]. The WRKY33 (annotated as WRKY24 in v.4.1; SMEL4.1_06g016680.1) transcription factor have been associated with MAP kinases, YLS9, and auxin-responsive family genes upregulation, potentially promoting the development of a more efficient root system, as confirmed by overexpressing the orthologue transcription factor in Arabidopsis.
10. Concluding Remarks
In the last few decades, the genetic basis of eggplant traits in modern cultivars and their relatives have been investigated by several publications, but methodological differences have made it difficult to efficiently compare their outputs. QGRs here defined represent the regions that most likely contain genetic elements that regulate eggplants’ phenotypes. However, the presence of large QGRs, probably linked to experimental and methodological limits, suggest a need for further dissection of these regions through high-resolution and fine-mapping approaches. This review summarized the state of the art on the understanding of the genetic mechanisms regulating the main agronomical, qualitative and resistance eggplant trait, and the data here organized might find application in future breeding challenges. The information on QTLs here provided can be employed to assist in marker-assisted breeding programs for introducing high-impact regions into superior germplasm. Furthermore, potential candidate genes found within QTL regions can be selected and their effects can be examined in vivo through techniques such as CRISPR-CAS9 gene editing or transient manipulation of gene expression. This can lead not only to the identification of the genes that control a particular trait, but also to the detection of genetic elements responsible for trait variation. These variations, known as functional markers, are the most efficient molecular markers for marker-assisted selection (MAS) because they are directly linked to the trait and, unlike genetically linked markers, do not require validation in other populations. Indeed, both genetic elements and their interactions can pose challenges in varietal development. Pleiotropic, dominant, and epistatic effects have been documented in the literature for multiple traits. For example, anthocyanin regulation is governed by a complex network of interactions and pleiotropic effects. Guan et al. [265] reported a major QTL responsible for both leaf vein pigmentation and pericarp color that explains over 50% of phenotypic variability, while Salgon et al. [40] identified epistasis affecting polygenic resistance to R. pseudosolanacearum, where the epistatic effect accounted for 35.7% of total phenotypic variance. Such interactions, especially those of low impact, have been reported to be potentially biased by background QTLs [266]. Hence, a thorough understanding of the parental genetic background is crucial for the development of trait-focused breeding programs, and relevant information can be obtained from the original articles cited in these reviews.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12051016/s1, Table S1: List of the manuscripts reporting QTLs, including information on the experimental populations; Table S2: List of the QTLs reported in literature; Table S3: List of the candidate genes reported in literature.
Author Contributions
Conceptualization, E.P. and L.B.; methodology, E.P., L.B., M.M. and L.G; software, L.G.; investigation, M.M. and L.G.; data curation, L.G. and M.M.; writing—original draft preparation, L.G. and M.M.; writing—review and editing, L.B. and E.P.; visualization, E.P., L.B., M.M. and L.G.; supervision, E.P. and L.B.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially funded by the European Union’s Horizon 2020 Research and Innovation Programme under the Grant Agreement number 677379 (G2P-SOL project: ‘linking genetic resources, genomes and phenotypes of solanaceous crops’). The overall work also partially fulfills some goals of the Agritech National Research Center and received funding from the European Union Next-Generation EU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR)–MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4—D.D. 1032 17/06/2022, CN00000022). In particular, our study represents a review paper within the Task 4.1.1. (Spoke 4) titled: ‘Next-generation genotyping and -omics technologies for the molecular prediction of multiple resilient traits in crop plants’.
Data Availability Statement
Data are available at their original publicly accessible repository.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. Available online: https://www.fao.org/faostat/en/#home (accessed on 17 January 2023).
- Spooner, D.M.; McLean, K.; Ramsay, G.; Waugh, R.; Bryan, G.J. A Single Domestication for Potato Based on Multilocus Amplified Fragment Length Polymorphism Genotyping. Proc. Natl. Acad. Sci. USA 2005, 102, 14694–14699. [Google Scholar] [CrossRef] [PubMed]
- Portis, E.; Nervo, G.; Cavallanti, F.; Barchi, L.; Lanteri, S. Multivariate Analysis of Genetic Relationships between Italian Pepper Landraces. Crop Sci. 2006, 46, 2517–2525. [Google Scholar] [CrossRef]
- Bai, Y.; Lindhout, P. Domestication and Breeding of Tomatoes: What Have We Gained and What Can We Gain in the Future? Ann. Bot. 2007, 100, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Razifard, H.; Ramos, A.; Della Valle, A.L.; Bodary, C.; Goetz, E.; Manser, E.J.; Li, X.; Zhang, L.; Visa, S.; Tieman, D.; et al. Genomic Evidence for Complex Domestication History of the Cultivated Tomato in Latin America. Mol. Biol. Evol. 2020, 37, 1118–1132. [Google Scholar] [CrossRef]
- Tripodi, P.; Rabanus-Wallace, M.T.; Barchi, L.; Kale, S.; Esposito, S.; Acquadro, A.; Schafleitner, R.; van Zonneveld, M.; Prohens, J.; Diez, M.J.; et al. Global Range Expansion History of Pepper (Capsicum spp.) Revealed by over 10,000 Genebank Accessions. Proc. Natl. Acad. Sci. USA 2021, 118, e2104315118. [Google Scholar] [CrossRef]
- Naeem, M.; Demirel, U.; Yousaf, M.F.; Caliskan, S.; Caliskan, M.E. Overview on Domestication, Breeding, Genetic Gain and Improvement of Tuber Quality Traits of Potato Using Fast Forwarding Technique (GWAS): A Review. Plant Breed. 2021, 140, 519–542. [Google Scholar] [CrossRef]
- Page, A.; Gibson, J.; Meyer, R.S.; Chapman, M.A. Eggplant Domestication: Pervasive Gene Flow, Feralization, and Transcriptomic Divergence. Mol. Biol. Evol. 2019, 36, 1359–1372. [Google Scholar] [CrossRef]
- Arnoux, S.; Fraïsse, C.; Sauvage, C. Genomic Inference of Complex Domestication Histories in Three Solanaceae Species. J. Evol. Biol. 2021, 34, 270–283. [Google Scholar] [CrossRef]
- Vorontsova, M.S.; Stern, S.; Bohs, L.; Knapp, S. African Spiny Solanum (subgenus Leptostemonum, Solanaceae): A Thorny Phylogenetic Tangle. Bot. J. Linn. Soc. 2013, 173, 176–193. [Google Scholar] [CrossRef]
- Knapp, S.; Vorontsova, M.S.; Prohens, J. Wild Relatives of the Eggplant (Solanum melongena L.: Solanaceae): New Understanding of Species Names in a Complex Group. PLoS ONE 2013, 8, e57039. [Google Scholar] [CrossRef]
- Barchi, L.; Acquadro, A.; Alonso, D.; Aprea, G.; Bassolino, L.; Demurtas, O.; Ferrante, P.; Gramazio, P.; Mini, P.; Portis, E.; et al. Single Primer Enrichment Technology (SPET) for High-Throughput Genotyping in Tomato and Eggplant Germplasm. Front. Plant Sci. 2019, 10, 1005. [Google Scholar] [CrossRef] [PubMed]
- Acquadro, A.; Barchi, L.; Gramazio, P.; Portis, E.; Vilanova, S.; Comino, C.; Plazas, M.; Prohens, J.; Lanteri, S. Coding SNPs Analysis Highlights Genetic Relationships and Evolution Pattern in Eggplant Complexes. PLoS ONE 2017, 12, e0180774. [Google Scholar] [CrossRef] [PubMed]
- Barchi, L.; Pietrella, M.; Venturini, L.; Minio, A.; Toppino, L.; Acquadro, A.; Andolfo, G.; Aprea, G.; Avanzato, C.; Bassolino, L.; et al. A Chromosome-Anchored Eggplant Genome Sequence Reveals Key Events in Solanaceae Evolution. Sci. Rep. 2019, 9, 11769. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, P.; Prohens, J.; Vilanova, S.; Gramazio, P.; Plazas, M. Phenotyping of Eggplant Wild Relatives and Interspecific Hybrids with Conventional and Phenomics Descriptors Provides Insight for Their Potential Utilization in Breeding. Front. Plant Sci. 2016, 7, 677. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Martínez, E.; Adalid-Martínez, A.M.; García-Martínez, M.D.; Mangino, G.; Raigón, M.D.; Plazas, M.; Gramazio, P.; Prohens, J.; Vilanova, S. Fruit Composition of Eggplant Lines with Introgressions from the Wild Relative S. incanum: Interest for Breeding and Safety for Consumption. Agronomy 2022, 12, 266. [Google Scholar] [CrossRef]
- Tan, S.; Cheng, J.-W.; Zhang, L.; Qin, C.; Nong, D.-G.; Li, W.-P.; Tang, X.; Wu, Z.-M.; Hu, K.-L. Construction of an Interspecific Genetic Map Based on InDel and SSR for Mapping the QTLs Affecting the Initiation of Flower Primordia in Pepper (Capsicum spp.). PLoS ONE 2015, 10, e0119389. [Google Scholar] [CrossRef] [PubMed]
- Barrantes, W.; López-Casado, G.; García-Martínez, S.; Alonso, A.; Rubio, F.; Ruiz, J.J.; Fernández-Muñoz, R.; Granell, A.; Monforte, A.J. Exploring New Alleles Involved in Tomato Fruit Quality in an Introgression Line Library of Solanum Pimpinellifolium. Front. Plant Sci. 2016, 7, 1172. [Google Scholar] [CrossRef]
- Rambla, J.L.; Medina, A.; Fernández-del-Carmen, A.; Barrantes, W.; Grandillo, S.; Cammareri, M.; López-Casado, G.; Rodrigo, G.; Alonso, A.; García-Martínez, S.; et al. Identification, Introgression, and Validation of Fruit Volatile QTLs from a Red-Fruited Wild Tomato Species. J. Exp. Bot. 2017, 68, 429–442. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohens, J.; Plazas, M.; Mangino, G.; Herraiz, F.J.; Vilanova, S. Development and Genetic Characterization of Advanced Backcross Materials and An Introgression Line Population of Solanum incanum in a S. melongena Background. Front. Plant Sci. 2017, 8, 1477. [Google Scholar] [CrossRef]
- Siddique, M.I.; Lee, J.-H.; Ahn, J.-H.; Kusumawardhani, M.K.; Safitri, R.; Harpenas, A.; Kwon, J.-K.; Kang, B.-C. Genotyping-by-Sequencing-Based QTL Mapping Reveals Novel Loci for Pepper yellow leaf curl virus (PepYLCV) Resistance in Capsicum annuum. PLoS ONE 2022, 17, e0264026. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An Introduction to Markers, Quantitative Trait Loci (QTL) Mapping and Marker-Assisted Selection for Crop Improvement: The Basic Concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Li, H.; Hearne, S.; Bänziger, M.; Li, Z.; Wang, J. Statistical Properties of QTL Linkage Mapping in Biparental Genetic Populations. Heredity 2010, 105, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Doganlar, S.; Frary, A.; Daunay, M.-C.; Lester, R.N.; Tanksley, S.D. Conservation of Gene Function in the Solanaceae as Revealed by Comparative Mapping of Domestication Traits in Eggplant. Genetics 2002, 161, 1713–1726. [Google Scholar] [CrossRef] [PubMed]
- Frary, A.; Doganlar, S.; Daunay, M.C.; Tanksley, S.D. QTL Analysis of Morphological Traits in Eggplant and Implications for Conservation of Gene Function during Evolution of Solanaceous Species. Theor. Appl. Genet. 2003, 107, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Eannetta, N.T.; Xu, Y.; Tanksley, S.D. A Detailed Synteny Map of the Eggplant Genome Based on Conserved Ortholog Set II (COSII) Markers. Theor. Appl. Genet. 2009, 118, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Gramazio, P.; Prohens, J.; Plazas, M.; Andújar, I.; Herraiz, F.J.; Castillo, E.; Knapp, S.; Meyer, R.S.; Vilanova, S. Location of Chlorogenic Acid Biosynthesis Pathway and Polyphenol Oxidase Genes in a New Interspecific Anchored Linkage Map of Eggplant. BMC Plant Biol. 2014, 14, 350. [Google Scholar] [CrossRef]
- Nunome, T.; Ishiguro, K.; Yoshida, T.; Hirai, M. Mapping of Fruit Shape and Color Development Traits in Eggplant (Solanum melongena L.) Based on RAPD and AFLP Markers. Breed. Sci. 2001, 51, 19–26. [Google Scholar] [CrossRef]
- Nunome, T.; Suwabe, K.; Iketani, H.; Hirai, M.; Wricke, G. Identification and Characterization of Microsatellites in Eggplant. Plant Breed. 2003, 122, 256–262. [Google Scholar] [CrossRef]
- Nunome, T.; Negoro, S.; Kono, I.; Kanamori, H.; Miyatake, K.; Yamaguchi, H.; Ohyama, A.; Fukuoka, H. Development of SSR Markers Derived from SSR-Enriched Genomic Library of Eggplant (Solanum melongena L.). Theor. Appl. Genet. 2009, 119, 1143–1153. [Google Scholar] [CrossRef]
- Barchi, L.; Lanteri, S.; Portis, E.; Stàgel, A.; Valè, G.; Toppino, L.; Rotino, G.L. Segregation Distortion and Linkage Analysis in Eggplant (Solanum melongena L.). Genome 2010, 53, 805–815. [Google Scholar] [CrossRef]
- Fukuoka, H.; Miyatake, K.; Nunome, T.; Negoro, S.; Shirasawa, K.; Isobe, S.; Asamizu, E.; Yamaguchi, H.; Ohyama, A. Development of Gene-Based Markers and Construction of an Integrated Linkage Map in Eggplant by Using Solanum Orthologous (SOL) Gene Sets. Theor. Appl. Genet. 2012, 125, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Portis, E.; Barchi, L.; Toppino, L.; Lanteri, S.; Acciarri, N.; Felicioni, N.; Fusari, F.; Barbierato, V.; Cericola, F.; Valè, G.; et al. QTL Mapping in Eggplant Reveals Clusters of Yield-Related Loci and Orthology with the Tomato Genome. PLoS ONE 2014, 9, e89499. [Google Scholar] [CrossRef] [PubMed]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP Discovery and Genetic Mapping Using Sequenced RAD Markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Barchi, L.; Lanteri, S.; Portis, E.; Acquadro, A.; Valè, G.; Toppino, L.; Rotino, G.L. Identification of SNP and SSR Markers in Eggplant Using RAD Tag Sequencing. BMC Genom. 2011, 12, 304. [Google Scholar] [CrossRef]
- Barchi, L.; Lanteri, S.; Portis, E.; Valè, G.; Volante, A.; Pulcini, L.; Ciriaci, T.; Acciarri, N.; Barbierato, V.; Toppino, L.; et al. A RAD Tag Derived Marker Based Eggplant Linkage Map and the Location of QTLs Determining Anthocyanin Pigmentation. PLoS ONE 2012, 7, e43740. [Google Scholar] [CrossRef]
- Barchi, L.; Toppino, L.; Valentino, D.; Bassolino, L.; Portis, E.; Lanteri, S.; Rotino, G.L. QTL Analysis Reveals New Eggplant Loci Involved in Resistance to Fungal Wilts. Euphytica 2018, 214, 20. [Google Scholar] [CrossRef]
- Hirakawa, H.; Shirasawa, K.; Miyatake, K.; Nunome, T.; Negoro, S.; Ohyama, A.; Yamaguchi, H.; Sato, S.; Isobe, S.; Tabata, S.; et al. Draft Genome Sequence of Eggplant (Solanum melongena L.): The Representative Solanum Species Indigenous to the Old World. DNA Res. 2014, 21, 649–660. [Google Scholar] [CrossRef]
- Salgon, S.; Jourda, C.; Sauvage, C.; Daunay, M.-C.; Reynaud, B.; Wicker, E.; Dintinger, J. Eggplant Resistance to the Ralstonia Solanacearum Species Complex Involves Both Broad-Spectrum and Strain-Specific Quantitative Trait Loci. Front. Plant Sci. 2017, 8, 828. [Google Scholar] [CrossRef]
- Salgon, S.; Raynal, M.; Lebon, S.; Baptiste, J.-M.; Daunay, M.-C.; Dintinger, J.; Jourda, C. Genotyping by Sequencing Highlights a Polygenic Resistance to Ralstonia Pseudosolanacearum in Eggplant (Solanum melongena L.). Int. J. Mol. Sci. 2018, 19, 357. [Google Scholar] [CrossRef]
- Lebeau, A.; Gouy, M.; Daunay, M.C.; Wicker, E.; Chiroleu, F.; Prior, P.; Frary, A.; Dintinger, J. Genetic Mapping of a Major Dominant Gene for Resistance to Ralstonia Solanacearum in Eggplant. Theor. Appl. Genet. 2013, 126, 143–158. [Google Scholar] [CrossRef]
- Frary, A.; Frary, A.; Daunay, M.-C.; Huvenaars, K.; Mank, R.; Doğanlar, S. QTL Hotspots in Eggplant (Solanum melongena) Detected with a High Resolution Map and CIM Analysis. Euphytica 2014, 197, 211–228. [Google Scholar] [CrossRef]
- Miyatake, K.; Saito, T.; Negoro, S.; Yamaguchi, H.; Nunome, T.; Ohyama, A.; Fukuoka, H. Detailed Mapping of a Resistance Locus against Fusarium Wilt in Cultivated Eggplant (Solanum melongena). Theor. Appl. Genet. 2016, 129, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping Quantitative Trait Loci Affecting Biochemical and Morphological Fruit Properties in Eggplant (Solanum melongena L.). Front. Plant Sci. 2016, 7, 256. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, W.; Hu, T.; Hu, H.; Wang, J.; Bao, C. Construction of a SNP-Based Genetic Map Using SLAF-Seq and QTL Analysis of Morphological Traits in Eggplant. Front. Genet. 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Sulli, M.; Barchi, L.; Toppino, L.; Diretto, G.; Sala, T.; Lanteri, S.; Rotino, G.L.; Giuliano, G. An Eggplant Recombinant Inbred Population Allows the Discovery of Metabolic QTLs Controlling Fruit Nutritional Quality. Front. Plant Sci. 2021, 12, 638195. [Google Scholar] [CrossRef] [PubMed]
- Toppino, L.; Barchi, L.; Mercati, F.; Acciarri, N.; Perrone, D.; Martina, M.; Gattolin, S.; Sala, T.; Fadda, S.; Mauceri, A.; et al. A New Intra-Specific and High-Resolution Genetic Map of Eggplant Based on a RIL Population, and Location of QTLs Related to Plant Anthocyanin Pigmentation and Seed Vigour. Genes 2020, 11, 745. [Google Scholar] [CrossRef]
- Mangino, G.; Plazas, M.; Vilanova, S.; Prohens, J.; Gramazio, P. Performance of a Set of Eggplant (Solanum melongena) Lines With Introgressions From Its Wild Relative S. incanum Under Open Field and Screenhouse Conditions and Detection of QTLs. Agronomy 2020, 10, 467. [Google Scholar] [CrossRef]
- Bhanushree, N.; Saha, P.; Tomar, B.S.; Gopala Krishnan, S.; Gurung, B.; Ghoshal, C.; Das Munshi, A. Single Marker Analysis and Mapping of QTLs Governing Fruit Weight in Eggplant (Solanum melongena L.). Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 769–775. [Google Scholar] [CrossRef]
- Mangino, G.; Vilanova, S.; Plazas, M.; Prohens, J.; Gramazio, P. Fruit Shape Morphometric Analysis and QTL Detection in a Set of Eggplant Introgression Lines. Sci. Hortic. 2021, 282, 110006. [Google Scholar] [CrossRef]
- Villanueva, G.; Rosa-Martínez, E.; Şahin, A.; García-Fortea, E.; Plazas, M.; Prohens, J.; Vilanova, S. Evaluation of Advanced Backcrosses of Eggplant with Solanum elaeagnifolium Introgressions under Low N Conditions. Agronomy 2021, 11, 1770. [Google Scholar] [CrossRef]
- Qian, Z.; Ji, Y.; Li, R.; Lanteri, S.; Chen, H.; Li, L.; Jia, Z.; Cui, Y. Identifying Quantitative Trait Loci for Thousand Grain Weight in Eggplant by Genome Re-Sequencing Analysis. Front. Genet. 2022, 13, 841198. [Google Scholar] [CrossRef] [PubMed]
- Mangino, G.; Arrones, A.; Plazas, M.; Pook, T.; Prohens, J.; Gramazio, P.; Vilanova, S. Newly Developed MAGIC Population Allows Identification of Strong Associations and Candidate Genes for Anthocyanin Pigmentation in Eggplant. Front. Plant Sci. 2022, 13, 847789. [Google Scholar] [CrossRef] [PubMed]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-Wide Association Studies. Nat. Rev. Methods Prim. 2021, 1, 59. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sallam, A.; Stephen Baenziger, P.; Börner, A. GWAS: Fast-Forwarding Gene Identification and Characterization in Temperate Cereals: Lessons from Barley—A Review. J. Adv. Res. 2020, 22, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Sul, J.H.; Martin, L.S.; Eskin, E. Population Structure in Genetic Studies: Confounding Factors and Mixed Models. PLoS Genet. 2018, 14, e1007309. [Google Scholar] [CrossRef]
- Wang, M.; Fang, Z.; Yoo, B.; Bejerano, G.; Peltz, G. The Effect of Population Structure on Murine Genome-Wide Association Studies. Front. Genet. 2021, 12, 745361. [Google Scholar] [CrossRef]
- Mancin, E.; Lourenco, D.; Bermann, M.; Mantovani, R.; Misztal, I. Accounting for Population Structure and Phenotypes From Relatives in Association Mapping for Farm Animals: A Simulation Study. Front. Genet. 2021, 12, 642065. [Google Scholar] [CrossRef]
- Cano-Gamez, E.; Trynka, G. From GWAS to Function: Using Functional Genomics to Identify the Mechanisms Underlying Complex Diseases. Front. Genet. 2020, 11, 424. [Google Scholar] [CrossRef]
- Ge, H.Y.; Liu, Y.; Zhang, J.; Han, H.Q.; Li, H.Z.; Shao, W.T.; Chen, H.Y. Simple Sequence Repeat-Based Association Analysis of Fruit Traits in Eggplant (Solanum melongena). Genet. Mol. Res. 2013, 12, 5651–5663. [Google Scholar] [CrossRef]
- Cericola, F.; Portis, E.; Toppino, L.; Barchi, L.; Acciarri, N.; Ciriaci, T.; Sala, T.; Rotino, G.L.; Lanteri, S. The Population Structure and Diversity of Eggplant from Asia and the Mediterranean Basin. PLoS ONE 2013, 8, e73702. [Google Scholar] [CrossRef]
- Cericola, F.; Portis, E.; Lanteri, S.; Toppino, L.; Barchi, L.; Acciarri, N.; Pulcini, L.; Sala, T.; Rotino, G.L. Linkage Disequilibrium and Genome-Wide Association Analysis for Anthocyanin Pigmentation and Fruit Color in Eggplant. BMC Genom. 2014, 15, 896. [Google Scholar] [CrossRef]
- Portis, E.; Cericola, F.; Barchi, L.; Toppino, L.; Acciarri, N.; Pulcini, L.; Sala, T.; Lanteri, S.; Rotino, G.L. Association Mapping for Fruit, Plant and Leaf Morphology Traits in Eggplant. PLoS ONE 2015, 10, e0135200. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome Analysis of Multiple Pathogenic Isolates of Streptococcus Agalactiae: Implications for the Microbial “Pan-Genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef]
- Ou, L.; Li, D.; Lv, J.; Chen, W.; Zhang, Z.; Li, X.; Yang, B.; Zhou, S.; Yang, S.; Li, W.; et al. Pan-Genome of Cultivated Pepper (Capsicum) and Its Use in Gene Presence–Absence Variation Analyses. New Phytol. 2018, 220, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Gonda, I.; Sun, H.; Ma, Q.; Bao, K.; Tieman, D.M.; Burzynski-Chang, E.A.; Fish, T.L.; Stromberg, K.A.; Sacks, G.L.; et al. The Tomato Pan-Genome Uncovers New Genes and a Rare Allele Regulating Fruit Flavor. Nat. Genet. 2019, 51, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Z.; Bao, Z.; Li, H.; Lyu, Y.; Zan, Y.; Wu, Y.; Cheng, L.; Fang, Y.; Wu, K.; et al. Graph Pangenome Captures Missing Heritability and Empowers Tomato Breeding. Nature 2022, 606, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Barchi, L.; Rabanus-Wallace, M.T.; Prohens, J.; Toppino, L.; Padmarasu, S.; Portis, E.; Rotino, G.L.; Stein, N.; Lanteri, S.; Giuliano, G. Improved Genome Assembly and Pan-genome Provide Key Insights into Eggplant Domestication and Breeding. Plant J. 2021, 107, 579–596. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, J.; Wang, W.; Hu, T.; Hu, H.; Bao, C. A High-Quality Chromosome-Level Genome Assembly Reveals Genetics for Important Traits in Eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [CrossRef]
- Li, D.; Qian, J.; Li, W.; Yu, N.; Gan, G.; Jiang, Y.; Li, W.; Liang, X.; Chen, R.; Mo, Y.; et al. A High-Quality Genome Assembly of the Eggplant Provides Insights into the Molecular Basis of Disease Resistance and Chlorogenic Acid Synthesis. Mol. Ecol. Resour. 2021, 21, 1274–1286. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Martina, M.; Tikunov, Y.; Portis, E.; Bovy, A.G. The Genetic Basis of Tomato Aroma. Genes 2021, 12, 226. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Luo, S.; Li, Q.; Li, N.; Du, W.; Yu, P.; Wang, X.; Zhang, W.; Xuan, S.; Zhou, X.; et al. Phenotypic Characterization and Differential Gene Expression Analysis Reveal That Dwarf Mutant Dwf Dwarfism Is Associated with Gibberellin in Eggplant. Horticulturae 2021, 7, 114. [Google Scholar] [CrossRef]
- Lu, Y.; Luo, S.; Li, N.; Li, Q.; Du, W.; Zhang, W.; Yu, P.; Xuan, S.; Wang, Y.; Zhao, J.; et al. Candidate Gene, SmCPR1, Encoding CPR1 Related to Plant Height of the Eggplant Dwarf Mutant Dwf. Horticulturae 2021, 7, 196. [Google Scholar] [CrossRef]
- Du, W.; Lu, Y.; Luo, S.; Yu, P.; Shen, J.; Wang, X.; Xuan, S.; Wang, Y.; Zhao, J.; Li, N.; et al. Genome-Wide Transcriptome Analysis Reveals That Upregulated Expression of Aux/IAA Genes Is Associated with Defective Leaf Growth of the Slf Mutant in Eggplant. Agronomy 2022, 12, 2647. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, A.; Li, W.; Jiang, Y.; Song, S.; Li, Y.; Chen, R. Comparative Proteomic Analysis of Eggplant (Solanum melongena L.) Heterostylous Pistil Development. PLoS ONE 2017, 12, e0179018. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, H.; Xu, T.; Shi, T.; Li, Z.; Hou, W. Comparative Transcriptome Analysis Reveals Key Genes and Pathways Involved in Prickle Development in Eggplant. Genes 2021, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Zhang, B.; Chen, H.; Lu, L.; Duan, M.; Zhou, J.; Cui, Y.; Li, D. Identification of Quantitative Trait Loci Controlling the Development of Prickles in Eggplant by Genome Re-Sequencing Analysis. Front. Plant Sci. 2021, 12, 731079. [Google Scholar] [CrossRef]
- Miyatake, K.; Saito, T.; Nunome, T.; Yamaguchi, H.; Negoro, S.; Ohyama, A.; Wu, J.; Katayose, Y.; Fukuoka, H. Fine Mapping of a Major Locus Representing the Lack of Prickles in Eggplant Revealed the Availability of a 0.5-Kb Insertion/Deletion for Marker-Assisted Selection. Breed. Sci. 2020, 70, 438–448. [Google Scholar] [CrossRef]
- Oppenheimer, D.G.; Herman, P.L.; Sivakumaran, S.; Esch, J.; Marks, M.D. A Myb Gene Required for Leaf Trichome Differentiation in Arabidopsis Is Expressed in Stipules. Cell 1991, 67, 483–493. [Google Scholar] [CrossRef]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 Locus, Which Regulates Trichome Differentiation and Anthocyanin Biosynthesis in Arabidopsis, Encodes a WD40 Repeat Protein. Plant Cell 1999, 11, 1337–1349. [Google Scholar] [CrossRef]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 Encodes a BHLH Protein That Regulates Trichome Development in Arabidopsis Through Interaction With GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar] [CrossRef]
- Zhang, X.; Oppenheimer, D.G. A Simple and Efficient Method for Isolating Trichomes for Downstream Analyses. Plant Cell Physiol. 2004, 45, 221–224. [Google Scholar] [CrossRef]
- Chang, Z.; Chen, Z.; Wang, N.; Xie, G.; Lu, J.; Yan, W.; Zhou, J.; Tang, X.; Deng, X.W. Construction of a Male Sterility System for Hybrid Rice Breeding and Seed Production Using a Nuclear Male Sterility Gene. Proc. Natl. Acad. Sci. USA 2016, 113, 14145–14150. [Google Scholar] [CrossRef]
- Picarella, M.E.; Mazzucato, A. The Occurrence of Seedlessness in Higher Plants; Insights on Roles and Mechanisms of Parthenocarpy. Front. Plant Sci. 2019, 9, 1997. [Google Scholar] [CrossRef]
- Schnable, P.S.; Wise, R.P. The Molecular Basis of Cytoplasmic Male Sterility and Fertility Restoration. Trends Plant Sci. 1998, 3, 175–180. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.-G. Male Sterility and Fertility Restoration in Crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Zhou, K.; Liu, Y.; Deng, L.; Zhang, X.; Lin, L.; Zhou, M.; Zhao, W.; Wen, C.; Xing, J.; et al. A Biotechnology-Based Male-Sterility System for Hybrid Seed Production in Tomato. Plant J. 2020, 102, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Milner, M.J.; Craze, M.; Bowden, S.; Bates, R.; Wallington, E.J.; Keeling, A. Identification of Genes Involved in Male Sterility in Wheat (Triticum aestivum L.) Which Could Be Used in a Genic Hybrid Breeding System. Plant Direct 2020, 4, e00201. [Google Scholar] [CrossRef] [PubMed]
- Melonek, J.; Duarte, J.; Martin, J.; Beuf, L.; Murigneux, A.; Varenne, P.; Comadran, J.; Specel, S.; Levadoux, S.; Bernath-Levin, K.; et al. The Genetic Basis of Cytoplasmic Male Sterility and Fertility Restoration in Wheat. Nat. Commun. 2021, 12, 1036. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Yang, X.; Zhao, N.; Hu, Z.; Mackenzie, S.A.; Zhang, M.; Yang, J. Exploiting Sterility and Fertility Variation in Cytoplasmic Male Sterile Vegetable Crops. Hortic. Res. 2022, 9, uhab039. [Google Scholar] [CrossRef] [PubMed]
- Phatak, S.C.; Liu, J.; Jaworski, C.A.; Sultanbawa, A.F. Functional Male Sterility in Eggplant: Inheritance and Linkage to the Purple Fruit Color Gene. J. Hered. 1991, 82, 81–83. [Google Scholar] [CrossRef]
- Khan, M.M.R.; Isshiki, S. Development of the Male-Sterile Line of Eggplant Utilizing the Cytoplasm of Solanum aethiopicum L. Aculeatum Group. J. Jpn. Soc. Hortic. Sci. 2010, 79, 348–353. [Google Scholar] [CrossRef]
- Khan, M.M.R.; Isshiki, S. Development of a Cytoplasmic Male-Sterile Line of Eggplant (Solanum melongena L.) with the Cytoplasm of Solanum anguivi. Plant Breed. 2011, 130, 256–260. [Google Scholar] [CrossRef]
- Hasnunnahar, M.; Khan, M.M.R.; Isshiki, S. Pollen and Seed Fertility of Three Functional Male-Sterile Lines of Eggplant with the Wild Solanum Cytoplasms. Sci. Hortic. 2012, 139, 58–61. [Google Scholar] [CrossRef]
- Cao, B.; Huang, Z.; Chen, G.; Lei, J. Restoring Pollen Fertility in Transgenic Male-Sterile Eggplant by Cre/Loxp-Mediated Site-Specific Recombination System. Genet. Mol. Biol. 2010, 33, 298–307. [Google Scholar] [CrossRef]
- Toppino, L.; Kooiker, M.; Lindner, M.; Dreni, L.; Rotino, G.L.; Kater, M.M. Reversible Male Sterility in Eggplant (Solanum melongena L.) by Artificial MicroRNA-Mediated Silencing of General Transcription Factor Genes. Plant Biotechnol. J. 2011, 9, 684–692. [Google Scholar] [CrossRef]
- Yang, Y.; Bao, S.; Zhou, X.; Liu, J.; Zhuang, Y. The Key Genes and Pathways Related to Male Sterility of Eggplant Revealed by Comparative Transcriptome Analysis. BMC Plant Biol. 2018, 18, 209. [Google Scholar] [CrossRef]
- Li, B.; Chen, X.; Wu, Y.; Gu, A.; Zhang, J.; Luo, S.; Gao, X.; Zhao, J.; Pan, X.; Shen, S. Gene Characterization and Molecular Pathway Analysis of Reverse Thermosensitive Genic Male Sterility in Eggplant (Solanum melongena L.). Hortic. Res. 2019, 6, 118. [Google Scholar] [CrossRef]
- Wang, Z.; Yuan, C.; Zhang, S.; Tian, S.; Tang, Q.; Wei, D.; Niu, Y. Screening and Interaction Analysis Identify Genes Related to Anther Dehiscence in Solanum melongena L. Front. Plant Sci. 2021, 12, 648193. [Google Scholar] [CrossRef]
- Yuan, C.; Zhang, S.; Hu, R.; Wei, D.; Tang, Q.; Wang, Y.; Tian, S.; Niu, Y.; Wang, Z. Comparative Transcriptome Analysis Provides Insight into the Molecular Mechanisms of Anther Dehiscence in Eggplant (Solanum melongena L.). Genomics 2021, 113, 497–506. [Google Scholar] [CrossRef]
- Zhang, S.-W.; Yuan, C.; An, L.-Y.; Niu, Y.; Song, M.; Tang, Q.-L.; Wei, D.-Y.; Tian, S.-B.; Wang, Y.-Q.; Yang, Y.; et al. SmCOI1 Affects Anther Dehiscence in a Male-Sterile Solanum melongena Line. Plant Biotechnol. 2020, 37, 1–8. [Google Scholar] [CrossRef]
- Wang, Z.; Yuan, C.; Ding, Z.; Hu, R.; Niu, Y.; Tang, Q.; Wei, D.; Song, M.; Wang, Y.; Tian, S. Analysis of differential genes and metabolic pathway related to functional male sterility in eggplant. Sheng Wu Gong Cheng Xue Bao 2021, 37, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Acciarri, N.; Restaino, F.; Vitelli, G.; Perrone, D.; Zottini, M.; Pandolfini, T.; Spena, A.; Rotino, G.L. Genetically Modified Parthenocarpic Eggplants: Improved Fruit Productivity under Both Greenhouse and Open Field Cultivation. BMC Biotechnol. 2002, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Aubert, S.; Daunay, M.C.; Pochard, E. Steroidic Saponins from Egg-Plant (Solanum melongena L.) II—Effects of Cultural Conditions, Genotypes and Parthenocarpy. Agronomie 1989, 9, 641–651. [Google Scholar] [CrossRef]
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 607161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gu, X.; Shao, J.; Hu, Z.; Yang, W.; Wang, L.; Su, H.; Zhu, L. Auxin Metabolism Is Involved in Fruit Set and Early Fruit Development in the Parthenocarpic Tomato “R35-P”. Front. Plant Sci. 2021, 12, 671713. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, K.; Saito, T.; Negoro, S.; Yamaguchi, H.; Nunome, T.; Ohyama, A.; Fukuoka, H. Development of Selective Markers Linked to a Major QTL for Parthenocarpy in Eggplant (Solanum melongena L.). Theor. Appl. Genet. 2012, 124, 1403–1413. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, M.; Tan, J.; Huang, S.; Wang, C.; Zhang, H.; Tan, T. Comparative Transcriptome Analysis Provides Insights into Molecular Mechanisms for Parthenocarpic Fruit Development in Eggplant (Solanum melongena L.). PLoS ONE 2017, 12, e0179491. [Google Scholar] [CrossRef]
- Du, L.; Bao, C.; Hu, T.; Zhu, Q.; Hu, H.; He, Q.; Mao, W. SmARF8, a Transcription Factor Involved in Parthenocarpy in Eggplant. Mol. Genet. Genom. 2016, 291, 93–105. [Google Scholar] [CrossRef]
- Matsuo, S.; Miyatake, K.; Endo, M.; Urashimo, S.; Kawanishi, T.; Negoro, S.; Shimakoshi, S.; Fukuoka, H. Loss of Function of the Pad-1 Aminotransferase Gene, Which Is Involved in Auxin Homeostasis, Induces Parthenocarpy in Solanaceae Plants. Proc. Natl. Acad. Sci. USA 2020, 117, 12784–12790. [Google Scholar] [CrossRef]
- Avana-Tientcheu, M.L.; Tiambo, C.K. Breeding and Productivity in Ending Hunger and Achieving Food Security and Nutrition. In Zero Hunger; Leal Filho, W., Azul, A.M., Brandli, L., Özuyar, P.G., Wall, T., Eds.; Encyclopedia of the UN Sustainable Development Goals; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–18. ISBN 978-3-319-69626-3. [Google Scholar]
- Gaikwad, K.B.; Rani, S.; Kumar, M.; Gupta, V.; Babu, P.H.; Bainsla, N.K.; Yadav, R. Enhancing the Nutritional Quality of Major Food Crops Through Conventional and Genomics-Assisted Breeding. Front. Nutr. 2020, 7, 533453. [Google Scholar] [CrossRef] [PubMed]
- Docimo, T.; Francese, G.; Ruggiero, A.; Batelli, G.; De Palma, M.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Mennella, G.; Tucci, M. Phenylpropanoids Accumulation in Eggplant Fruit: Characterization of Biosynthetic Genes and Regulation by a MYB Transcription Factor. Front. Plant Sci. 2016, 6, 1233. [Google Scholar] [CrossRef] [PubMed]
- Lelario, F.; De Maria, S.; Rivelli, A.R.; Russo, D.; Milella, L.; Bufo, S.A.; Scrano, L. A Complete Survey of Glycoalkaloids Using LC-FTICR-MS and IRMPD in a Commercial Variety and a Local Landrace of Eggplant (Solanum melongena L.) and Their Anticholinesterase and Antioxidant Activities. Toxins 2019, 11, 230. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Qian, Z.; Zhang, J.; Yang, J.; Wu, M.; Barchi, L.; Zhao, H.; Sun, H.; Cui, Y.; Wen, C. Impact of Fruit Shape Selection on Genetic Structure and Diversity Uncovered from Genome-Wide Perfect SNPs Genotyping in Eggplant. Mol. Breed. 2019, 39, 140. [Google Scholar] [CrossRef]
- Shi, S.; Li, D.; Li, S.; Wang, Y.; Tang, X.; Liu, Y.; Ge, H.; Chen, H. Comparative Transcriptomic Analysis of Early Fruit Development in Eggplant (Solanum Melongena L.) and Functional Characterization of SmOVATE5. Plant Cell Rep. 2023, 1–16. [Google Scholar] [CrossRef]
- Rodríguez, G.R.; Muños, S.; Anderson, C.; Sim, S.-C.; Michel, A.; Causse, M.; Gardener, B.B.M.; Francis, D.; van der Knaap, E. Distribution of SUN, OVATE, LC, and FAS in the Tomato Germplasm and the Relationship to Fruit Shape Diversity1[C][W][OA]. Plant Physiol. 2011, 156, 275–285. [Google Scholar] [CrossRef]
- Acquadro, A.; Barchi, L.; Portis, E.; Nourdine, M.; Carli, C.; Monge, S.; Valentino, D.; Lanteri, S. Whole Genome Resequencing of Four Italian Sweet Pepper Landraces Provides Insights on Sequence Variation in Genes of Agronomic Value. Sci. Rep. 2020, 10, 9189. [Google Scholar] [CrossRef]
- Borovsky, Y.; Raz, A.; Doron-Faigenboim, A.; Zemach, H.; Karavani, E.; Paran, I. Pepper Fruit Elongation Is Controlled by Capsicum annuum Ovate Family Protein 20. Front. Plant Sci. 2022, 12, 815589. [Google Scholar] [CrossRef]
- Cong, B.; Barrero, L.S.; Tanksley, S.D. Regulatory Change in YABBY-like Transcription Factor Led to Evolution of Extreme Fruit Size during Tomato Domestication. Nat. Genet. 2008, 40, 800–804. [Google Scholar] [CrossRef]
- Xiao, H.; Jiang, N.; Schaffner, E.; Stockinger, E.J.; van der Knaap, E. A Retrotransposon-Mediated Gene Duplication Underlies Morphological Variation of Tomato Fruit. Science 2008, 319, 1527–1530. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Zhang, L.; Sapkota, M.; Ramos, A.; Razifard, H.; Caicedo, A.L.; van der Knaap, E. Unraveling the Genetics of Tomato Fruit Weight during Crop Domestication and Diversification. Theor. Appl. Genet. 2021, 134, 3363–3378. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Huang, Z.; Chakrabarti, M.; Illa-Berenguer, E.; Liu, X.; Wang, Y.; Ramos, A.; van der Knaap, E. Fruit Weight Is Controlled by Cell Size Regulator Encoding a Novel Protein That Is Expressed in Maturing Tomato Fruits. PLoS Genet. 2017, 13, e1006930. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Feng, Q.; Snouffer, A.; Zhang, B.; Rodríguez, G.R.; van der Knaap, E. Increasing Fruit Weight by Editing a Cis-Regulatory Element in Tomato KLUH Promoter Using CRISPR/Cas9. Front. Plant Sci. 2022, 13, 879642. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Zhang, N.; Sauvage, C.; Muños, S.; Blanca, J.; Cañizares, J.; Diez, M.J.; Schneider, R.; Mazourek, M.; McClead, J.; et al. A Cytochrome P450 Regulates a Domestication Trait in Cultivated Tomato. Proc. Natl. Acad. Sci. USA 2013, 110, 17125–17130. [Google Scholar] [CrossRef]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major Impacts of Widespread Structural Variation on Gene Expression and Crop Improvement in Tomato. Cell 2020, 182, 145–161.e23. [Google Scholar] [CrossRef]
- Arrones, A.; Mangino, G.; Alonso, D.; Plazas, M.; Prohens, J.; Portis, E.; Barchi, L.; Giuliano, G.; Vilanova, S.; Gramazio, P. Mutations in the SmAPRR2 Transcription Factor Suppressing Chlorophyll Pigmentation in the Eggplant Fruit Peel Are Key Drivers of a Diversified Colour Palette. Front. Plant Sci. 2022, 13, 1025951. [Google Scholar] [CrossRef]
- Powell, A.L.T.; Nguyen, C.V.; Hill, T.; Cheng, K.L.; Figueroa-Balderas, R.; Aktas, H.; Ashrafi, H.; Pons, C.; Fernández-Muñoz, R.; Vicente, A.; et al. Uniform Ripening Encodes a Golden 2-like Transcription Factor Regulating Tomato Fruit Chloroplast Development. Science 2012, 336, 1711–1715. [Google Scholar] [CrossRef]
- Nguyen, C.V.; Vrebalov, J.T.; Gapper, N.E.; Zheng, Y.; Zhong, S.; Fei, Z.; Giovannoni, J.J. Tomato GOLDEN2-LIKE Transcription Factors Reveal Molecular Gradients That Function during Fruit Development and Ripening. Plant Cell 2014, 26, 585–601. [Google Scholar] [CrossRef]
- Pan, Y.; Bradley, G.; Pyke, K.; Ball, G.; Lu, C.; Fray, R.; Marshall, A.; Jayasuta, S.; Baxter, C.; van Wijk, R.; et al. Network Inference Analysis Identifies an APRR2-Like Gene Linked to Pigment Accumulation in Tomato and Pepper Fruits. Plant Physiol. 2013, 161, 1476–1485. [Google Scholar] [CrossRef]
- Liu, D.; Li, W.; Cheng, J. The Novel Protein DELAYED PALE-GREENING1 Is Required for Early Chloroplast Biogenesis in Arabidopsis Thaliana. Sci. Rep. 2016, 6, 25742. [Google Scholar] [CrossRef]
- Oren, E.; Tzuri, G.; Vexler, L.; Dafna, A.; Meir, A.; Faigenboim, A.; Kenigswald, M.; Portnoy, V.; Schaffer, A.A.; Levi, A.; et al. The Multi-Allelic APRR2 Gene Is Associated with Fruit Pigment Accumulation in Melon and Watermelon. J. Exp. Bot. 2019, 70, 3781. [Google Scholar] [CrossRef]
- Cominelli, E.; Sala, T.; Calvi, D.; Gusmaroli, G.; Tonelli, C. Over-Expression of the Arabidopsis AtMYB41 Gene Alters Cell Expansion and Leaf Surface Permeability. Plant J. 2007, 53, 53–64. [Google Scholar] [CrossRef]
- Raffaele, S.; Vailleau, F.; Léger, A.; Joubès, J.; Miersch, O.; Huard, C.; Blée, E.; Mongrand, S.; Domergue, F.; Roby, D. A MYB Transcription Factor Regulates Very-Long-Chain Fatty Acid Biosynthesis for Activation of the Hypersensitive Cell Death Response in Arabidopsis. Plant Cell 2008, 20, 752–767. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Li, S.; He, S.; Waßmann, F.; Yu, C.; Qin, G.; Schreiber, L.; Qu, L.-J.; Gu, H. CFL1, a WW Domain Protein, Regulates Cuticle Development by Modulating the Function of HDG1, a Class IV Homeodomain Transcription Factor, in Rice and Arabidopsis. Plant Cell 2011, 23, 3392–3411. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-L.; Zhang, C.-L.; Wang, G.-L.; Wang, Y.-X.; Qi, C.-H.; Zhao, Q.; You, C.-X.; Li, Y.-Y.; Hao, Y.-J. The R2R3 MYB Transcription Factor MdMYB30 Modulates Plant Resistance against Pathogens by Regulating Cuticular Wax Biosynthesis. BMC Plant Biol. 2019, 19, 362. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Xie, Q.; Yang, Q.; Sun, P.; Gao, S.; Li, H.; Zhang, J.; Wang, T.; Ye, Z.; Yang, C. WOOLLY, Interacting with MYB Transcription Factor MYB31, Regulates Cuticular Wax Biosynthesis by Modulating CER6 Expression in Tomato. Plant J. 2020, 103, 323–337. [Google Scholar] [CrossRef]
- Zhang, A.; Huang, Q.; Li, J.; Zhu, W.; Liu, X.; Wu, X.; Zha, D. Comparative Transcriptome Analysis Reveals Gene Expression Differences in Eggplant (Solanum melongena L.) Fruits with Different Brightness. Foods 2022, 11, 2506. [Google Scholar] [CrossRef]
- Todd, J.; Post-Beittenmiller, D.; Jaworski, J.G. KCS1 Encodes a Fatty Acid Elongase 3-Ketoacyl-CoA Synthase Affecting Wax Biosynthesis in Arabidopsis Thaliana. Plant J. 1999, 17, 119–130. [Google Scholar] [CrossRef]
- Yang, T.; Li, Y.; Liu, Y.; He, L.; Liu, A.; Wen, J.; Mysore, K.S.; Tadege, M.; Chen, J. The 3-Ketoacyl-CoA Synthase WFL Is Involved in Lateral Organ Development and Cuticular Wax Synthesis in Medicago Truncatula. Plant Mol. Biol. 2021, 105, 193–204. [Google Scholar] [CrossRef]
- Lee, S.-B.; Jung, S.-J.; Go, Y.-S.; Kim, H.-U.; Kim, J.-K.; Cho, H.-J.; Park, O.K.; Suh, M.-C. Two Arabidopsis 3-Ketoacyl CoA Synthase Genes, KCS20 and KCS2/DAISY, Are Functionally Redundant in Cuticular Wax and Root Suberin Biosynthesis, but Differentially Controlled by Osmotic Stress. Plant J. 2009, 60, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jung, J.H.; Lee, S.B.; Go, Y.S.; Kim, H.J.; Cahoon, R.; Markham, J.E.; Cahoon, E.B.; Suh, M.C. Arabidopsis 3-Ketoacyl-Coenzyme A Synthase9 Is Involved in the Synthesis of Tetracosanoic Acids as Precursors of Cuticular Waxes, Suberins, Sphingolipids, and Phospholipids1[W]. Plant Physiol. 2013, 162, 567–580. [Google Scholar] [CrossRef]
- Huang, H.; Ayaz, A.; Zheng, M.; Yang, X.; Zaman, W.; Zhao, H.; Lü, S. ArabidopsisKCS5 and KCS6 Play Redundant Roles in Wax Synthesis. Int. J. Mol. Sci. 2022, 23, 4450. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Mumtaz, A.; Gonzales-Vigil, E. Evolution and Molecular Basis of Substrate Specificity in a 3-Ketoacyl-CoA Synthase Gene Cluster from Populus Trichocarpa. J. Biol. Chem. 2022, 298, 102496. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Tang, H.; Wei, X.; Zhao, Y.; Wang, Z.; Su, H.; Niu, L.; Yuan, Y.; Zhang, X. BrWAX3, Encoding a β-Ketoacyl-CoA Synthase, Plays an Essential Role in Cuticular Wax Biosynthesis in Chinese Cabbage. Int. J. Mol. Sci. 2022, 23, 10938. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of Plant Pigments: Anthocyanins, Betalains and Carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Jiao, Y.; Yang, Z.; Jiang, Y.; Zhai, W. Study on Chemical Constituents and Antioxidant Activity of Anthocyanins from Purple Sweet Potato (Ipomoea batatas L.). Int. J. Food Eng. 2012, 54, 2604–2614. [Google Scholar] [CrossRef]
- Zhao, Z.; Yan, H.; Zheng, R.; Khan, M.S.; Fu, X.; Tao, Z.; Zhang, Z. Anthocyanins Characterization and Antioxidant Activities of Sugarcane (Saccharum officinarum L.) Rind Extracts. Ind. Crops Prod. 2018, 113, 38–45. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- van Eck, H.J.; Jacobs, J.M.E.; van den Berg, P.M.M.M.; Stiekema, W.J.; Jacobsen, E. The Inheritance of Anthocyanin Pigmentation in Potato (Solanum tuberosum L.) and Mapping of Tuber Skin Colour Loci Using RFLPs. Heredity 1994, 73, 410–421. [Google Scholar] [CrossRef]
- Chaim, A.B.; Borovsky, Y.; De Jong, W.; Paran, I. Linkage of the A Locus for the Presence of Anthocyanin and Fs10.1, a Major Fruit-Shape QTL in Pepper. Theor. Appl. Genet. 2003, 106, 889–894. [Google Scholar] [CrossRef]
- Borovsky, Y.; Oren-Shamir, M.; Ovadia, R.; De Jong, W.; Paran, I. The A Locus That Controls Anthocyanin Accumulation in Pepper Encodes a MYB Transcription Factor Homologous to Anthocyanin2 of Petunia. Theor. Appl. Genet. 2004, 109, 23–29. [Google Scholar] [CrossRef]
- Bovy, A.; Schijlen, E.; Hall, R.D. Metabolic Engineering of Flavonoids in Tomato (Solanum lycopersicum): The Potential for Metabolomics. Metabolomics 2007, 3, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Gonzali, S.; Mazzucato, A.; Perata, P. Purple as a Tomato: Towards High Anthocyanin Tomatoes. Trends Plant Sci. 2009, 14, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Stommel, J.R.; Dumm, J.M. Coordinated Regulation of Biosynthetic and Regulatory Genes Coincides with Anthocyanin Accumulation in Developing Eggplant Fruit. J. Am. Soc. Hortic. Sci. 2015, 140, 129–135. [Google Scholar] [CrossRef]
- Condurache (Lazăr), N.-N.; Croitoru, C.; Enachi, E.; Bahrim, G.-E.; Stănciuc, N.; Râpeanu, G. Eggplant Peels as a Valuable Source of Anthocyanins: Extraction, Thermal Stability and Biological Activities. Plants 2021, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Florio, F.E.; Gattolin, S.; Toppino, L.; Bassolino, L.; Fibiani, M.; Lo Scalzo, R.; Rotino, G.L. A SmelAAT Acyltransferase Variant Causes a Major Difference in Eggplant (Solanum melongena L.) Peel Anthocyanin Composition. Int. J. Mol. Sci. 2021, 22, 9174. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB Transcription Factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yu, W.; Xu, J.; Lu, X.; Liu, Y. Anthocyanin Biosynthesis Induced by MYB Transcription Factors in Plants. Int. J. Mol. Sci. 2022, 23, 11701. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Luo, S.; Ma, W.; Li, N.; Zhang, W.; Tikunov, Y.; Xuan, S.; Zhao, J.; Wang, Y.; et al. Discovery of a DFR Gene That Controls Anthocyanin Accumulation in the Spiny Solanum Group: Roles of a Natural Promoter Variant and Alternative Splicing. Plant J. 2022, 111, 1096–1109. [Google Scholar] [CrossRef]
- Li, J.; He, Y.-J.; Zhou, L.; Liu, Y.; Jiang, M.; Ren, L.; Chen, H. Transcriptome Profiling of Genes Related to Light-Induced Anthocyanin Biosynthesis in Eggplant (Solanum melongena L.) before Purple Color Becomes Evident. BMC Genom. 2018, 19, 201. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; He, Y.; Li, J.; Liu, Y.; Chen, H. CBFs Function in Anthocyanin Biosynthesis by Interacting with MYB113 in Eggplant (Solanum melongena L.). Plant Cell Physiol. 2020, 61, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, S.; Yang, Y.; Liu, J.; Zhuang, Y. Integrated Metabolome and Transcriptome Analysis Reveals a Regulatory Network of Fruit Peel Pigmentation in Eggplant (Solanum melongena L.). Int. J. Mol. Sci. 2022, 23, 13475. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, H.; Zhou, L.; Liu, Y.; Chen, H. Comparative Transcription Analysis of Photosensitive and Non-Photosensitive Eggplants to Identify Genes Involved in Dark Regulated Anthocyanin Synthesis. BMC Genom. 2019, 20, 678. [Google Scholar] [CrossRef]
- He, Y.; Wang, Z.; Ge, H.; Liu, Y.; Chen, H. Weighted Gene Co-Expression Network Analysis Identifies Genes Related to Anthocyanin Biosynthesis and Functional Verification of Hub Gene SmWRKY44. Plant Sci. 2021, 309, 110935. [Google Scholar] [CrossRef]
- Andrea, M.; Francesco, E.F.; Sergio, I.; Alessandra, G.; Maria, A.M.; Cinzia, C.; Lorenzo, B.; Arianna, M.; Cecilia, C.; Patrizia, R.; et al. Identification of a New R3 MYB Type Repressor and Functional Characterization of the Members of the MBW Transcriptional Complex Involved in Anthocyanin Biosynthesis in Eggplant (S. melongena L.). PLoS ONE 2020, 15, e0232986. [Google Scholar] [CrossRef]
- Li, L.; Li, S.; Ge, H.; Shi, S.; Li, D.; Liu, Y.; Chen, H. A Light-Responsive Transcription Factor SmMYB35 Enhances Anthocyanin Biosynthesis in Eggplant (Solanum melongena L.). Planta 2021, 255, 12. [Google Scholar] [CrossRef]
- Guo, J.; Wang, M.-H. Ultraviolet A-Specific Induction of Anthocyanin Biosynthesis and PAL Expression in Tomato (Solanum lycopersicum L.). Plant Growth Regul. 2010, 62, 1–8. [Google Scholar] [CrossRef]
- Jiang, M.; Ren, L.; Lian, H.; Liu, Y.; Chen, H. Novel Insight into the Mechanism Underlying Light-Controlled Anthocyanin Accumulation in Eggplant (Solanum melongena L.). Plant Sci. 2016, 249, 46–58. [Google Scholar] [CrossRef]
- Qiu, Z.; Wang, X.; Gao, J.; Guo, Y.; Huang, Z.; Du, Y. The Tomato Hoffman’s Anthocyaninless Gene Encodes a BHLH Transcription Factor Involved in Anthocyanin Biosynthesis That Is Developmentally Regulated and Induced by Low Temperatures. PLoS ONE 2016, 11, e0151067. [Google Scholar] [CrossRef]
- Maier, A.; Hoecker, U. COP1/SPA Ubiquitin Ligase Complexes Repress Anthocyanin Accumulation under Low Light and High Light Conditions. Plant Signal. Behav. 2014, 10, e970440. [Google Scholar] [CrossRef]
- Meng, L.-S.; Liu, A. Light Signaling Induces Anthocyanin Biosynthesis via AN3 Mediated COP1 Expression. Plant Signal. Behav. 2015, 10, e1001223. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, S.; Dong, Y.; Zhang, X.; Li, D.; Liu, Y.; Chen, H. Fine Mapping and Characterization of the Dominant Gene SmFTSH10 Conferring Non-Photosensitivity in Eggplant (Solanum melongena L.). Theor. Appl. Genet. 2022, 135, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ren, L.; Gao, Z.; Jiang, M.; Liu, Y.; Zhou, L.; He, Y.; Chen, H. Combined Transcriptomic and Proteomic Analysis Constructs a New Model for Light-Induced Anthocyanin Biosynthesis in Eggplant (Solanum melongena L.). Plant Cell Environ. 2017, 40, 3069–3087. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Phukan, T.; Kabyashree, K.; Singh, R.; Sharma, P.L.; Singh, N.; Barman, A.; Jena, B.R.; Ray, S.K. Ralstonia Solanacearum Virulence in Eggplant Seedlings by the Leaf-Clip Inoculation. Phytopathol. Res. 2019, 1, 23. [Google Scholar] [CrossRef]
- Xiao, X.; Lin, W.; Feng, E.; Wu, C.; Ou, X. Genome-Wide Identification of Binding Sites for SmTCP7a Transcription Factors of Eggplant during Bacterial Wilt Resistance by ChIP-Seq. Int. J. Mol. Sci. 2022, 23, 6844. [Google Scholar] [CrossRef] [PubMed]
- Khoury, C.K.; Brush, S.; Costich, D.E.; Curry, H.A.; de Haan, S.; Engels, J.M.M.; Guarino, L.; Hoban, S.; Mercer, K.L.; Miller, A.J.; et al. Crop Genetic Erosion: Understanding and Responding to Loss of Crop Diversity. New Phytol. 2022, 233, 84–118. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial Activity and Mode of Action of Ferulic and Gallic Acids against Pathogenic Bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Paniagua, C.; Santiago-Doménech, N.; Kirby, A.R.; Gunning, A.P.; Morris, V.J.; Quesada, M.A.; Matas, A.J.; Mercado, J.A. Structural Changes in Cell Wall Pectins during Strawberry Fruit Development. Plant Physiol. Biochem. 2017, 118, 55–63. [Google Scholar] [CrossRef]
- Li, N.; Zhao, M.; Liu, T.; Dong, L.; Cheng, Q.; Wu, J.; Wang, L.; Chen, X.; Zhang, C.; Lu, W.; et al. A Novel Soybean Dirigent Gene GmDIR22 Contributes to Promotion of Lignan Biosynthesis and Enhances Resistance to Phytophthora Sojae. Front. Plant Sci. 2017, 8, 1185. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Mehra, S.; Chatterjee, S.; Purty, R.S. In Silico Identification and Validation of MiRNA and Their DIR Specific Targets in Oryza Sativa Indica under Abiotic Stress. Non-Coding RNA Res. 2020, 5, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Shen, Y.; Ahmad, B.; Yao, L.; He, T.; Fan, J.; Liu, Y.; Chen, Q.; Wen, Z. Genome-Wide Identification and Expression Analysis of Dirigent Gene Family in Strawberry (Fragaria Vesca) and Functional Characterization of FvDIR13. Sci. Hortic. 2022, 297, 110913. [Google Scholar] [CrossRef]
- Zhang, K.; Xing, W.; Sheng, S.; Yang, D.; Zhen, F.; Jiang, H.; Yan, C.; Jia, L. Genome-Wide Identification and Expression Analysis of Eggplant DIR Gene Family in Response to Biotic and Abiotic Stresses. Horticulturae 2022, 8, 732. [Google Scholar] [CrossRef]
- Peng, J.; Wang, P.; Fang, H.; Zheng, J.; Zhong, C.; Yang, Y.; Yu, W. Weighted Gene Co-Expression Analysis Network-Based Analysis on the Candidate Pathways and Hub Genes in Eggplant Bacterial Wilt-Resistance: A Plant Research Study. Int. J. Mol. Sci. 2021, 22, 13279. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Wang, X.; Wang, D.; Xu, F.; Ding, X.; Zhang, Z.; Bi, D.; Cheng, Y.T.; Chen, S.; Li, X.; et al. Regulation of Cell Death and Innate Immunity by Two Receptor-like Kinases in Arabidopsis. Cell Host Microbe 2009, 6, 34–44. [Google Scholar] [CrossRef]
- Van der Does, D.; Boutrot, F.; Engelsdorf, T.; Rhodes, J.; McKenna, J.F.; Vernhettes, S.; Koevoets, I.; Tintor, N.; Veerabagu, M.; Miedes, E.; et al. The Arabidopsis Leucine-Rich Repeat Receptor Kinase MIK2/LRR-KISS Connects Cell Wall Integrity Sensing, Root Growth and Response to Abiotic and Biotic Stresses. PLoS Genet. 2017, 13, e1006832. [Google Scholar] [CrossRef]
- Xi’ou, X.; Bihao, C.; Guannan, L.; Jianjun, L.; Qinghua, C.; Jin, J.; Yujing, C. Functional Characterization of a Putative Bacterial Wilt Resistance Gene (RE-Bw) in Eggplant. Plant Mol. Biol. Report. 2015, 4, 1058–1073. [Google Scholar] [CrossRef]
- Qiu, Z.; Yan, S.; Xia, B.; Jiang, J.; Yu, B.; Lei, J.; Chen, C.; Chen, L.; Yang, Y.; Wang, Y.; et al. The Eggplant Transcription Factor MYB44 Enhances Resistance to Bacterial Wilt by Activating the Expression of Spermidine Synthase. J. Exp. Bot. 2019, 70, 5343–5354. [Google Scholar] [CrossRef]
- Tassone, M.R.; Bagnaresi, P.; Desiderio, F.; Bassolino, L.; Barchi, L.; Florio, F.E.; Sunseri, F.; Sirangelo, T.M.; Rotino, G.L.; Toppino, L. A Genomic BSAseq Approach for the Characterization of QTLs Underlying Resistance to Fusarium Oxysporum in Eggplant. Cells 2022, 11, 2548. [Google Scholar] [CrossRef]
- Agurto, M.; Schlechter, R.O.; Armijo, G.; Solano, E.; Serrano, C.; Contreras, R.A.; Zúñiga, G.E.; Arce-Johnson, P. RUN1 and REN1 Pyramiding in Grapevine (Vitis Vinifera Cv. Crimson Seedless) Displays an Improved Defense Response Leading to Enhanced Resistance to Powdery Mildew (Erysiphe Necator). Front. Plant Sci. 2017, 8, 758. [Google Scholar] [CrossRef]
- Sujayanand, G.K.; Sharma, R.K.; Shankarganesh, K.; Saha, S.; Tomar, R.S. Crop Diversification for Sustainable Insect Pest Management in Eggplant (Solanales: Solanaceae). Fla. Entomol. 2015, 98, 305–314. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological Control of Nematodes Infecting Eggplant in Egypt. Bull. Natl. Res. Cent. 2021, 45, 6. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Zhang, J.; He, T.; Huang, J.; Zhang, Z.; Wang, Y.; Hafeez, M.; Zhou, S.; Ren, X.; et al. Comprehensive Metabolome and Volatilome Analyses in Eggplant and Tomato Reveal Their Differential Responses to Tuta Absoluta Infestation. Front. Plant Sci. 2021, 12, 757230. [Google Scholar] [CrossRef] [PubMed]
- Loyani, L.K.; Bradshaw, K.; Machuve, D. Segmentation of Tuta Absoluta’s Damage on Tomato Plants: A Computer Vision Approach. Appl. Artif. Intell. 2021, 35, 1107–1127. [Google Scholar] [CrossRef]
- D’Esposito, D.; Manzo, D.; Ricciardi, A.; Garonna, A.P.; De Natale, A.; Frusciante, L.; Pennacchio, F.; Ercolano, M.R. Tomato Transcriptomic Response to Tuta Absoluta Infestation. BMC Plant Biol. 2021, 21, 358. [Google Scholar] [CrossRef] [PubMed]
- El-Remaly, E.; Osman, A.A.; El-Gawad, H.G.A.; Althobaiti, F.; Albogami, S.; Dessoky, E.S.; El-Mogy, M.M. Bio-Management of Root-Knot Nematodes on Cucumber Using Biocidal Effects of Some Brassicaceae Crops. Horticulturae 2022, 8, 699. [Google Scholar] [CrossRef]
- Elkelany, U.S.; El-Mougy, N.S.; Abdel-Kader, M.M. Management of Root-Knot Nematode Meloidogyne Incognita of Eggplant Using Some Growth-Promoting Rhizobacteria and Chitosan under Greenhouse Conditions. Egypt. J. Biol. Pest Control 2020, 30, 134. [Google Scholar] [CrossRef]
- Sikder, M.M.; Vestergård, M.; Kyndt, T.; Kudjordjie, E.N.; Nicolaisen, M. Phytohormones Selectively Affect Plant Parasitic Nematodes Associated with Arabidopsis Roots. New Phytol. 2021, 232, 1272–1285. [Google Scholar] [CrossRef]
- Sato, K.; Uehara, T.; Holbein, J.; Sasaki-Sekimoto, Y.; Gan, P.; Bino, T.; Yamaguchi, K.; Ichihashi, Y.; Maki, N.; Shigenobu, S.; et al. Transcriptomic Analysis of Resistant and Susceptible Responses in a New Model Root-Knot Nematode Infection System Using Solanum Torvum and Meloidogyne Arenaria. Front. Plant Sci. 2021, 12, 680151. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, H.; Tan, J.; Huang, S.; Chen, X.; Jiang, D.; Xiao, X. Transcriptome Analysis of Eggplant Root in Response to Root-Knot Nematode Infection. Pathogens 2021, 10, 470. [Google Scholar] [CrossRef]
- Song, L.-X.; Xu, X.-C.; Wang, F.-N.; Wang, Y.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Zhou, J.; Yu, J.-Q. Brassinosteroids act as a positive regulator for resistance against root-knot nematode involving RESPIRATORY BURST OXIDASE HOMOLOG-dependent activation of MAPKs in tomato. Plant Cell Environ. 2018, 41, 1113–1125. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, J.; Bao, S.; Yang, Y.; Zhuang, Y. Molecular Cloning and Characterization of a Wild Eggplant Solanum Aculeatissimum NBS-LRR Gene, Involved in Plant Resistance to Meloidogyne Incognita. Int. J. Mol. Sci. 2018, 19, 583. [Google Scholar] [CrossRef]
- Savvas, D.; Lenz, F. Effects of NaCl or Nutrient-Induced Salinity on Growth, Yield, and Composition of Eggplants Grown in Rockwool. Sci. Hortic. 2000, 84, 37–47. [Google Scholar] [CrossRef]
- Daunay, M.C.; Hazra, P. Eggplant. In Handbook of Vegetables; Peter, K.V., Hazra, P., Eds.; Studium Press: Houston, TX, USA, 2012; pp. 257–322. [Google Scholar]
- Wu, X.; He, J.; Chen, J.; Yang, S.; Zha, D. Alleviation of Exogenous 6-Benzyladenine on Two Genotypes of Eggplant (Solanum melongena Mill.) Growth under Salt Stress. Protoplasma 2014, 251, 169–176. [Google Scholar] [CrossRef]
- Díaz-Pérez, J.C.; Eaton, T.E. Eggplant (Solanum melongena L.) Plant Growth and Fruit Yield as Affected by Drip Irrigation Rate. HortScience 2015, 50, 1709–1714. [Google Scholar] [CrossRef]
- Plazas, M.; Nguyen, H.T.; González-Orenga, S.; Fita, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Comparative Analysis of the Responses to Water Stress in Eggplant (Solanum melongena) Cultivars. Plant Physiol. Biochem. 2019, 143, 72–82. [Google Scholar] [CrossRef]
- Toppino, L.; Barchi, L.; Rotino, G.L. Next Generation Breeding for Abiotic Stress Resistance in Eggplant. In Genomic Designing for Abiotic Stress Resistant Vegetable Crops; Kole, C., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 115–151. ISBN 978-3-031-03964-5. [Google Scholar]
- Fu, Q.S.; Yang, R.C.; Wang, H.S.; Zhao, B.; Zhou, C.L.; Ren, S.X.; Guo, Y.-D. Leaf Morphological and Ultrastructural Performance of Eggplant (Solanum melongena L.) in Response to Water Stress. Photosynthetica 2013, 51, 109–114. [Google Scholar] [CrossRef]
- Ramachandra Reddy, A.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Amiri Rodan, M.; Hassandokht, M.R.; Sadeghzadeh-Ahari, D.; Mousavi, A. Mitigation of Drought Stress in Eggplant by Date Straw and Plastic Mulches. J. Saudi Soc. Agric. Sci. 2020, 19, 492–498. [Google Scholar] [CrossRef]
- Tamang, B.G.; Fukao, T. Plant Adaptation to Multiple Stresses during Submergence and Following Desubmergence. Int. J. Mol. Sci. 2015, 16, 30164–30180. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Win, K.T.; Oo, A.Z.; Bellingrath-Kimura, S.D. Influence of Soil Types and Osmotic Pressure on Growth and 137Cs Accumulation in Blackgram (Vigna mungo L.). J. Environ. Radioact. 2017, 169–170, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.R.; Belo, J.; Beeckman, T.; Barros, P.M.; Oliveira, M.M. The Combined Effect of Heat and Osmotic Stress on Suberization of Arabidopsis Roots. Cells 2022, 11, 2341. [Google Scholar] [CrossRef]
- Swindell, W.R. The Association Among Gene Expression Responses to Nine Abiotic Stress Treatments in Arabidopsis Thaliana. Genetics 2006, 174, 1811–1824. [Google Scholar] [CrossRef]
- Biniaz, Y.; Tahmasebi, A.; Afsharifar, A.; Tahmasebi, A.; Poczai, P. Meta-Analysis of Common and Differential Transcriptomic Responses to Biotic and Abiotic Stresses in Arabidopsis Thaliana. Plants 2022, 11, 502. [Google Scholar] [CrossRef]
- Chen, J.; Wang, S.; Wu, F.; Wei, M.; Li, J.; Yang, F. Genome–Wide Identification and Functional Characterization of Auxin Response Factor (ARF) Genes in Eggplant. Int. J. Mol. Sci. 2022, 23, 6219. [Google Scholar] [CrossRef]
- Na, C.; Shuanghua, W.; Jinglong, F.; Bihao, C.; Jianjun, L.; Changming, C.; Jin, J. Overexpression of the Eggplant (Solanum melongena) NAC Family Transcription Factor SmNAC Suppresses Resistance to Bacterial Wilt. Sci. Rep. 2016, 6, 31568. [Google Scholar] [CrossRef]
- Tani, E.; Kizis, D.; Markellou, E.; Papadakis, I.; Tsamadia, D.; Leventis, G.; Makrogianni, D.; Karapanos, I. Cultivar-Dependent Responses of Eggplant (Solanum melongena L.) to Simultaneous Verticillium Dahliae Infection and Drought. Front. Plant Sci. 2018, 9, 1181. [Google Scholar] [CrossRef]
- Moglia, A.; Gianoglio, S.; Acquadro, A.; Valentino, D.; Milani, A.M.; Lanteri, S.; Comino, C. Identification of DNA Methyltransferases and Demethylases in Solanum melongena L., and Their Transcription Dynamics during Fruit Development and after Salt and Drought Stresses. PLoS ONE 2019, 14, e0223581. [Google Scholar] [CrossRef]
- Shen, L.; Zhao, E.; Liu, R.; Yang, X. Transcriptome Analysis of Eggplant under Salt Stress: AP2/ERF Transcription Factor SmERF1 Acts as a Positive Regulator of Salt Stress. Plants 2022, 11, 2205. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Xu, Y.; Wang, S.; Gao, J.; Lu, D.; Zhou, C.; Liao, Y.; Ma, Y.; Zheng, Y. Identification and Expression Analysis of Zinc Finger A20/AN1 Stress-Associated Genes SmSAP Responding to Abiotic Stress in Eggplant. Horticulturae 2022, 8, 108. [Google Scholar] [CrossRef]
- Wan, F.; Gao, J.; Wang, G.; Niu, Y.; Wang, L.; Zhang, X.; Wang, Y.; Pan, Y. Genome-Wide Identification of NAC Transcription Factor Family and Expression Analysis of ATAF Subfamily Members under Abiotic Stress in Eggplant. Sci. Hortic. 2021, 289, 110424. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; He, Y.; Liu, Y.; Chen, H. Functional Characterization of SmCBF Genes Involved in Abiotic Stress Response in Eggplant (Solanum melongena). Sci. Hortic. 2018, 233, 14–21. [Google Scholar] [CrossRef]
- Hannachi, S.; Van Labeke, M.-C. Salt Stress Affects Germination, Seedling Growth and Physiological Responses Differentially in Eggplant Cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Al-Qurainy, F.; Harris, P.J.C. Salt Tolerance in Selected Vegetable Crops. Crit. Rev. Plant Sci. 2012, 31, 303–320. [Google Scholar] [CrossRef]
- Akinci, I.E.; Akinci, S.; Yilmaz, K.; Dikici, H. Response of Eggplant Varieties (Solanum melongena) to Salinity in Germination and Seedling Stages. N. Z. J. Crop Hortic. Sci. 2004, 32, 193–200. [Google Scholar] [CrossRef]
- Ünlükara, A.; Kurunç, A.; Kesmez, G.D.; Yurtseven, E.; Suarez, D.L. Effects of Salinity on Eggplant (Solanum melongena L.) Growth and Evapotranspiration. Irrig. Drain. 2010, 59, 203–214. [Google Scholar] [CrossRef]
- Li, J.; Gao, Z.; Zhou, L.; Li, L.; Zhang, J.; Liu, Y.; Chen, H. Comparative Transcriptome Analysis Reveals K+ Transporter Gene Contributing to Salt Tolerance in Eggplant. BMC Plant Biol. 2019, 19, 67. [Google Scholar] [CrossRef]
- Saeed, R.; Ahmad, R.; Mirbahar, A.A.; Jehan, B. Germination Indices of Egg Plant (Solanum melongena L.) under Sea Salt Salinity. Int. J. Biol. Biotechnol. 2014, 11, 51–55. [Google Scholar]
- Abbas, W.; Ashraf, M.; Akram, N.A. Alleviation of Salt-Induced Adverse Effects in Eggplant (Solanum melongena L.) by Glycinebetaine and Sugarbeet Extracts. Sci. Hortic. 2010, 125, 188–195. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yao, X.; Chen, J.; Zhu, Z.; Zhang, H.; Zha, D. Brassinosteroids Protect Photosynthesis and Antioxidant System of Eggplant Seedlings from High-Temperature Stress. Acta Physiol. Plant. 2014, 36, 251–261. [Google Scholar] [CrossRef]
- Lv, L.L.; Feng, X.F.; Li, W.; Li, K. High Temperature Reduces Peel Color in Eggplant (Solanum melongena) as Revealed by RNA-Seq Analysis. Genome 2019, 62, 503–512. [Google Scholar] [CrossRef]
- Zhang, A.; Zhu, Z.; Shang, J.; Zhang, S.; Shen, H.; Wu, X.; Zha, D. Transcriptome Profiling and Gene Expression Analyses of Eggplant (Solanum melongena L.) under Heat Stress. PLoS ONE 2020, 15, e0236980. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, S.; Liu, X.; Shang, J.; Zhang, A.; Zhu, Z.; Zha, D. Chalcone Synthase (CHS) Family Members Analysis from Eggplant (Solanum melongena L.) in the Flavonoid Biosynthetic Pathway and Expression Patterns in Response to Heat Stress. PLoS ONE 2020, 15, e0226537. [Google Scholar] [CrossRef]
- Rylander, M.N.; Feng, Y.; Bass, J.; Diller, K.R. Thermally Induced Injury and Heat-Shock Protein Expression in Cells and Tissues. Ann. N. Y. Acad. Sci. 2005, 1066, 222–242. [Google Scholar] [CrossRef]
- Wankhede, N.L.; Kale, M.B.; Upaganlawar, A.B.; Taksande, B.G.; Umekar, M.J.; Behl, T.; Abdellatif, A.A.H.; Bhaskaran, P.M.; Dachani, S.R.; Sehgal, A.; et al. Involvement of Molecular Chaperone in Protein-Misfolding Brain Diseases. Biomed. Pharmacother. 2022, 147, 112647. [Google Scholar] [CrossRef]
- Baird, N.A.; Turnbull, D.W.; Johnson, E.A. Induction of the Heat Shock Pathway during Hypoxia Requires Regulation of Heat Shock Factor by Hypoxia-Inducible Factor-1. J. Biol. Chem. 2006, 281, 38675–38681. [Google Scholar] [CrossRef]
- Gong, C.; Pang, Q.; Li, Z.; Li, Z.; Chen, R.; Sun, G.; Sun, B. Genome-Wide Identification and Characterization of Hsf and Hsp Gene Families and Gene Expression Analysis under Heat Stress in Eggplant (Solanum Melongema L.). Horticulturae 2021, 7, 149. [Google Scholar] [CrossRef]
- Wang, J.; Hu, H.; Wang, W.; Wei, Q.; Hu, T.; Bao, C. Genome-Wide Identification and Functional Characterization of the Heat Shock Factor Family in Eggplant (Solanum melongena L.) under Abiotic Stress Conditions. Plants 2020, 9, 915. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Pan, Y.; Li, J.; Chen, X.; Pan, Y.; Wang, Y.; Tian, S.; Zhang, X. Heterologous Expression of Arabidopsis C-Repeat Binding Factor 3 (AtCBF3) and Cold-Regulated 15A (AtCOR15A) Enhanced Chilling Tolerance in Transgenic Eggplant (Solanum melongena L.). Plant Cell Rep. 2014, 33, 1951–1961. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, B.; Chen, Z.; Liu, Y. Low temperature injury and cold resistance mechanism of eggplant. Fujian J. Agric. Sci. 2000, 15, 40–42. [Google Scholar]
- Fallik, E.; Temkin-Gorodeiski, N.; Grinberg, S.; Davidson, H. Prolonged Low-Temperature Storage of Eggplants in Polyethylene Bags. Postharvest Biol. Technol. 1995, 5, 83–89. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, J.; Zhou, X.; Liu, S.; Zhuang, Y. Transcriptomics Analysis Unravels the Response to Low Temperature in Sensitive and Tolerant Eggplants. Sci. Hortic. 2020, 271, 109468. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, J.; Zhou, X.; Liu, S.; Zhuang, Y. Identification of WRKY Gene Family and Characterization of Cold Stress-Responsive WRKY Genes in Eggplant. PeerJ 2020, 8, e8777. [Google Scholar] [CrossRef]
- Pandey, V.K.; Gopal, R. Nickel Toxicity Effects on Growth and Metabolism of Eggplant. Int. J. Veg. Sci. 2010, 16, 351–360. [Google Scholar] [CrossRef]
- Yuan, H.; Sun, L.; Tai, P.; Liu, W.; Li, X.; Hao, L. Effects of Grafting on Root-to-Shoot Cadmium Translocation in Plants of Eggplant (Solanum melongena) and Tomato (Solanum lycopersicum). Sci. Total Environ. 2019, 652, 989–995. [Google Scholar] [CrossRef]
- Saeedifar, F.; Ziarati, P.; Ramezan, Y. Nitrate and Heavy Metal Contents in Eggplant (Solanum melongena) Cultivated in the Farmlands in the South of Tehran-Iran. Int. J. Farming Allied Sci. 2014, 3, 60–65. [Google Scholar]
- Ye, X.; Xiao, W.; Zhang, Y.; Zhao, S.; Wang, G.; Zhang, Q.; Wang, Q. Assessment of Heavy Metal Pollution in Vegetables and Relationships with Soil Heavy Metal Distribution in Zhejiang Province, China. Environ. Monit. Assess. 2015, 187, 378. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Fukuoka, H.; Arao, T.; Ohyama, A.; Nunome, T.; Miyatake, K.; Negoro, S. Gene Expression Analysis in Cadmium-Stressed Roots of a Low Cadmium-Accumulating Solanaceous Plant, Solanum torvum. J. Exp. Bot. 2010, 61, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Cerruti, E.; Gisbert, C.; Drost, H.-G.; Valentino, D.; Portis, E.; Barchi, L.; Prohens, J.; Lanteri, S.; Comino, C.; Catoni, M. Grafting Vigour Is Associated with DNA De-Methylation in Eggplant. Hortic. Res. 2021, 8, 241. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Wei, S.; Twardowska, I.; Zhang, Q. In Search of the Exclusion/Low-Accumulation Mechanisms: Cadmium Uptake and Accumulation from Soil by Cultivated (Solanum melongena L.) and Wild Eggplants (Solanum torvum L.). J. Clean. Prod. 2021, 323, 129141. [Google Scholar] [CrossRef]
- Cui, W.; Tai, P.; Li, X.; Jia, C.; Yuan, H.; He, L.; Sun, L. A Reduction in Cadmium Accumulation and Sulphur Containing Compounds Resulting from Grafting in Eggplants (Solanum melogena) Is Associated with DNA Methylation. Plant Soil 2021, 468, 183–196. [Google Scholar] [CrossRef]
- Rosati, A.; Badeck, F.W.; Dejong, T.M. Estimating Canopy Light Interception and Absorption Using Leaf Mass Per Unit Leaf Area in Solanum melongena. Ann. Bot. 2001, 88, 101–109. [Google Scholar] [CrossRef]
- Wasiu Babatunde, A.; Togun, A.; Olaniran, O.; Akinfasoye, J.O.; Tairu, F.M. Physico-Chemical Properties of Egg Plant (Solanum melongena L.) Fruit in Response to Nitrogen Fertilizer and Fruit Size. Agric. J. 2007, 2, 140–148. [Google Scholar]
- Pal, S.; Saimbhi, M.S.; Bal, S. Effect of Nitrogen and Phosphorus Levels on Growth and Yield of Brinjal Hybrids(Solanum melongena). Veg. Sci. 2002, 29, 90–91. [Google Scholar]
- Mauceri, A.; Bassolino, L.; Lupini, A.; Badeck, F.; Rizza, F.; Schiavi, M.; Toppino, L.; Abenavoli, M.R.; Rotino, G.L.; Sunseri, F. Genetic Variation in Eggplant for Nitrogen Use Efficiency under Contrasting NO3—Supply. J. Integr. Plant Biol. 2020, 62, 487–508. [Google Scholar] [CrossRef]
- Mauceri, A.; Abenavoli, M.R.; Toppino, L.; Panda, S.; Mercati, F.; Aci, M.M.; Aharoni, A.; Sunseri, F.; Rotino, G.L.; Lupini, A. Transcriptomics Reveal New Insights into Molecular Regulation of Nitrogen Use Efficiency in Solanum melongena. J. Exp. Bot. 2021, 72, 4237–4253. [Google Scholar] [CrossRef]
- Mauceri, A.; Aci, M.M.; Toppino, L.; Panda, S.; Meir, S.; Mercati, F.; Araniti, F.; Lupini, A.; Panuccio, M.R.; Rotino, G.L.; et al. Uncovering Pathways Highly Correlated to NUE through a Combined Metabolomics and Transcriptomics Approach in Eggplant. Plants 2022, 11, 700. [Google Scholar] [CrossRef]
- Guan, W.; Ke, C.; Tang, W.; Jiang, J.; Xia, J.; Xie, X.; Yang, M.; Duan, C.; Wu, W.; Zheng, Y. Construction of a High-Density Recombination Bin-Based Genetic Map Facilitates High-Resolution Mapping of a Major QTL Underlying Anthocyanin Pigmentation in Eggplant. Int. J. Mol. Sci. 2022, 23, 10258. [Google Scholar] [CrossRef] [PubMed]
- Al-sane, K.O.; Povero, G.; Perata, P. Anthocyanin Tomato Mutants: Overview and Characterization of an Anthocyanin-Less Somaclonal Mutant. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2011, 145, 436–444. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).